
Salt Sensitive Tet-Off-Like Systems to Knockdown Primordial Germ Cell Genes for Repressible Transgenic Sterilization in Channel Catfish, Ictalurus punctatus

 ,
,
Abstract
:1. Introduction
2. Results
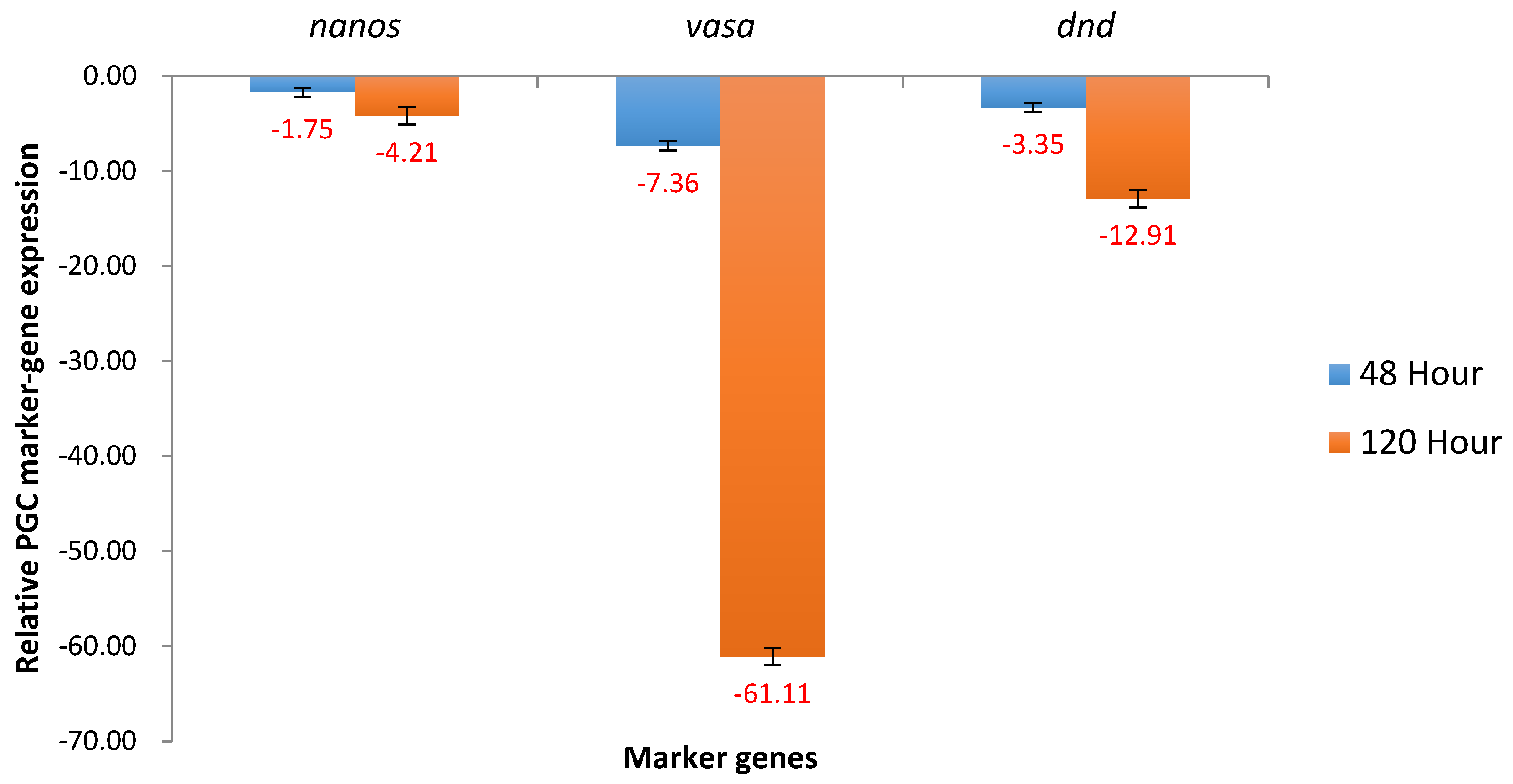
