Oxysterols from a Marine Sponge Inflatella sp. and Their Action in 6-Hydroxydopamine-Induced Cell Model of Parkinson’s Disease

 ,
, 

Abstract
1. Introduction
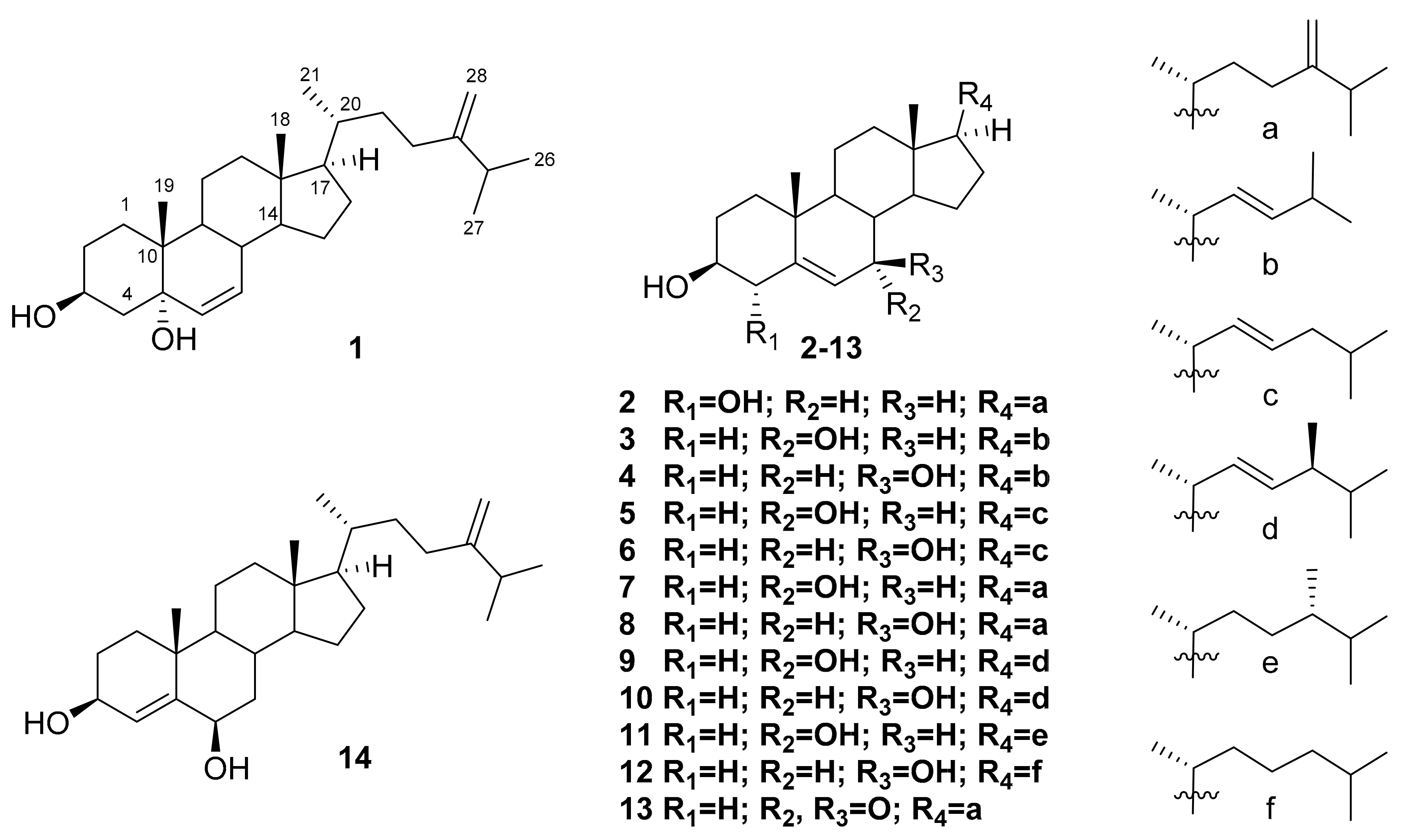
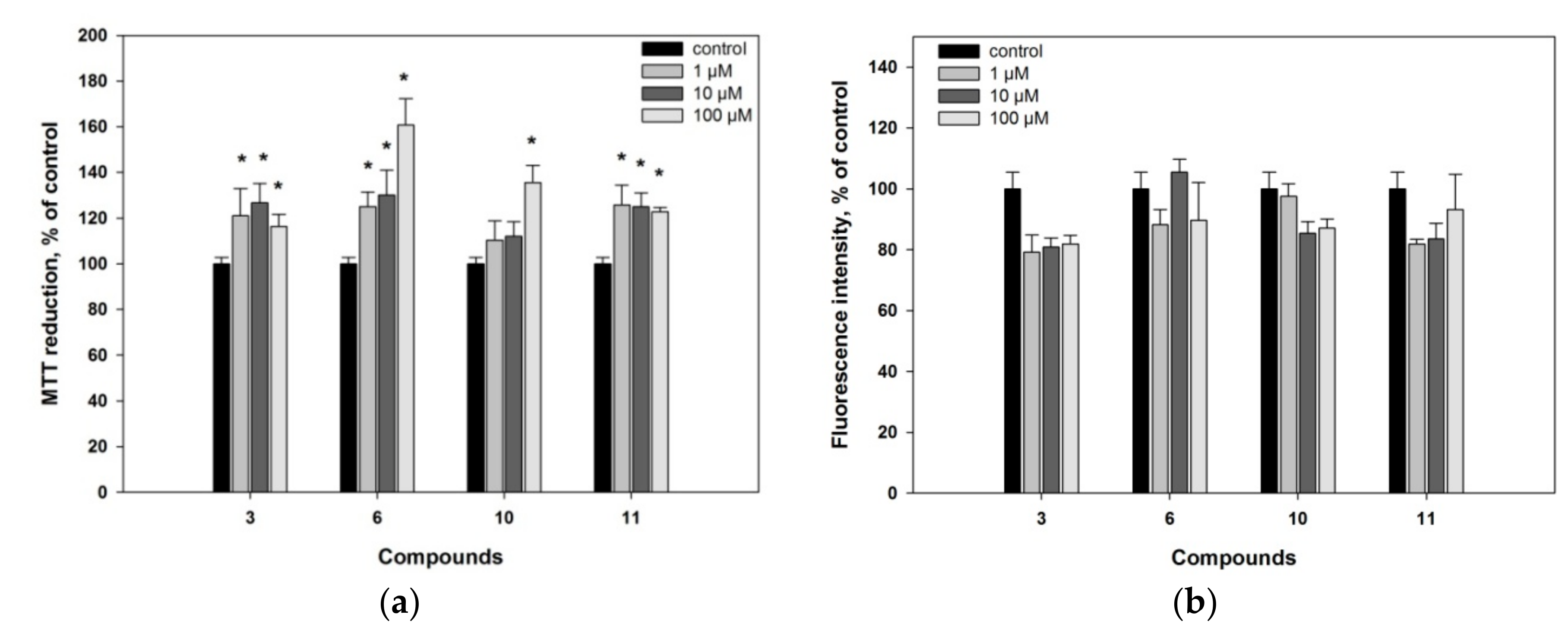
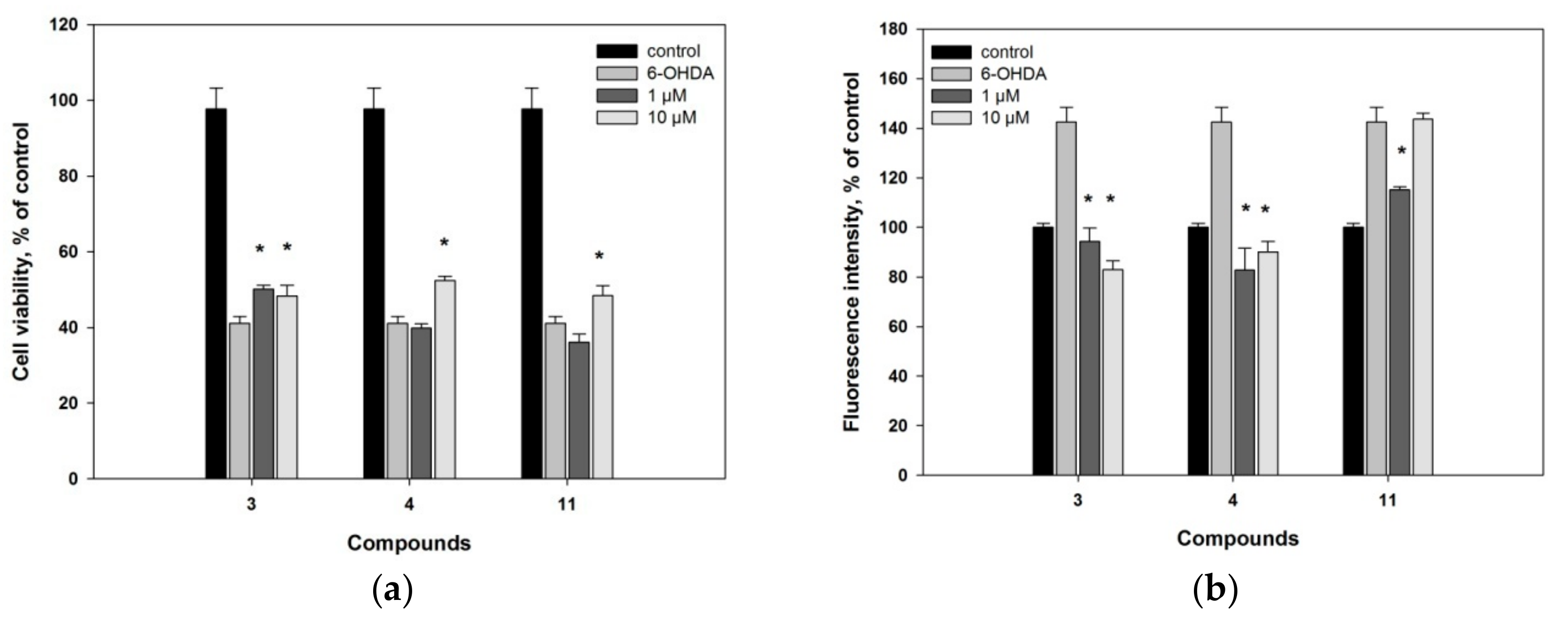
2. Results and Discussion
3. Materials and Methods
3.1. General Methods
3.2. Animal Material
3.3. Extraction and Isolation
3.3.1. 24-Methylcholesta-6,24(28)-diene-3β,5α-diol (1)
3.3.2. 24-Methylcholesta-5,24(28)-diene-3β,4α-diol (2)
3.3.3. (22E)-24-Nor-cholesta-5,22-diene-3β,7α-diol (3)
3.3.4. (22E)-24-Nor-cholesta-5,22-diene-3β,7β-diol (4)
3.4. Biological Activities
3.4.1. Cell Line and Culture Condition
3.4.2. MTT Assay
3.4.3. Nonspecific Esterase Activity Assay
3.4.4. 6-OHDA-Induced In Vitro Model of Parkinson’s Disease
3.4.5. Reactive Oxygen Species (ROS) Level Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schroepfer, G.J., Jr. Oxysterols: Modulators of cholesterol metabolism and other processes. Physiol. Revs. 2000, 80, 361–554. [Google Scholar] [CrossRef] [PubMed]
- Doria, M.; Maugest, L.; Moreau, T.; Lizard, G.; Vejux, A. Contribution of cholesterol and oxysterols to the pathophysiology of Parkinson’s disease. Free Radic. Biol. Med. 2016, 101, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, W.J.; Abdel-Khalik, J.T.; Yutuc, H.E.; Morgan, A.H.; Wang, Y. Current trends in oxysterol research. Biochem. Soc. Trans. 2016, 44, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.Y.J.; Seet, R.C.S.; Huang, S.H.; Long, L.H.; Halliwell, B. Different patterns of oxidized lipid products in plasma and urine of dengue fever, stroke, and Parkinson’s disease patients: Cautions in the use of biomarkers of oxidative stress. Antioxyd. Redox Signal. 