Does Osmotic Stress Affect Natural Product Expression in Fungi?

 , , ,
, , ,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Strain and Medium Selection
2.2. Multi-Site Study
2.3. Single-Site Study
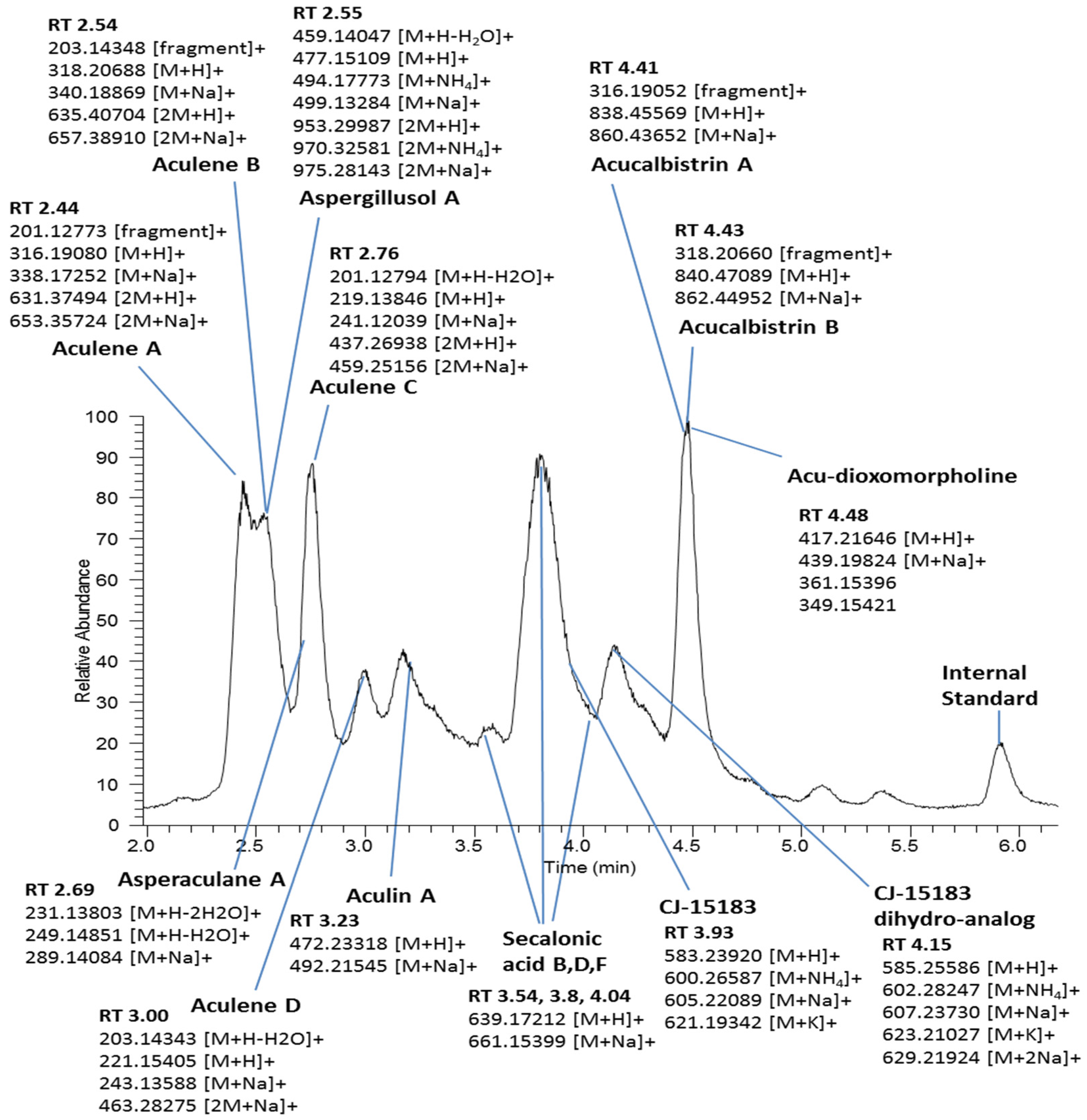
2.4. Extraction and UPLC-HRMS Analysis
2.5. Data Processing
3. Results
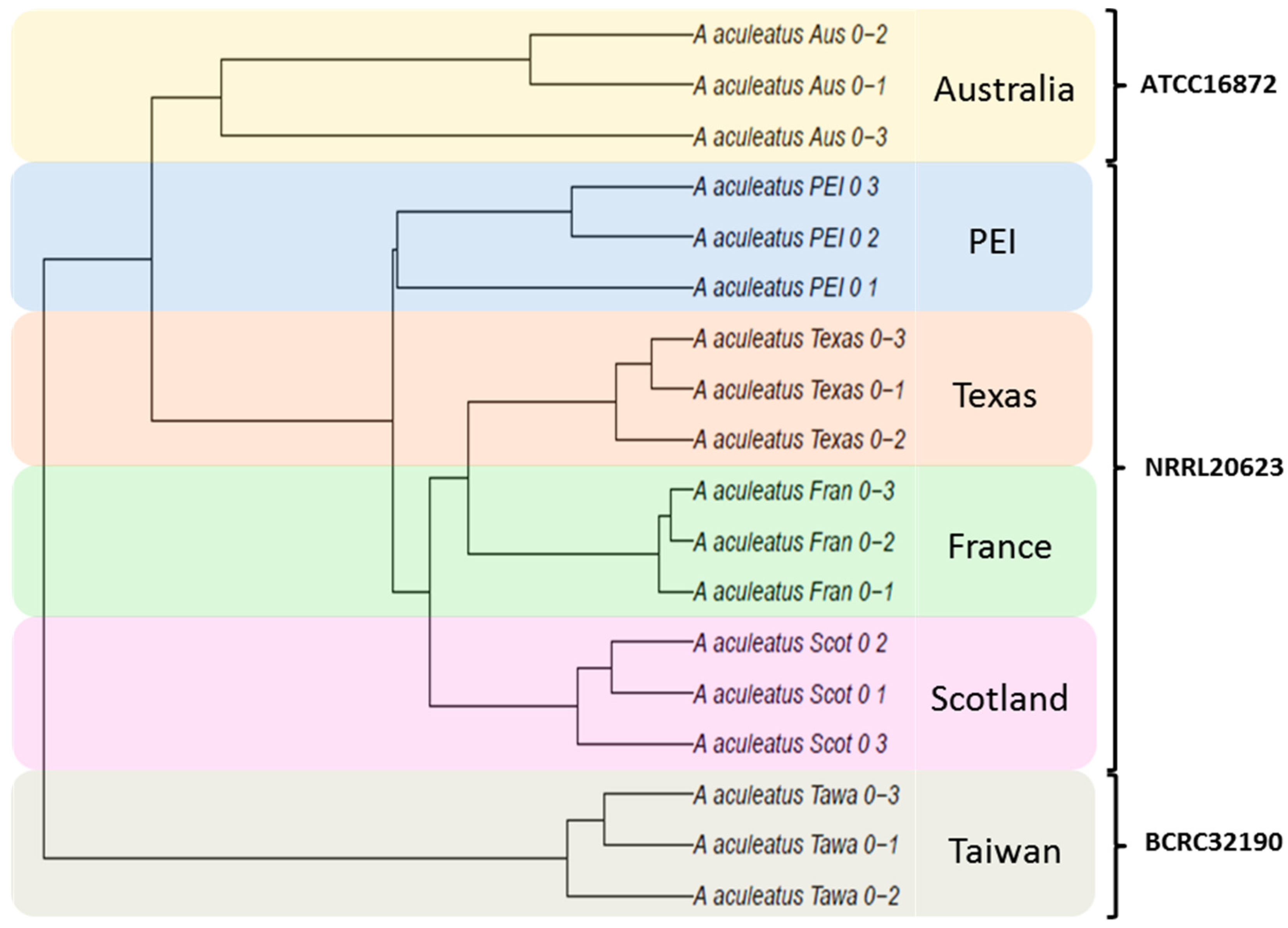
3.1. Characterization of A. aculeatus Ex-Type Strains
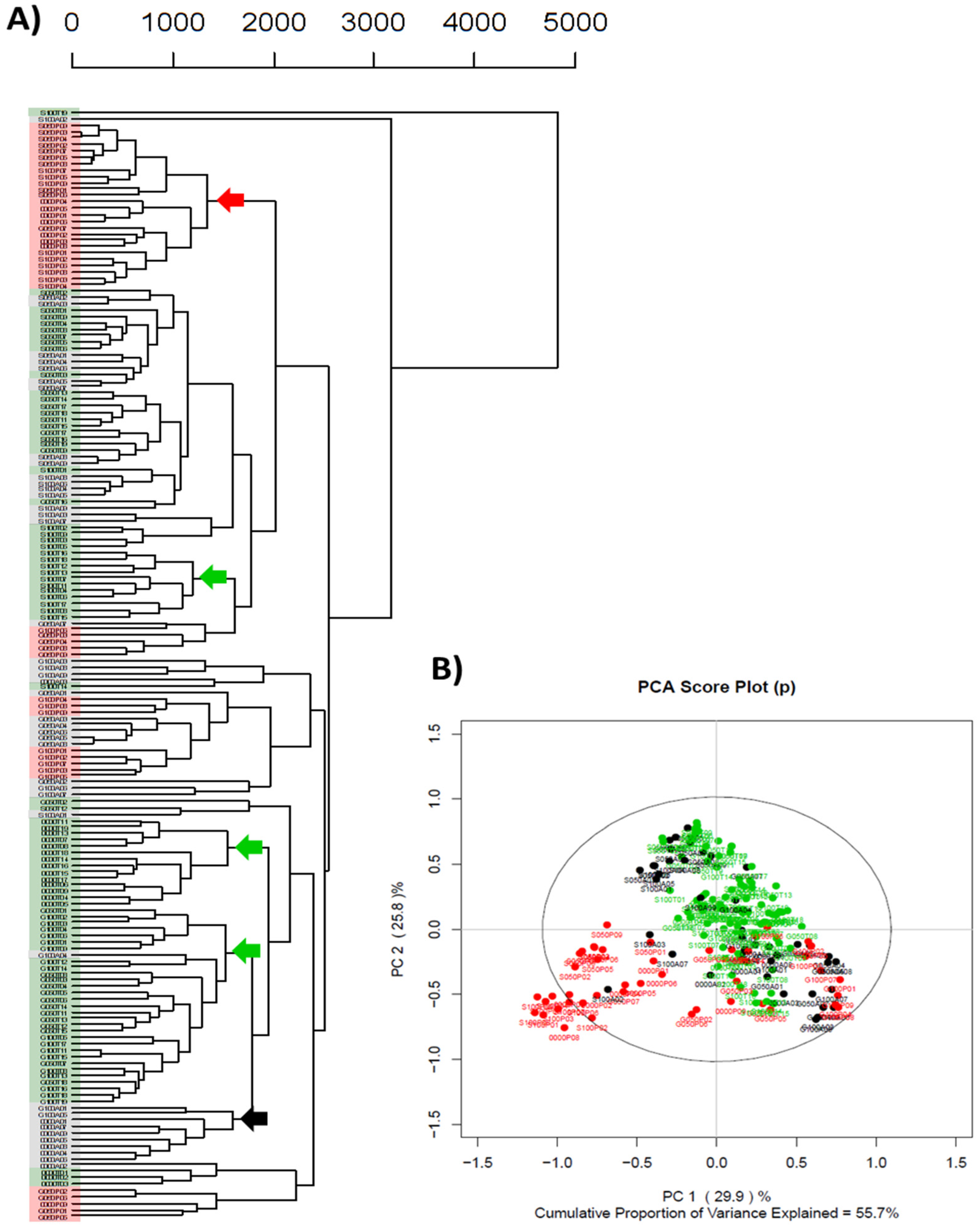
3.2. Multi-Site Study: Summary
3.3. Single-Site Study: Summary
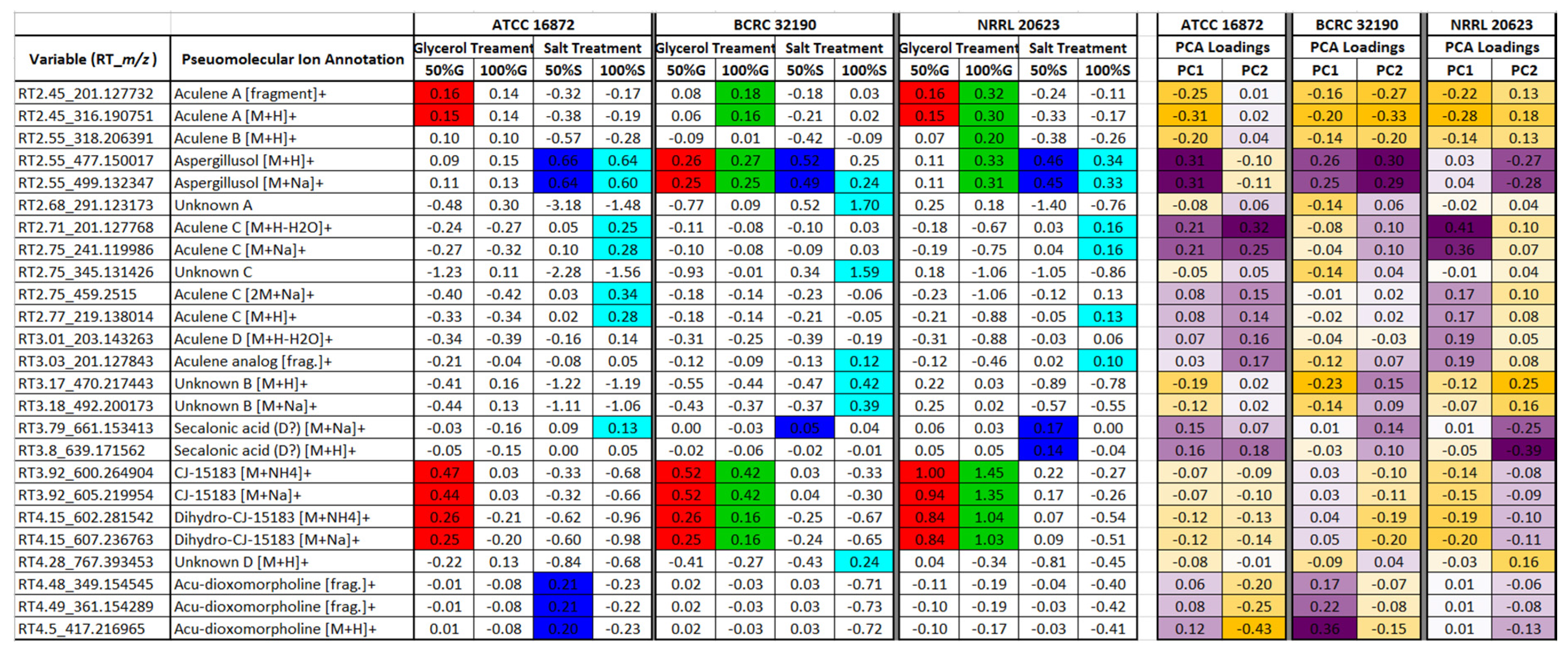
3.4. Single-Site Study: Effects of Osmolite Concentration upon Secondary Metabolism
4. Discussion
4.1. Site-to-Site Variability
4.2. Differences in Ex-Type Strain Secondary Metabolism
4.3. Effect of Increased Salinity/Osmotic Stress on Secondary Metabolism
