Evaluation of Rapid, Early Warning Approaches to Track Shellfish Toxins Associated with Dinophysis and Alexandrium Blooms

Abstract
1. Introduction
2. Materials and Methods
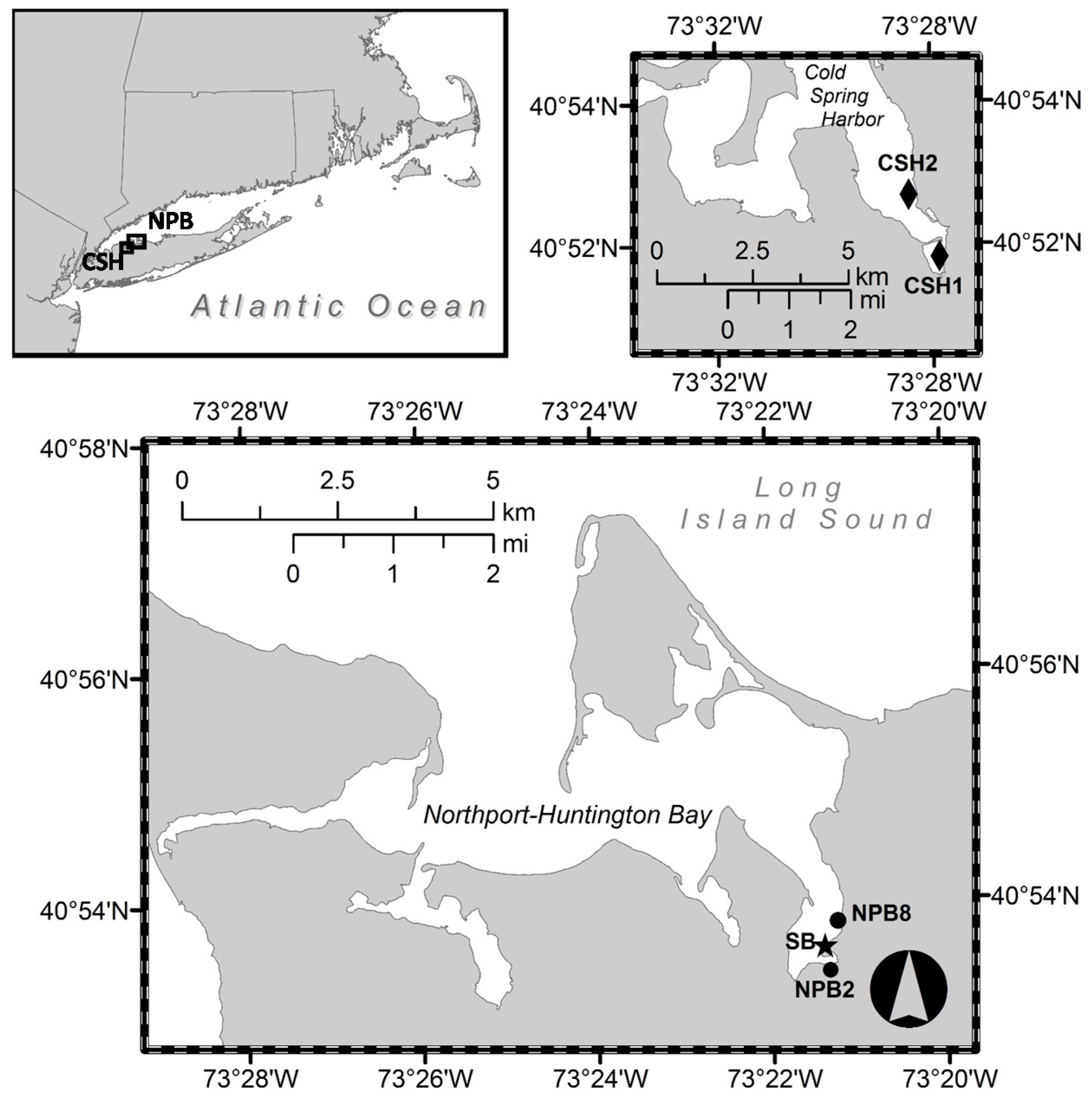
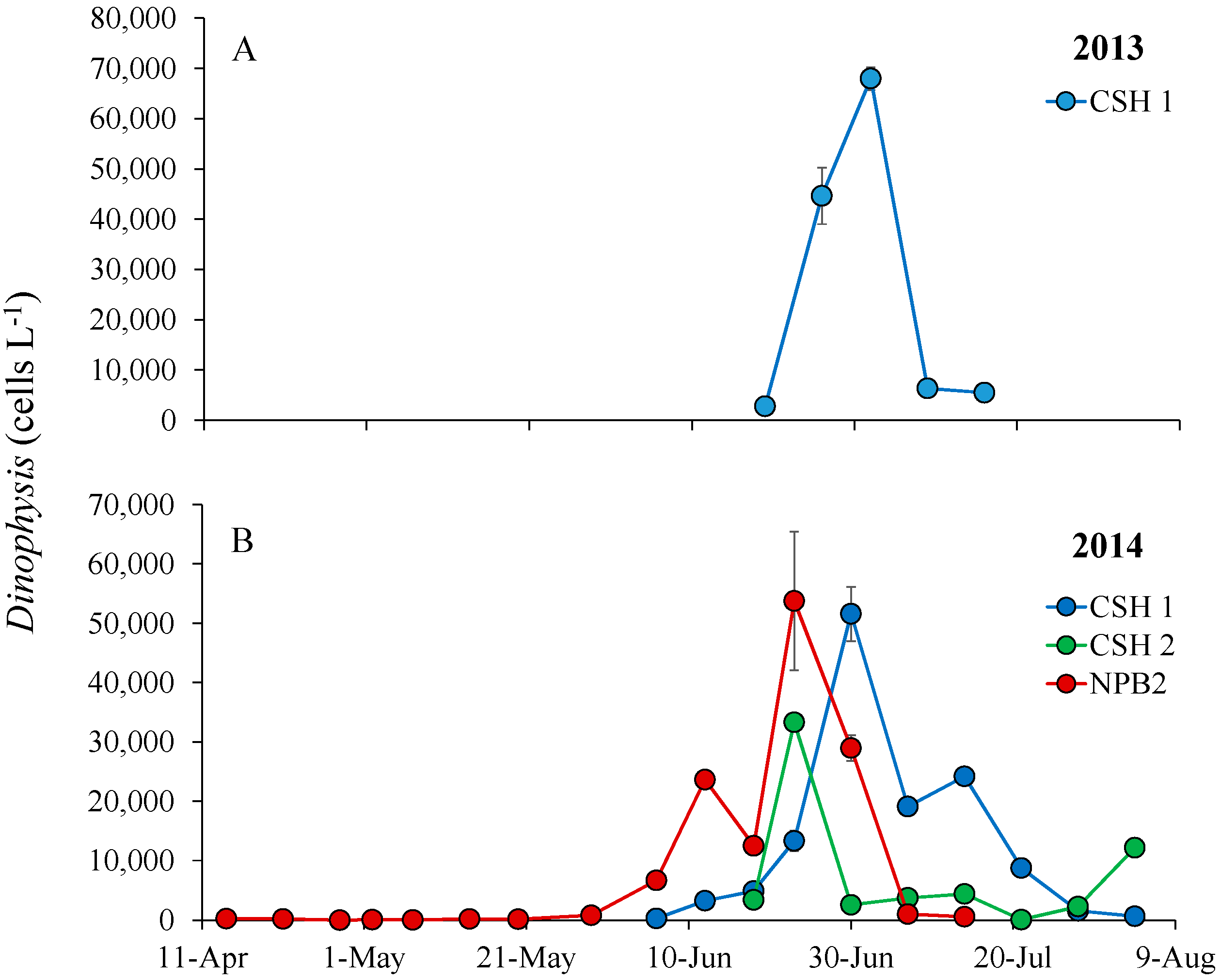
2.1. Time Series Field Sampling
2.2. Shellfish and SPATT Sampler Deployment and Collection
2.3. Analysis of PSP Toxins in Algal Pellets, Shellfish and SPATT
2.4. Analysis of DSP Toxins in Algal Pellets, Shellfish and SPATT
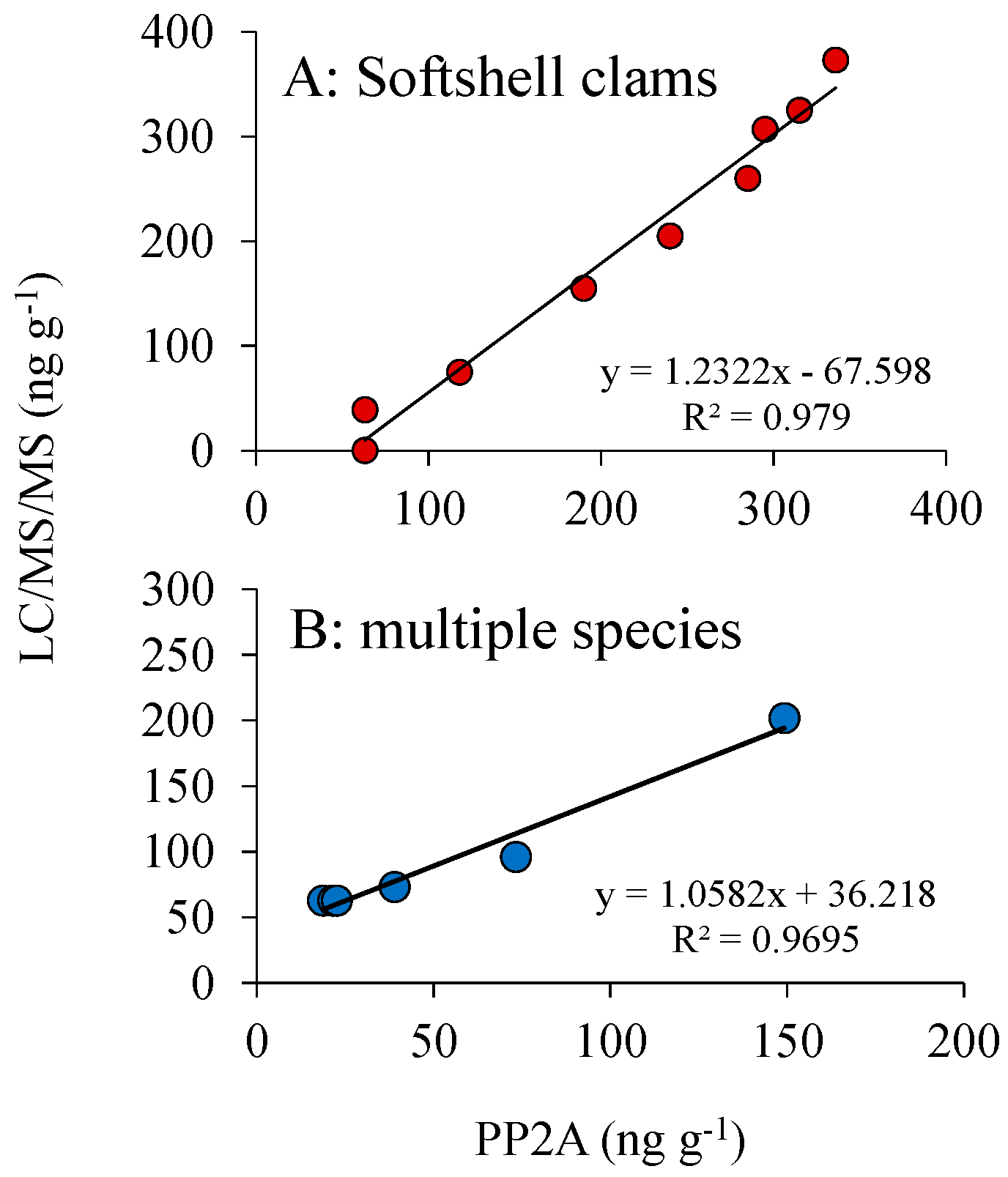
2.5. Comparison of DSP Toxin Analysis Methodologies, LC/MS and PP2A
2.6. Statistical Analysis
3. Results
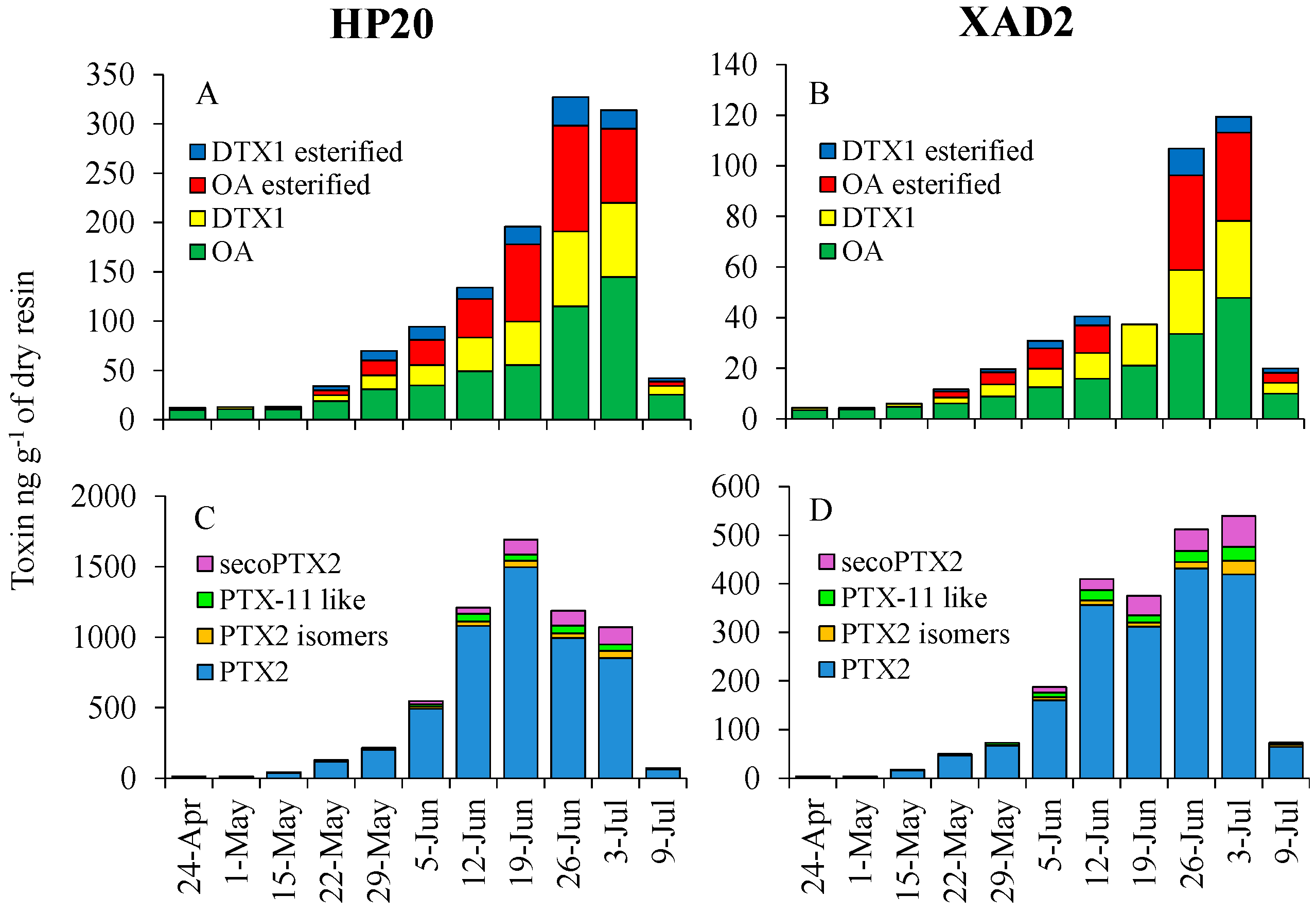
3.1. SPATT Technology: Comparison of HP20 and XAD2 Resins
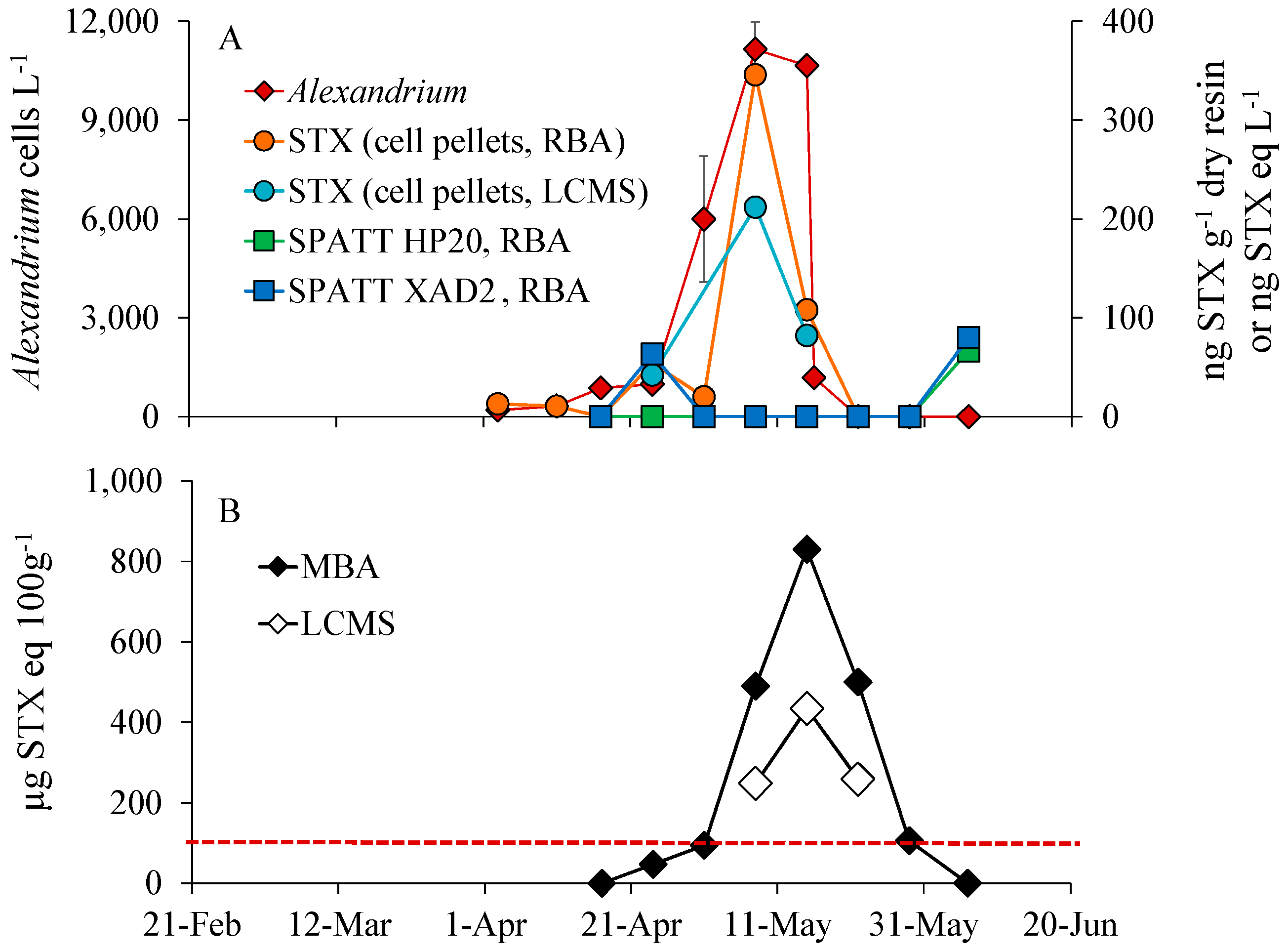