2009, 11, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Marwarha, G.; Rhen, T.; Schommer, T.; Ghribi, O. The oxysterol 27-hydroxycholesterol regulates α-synuclein and tyrosine hydroxylase expression levels in human neuroblastoma cells through modulation of liver X receptors and estrogen receptors–relevance to Parkinson’s disease. J. Neurochem. 2011, 119, 1119–1136. [Google Scholar] [CrossRef] [PubMed]
- Iuliano, L.; Crick, P.J.; Zerbinati, C.; Tritapepe, L.; Abdel-Khalik, J.; Poirot, M.; Wang, Y.; Griffiths, W.J. Cholesterol metabolites exported from human brain. Steroids 2015, 99, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Crick, P.J.; Bentley, T.W.; Abdel-Khalik, J.; Matthews, I.; Clayton, P.T.; Morris, A.A.; Bigger, B.W.; Zerbinati, C.; Tritapepe, L.; Iuliano, L.; et al. Quantitative charge-tags for sterol and oxysterol analysis. Clin. Chem. 2015, 61, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.Z.; Liu, J.W.; Wang, X.; Jeong, I.H.; Ahn, Y.J.; Zhang, C.J. Anti-BACE1 and antimicrobial activities of steroidal compounds isolated from marine Urechis unicinctus. Mar. Drugs 2018, 16, 94. [Google Scholar] [CrossRef] [PubMed]
- Kolesnikova, S.A.; Lyakhova, E.G.; Kalinovsky, A.I.; Pushilin, M.A.; Afiyatullov, S.S.; Yurchenko, E.A.; Dyshlovoy, S.A.; Minh, C.V.; Stonik, V.A. Isolation, structures, and biological activities of triterpenoids from a Penares sp. marine sponge. J. Nat. Prod. 2013, 76, 1746–1752. [Google Scholar] [CrossRef] [PubMed]
- Lyakhova, E.G.; Kolesnikova, S.A.; Kalinovskii, A.I.; Kim, N.Y.; Krasokhin, V.B.; Stonik, V.A. Secondary metabolites of the marine sponge Penares cf. schulzei, fluorescence properties of 24-methylenecholesta-4,6,8(14)-trien-3-one. Chem. Nat. Comp. 2015, 51, 334–335. [Google Scholar] [CrossRef]