2.1. Expression of Nanos, Vasa and Dead End in Channel Catfish Embryos
2.2. P1 Spawning
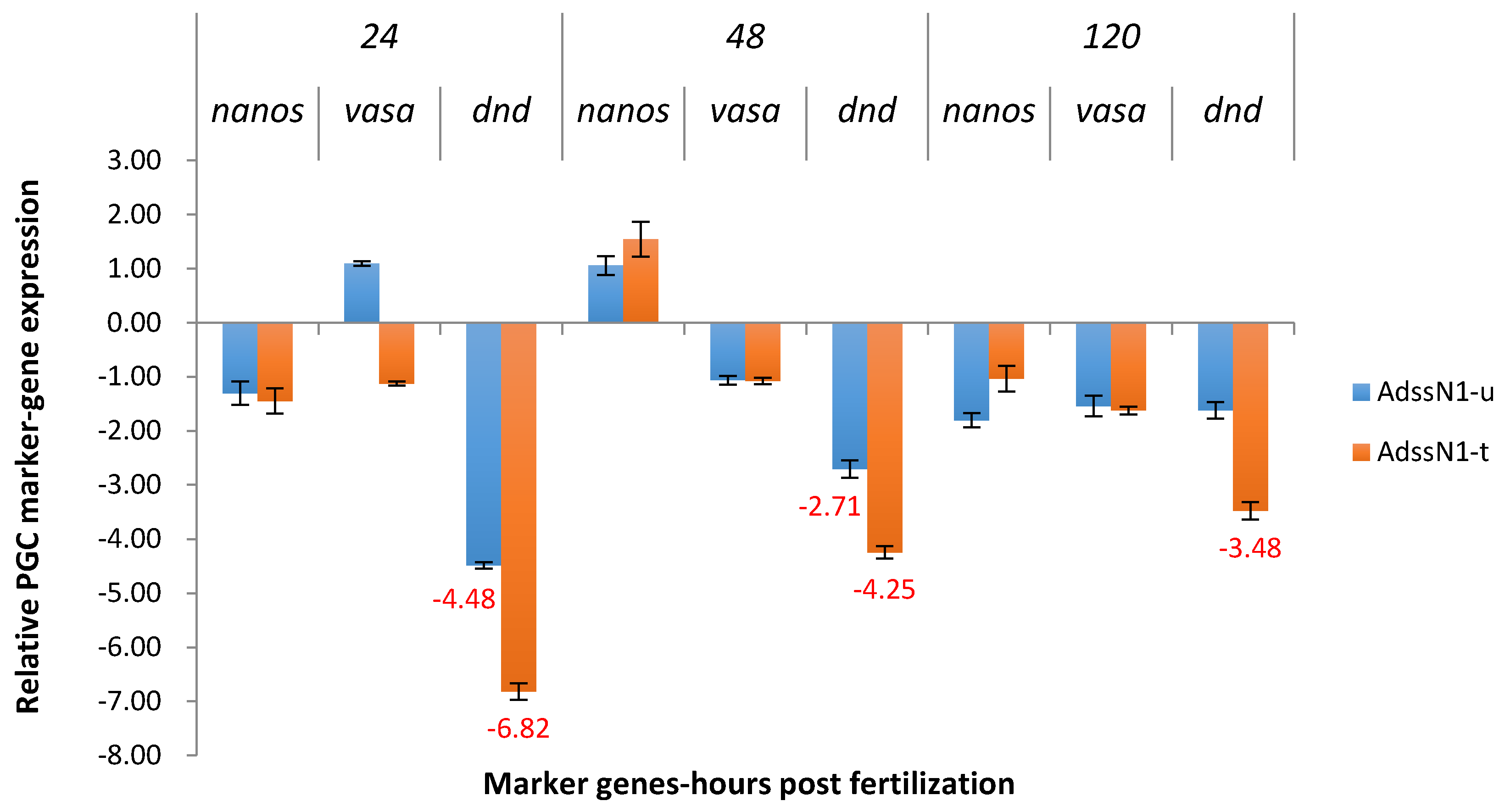
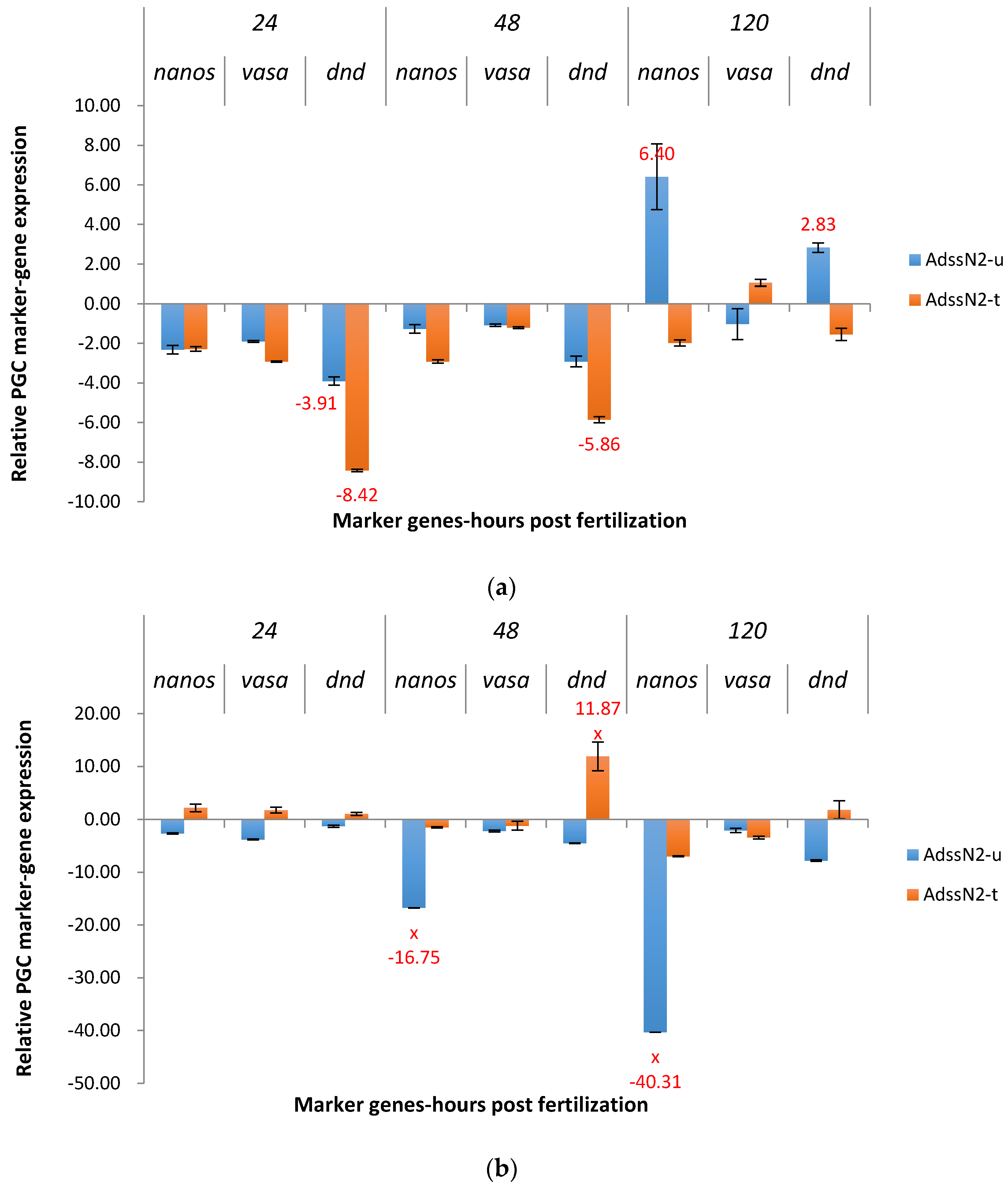
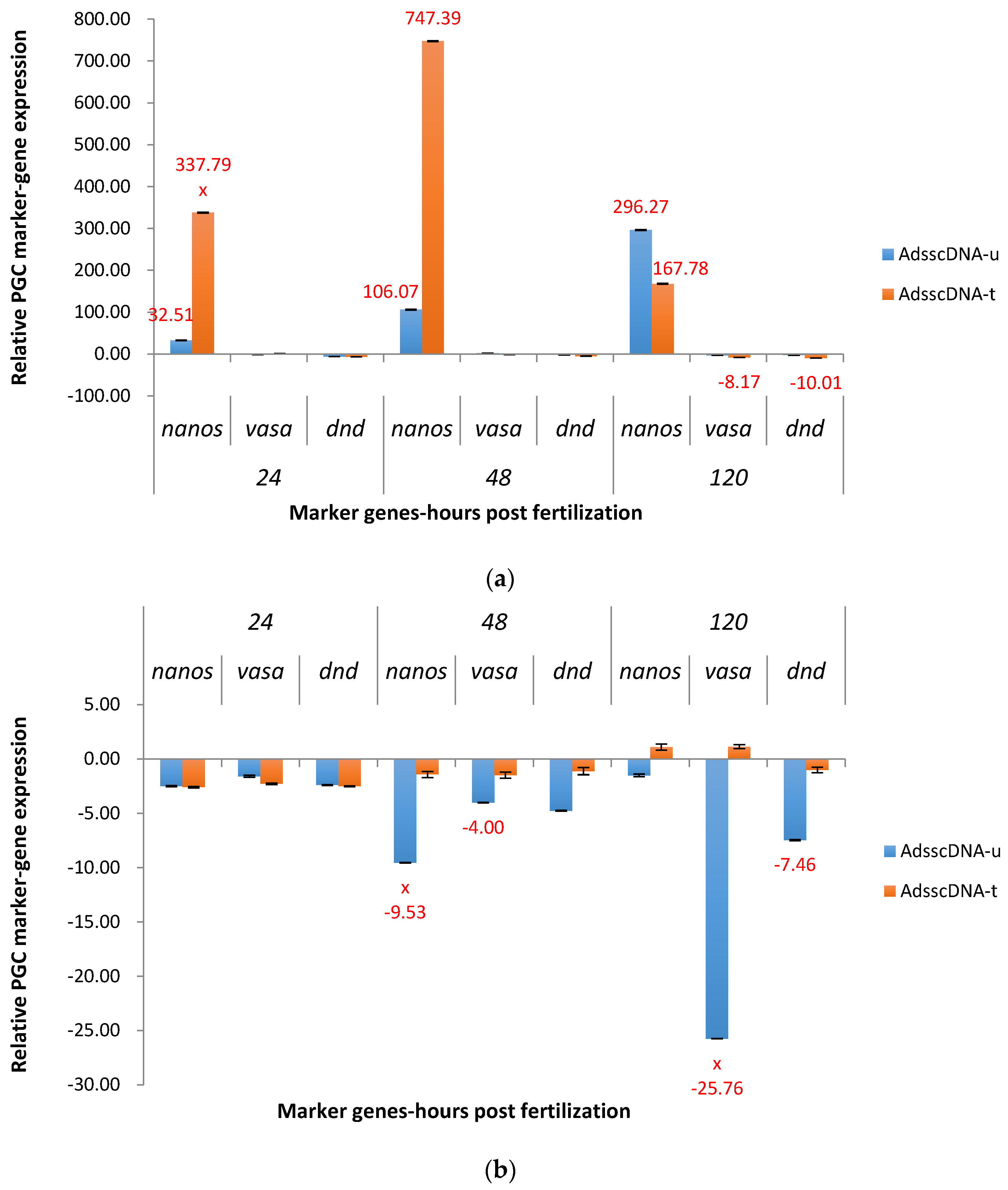