3.2. Alexandrium Bloom Dynamics and PSP Toxins in Phytoplankton Concentrates, Shellfish and SPATT
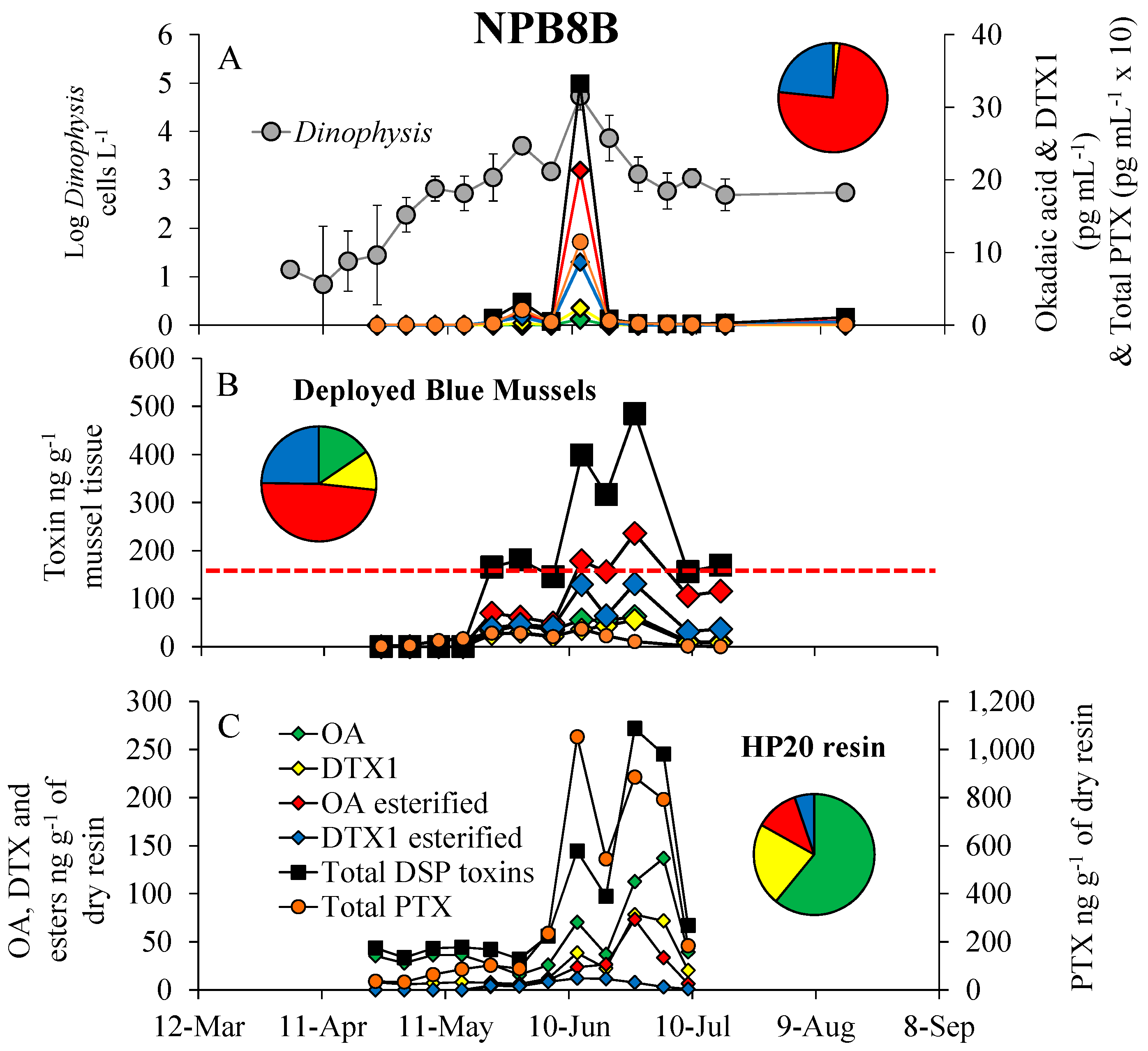
3.3. Dinophysis Bloom Dynamics and DSP Toxins in Phytoplankton Concentrates, Shellfish and SPATT
3.4. DSP Toxin Analysis: LC/MS vs. PP2A
4. Discussion
4.1. DSP SPATT Samplers
4.2. DSP in SPATT, Bivalves and Cells
4.3. PSP SPATT Samplers and Toxicity
4.4. The Co-Occurrence of DSP and PSP Toxins in Shellfish
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hallegraeff, G.M. A review of harmful algal blooms and their apparent global increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef]
- Glibert, P.M.; Anderson, D.M.; Gentien, P.; Granéli, E.; Sellner, K.G. The global, complex phenomena of harmful algal blooms. Oceanography 2005, 18, 136–147. [Google Scholar] [CrossRef]