- Lyakhova, E.G.; Kolesnikova, S.A.; Kalinovsky, A.I.; Dmitrenok, P.S.; Nam, N.H.; Stonik, V.A. Further study on Penares sp. from Vietnamese waters: minor lanostane and nor-lanostane triterpenes. Steroids 2015, 96, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Berne, S.; Kalauz, M.; Lapat, M.; Savin, L.; Janussen, D.; Kersken, D.; Avguštin, J.A.; Jokhadar, Š.Z.; Jaklič, D.; Gunde-Cimerman, N.; et al. Screening of the Antarctic marine sponges (Porifera) as a source of bioactive compounds. Polar Biol. 2016, 39, 947–959. [Google Scholar] [CrossRef]
- Holland, H.L.; Jahangir. Reactions of steroidal 4,5- and 5,6- epoxides with strong bases. Can. J. Chem. 1983, 61, 2165–2170. [Google Scholar] [CrossRef]
- Della Greca, M.; Fiorentino, A.; Molinaro, A.; Monaco, P.; Previtera, L. Hydroperoxysterols in Arum italicum. Nat. Prod. Lett. 1994, 5, 7–14. [Google Scholar] [CrossRef]
- Zielinski, Z.A.M.; Pratt, D.A. Cholesterol autoxidation revisited: debunking the dogma associated with the most vilified of lipids. J. Am. Chem. Soc. 2016, 138, 6932–6935. [Google Scholar] [CrossRef] [PubMed]
- Notaro, G.; Piccialli, V.; Sica, D. New steroidal hydroxyketones and closely related diols from the marine sponge Cliona copiosa. J. Nat. Prod. 1992, 55, 1588–1594. [Google Scholar] [CrossRef]
- Liu, T.F.; Lu, X.; Tang, H.; Zhang, M.M.; Wang, P.; Sun, P.; Liu, Z.Y.; Wang, Z.L.; Li, L.; Rui, Y.C.; et al. 3β,5α,6β-Oxygenated sterols from the South China Sea gorgonian Muriceopsis flavida and their tumor cell growth inhibitory activity and apoptosis-inducing function. Steroids 2013, 78, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Findlay, J.A.; Patil, A.D. Novel sterols from the finger sponge Haliclona oculata. Can. J. Chem. 1985, 63, 2406–2410. [Google Scholar] [CrossRef]