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Secondary Metabolism and Differentiation in Fungi; Bennett, J.W., Ciegler, A., Eds.; Marcel Dekker: New York, NY, USA, 1983. [Google Scholar]
- Demain, A.L. Regulation of secondary metabolism in fungi. Pure Appl. Chem. 1986, 58, 219–226. [Google Scholar] [CrossRef]
- Lim, F.; Sanchez, J.; Wang, C.C.; Keller, N. Toward awakening cryptic secondary metabolite gene clusters in filamentous fungi. Methods Enzymol. 2012, 517, 303–324. [Google Scholar] [PubMed]
- Keller, N.P. Translating biosynthetic gene clusters into fungal armor and weaponry. Nat. Chem. Biol. 2015, 11, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Pang, K.-L.; Overy, D.P.; Jones, E.B.G.; Calado, M.; Burgaud, G.; Walker, A.K.; Johnson, J.A.; Kerr, R.G.; Cha, H.-J.; Bills, G.F. ‘Marine fungi’ and ‘marine-derived fungi’ in natural product chemistry research: Toward a new consensual definition. Fungal Biol. Rev. 2016, 30, 163–175. [Google Scholar] [CrossRef]
- Lenassi, M.; Gostinčar, C.; Jackman, S.; Turk, M.; Sadowski, I.; Nislow, C.; Jones, S.; Birol, I.; Gunde-Cimerman, N. Whole genome duplication and enrichment of metal cation transporters revealed by de novo genome sequencing of extremely halotolerant black yeast Hortaea werneckii. PLoS ONE 2013, 8, e71328. [Google Scholar] [CrossRef] [PubMed]
- Zajc, J.; Liu, Y.; Dai, W.; Yang, Z.; Hu, J.; Gostincar, C.; Gunde-Cimerman, N. Genome and transcriptome sequencing of the halophilic fungus Wallemia ichthyophaga : Haloadaptations present and absent. BMC Genom. 2013, 14, 617. [Google Scholar] [CrossRef] [PubMed]
- Kis-Papo, T.; Weig, A.R.; Riley, R.; Peršoh, D.; Salamov, A.; Sun, H.; Lipzen, A.; Wasser, S.P.; Rambold, G.; Grigoriev, I.V.; et al. Genomic adaptations of the halophilic Ded Sea filamentous fungus Eurotium rubrum. Nat. Commun. 