- Hallegraeff, G.M. Ocean climate change, phytoplankton community responses, and harmful algal blooms: A formidable predictive challenge. J. Phycol. 2010, 46, 220–235. [Google Scholar] [CrossRef]
- Anderson, D.M.; Burkholder, J.M.; Cochlan, W.P.; Glibert, P.M.; Gobler, C.J.; Heil, C.A.; Kudela, R.M.; Parsons, M.L.; Rensel, J.; Townsend, D.W. Harmful algal blooms and eutrophication: Examining linkages from selected coastal regions of the United States. Harmful Algae 2008, 8, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Cembella, A.D.; Hallegraeff, G.M. Progress in understanding harmful algal blooms: Paradigm shifts and new technologies for research, monitoring, and management. Ann. Rev. Mar. Sci. 2012, 4, 143–176. [Google Scholar] [CrossRef] [PubMed]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, J.; Hoogenboom, R.L.A.P.; Hendriksen, P.J.M.; Bodero, M.; Bovee, T.F.H.; Rietjens, I.M.C.M.; Gerssen, A. Marine biotoxins and associated outbreaks following seafood consumption: Prevention and surveillance in the 21st century. Glob. Food Secur. 2017, 15, 11–21. [Google Scholar] [CrossRef]
- Lewitus, A.J.; Horner, R.A.; Caron, D.A.; Garcia-Mendoza, E.; Hickey, B.M.; Hunter, M.; Huppert, D.D.; Kudela, R.M.; Langlois, G.W.; Largier, J.L.; et al. Harmful algal blooms along the North American west coast region: History, trends, causes, and impacts. Harmful Algae 2012, 19, 133–159. [Google Scholar] [CrossRef]
- Feifel, K.M.; Moore, S.K.; Horner, R.A. An Alexandrium spp. cyst record from Sequim Bay, Washington state, USA, and its relation to past climate variability. J. Phycol. 2012, 48, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Horner, R.A.; Garrison, D.L.; Plumley, F.G. Harmful algal blooms and red tide problems on the U.S. west coast. Limnol. Oceanogr. 1997, 42, 1076–1088. [Google Scholar] [CrossRef]
- Anderson, D.M. Bloom dynamics of toxic Alexandrium species in the northeastern U.S. Limnol. Oceanogr. 1997, 42, 1009–1022. [Google Scholar] [CrossRef]
- Trainer, V.; Moore, L.; Bill, B.; Adams, N.; Harrington, N.; Borchert, J.; da Silva, D.; Eberhart, B.-T. Diarrhetic shellfish toxins and other lipophilic toxins of human health concern in Washington State. Mar. Drugs 2013, 11, 1815–1835. [Google Scholar] [CrossRef] [PubMed]
- Massachusetts Division of Marine Fisheries. Massachusetts Division of Marine Fisheries 2015 Annual Report; Department of Fish and Game: Boston, MA, USA, 2015; p. 110.