- Yang, F.; Zhang, H.-J.; Liu, X.-F.; Chen, W.-S.; Tang, H.-F.; Lin, H.-W. Oxygenated steroids from marine bryozoan Biflustra grandicella. Biochem. System. Ecol. 2009, 37, 686–689. [Google Scholar] [CrossRef]
- Anjaneyulu, V.; Babu, B.H.; Rao, K.M.C.A.; Rao, K.N. 24-Methylenecholest-4-ene-3β,6β-diol from a soft coral Sinularia ovispiculata of the Andaman and Nicobar Islands. Ind. J. Chem. B 1994, 33B, 806–808. [Google Scholar]
- Deghrigue, M.; Festa, C.; Ghribi, L.; D’Auria, M.V.; De Marino, S.; Ben Jannet, H.; Bouraoui, A. Anti-inflammatory and analgesic activities with gastroprotective effect of semi-purified fractions and isolation of pure compounds from Mediterranean gorgonian Eunicella singularis. Asian Pac. J. Trop. Med. 2015, 8, 606–611. [Google Scholar] [CrossRef] [PubMed]
- Slattery, M.; Hamann, M.T.; McClintock, J.B.; Perry, T.L.; Puglisi, M.P.; Yoshida, W.Y. Ecological role of water-borne metabolites from Antarctic soft corals. Mar. Ecol. Prog. Ser. 1997, 161, 133–144. [Google Scholar] [CrossRef]
- Gunatilaka, A.A.L.; Samaranayake, G.; Kingston, D.G.I.; Hoffmann, G.; Johnson, R.K. Bioactive ergost-5-ene-3β,7α-diol derivatives from Pseudobersama mossambicensis. J. Nat. Prod. 1992, 55, 1648–1654. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Zhang, C.; Zheng, K.; Su, J.; Zeng, L. Spectral characteristics and cholinesterase inhibitory activities of some Δ5-3β,7β-dihydroxy sterols and their C-7 epimers. Tianran Chanwu Yanjiu Yu Kaifa 2006, 18, 893–895. [Google Scholar]
- Heltzel, C.E.; Gunatilaka, A.A.L.; Kingston, D.G.I.; Hofmann, G.A.; Johnson, R.K. Synthesis and structure-activity relationships of cytotoxic 7-hydroxy sterols. J. Nat. Prod. 1994, 57, 620–628. [Google Scholar] [CrossRef] [PubMed]
- Kodai, T.; Umebayashi, K.; Nakatani, T.; Ishiyama, K.; Noda, N. Compositions of royal jelly II. Organic acid glycosides and sterols of the royal jelly of honeybees (Apis mellifera). Chem. Pharm. Bull. 2007, 55, 1528–1531. [Google Scholar] [CrossRef] [PubMed]