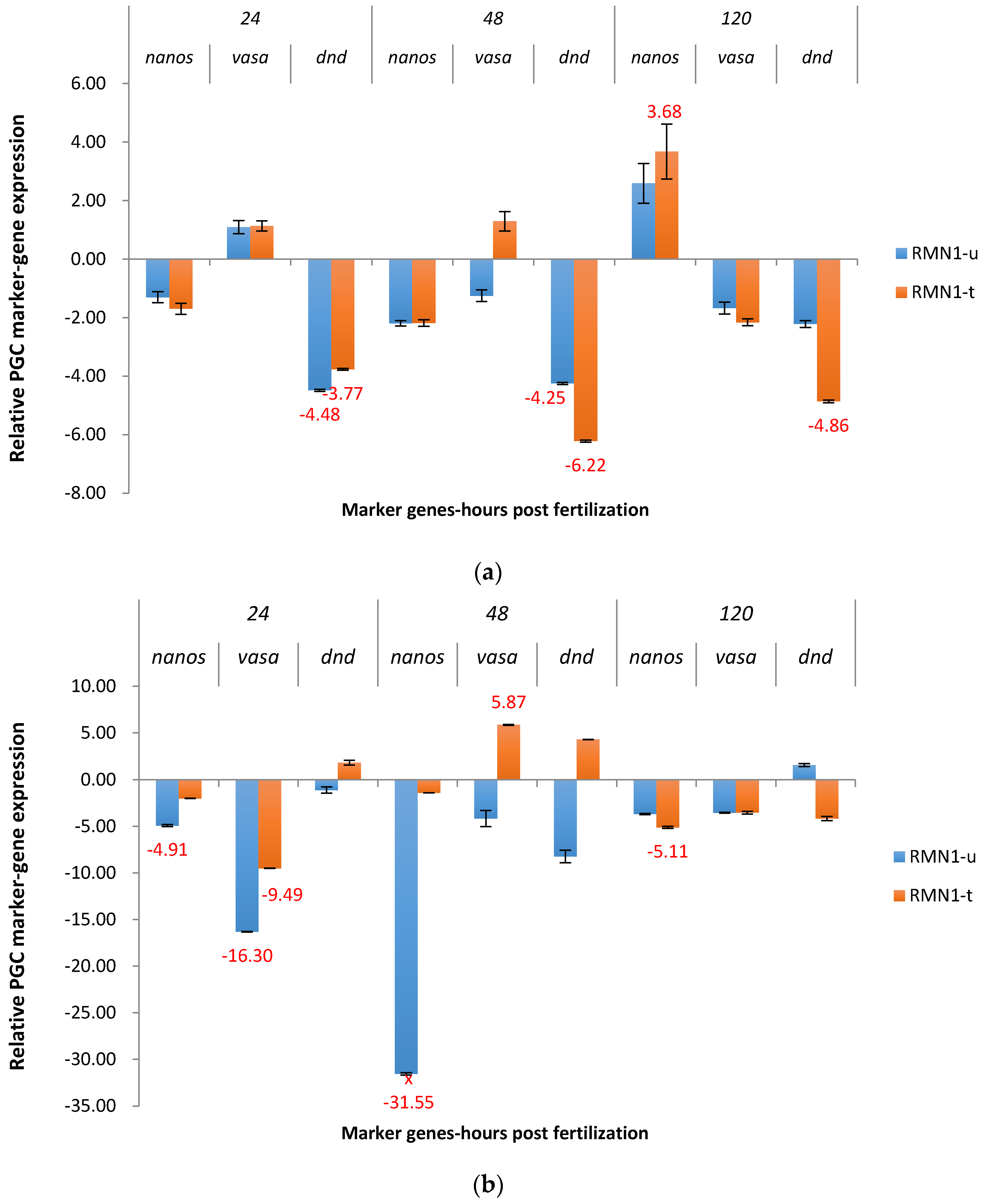
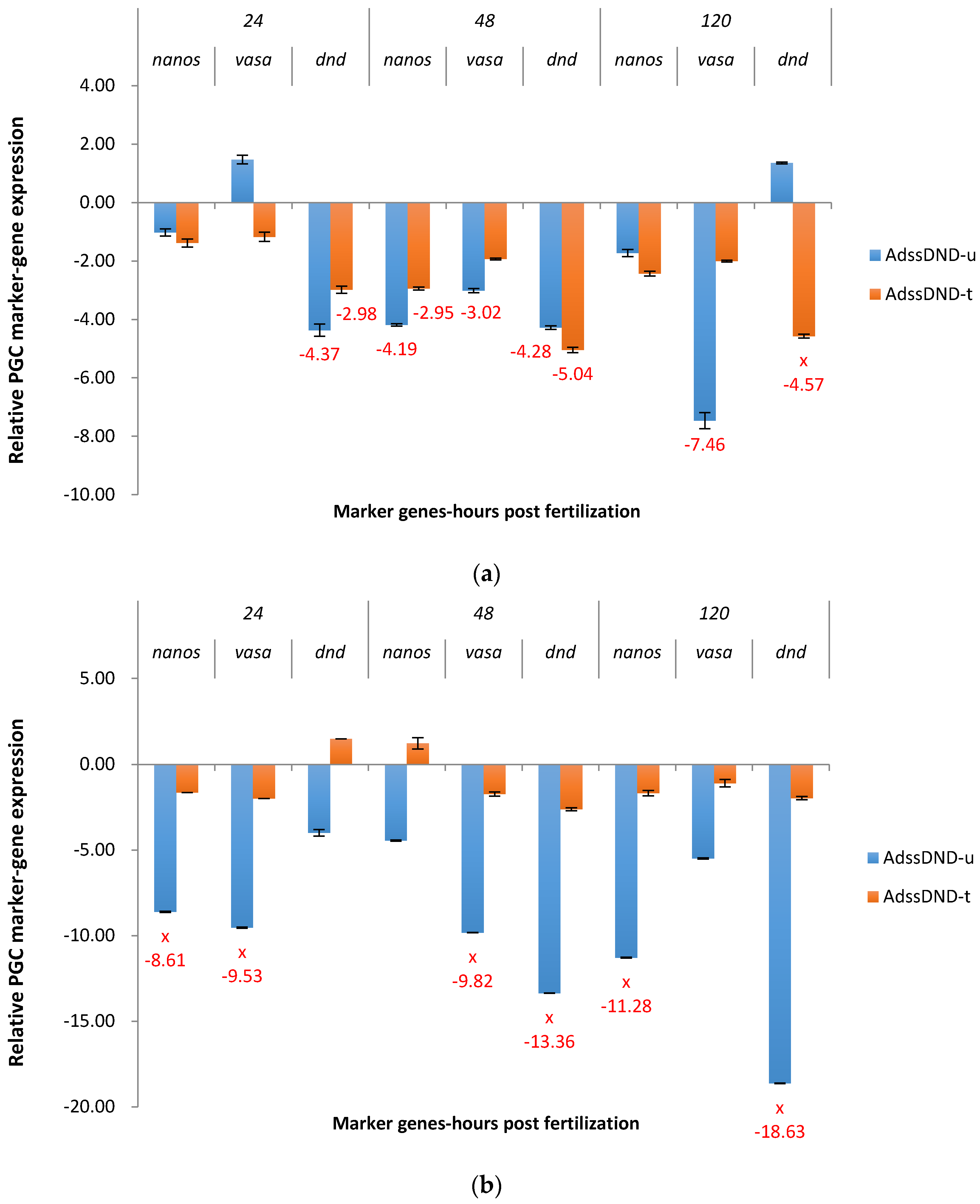
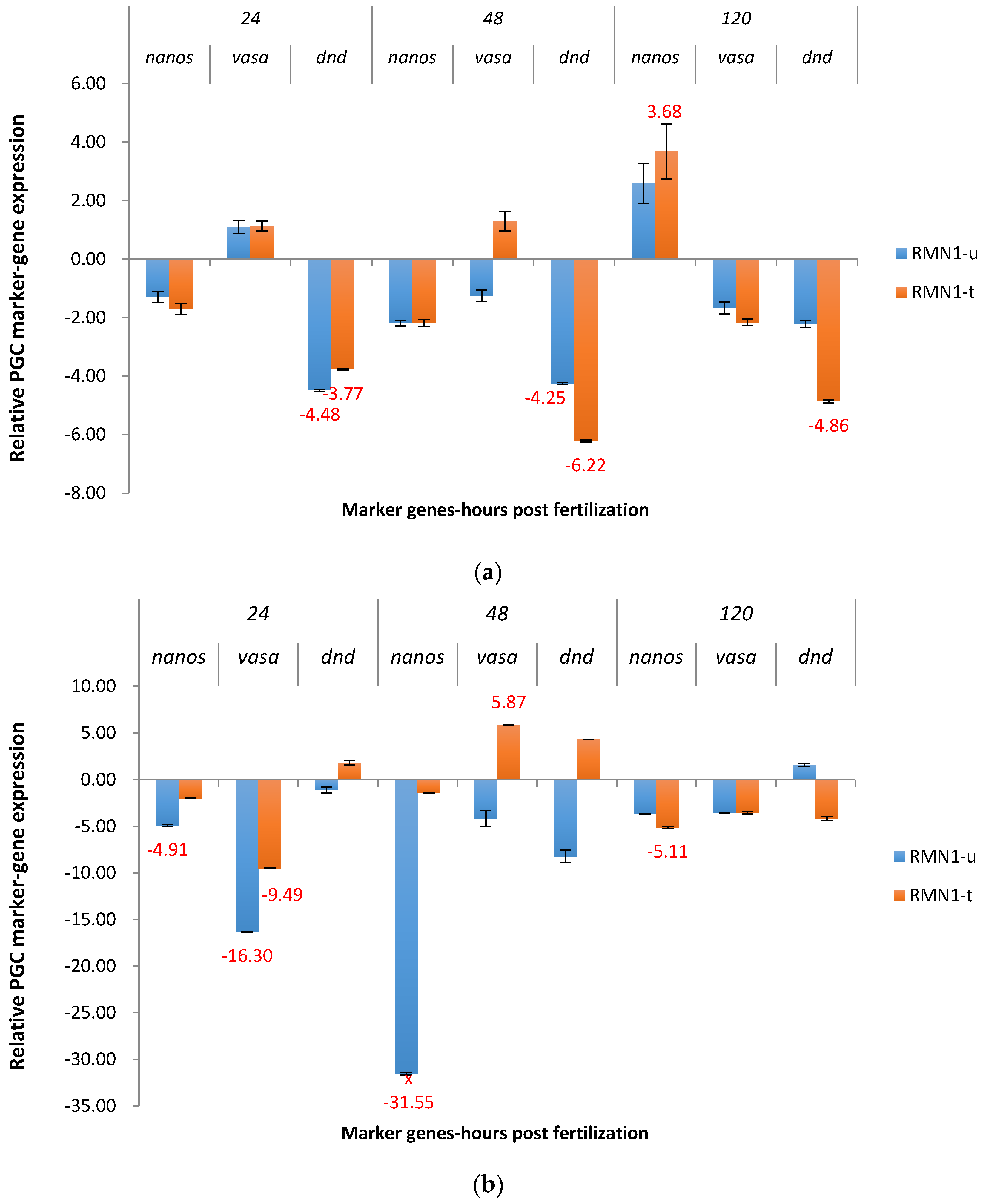
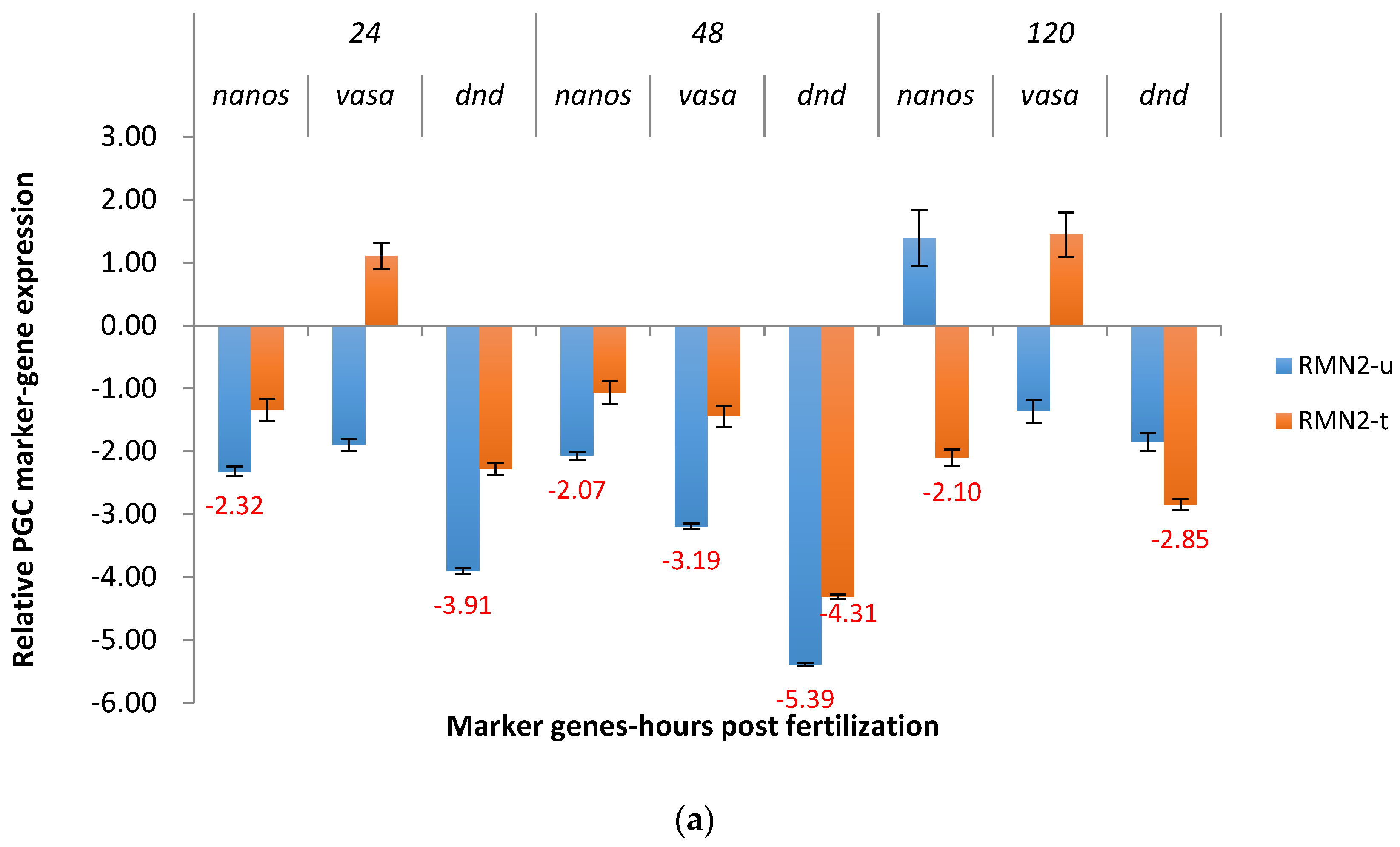
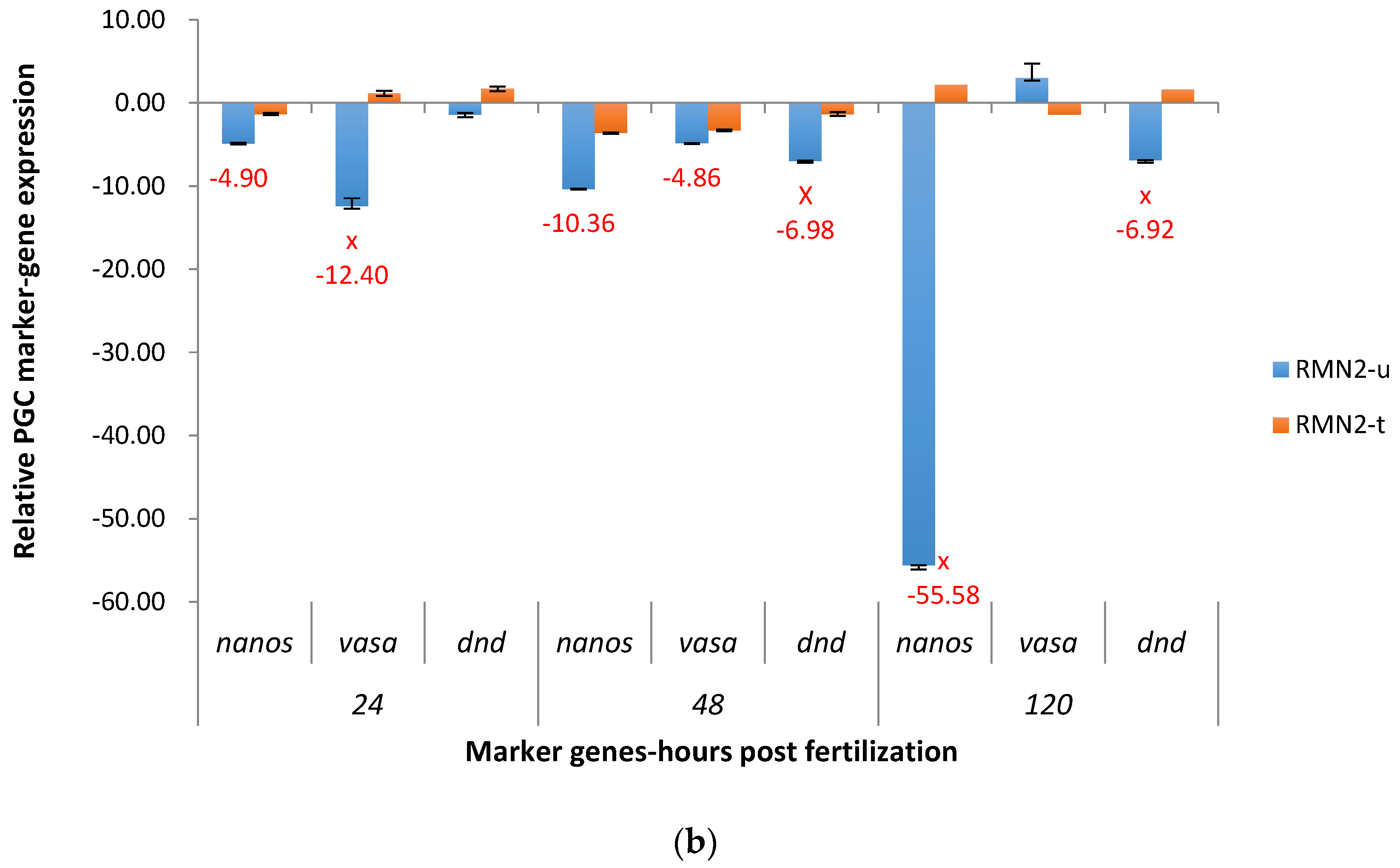