- Hattenrath-Lehmann, T.K.; Marcoval, M.A.; Berry, D.L.; Fire, S.; Wang, Z.; Morton, S.L.; Gobler, C.J. The emergence of Dinophysis acuminata blooms and DSP toxins in shellfish in New York waters. Harmful Algae 2013, 26, 33–44. [Google Scholar] [CrossRef]
- Campbell, L.; Olson, R.J.; Sosik, H.M.; Abraham, A.; Henrichs, D.W.; Hyatt, C.J.; Buskey, E.J. First harmful Dinophysis (Dinophyceae, Dinophysiales) bloom in the U.S. is revealed by automated imaging flow cytometry. J. Phycol. 2010, 46, 66–75. [Google Scholar] [CrossRef]
- Swanson, K.M.; Flewelling, L.J.; Byrd, M.; Nunez, A.; Villareal, T.A. The 2008 Texas Dinophysis ovum bloom: Distribution and toxicity. Harmful Algae 2010, 9, 190–199. [Google Scholar] [CrossRef]
- Deeds, J.R.; Wiles, K.; Heideman, G.B.; White, K.D.; Abraham, A. First US report of shellfish harvesting closures due to confirmed okadaic acid in Texas Gulf coast oysters. Toxicon 2010, 55, 1138–1146. [Google Scholar] [CrossRef] [PubMed]
- Hoagland, P.; Anderson, D.; Kaoru, Y.; White, A. The economic effects of harmful algal blooms in the United States: Estimates, assessment issues, and information needs. Estuaries 2002, 25, 819–837. [Google Scholar] [CrossRef]
- Jin, D.; Hoagland, P. The value of harmful algal bloom predictions to the nearshore commercial shellfish fishery in the Gulf of Maine. Harmful Algae 2008, 7, 772–781. [Google Scholar] [CrossRef]
- Jin, D.; Thunberg, E.; Hoagland, P. Economic impact of the 2005 red tide event on commercial shellfish fisheries in New England. Ocean Coast. Manag. 2008, 51, 420–429. [Google Scholar] [CrossRef]
- Koukaras, K.; Nikolaidis, G. Dinophysis blooms in Greek coastal waters (Thermaikos Gulf, NW Aegean Sea). J. Plankton Res. 2004, 26, 445–457. [Google Scholar] [CrossRef]
- MacKenzie, L.; Beuzenberg, V.; Holland, P.; McNabb, P.; Selwood, A. Solid phase adsorption toxin tracking (SPATT): A new monitoring tool that simulates the biotoxin contamination of filter feeding bivalves. Toxicon 2004, 44, 901–918. [Google Scholar] [CrossRef] [PubMed]
- Kudela, R.M. Characterization and deployment of Solid Phase Adsorption Toxin Tracking (SPATT) resin for monitoring of microcystins in fresh and saltwater. Harmful Algae 2011, 11, 117–125. [Google Scholar] [CrossRef]
- Gibble, C.M.; Kudela, R.M. Detection of persistent microcystin toxins at the land–sea interface in Monterey Bay, California. Harmful Algae 2014, 39, 146–153. [Google Scholar] [CrossRef]
- Lane, J.Q.; Roddam, C.M.; Langlois, G.W.; Kudela, R.M. Application of solid phase adsorption toxin tracking (SPATT) for field detection of the hydrophilic phycotoxins domoic acid and saxitoxin in coastal California. Limnol. Oceanogr. Methods 2010, 8, 645–660. [Google Scholar] [CrossRef]
- Rodríguez, P.; Alfonso, A.; Turrell, E.; Lacaze, J.-P.; Botana, L.M. Study of solid phase adsorption of paralytic shellfish poisoning toxins (PSP) onto different resins. Harmful Algae 2011, 10, 447–455. [Google Scholar] [CrossRef]
- McCarthy, M.; van Pelt, F.N.; Bane, V.; O’Halloran, J.; Furey, A. Application of passive (SPATT) and active sampling methods in the profiling and monitoring of marine biotoxins. Toxicon 2014, 89, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, G.; Morono, A.; Paz, B.; Franco, J.M.; Pazos, Y.; Reguera, B. Evaluation of passive samplers as a monitoring tool for early warning of Dinophysis toxins in shellfish. Mar. Drugs 2013, 11, 3823–3845. [Google Scholar] [CrossRef] [PubMed]
- Fux, E.; Marcaillou, C.; Mondeguer, F.; Bire, R.; Hess, P. Field and mesocosm trials on passive sampling for the study of adsorption and desorption behaviour of lipophilic toxins with a focus on OA and DTX1. Harmful Algae 2008, 7, 574–583. [Google Scholar] [CrossRef]
- Rundberget, T.; Gustad, E.; Samdal, I.A.; Sandvik, M.; Miles, C.O. A convenient and cost-effective method for monitoring marine algal toxins with passive samplers. Toxicon 2009, 53, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Zhang, J. Current techniques for detecting and monitoring algal toxins and causative harmful algal blooms. J. Environ. Anal. Chem. 2015, 2, 123. [Google Scholar]
- Hattenrath, T.K.; Anderson, D.M.; Gobler, C.J. The influence of anthropogenic nitrogen loading and meteorological conditions on the dynamics and toxicity of Alexandrium fundyense blooms in a New York (USA) estuary. Harmful Algae 2010, 9, 402–412. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Bill, B.D.; Erickson, A.; Baugh, K.A.; O’Rourke, L.; Costa, P.R.; Nance, S.; Trainer, V.L. Characterization of intracellular and extracellular saxitoxin levels in both field and cultured Alexandrium spp. samples from Sequim Bay, Washington. Mar. Drugs 2008, 6, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Reverté, L.; Soliño, L.; Carnicer, O.; Diogène, J.; Campàs, M. Alternative methods for the detection of emerging marine toxins: Biosensors, biochemical assays and cell-based assays. Mar. Drugs 2014, 12, 5719–5763. [Google Scholar] [CrossRef] [PubMed]
- USFDA. Fish and Fisheries Products Hazards and Controls Guidance, 4th ed.; Department of Health and Human Services, Public Health Service, Food and Drug Administration, Center for Food Safety and Applied Nutrition: Washington, DC, USA, 2011.
- ISSC. Interstate Shellfish Sanitation Conference. National Shellfish Sanitation Program (NSSP): Guide for the Control of Molluscan Shellfish, 2015 Revision. Available online: https://www.fda.gov/downloads/Food/GuidanceRegulation/FederalStateFoodPrograms/UCM505093.pdf (accessed on 9 January 2018).