- Abdelhameed, R.F.; Ibrahim, A.K.; Yamada, K.; Ahmed, S.A. Cytotoxic and anti-inflammatory compounds from Red Sea grass Thalassodendron ciliatum. Med. Chem. Res. 2018, 27, 1238–1244. [Google Scholar] [CrossRef]
- Ward, L.J.; Ljunggren, S.A.; Karlsson, H.; Li, W.; Yuan, X.M. Exposure to atheroma-relevant 7-oxysterols causes proteomic alterations in cell death, cellular longevity, and lipid metabolism in THP-1 macrophages. PLoS ONE 2017, 12, e0174475/1–e0174475/17. [Google Scholar] [CrossRef] [PubMed]
- Thao, N.P.; Nam, N.H.; Cuong, N.X.; Tai, B.H.; Quang, T.H.; Ngan, N.T.T.; Luyen, B.T.T.; Yang, S.Y.; Choi, C.H.; Kim, S.; et al. Steroidal constituents from the soft coral Sinularia dissecta and their inhibitory effects on lipopolysaccharide-stimulated production of pro-inflammatory cytokines in bone marrow-derived dendritic cells. Bull. Kor. Chem. Soc. 2013, 34, 949–952. [Google Scholar]
- Zeng, L.; Li, X.; Su, J.; Fu, X.; Schmitz, F.J. A new cytotoxic dihydroxy sterol from the soft coral Alcyonium patagonicum. J. Nat.Prod. 1995, 58, 296–298. [Google Scholar] [CrossRef] [PubMed]
- Pasban-Aliabadi, H.; Esmaeili-Mahani, S.; Abbasnejad, M. Orexin-A protects human neuroblastoma SH-SY5Y cells against 6-hydroxydopamine-induced neurotoxicity: involvement of PKC and PI3K signaling pathways. Rejuvenation Res. 2017, 20, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Stockert, J.C.; Horobin, R.W.; Colombo, L.L.; Blázquez-Castro, A. Tetrazolium salts and formazan products in cell biology: Viability assessment, fluorescence imaging, and labeling perspectives. Acta Histochem. 