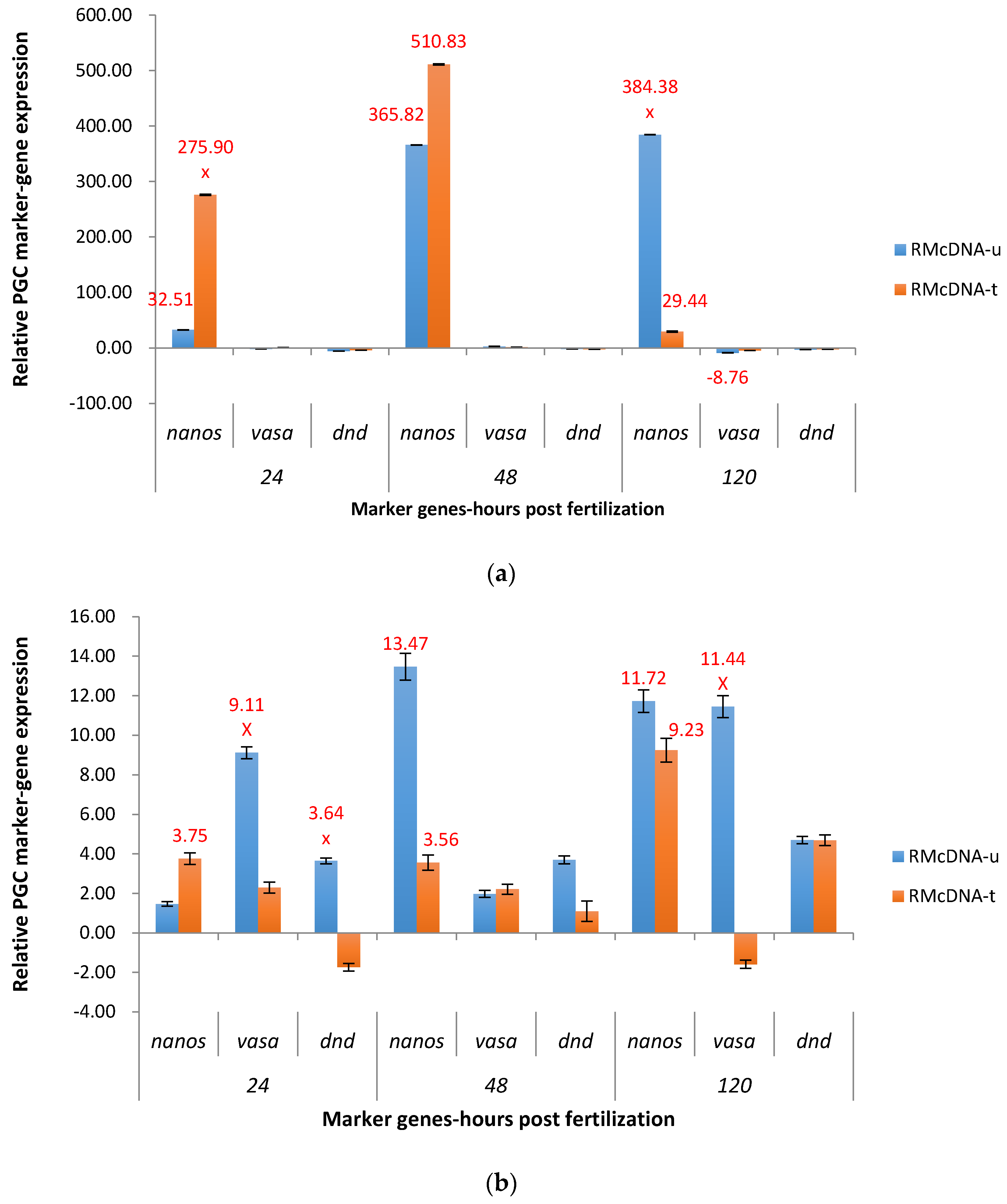
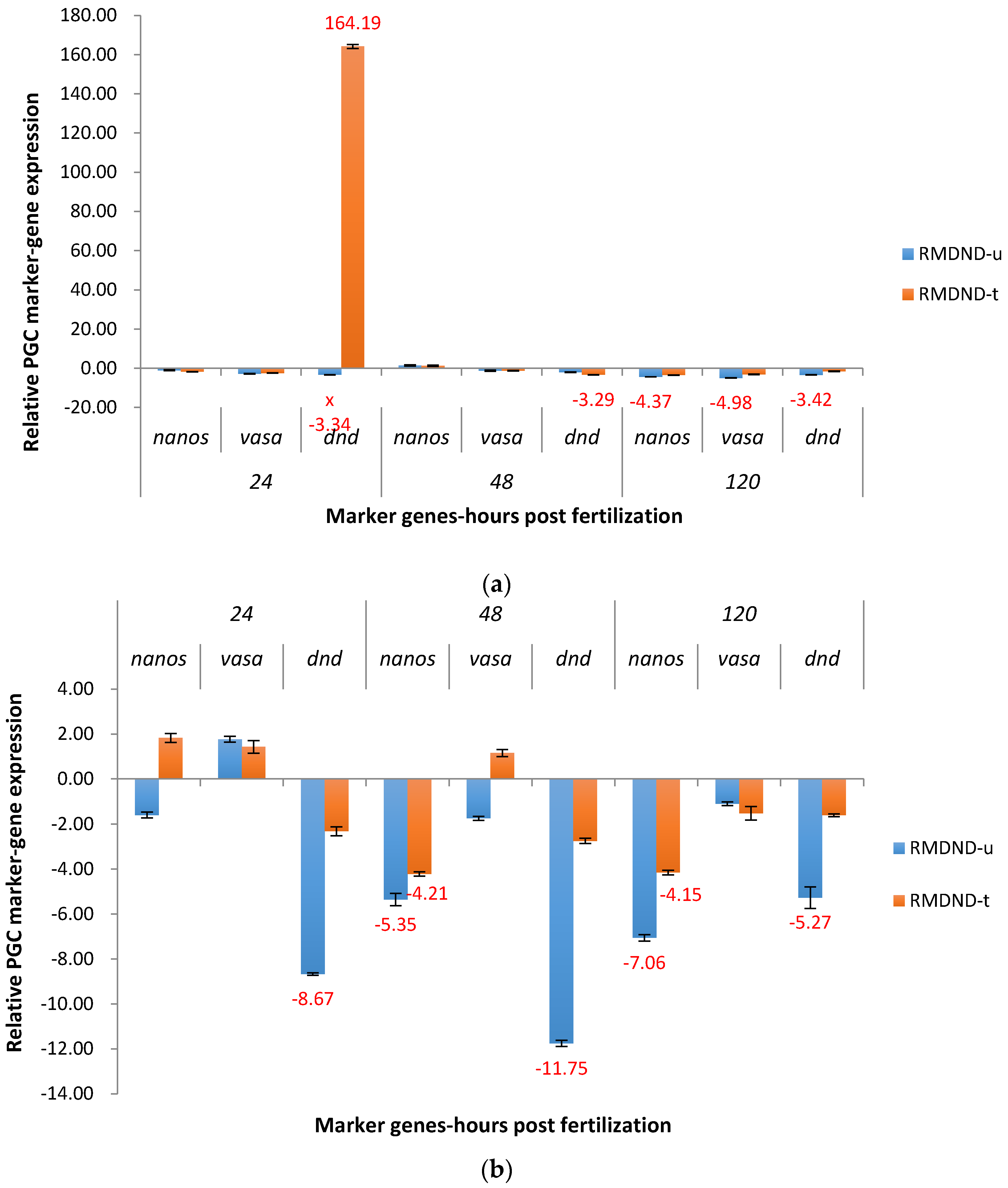
2.3. Real-Time Quantitative PCR Results for the P1 and F1 Generation Transgenic Channel Catfish
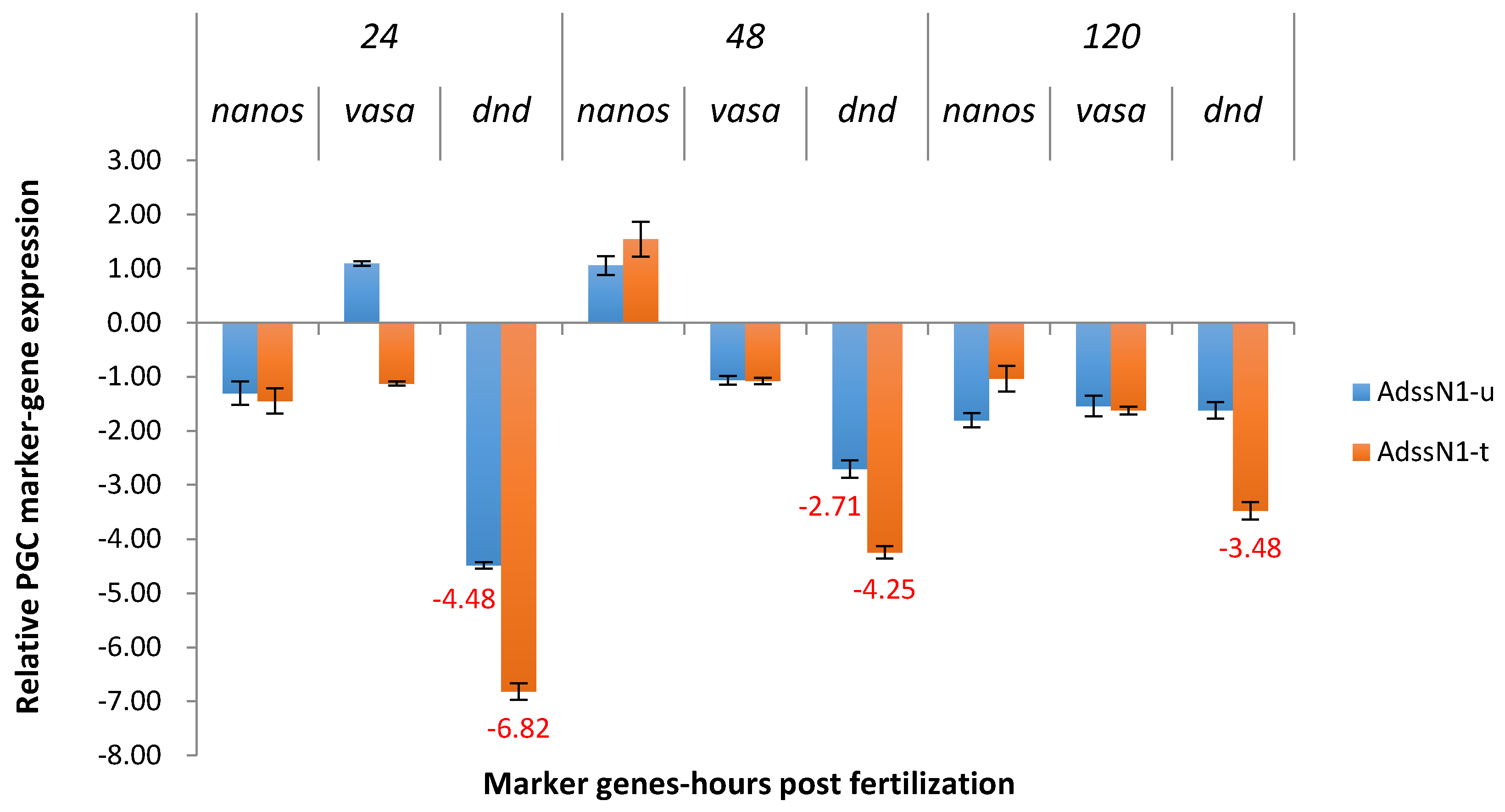