2014, 5, 3745. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lu, Z.; Sun, K.; Zhu, W. Effects of high salt stress on secondary metabolite production in the marine-derived fungus Spicaria elegans. Marine Drugs 2011, 9, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zheng, J.; Liu, P.; Wang, W.; Zhu, W. Three new compounds from Aspergillus terreus PT06–2 grown in a high salt medium. Marine Drugs 2011, 9, 1368–1378. [Google Scholar] [CrossRef] [PubMed]
- Bugni, T.; Ireland, C. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2012, 28, 290–344. [Google Scholar] [CrossRef] [PubMed]
- Overy, D.P.; Bayman, P.; Kerr, R.G.; Bills, G.F. An assessment of natural product discovery from marine (sensu strictu) and marine-derived fungi. Mycology 2014, 5, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Bérdy, J. Thoughts and facts about antibiotics: Where we are now and where we are heading. J. Antibiot. 2012, 65, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Capon, R.J.; Ratnayake, R.; Stewart, M.; Lacey, E.; Tennant, S.; Gill, J.H. Aspergillaxines A–E: Novel heterocyclic dipeptides from an Australian strain of Aspergillus unilateralis. Org. Biomol. Chem. 2005, 3, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.Y.S.; Arbell, L.M.; Avelar, A.; Borgeson, B.M.; Crews, P. New hisutane based sesquiterpenes from salt water cultures of a marine sponge-derived fungus and the terrestrial fungus Coriolus consors. Tetrahedron 1998, 54, 7335–7342. [Google Scholar] [CrossRef]
- The draft genome sequence of A. aculeatus. Available online: http://genome.jgi.doe.gov/Aspac1/Aspac1.home.html (accessed on 15 August 2016).