2018, 120, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Stepanenko, A.A.; Dmitrenko, V.V. Pitfalls of the MTT assay: Direct and off-target effects of inhibitors can result in over/under estimation of cell viability. Gene 2015, 574, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Bernhard, D.; Schwaiger, W.; Crazzolara, R.; Tinhofer, I.; Kofler, R.; Csordas, A. Enhanced MTT-reducing activity under growth inhibition by resveratrol in CEM-C7H2 lymphocytic leukemia cells. Cancer Lett. 2003, 195, 193–199. [Google Scholar] [CrossRef]
- Bitterman, J.L.; Chung, J.H. Metabolic effects of resveratrol: Addressing the controversies. Cell. Mol. Life Sci. 2015, 72, 1473–1488. [Google Scholar] [CrossRef] [PubMed]
- Mhyre, T.R.; Boyd, J.T.; Hamill, R.W.; Maguire-Zeiss, K.A. Parkinson’s Disease. Subcell. Biochem. 2012, 65, 389–455. [Google Scholar] [PubMed]
- Pan, Z.; Niu, Y.; Liang, Y.; Zhang, X.; Dong, M. β-Ecdysterone protects SH-SY5Y cells against 6-hydroxydopamine-induced apoptosis via mitochondria-dependent mechanism: Involvement of p38(MAPK)-p53 signaling pathway. Neurotox. Res. 2016, 30, 453–466. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| No. | 1 a | 2 a | 3 b | 4 a |
|---|---|---|---|---|
| 1 | 28.5 | 36.7 | 37.0 | 37.0 |
| 2 | 30.8 | 28.1 | 31.4 | 31.6 |
| 3 | 67.4 | 76.6 | 71.4 | 71.5 |
| 4 | 41.2 | 75.2 | 42.1 | 41.8 |
| 5 | 74.2 | 142.1 | 146.2 | 143.7 |
| 6 | 133.5 | 117.8 | 123.9 | 125.5 |
| 7 | 133.4 | 31.6 | 65.3 | 73.4 |
| 8 | 38.7 | 31.5 | 37.5 | 40.9 |
| 9 | 45.3 | 50.5 | 42.3 | 48.3 |
| 10 | 38.3 | 38.1 | 37.4 | 36.5 |
| 11 | 21.3 | 20.9 | 20.7 | 21.1 |
| 12 | 40.3 | 39.8 | 39.1 | 39.5 |
| 13 | 44.0 | 42.3 | 42.0 | 42.8 |
| 14 | 54.2 | 56.8 | 49.5 | 56.0 |
| 15 | 24.1 | 24.3 | 24.2 | 26.3 |
| 16 | 28.8 | 28.2 | 28.4 | 28.6 |
| 17 | 56.1 | 56.0 | 55.8 | 55.4 |
| 18 | 12.4 | 11.9 | 11.9 | 12.1 |
| 19 | 14.9 | 20.2 | 18.2 | 19.2 |
| 20 | 36.0 | 35.8 | 39.8 | 39.7 |