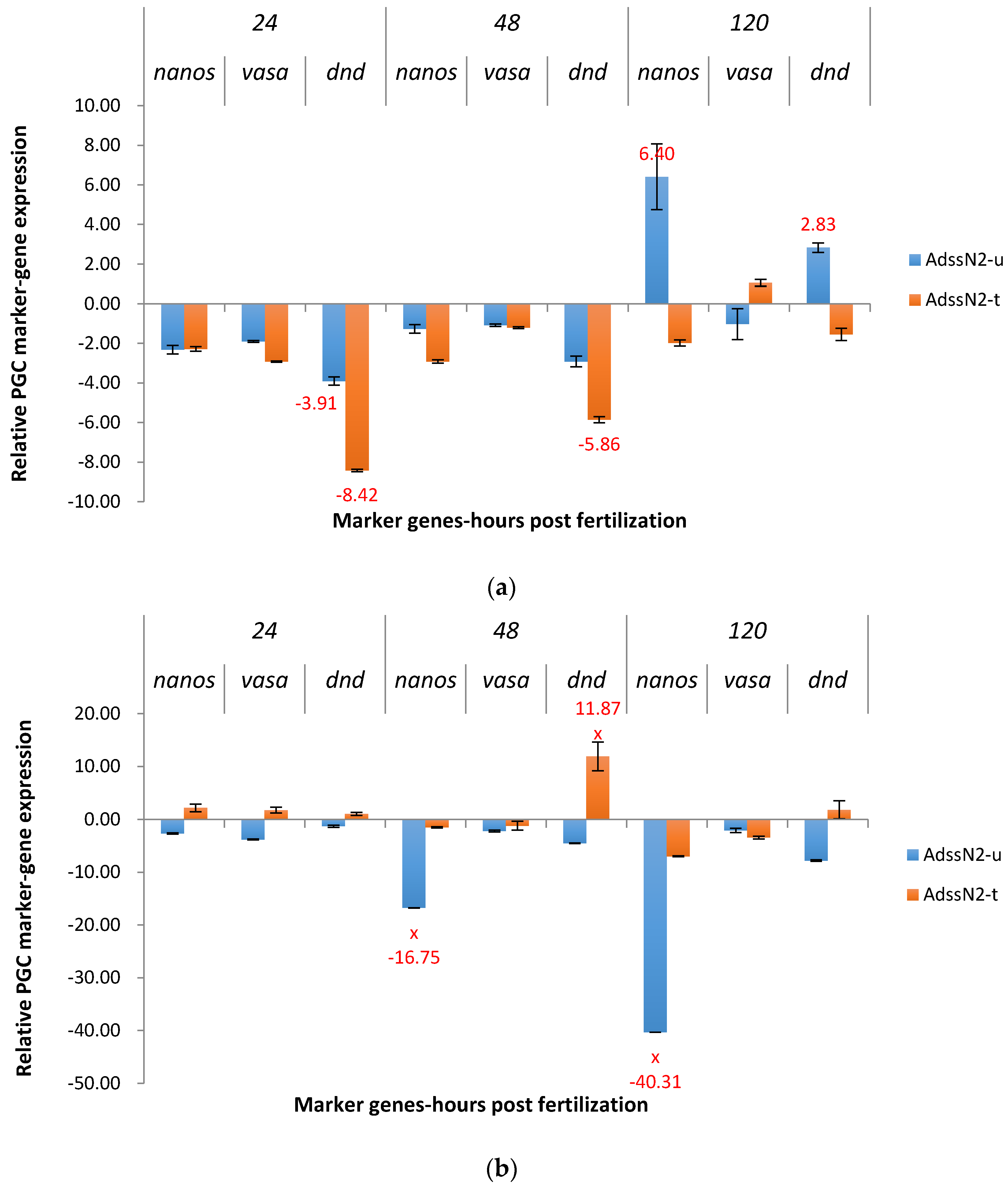
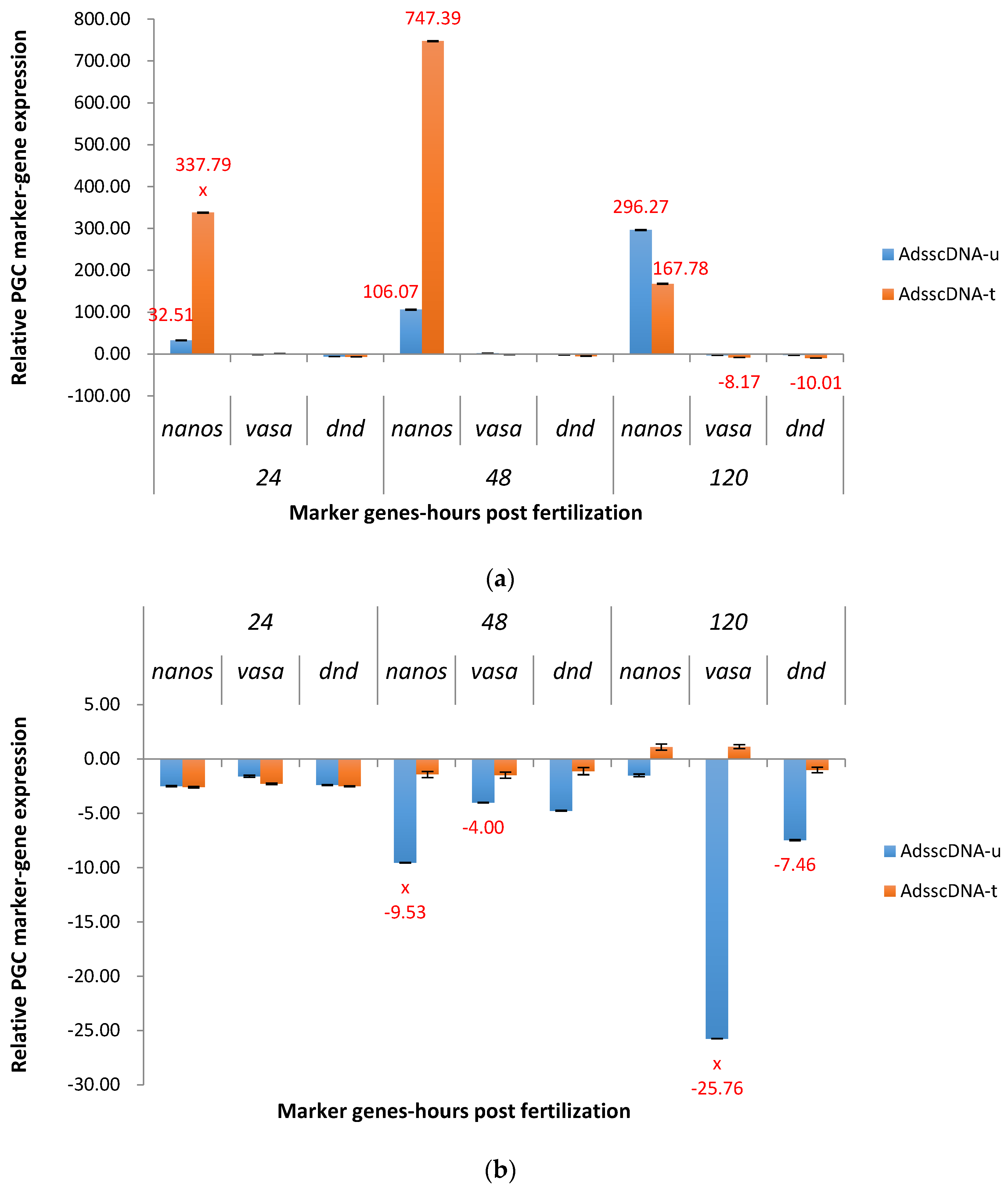
2.3.1. ADSS System
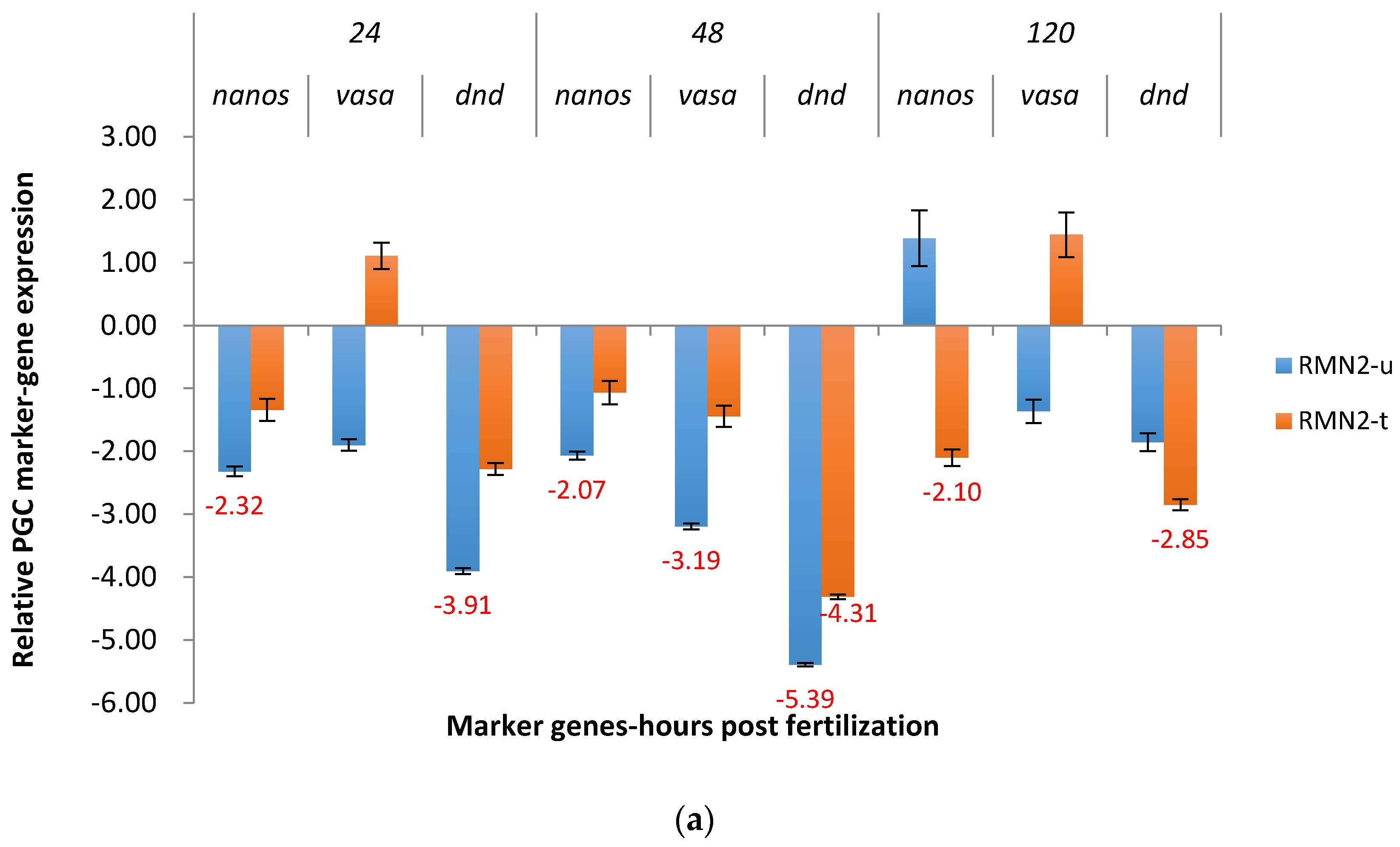
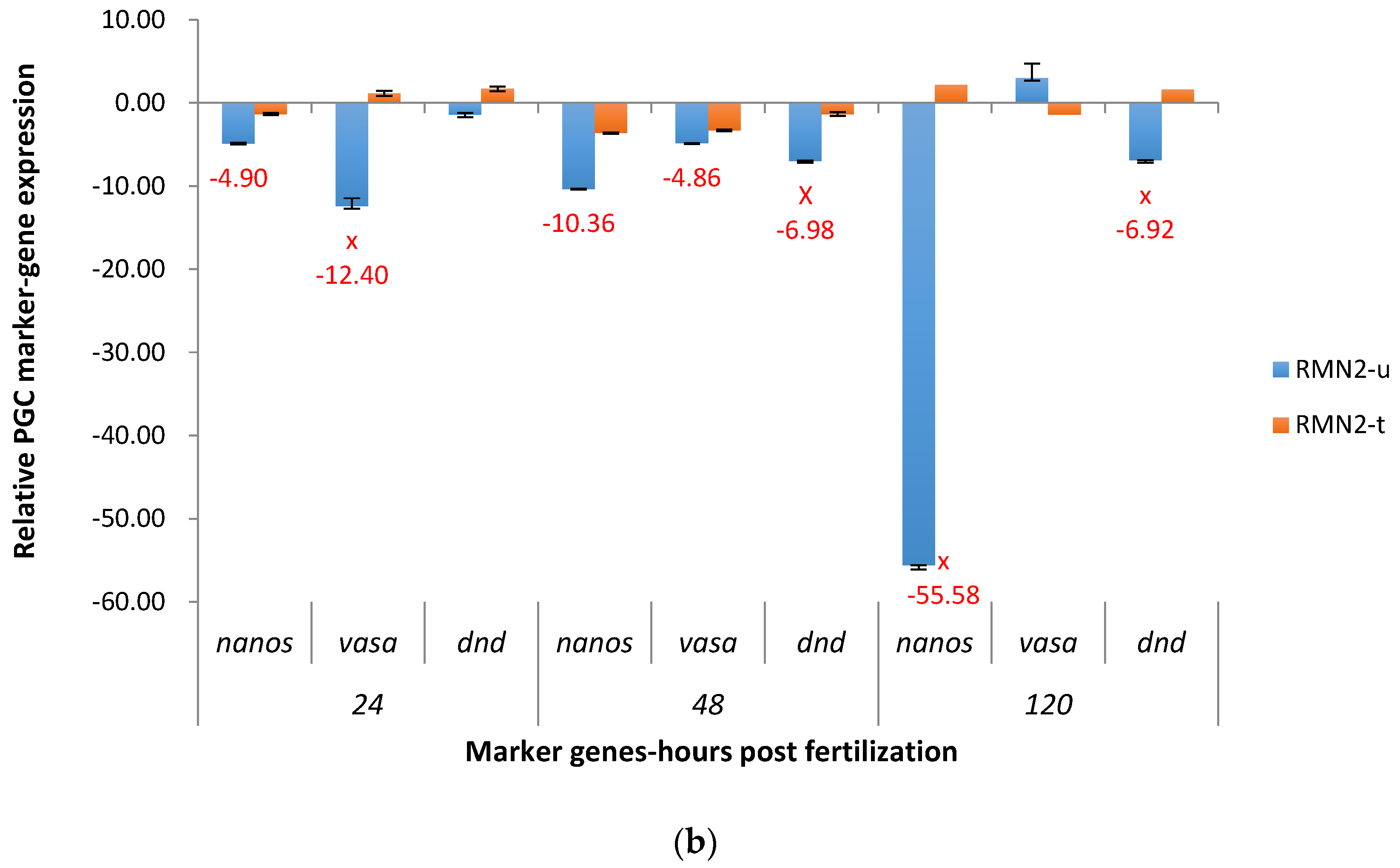
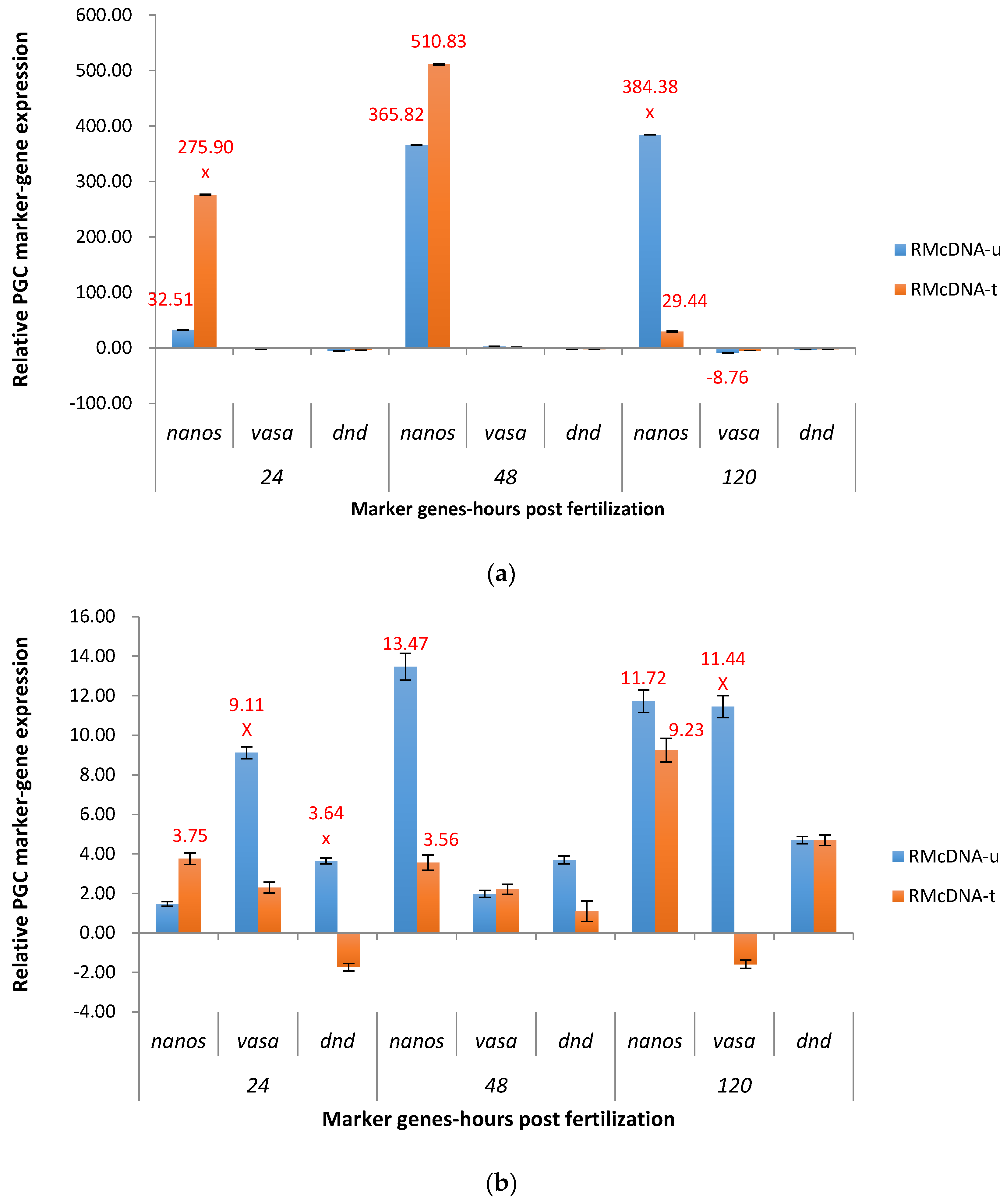