- De Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [CrossRef] [PubMed]
- Gaude, E.; Chignola, F.; Spiliotopoulus, D.; Spitaleri, A.; Ghitti, M.; Garcia-Manteiga, J.M.; Mari, S.; Musco, G. muma, an R package for metabolomics univariate and multivariate statistical analysis. Curr. Metab. 2013, 1, 180–189. [Google Scholar] [CrossRef]
- Anderson, R.; Buchi, G.; Kobbe, B.; Demain, A.L. Secalonic acids D and F are toxic metabolites of Aspergillus aculeatus. J. Org. Chem. 1977, 42, 352–353. [Google Scholar] [CrossRef]
- Gao, Y.-Q.; Guo, C.-J.; Zhang, Q.; Zhou, W.-M.; Wang, C.C.C.; Gao, J.-M. Asperaculanes A and, B.; two sesquiterpenoids from the fungus Aspergillus aculeatus. Molecules 2015, 20, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Ingavat, N.; Dobereiner, J.; Wiyakrutta, S.; Mahidol, C.; Ruchirawat, S.; Kittakoop, P. Aspergillusol A., a α-glucosidase inhibitor from the marine-derived fungus Aspergillus aculeatus. J. Nat. Prod. 2009, 72, 2049–2052. [Google Scholar] [CrossRef] [PubMed]
- Kurobane, I.; Vining, L.C. Biosynthetic realtionships among the secalonic acids. Isolation of emodin, endocrocin and secalonic acids from Pyrenochaeta terrestris and Aspergillus aculeatus. J. Antibiot. 1979, 32, 1256–1266. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.M.; Hoeck, C.; Frisvad, J.C.; Gotfredsen, C.H.; Larsen, T.O. Dereplication guided discovery of secondary metabolites of mixed biosynthetic origin from Aspergillus aculeatus. Molecules 2014, 19, 10898–10921. [Google Scholar] [CrossRef] [PubMed]
- Petersen, L.M.; Holm, D.K.; Gotfredsen, C.H.; Mortensen, U.H.; Larsen, T.O. Investigation of a 6-MSA synthase gene cluster in Aspergillus aculeatus reveals 6-MSA-derived aculinic acid, aculins A–B and epi-aculin, A. ChemBioChem 2015, 16, 2200–2204. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Hirai, H.; Ishiguro, M.; Kambara, T.; Kojima, Y.; Matsunaga, T.; Nishida, H.; Suzuki, Y.; Sugiura, A.; Harwood, H.J.; et al. CJ-15,183, a new inhibitor of squalene synthase produced by a fungus, Aspergillus aculeatus. J. Antibiot. 2001, 54, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Keller, B.O.; Sui, J.; Young, A.B.; Whittal, R.M. Interferences and contaminants encountered in modern mass spectrometery. Anal. Chim. Acta 2008, 627, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.J.; Smith, D. Fungal genetic resource centres and the genomic challenge. Mycol. Res. 2004, 1088, 1351–1362. [Google Scholar] [CrossRef]
- Kuhls, K.; Lieckfeldt, E.; Börner, T. PCR-fingerprinting used for comparison of ex type strains of Trichoderma species deposited in different culture collections. Microbiol. Res. 1995, 150, 363–371. [Google Scholar] [CrossRef]
- Samson, R.A.; Noonim, P.; Meijer, M.; Houbraken, J.; Frisvad, J.C.; Varga, J. Diagnostic tools to identify black aspergilli. Stud. Mycol. 2007, 59, 129–145. [Google Scholar] [CrossRef] [PubMed]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Bills, G.F.; Platas, G.; Fillola, A.; Jiménez, M.R.; Collado, J.; Vicente, F.; Martín, J.; González, A.; Bur-Zimmermann, J.; Tormo, J.R.; et al. Enhancement of antibiotic and secondary metabolite detection from filamentous fungi by growth on nutritional arrays. J. Appl. Microbiol. 2008, 104, 1644–1658. [Google Scholar] [CrossRef] [PubMed]
- Filtenborg, O.; Frisvad, J.C.; Thrane, U. The significance of yeast extract composition on metabolite production in Penicillium. In Modern Concepts in Penicillium and Aspergillus Classification; Samson, R.A., Pitt, J.I., Eds.; Plenum Press: New York, NY, USA, 1990; pp. 433–441. [Google Scholar]