| 21 | 18.9 | 18.7 | 20.8 | 20.9 |
| 22 | 34.9 | 34.7 | 133.6 | 133.5 |
| 23 | 31.3 | 31.0 | 134.9 | 135.0 |
| 24 | 157.1 | 156.9 | ||
| 25 | 34.1 | 33.8 | 30.9 | 30.9 |
| 26 | 22.1 | 21.9 | 22.7 | 22.7 |
| 27 | 22.2 | 22.0 | 22.7 | 22.7 |
| 28 | 106.3 | 106.0 |
| No. a | 1 b | 2 c | 3 c | 4 b |
|---|---|---|---|---|
| 1 | α: 1.62, m β: 1.37, m | α: 1.14, m β: 1.84, m | α: 1.12, m β: 1.87, m | α: 1.06, m β: 1.85, m |
| 2 | α: 1.89, m β: 1.55, m | α: 1.89, m β: 1.60, m | α: 1.86, m β: 1.52, m | α: 1.85, m β: 1.52, m |
| 3 | 4.12, m | 3.27, td (10.5, 4.9) | 3.39, m | 3.55, m |
| 4 | α: 1.80, dd (12.4, 4.9) β: 1.64, m | 4.06, m | α: 2.34, ddd (13.2, 4.9, 2.2) β: 2.29, ddt (13.2, 11.3, 1.8) | α: 2.34, ddd (13.2, 5.2, 2.3) β: 2.26, ddt (13.2, 11.3, 2.2) |
| 5 | ||||
| 6 | 5.57, dd (9.8, 2.7) | 5.74, dt (5.5, 2.2) | 5.60, dd (5.3, 1.6) | 5.29, br t (2.2) |
| 7 | 5.63, dd (9.8, 1.6) | α: 1.58, m β: 2.10, m | 3.85, br s | 3.84, dt (8.3, 2.2) |
| 8 | 1.93, m | 1.44, m | 1.47, m | 1.40, m |
| 9 | 1.48, dd (11.6, 3.9) | 0.99, m | 1.23, m | 1.03, m |
| 10 | ||||
| 11 | α: 1.44, m β: 1.31, m | α: 1.01, m β: 1.48, m | α: 1.54, m β: 1.49, m | α: 1.53, m β: 1.47, m |
| 12 | α: 1.23, m β: 2.05, dt (12.5, 3.1) | α: 1.18, td (12.7, 5.2) β: 2.03, dt (12.7, 3.7) | α: 1.19, m β: 1.98, dt (12.6, 3.5) | α: 1.15, m β: 1.99, m |
| 13 | ||||
| 14 | 1.23, m | 1.00, m | 1.44, m | 1.16, m |
| 15 | α: 1.67, m β: 1.22, m | α: 1.61, m β: 1.10, m | α: 1.67, m β: 1.12, m | α: 1.77, m β: 1.40, m |
| 16 | α: 1.89, m β: 1.32, m | α: 1.87, m β: 1.29, m | α: 1.73, m β: 1.29, m | α: 1.71, m β: 1.30, m |
| 17 | 1.17, m | 1.14, m | 1.20, m | 1.12, m |
| 18 | 0.71, s | 0.69, s | 0.70, s | 0.70, s |
| 19 | 0.92, s | 1.03, s | 1.00, s | 1.35, s |
| 20 | 1.43, m | 1.42, m | 2.01, m | 2.00, m |