- Hattenrath-Lehmann, T.K.; Marcoval, M.A.; Mittlesdorf, H.; Goleski, J.A.; Wang, Z.; Haynes, B.; Morton, S.L.; Gobler, C.J. Nitrogenous nutrients promote the growth and toxicity of Dinophysis acuminata during estuarine bloom events. PLoS ONE 2015, 10, e0124148. [Google Scholar] [CrossRef] [PubMed]
- Prud’homme van Reine, W.F. Report of the Nomenclature Committee for Algae: 15. Taxon 2017, 66, 191–192. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Keafer, B.A.; Gribble, K.E.; Marin, R.; Scholin, C.A. Identification and enumeration of Alexandrium spp. from the Gulf of Maine using molecular probes. Deep Sea Res. Part I 2005, 52, 2467–2490. [Google Scholar] [CrossRef]
- John, U.; Litaker, R.W.; Montresor, M.; Murray, S.; Brosnahan, M.L.; Anderson, D.M. Formal revision of the Alexandrium tamarense species complex (Dinophyceae) taxonomy: The introduction of five species with emphasis on molecular-based (rDNA) classification. Protist 2014, 165, 779–804. [Google Scholar] [CrossRef] [PubMed]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis; Horowitz, W., Ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1990; pp. 881–882. [Google Scholar]
- IAEA-TECDOC-1729. RBA for paralytic shellfish poisoning toxins: Saxitoxins. In Detection of Harmful Algal Toxins Using the Radioligand Receptor Binding Assay; International Atomic Energy Agency: Vienna, Austria, 2013. [Google Scholar]
- Lawrence, J.F.; Niedzwiadek, B.; Menard, C. Quantitative determination of paralytic shellfish poisoning toxins in shellfish using prechromatographic oxidation and liquid chromatography with fluorescence detection: Collaborative study. J. AOAC Int. 2005, 88, 1714–1732. [Google Scholar] [PubMed]
- Boundy, M.J.; Selwood, A.I.; Harwood, D.T.; McNabb, P.S.; Turner, A.D. Development of a sensitive and selective liquid chromatography-mass spectrometry method for high throughput analysis of paralytic shellfish toxins using graphitised carbon solid phase extraction. J. Chromatogr. A 2015, 1387, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; McNabb, P.S.; Harwood, D.T.; Selwood, A.I.; Boundy, M.J. Single-laboratory validation of a multitoxin ultra-performance LC-hydrophilic interaction LC-MS/MS method for quantitation of paralytic shellfish toxins in bivalve shellfish. J. AOAC Int. 2015, 98, 609–621. [Google Scholar] [CrossRef] [PubMed]
- Gerssen, A.; Mulder, P.P.J.; McElhinney, M.A.; de Boer, J. Liquid chromatography-tandem mass spectrometry method for the detection of marine lipophilic toxins under alkaline conditions. J. Chromatogr. A 2009, 1216, 1421–1430. [Google Scholar] [CrossRef] [PubMed]
- Gerssen, A.; van Olst, E.H.W.; Mulder, P.P.J.; de Boer, J. In-house validation of a liquid chromatography tandem mass spectrometry method for the analysis of lipophilic marine toxins in shellfish using matrix-matched calibration. Anal. Bioanal. Chem. 2010, 397, 3079–3088. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Quilliam, M.A. LC-MS/MS analysis of Diarrhetic Shellfish Poisoning (DSP) toxins, okadaic acid and Dinophysistoxin analogues, and other lipophilic toxins. Anal. Sci. 2011, 27, 571–584. [Google Scholar] [CrossRef] [PubMed]
- Mountfort, D.O.; Suzuki, T.; Truman, P. Protein phosphatase inhibition assay adapted for determination of total DSP in contaminated mussels. Toxicon 2001, 39, 383–390. [Google Scholar] [CrossRef]
- EU-RL-MB (European Union Reference Laboratory for Marine Biotoxin). EU-Harmonised Standard Operating Procedures for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS; Agencia Española de Consumo, Seguridad Alimentaria y Nutrición (AECOSAN), Ed.; Version 5; Campus Universitario de VIgo. As Lagoas-Marcosende: Vigo, Spain, 2015. [Google Scholar]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Staedler, M.; Miller, W.A.; Toy-Choutka, S.; Dominik, C. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5, e12576. [Google Scholar] [CrossRef] [PubMed]
- EFSA, E. Marine biotoxins in shellfish–okadaic acid and analogues. Scientific Opinion of the Panel on Contaminants in the Food chain. EFSA J. 2008, 589, 1–62. [Google Scholar]
- Smith, J.L.; Tong, M.M.; Fux, E.; Anderson, D.M. Toxin production, retention, and extracellular release by Dinophysis acuminata during extended stationary phase and culture decline. Harmful Algae 2012, 19, 125–132. [Google Scholar] [CrossRef]
- Nagai, S.; Suzuki, T.; Nishikawa, T.; Kamiyama, T. Differences in the production and excretion kinetics of okadaic acid, Dinophysistoxin-1, and pectenotoxin-2 between cultures of Dinophysis acuminata and Dinophysis fortii isolated from western Japan. J. Phycol. 2011, 47, 1326–1337. [Google Scholar] [CrossRef] [PubMed]
- Manage, P.M.; Edwards, C.; Singh, B.K.; Lawton, L.A. Isolation and identification of novel microcystin-degrading bacteria. Appl. Environ. Microbiol. 2009, 75, 6924–6928. [Google Scholar] [CrossRef] [PubMed]
- Shetty, K.G.; Huntzicker, J.V.; Rein, K.S.; Jayachandran, K. Biodegradation of polyether algal toxins–Isolation of potential marine bacteria. J. Environ. Sci. Health Part A 2010, 45, 1850–1857. [Google Scholar] [CrossRef] [PubMed]