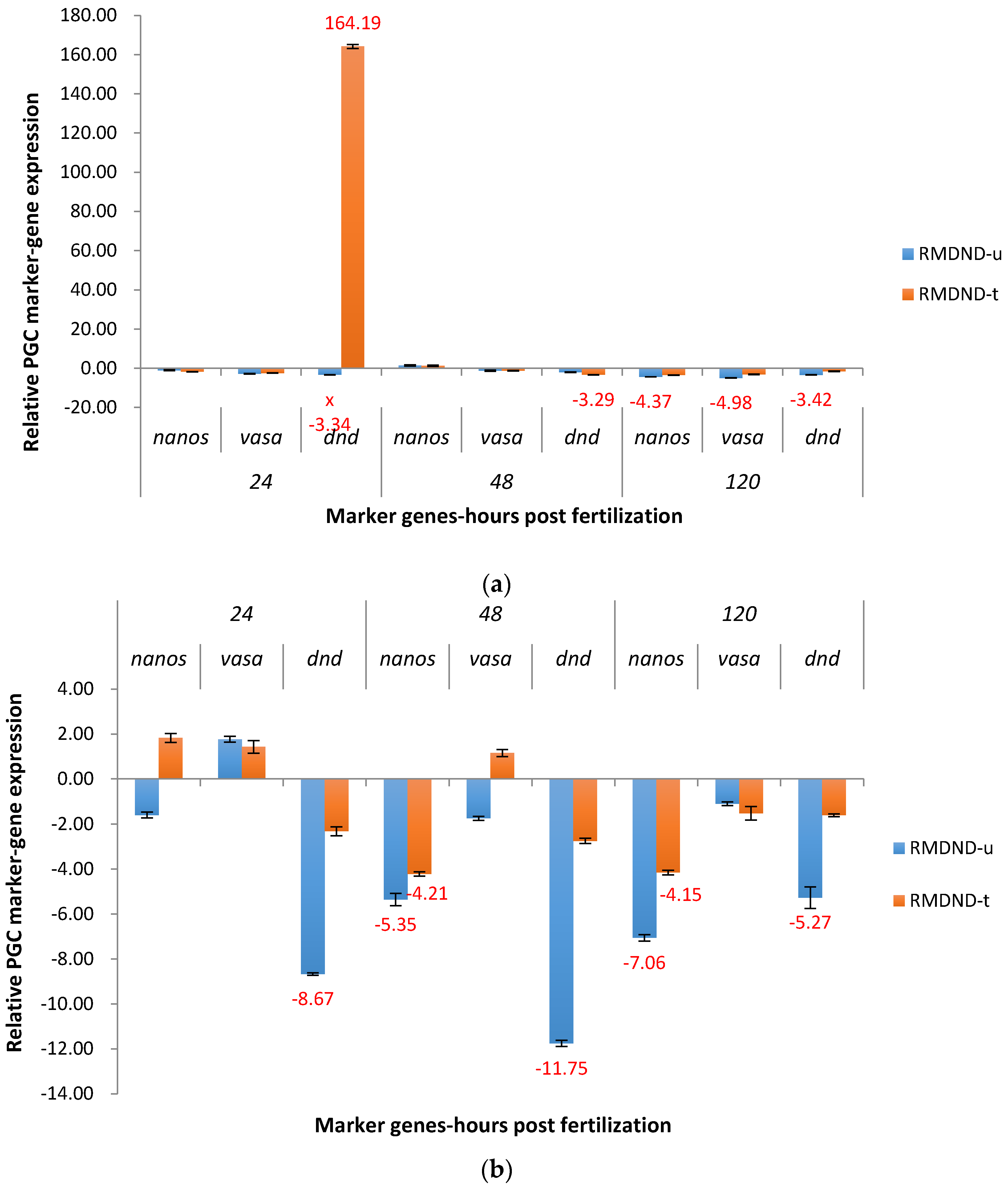
2.3.2. RM System
3. Discussion
4. Materials and Methods
4.1. Construction of Plasmids
4.2. P1 Fish
4.3. Artificial Spawning and Embryo Culture
4.4. Embryo Sample Collection and RNA Extraction
4.5. PCR
4.6. Real-Time PCR
4.7. Spawning Evaluation
4.8. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- AquaBounty Technologies. Available online: https://aquabounty.com/innovation/technology (accessed on 25 January 2017).