- Ryan, M.J.; Bridge, P.D.; Smith, D.; Jeffries, P. Phenotypic degeneration occurs during sector formation in Metarhizium anisopliae. J. Appl. Microbiol. 2002, 93, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.J.; Smith, D.; Bridge, P.D.; Jeffries, P. The relationship between fungal preservation method and secondary metabolite production in Metarhizium anisopliae and Fusarium oxysporum. World J. Microbiol. Biotechnol. 2003, 19, 839–844. [Google Scholar] [CrossRef]
- Santos, I.M.; Abrunhosa, L.; Venancio, A.; Lima, N. The effect of culture preservation techniques on patulin and citrinin production by Penicillium expansum Link. Lett. Appl. Microbiol. 2002, 35, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Wing, N.; Burgess, L.W.; Bryden, W.L. Cultural degeneration in two Fusarium species and its effects on toxigenicity and cultural morphology. Mycol. Res. 1995, 99, 615–620. [Google Scholar] [CrossRef]
- Wicklow, D.T. Metabolites in the coevolution of fungal chemical defence systems. In Coevolution of Fungi with Plants and Animals; Pirozynski, K.A., Hawksworth, D.L., Eds.; Academic Press Limited: London, UK, 1988; pp. 173–201. [Google Scholar]
- Anderson, R.F.; Whitmore, L.M.; Brown, W.E.; Peterson, W.H.; Churchill, B.W.; Roegner, F.R.; Campbell, T.H.; Backus, M.P.; Stauffer, J.F. Penicillin production by pigment-free molds. Ind. Eng. Chem. 1953, 45, 768–773. [Google Scholar] [CrossRef]
- Backus, M.P.; Stauffer, J.F. The production and selection of a family of strains in Penicillium chrysogenum. Mycologia 1955, 47, 429–463. [Google Scholar] [CrossRef]
- Raper, K.B.; Fennel, D.I. The Genus Aspergillus; Williams & Wilkins Co.: Baltimore, MD, USA, 1965; pp. 1–686. [Google Scholar]
- Yue, Q.; Chen, L.; Zhang, X.; Li, K.; Sun, J.; Liu, X.; An, Z.; Bills, G.F. Evolution of chemical diversity in the echinocandin lipopeptide antifungal metabolites. Eukaryot. Cell 2015, 14, 698–718. [Google Scholar] [CrossRef] [PubMed]
- Jennings, D.H. Fungal growth in the sea. In The Biology of Marine Fungi; Moss, S.T., Ed.; Cambridge University Press: Cambridge, UK, 1986; pp. 1–10. [Google Scholar]
- Liu, S.; Li, J.; Wu, Y.; Ren, Y.; Liu, Q.; Wang, Q.; Zhou, X.; Cai, M.; Zhang, Y. De novo transcriptome sequencing of marine-derived Aspergillus glaucus and comparative analysis of metabolic and developmental variations in response to salt stress. Genes Genom. 2017, 39, 317–329. [Google Scholar] [CrossRef]












© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Overy, D.; Correa, H.; Roullier, C.; Chi, W.-C.; Pang, K.-L.; Rateb, M.; Ebel, R.; Shang, Z.; Capon, R.; Bills, G.; et al. Does Osmotic Stress Affect Natural Product Expression in Fungi? Mar. Drugs 2017, 15, 254. https://doi.org/10.3390/md15080254
Overy D, Correa H, Roullier C, Chi W-C, Pang K-L, Rateb M, Ebel R, Shang Z, Capon R, Bills G, et al. Does Osmotic Stress Affect Natural Product Expression in Fungi? Marine Drugs. 2017; 15(8):254. https://doi.org/10.3390/md15080254
Chicago/Turabian StyleOvery, David, Hebelin Correa, Catherine Roullier, Wei-Chiung Chi, Ka-Lai Pang, Mostafa Rateb, Rainer Ebel, Zhuo Shang, Rob Capon, Gerald Bills, and et al. 2017. "Does Osmotic Stress Affect Natural Product Expression in Fungi?" Marine Drugs 15, no. 8: 254. https://doi.org/10.3390/md15080254
APA StyleOvery, D., Correa, H., Roullier, C., Chi, W.-C., Pang, K.-L., Rateb, M., Ebel, R., Shang, Z., Capon, R., Bills, G., & Kerr, R. (2017). Does Osmotic Stress Affect Natural Product Expression in Fungi? Marine Drugs, 15(8), 254. https://doi.org/10.3390/md15080254