- Hattenrath-Lehmann, T.K.; Smith, J.L.; Wallace, R.B.; Merlo, L.R.; Koch, F.; Mittelsdorf, H.; Goleski, J.A.; Anderson, D.M.; Gobler, C.J. The effects of elevated CO2 on the growth and toxicity of field populations and cultures of the saxitoxin-producing dinoflagellate, Alexandrium fundyense. Limnol. Oceanogr. 2015, 60, 198–214. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.M.; Kulis, D.M.; Doucette, G.J.; Gallagher, J.C.; Balech, E. Biogeography of toxic dinoflagellates in the genus Alexandrium from the northeastern United States and Canada. Mar. Biol. 1994, 120, 467–478. [Google Scholar] [CrossRef]
- Gago-Martinez, A.; Rodriguez-Vazquez, J.A.; Thibault, P.; Quilliam, M.A. Simultaneous occurrence of diarrhetic and paralytic shellfish poisoning toxins in Spanish mussels in 1993. Natl. Toxins 1996, 4, 72–79. [Google Scholar] [CrossRef]
- Garcia, C.; Mardones, P.; Sfeir, A.; Lagos, N. Simultaneous presence of Paralytic and Diarrheic Shellfish Poisoning toxins in Mytilus chilensis samples collected in the Chiloe Island, Austral Chilean fjords. Biol. Res. 2004, 37, 721–731. [Google Scholar] [CrossRef]
- Jester, R.; Lefebvre, K.; Langlois, G.; Vigilant, V.; Baugh, K.; Silver, M.W. A shift in the dominant toxin-producing algal species in central California alters phycotoxins in food webs. Harmful Algae 2009, 8, 291–298. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Quakenbush, L.; Frame, E.; Huntington, K.B.; Sheffield, G.; Stimmelmayr, R.; Bryan, A.; Kendrick, P.; Ziel, H.; Goldstein, T.; et al. Prevalence of algal toxins in Alaskan marine mammals foraging in a changing arctic and subarctic environment. Harmful Algae 2016, 55, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean warming since 1982 has expanded the niche of toxic algal blooms in the North Atlantic and North Pacific oceans. Proc. Natl. Acad. Sci. USA 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NPB2B | Particulate DSP Toxin | Dinophysis Densities | DSP Toxins in Shellfish |
|---|---|---|---|
| Particulate DSP toxin | - | - | - |
| Dinophysis densities | 0.99 (p < 0.001, n = 26) | - | - |
| DSP toxins in shellfish | 0.35 (p > 0.05, n = 9) | 0.38 (p < 0.05, n = 9) | - |
| DSP toxins in SPATT | 0 (p > 0.05, n = 11) | 0 (p > 0.05, n = 11) | 0.89 (p < 0.001, n = 8) |
| NPB8B | |||
| Particulate DSP toxin | - | - | - |
| Dinophysis densities | 0.99 (p < 0.001, n = 10) | - | - |
| DSP toxins in shellfish | 0.05 (p > 0.05, n = 8) | 0.18 (p > 0.05, n = 12) | - |
| DSP toxins in SPATT | 0 (p > 0.05, n = 8) | 0 (p > 0.05, n = 12) | 0.70 (p < 0.001, n = 11) |
| Sample ID | Total OA | Recovery |
|---|---|---|
| CRM-OA-c | 1.7 ± 0.1 nM | 113% |
| CRM-DSP-mus-c 1 | 3003 ± 9.3 ng g−1 | 100% |
| CRM-DSP-mus-c 2 | 3013 ± 183 ng g−1 | 100% |
| CRM-DSP-mus-c 3 | 3159 ± 191 ng g−1 | 105% |
| CRM-zero-mus 1 | <63 ng g−1 | |
| CRM-zero-mus 2 | <63 ng g−1 | |
| CRM-zero-mus 3 | <63 ng g−1 |
| Bay | Site | Date | Species | Total DSP ng g−1 | ||
|---|---|---|---|---|---|---|
| PP2A | LCMS | |||||
| Double Exhaustive | Single Dispersive | Double Exhaustive | ||||
| Cold Spring Harbor | CSH1 | 7/10/2013 | Mytilus edulis | 168 | ||
| CSH1 | 7/10/2013 | Crassostrea virginica | 25 | |||
| CSH1 | 7/10/2013 | Mya arenaria | 78 | |||
| CSH1 | 7/7/2014 | Geukensia demissa | 202 | 176 | 149 | |
| CSH1 | 7/14/2014 | Geukensia demissa | 73 | 69 | 39 | |
| CSH1 | 7/21/2014 | Mytilus edulis | 96 | 90 | 73 | |
| CSH1 | 7/21/2014 | Geukensia demissa | <63 | 23 | ||
| CSH2 | 7/7/2014 | Mytilus edulis | 81 | |||
| CSH2 | 7/14/2014 | Mytilus edulis | 89 | |||
| Northport Bay | SB | 7/2/2014 | Geukensia demissa | <63 | 19 | |
| SB | 7/2/2014 | Mya arenaria | <63 | <63 | 22 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hattenrath-Lehmann, T.K.; Lusty, M.W.; Wallace, R.B.; Haynes, B.; Wang, Z.; Broadwater, M.; Deeds, J.R.; Morton, S.L.; Hastback, W.; Porter, L.; et al. Evaluation of Rapid, Early Warning Approaches to Track Shellfish Toxins Associated with Dinophysis and Alexandrium Blooms. Mar. Drugs 2018, 16, 28. https://doi.org/10.3390/md16010028
Hattenrath-Lehmann TK, Lusty MW, Wallace RB, Haynes B, Wang Z, Broadwater M, Deeds JR, Morton SL, Hastback W, Porter L, et al. Evaluation of Rapid, Early Warning Approaches to Track Shellfish Toxins Associated with Dinophysis and Alexandrium Blooms. Marine Drugs. 2018; 16(1):28. https://doi.org/10.3390/md16010028
Chicago/Turabian StyleHattenrath-Lehmann, Theresa K., Mark W. Lusty, Ryan B. Wallace, Bennie Haynes, Zhihong Wang, Maggie Broadwater, Jonathan R. Deeds, Steve L. Morton, William Hastback, Leonora Porter, and et al. 2018. "Evaluation of Rapid, Early Warning Approaches to Track Shellfish Toxins Associated with Dinophysis and Alexandrium Blooms" Marine Drugs 16, no. 1: 28. https://doi.org/10.3390/md16010028
APA StyleHattenrath-Lehmann, T. K., Lusty, M. W., Wallace, R. B., Haynes, B., Wang, Z., Broadwater, M., Deeds, J. R., Morton, S. L., Hastback, W., Porter, L., Chytalo, K., & Gobler, C. J. (2018). Evaluation of Rapid, Early Warning Approaches to Track Shellfish Toxins Associated with Dinophysis and Alexandrium Blooms. Marine Drugs, 16(1), 28. https://doi.org/10.3390/md16010028