| 21 | 0.96, d (6.5) | 0.95, d (6.6) | 1.01, d (6.7) | 1.01, d (6.6) |
| 22 | 1.16, m 1.55, m | 1.16, m 1.55, m | 5.18, dd (15.4, 8.4) | 5.18, dd (15.4, 8.4) |
| 23 | 1.88, m 2.10, ddd (15.4, 11.5, 4.8) | 1.88, m 2.09, m | 5.27, dd (15.3, 6.6) | 5.28, dd (15.4, 6.5) |
| 24 | ||||
| 25 | 2.23, sept (6.8) | 2.23, sept (7.0) | 2.19, br sept (6.7) | 2.19, br sept (6.7) |
| 26 | 1.03, d (6.8) | 1.02, d (7.0) | 0.94, d (6.7) | 0.95, d (6.7) |
| 27 | 1.02, d (6.8) | 1.03, d (7.0) | 0.94, d (6.7) | 0.95, d (6.7) |
| 28 | 4.66, s 4.72, s | 4.66, s 4.72, s |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kolesnikova, S.A.; Lyakhova, E.G.; Kalinovsky, A.I.; Popov, R.S.; Yurchenko, E.A.; Stonik, V.A. Oxysterols from a Marine Sponge Inflatella sp. and Their Action in 6-Hydroxydopamine-Induced Cell Model of Parkinson’s Disease. Mar. Drugs 2018, 16, 458. https://doi.org/10.3390/md16110458
Kolesnikova SA, Lyakhova EG, Kalinovsky AI, Popov RS, Yurchenko EA, Stonik VA. Oxysterols from a Marine Sponge Inflatella sp. and Their Action in 6-Hydroxydopamine-Induced Cell Model of Parkinson’s Disease. Marine Drugs. 2018; 16(11):458. https://doi.org/10.3390/md16110458
Chicago/Turabian StyleKolesnikova, Sophia A., Ekaterina G. Lyakhova, Anatoly I. Kalinovsky, Roman S. Popov, Ekaterina A. Yurchenko, and Valentin A. Stonik. 2018. "Oxysterols from a Marine Sponge Inflatella sp. and Their Action in 6-Hydroxydopamine-Induced Cell Model of Parkinson’s Disease" Marine Drugs 16, no. 11: 458. https://doi.org/10.3390/md16110458
APA StyleKolesnikova, S. A., Lyakhova, E. G., Kalinovsky, A. I., Popov, R. S., Yurchenko, E. A., & Stonik, V. A. (2018). Oxysterols from a Marine Sponge Inflatella sp. and Their Action in 6-Hydroxydopamine-Induced Cell Model of Parkinson’s Disease. Marine Drugs, 16(11), 458. https://doi.org/10.3390/md16110458