- Devlin, R.H.; Sundström, L.F.; Leggatt, R.A. Assessing ecological and evolutionary consequences of growth-accelerated genetically engineered fishes. BioScience 2015, 65, 685–700. [Google Scholar] [CrossRef]
- Rasmussen, R.S.; Morrissey, M.T. Biotechnology in aquaculture: Transgenics and polyploidy. Compr. Rev. Food Sci. Food Saf. 2007, 6, 2–16. [Google Scholar] [CrossRef]
- Mair, G.C.; Nam, Y.K.; Solar, I. Environmental Risk Assessment of Genetically Modified Organisms, Volume 3. In Methodologies for Transgenic Fish; School of Biological Sciences, Flinders University: Adelaide, Australia, 2007. [Google Scholar]
- Molyneaux, K.; Wylie, C. Primordial germ cell migration. Int. J. Dev. Biol. 2004, 48, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Köprunner, M.; Thisse, C.; Thisse, B.; Raz, E. A zebrafish nanos-related gene is essential for the development of primordial germ cells. Genes Dev. 2001, 15, 2877–2885. [Google Scholar] [PubMed]
- Li, M.; Hong, N.; Xu, H.; Yi, M.; Li, C.; Gui, J.; Hong, Y. Medaka vasa is required for migration but not survival of primordial germ cells. Mech. Dev. 2009, 126, 366–381. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.E.; Lehmann, R. Mechanisms guiding primordial germ cell migration: Strategies from different organisms. Nat. Rev. Mol. Cell Biol. 2010, 11, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Fujimoto, T.; Maegawa, S.; Inoue, K.; Tanaka, M.; Arai, K.; Yamaha, E. Visualization of primordial germ cells in vivo using GFP-nos1 3′UTR mRNA. Int. J. Dev. Biol. 2006, 50, 691–699. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.T.; Collodi, P. Inducible sterilization of zebrafish by disruption of primordial germ cell migration. PLoS ONE 2013, 8, e68455. [Google Scholar] [CrossRef] [PubMed]
- Slanchev, K.; Stebler, J.; de la Cueva-Méndez, G.; Raz, E. Development without germ cells: The role of the germ line in zebrafish sex differentiation. Proc. Nat. Acad. Sci. USA 2005, 102, 4074–4079. [Google Scholar] [CrossRef] [PubMed]
- Weidinger, G.; Stebler, J.; Slanchev, K.; Dumstrei, K.; Wise, C.; Lovell-Badge, R.; Thisse, C.; Thisse, C.; Raz, E. Dead end, a novel vertebrate germ plasm component, is required for zebrafish primordial germ cell migration and survival. Curr. Biol. 2003, 13, 1429–1434. [Google Scholar] [CrossRef]
- Fan, L.; Moon, J.; Wong, T.T.; Crodian, J.; Collodi, P. Zebrafish primordial germ cell cultures derived from vasa::RFP transgenic embryos. Stem Cells Dev. 2008, 17, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.; Kawakami, K.; Hopkins, N. Zebrafish vasa homologue RNA is localized to the cleavage planes of 2- and 4-cell-stage embryos and is expressed in the primordial germ cells. Development 1997, 124, 3157–3165. [Google Scholar] [PubMed]
- Herpin, A.; Rohr, S.; Riedel, D.; Kluever, N.; Raz, E.; Schartl, M. Specification of primordial germ cells in medaka (Oryzias latipes). BMC Dev. Biol. 2007, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Braat, A.K.; van de Water, S.; Korving, J.; Zivkovic, D. A zebrafish vasa morphant abolishes vasa protein but does not affect the establishment of the germline. Genesis 2001, 30, 183–185. [Google Scholar] [CrossRef] [PubMed]
- Gavis, E.R.; Lunsford, L.; Bergsten, S.E.; Lehmann, R. A conserved 90 nucleotide element mediates translational repression of nanos RNA. Development 1996, 122, 2791–2800. [Google Scholar] [PubMed]
- Kedde, M.; Strasser, M.J.; Boldajipour, B.; Vrielink, J.A.; Slanchev, K.; le Sage, C.; Nagel, R.; Voorhoeve, P.M.; van Duijse, J.; Orom, U.A.; et al. RNA-binding protein Dnd1 inhibits microRNA access to target mRNA. Cell 2007, 131, 1273–1286. [Google Scholar] [CrossRef] [PubMed]
- Saurabh, S.; Vidyarthi, A.S.; Prasad, D. RNA interference: Concept to reality in crop improvement. Planta 2014, 239, 543–564. [Google Scholar] [CrossRef] [PubMed]
- Daneholt, B. Advanced information: RNA interference. The Nobel Prize in Physiology or Medicine 2006. Archived from the Original on 20 January 2007.
- Thresher, R.; Grewe, P.; Patil, J.G.; Whyard, S.; Templeton, C.M.; Chaimongol, A.; Hardy, C.; Hinds, L.A.; Dunham, R.A. Development of repressible sterility to prevent the establishment of feral populations of exotic and genetically modified animals. Aquaculture 2009, 290, 104–109. [Google Scholar] [CrossRef]
- Bernick, E.P.; Zhang, P.J.; Du, S. Knockdown and overexpression of Unc-45b result in defective myofibril organization in skeletal muscles of zebrafish embryos. BMC Cell Biol. 2010, 11, 70. [Google Scholar] [CrossRef] [PubMed]
- Prelich, G. Gene Overexpression: Uses, mechanisms, and interpretation. Genetics 2012, 190, 841–854. [Google Scholar] [CrossRef] [PubMed]
- Thresher, R.; Hinds, L.A.; Hardy, C.; Whyard, S.; Vignarajan, S.; Grewe, P.; Patil, J. Repressible Sterility of Animals. U.S. Patent 20050071891, 31 March 2005. [Google Scholar]
- Borode, A.O.; Balogun, A.M.; Omoyeni, B.A. Effect of salinity on embryonic development, hatchability, and growth of african catfish, Clarias gariepinus, eggs and larvae. J. Appl. Aquac. 2008, 12, 89–93. [Google Scholar] [CrossRef]
- Swann, L.; Fitzgerald, S. The Use and Application of Salt in Aquaculture; Purdue University: West Lafayette, IN, USA, 1992. [Google Scholar]
- Hoshijima, K.; Hirose, S. Expression of endocrine genes in zebrafish larvae in response to environmental salinity. J. Endocrinol. 2007, 193, 481–491. [Google Scholar] [CrossRef] [PubMed]
- Powers, D.A.; Cole, T.; Creech, K.; Chen, T.T.; Lin, C.M.; Kight, K.; Dunham, R.A. Electroporation: A method for transferring genes into the gametes of zebrafish, Danio rerio, channel catfish, Ictalurus punctatus, and common carp, Cyprinus carpio. Mol. Mar. Biol. Biotechnol. 1992, 1, 301–309. [Google Scholar] [PubMed]
- Cheng, Q.; Su, B.F.; Qin, Z.K.; Weng, C.C.; Yin, F.; Zhou, Y.G.; Fobes, M.; Rex, D.A. Interaction of diet and the masou salmon Δ5-desaturase transgene on Δ6-desaturase and stearoyl-CoA desaturase gene expression and N-3 fatty acid level in common carp (Cyprinus carpio). Transgen. Res. 2014, 23, 729–742. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, P.D.; Schmale, M.C. GFP as a genetic marker scorable throughout the life cycle of transgenic zebra fish. Mar. Biotechnol. 2000, 2, 107–125. [Google Scholar] [PubMed]
- Xu, H.; Lim, M.; Dwarakanath, M.; Hong, Y. Vasa identifies germ cells and critical stages of oogenesis in the asian seabass. Int. J. Biol. Sci. 2004, 10, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Su, B.F.; Shang, M.; Grewe, P.M.; Patil, J.G.; Peatman, E.; Perera, D.A.; Cheng, Q.; Li, C.; Weng, C.C.; Li, P.; et al. Suppression and restoration of primordial germ cell marker gene expression in channel catfish, Ictalurus punctatus, using knockdown constructs regulated by copper transport protein gene promoters: Potential for reversible transgenic sterilization. Theriogenology 2015, 84, 1499–1512. [Google Scholar] [CrossRef] [PubMed]
- Su, B.F.; Peatman, E.; Shang, M.; Thresher, R.; Grewe, P.; Patil, J.; Pinkert, C.A.; Irwin, M.H.; Li, C.; Perera, D.A.; et al. Expression and knockdown of primordial germ cell genes, vasa, nanos and dead end in common carp (Cyprinus carpio) embryos for transgenic sterilization and reduced sexual maturity. Aquaculture 2014, 420–421, 72–84. [Google Scholar] [CrossRef]
- Zhang, D. Transgenic Disruption of Aromatase Using the Daughterless Construct to Alter Sex Ratio in Common Carp, Cyprinus carpio. Master’s Thesis, Auburn University, Auburn, AL, USA, 1 August 2016. [Google Scholar]
- Saxena, S.; Jonsson, Z.O.; Dutta, A. Small RNAs with imperfect match to endogenous mRNA repress translation. J. Biol. Chem. 2003, 278, 44312–44319. [Google Scholar] [CrossRef] [PubMed]
- Scacheri, P.C.; Rozenblatt-Rosen, O.; Caplen, N.J.; Wolfsberg, T.G.; Umayam, L.; Lee, J.C.; Hughes, C.M.; Shanmugam, K.S.; Bhattacharjee, A.; Meyerson, M.; et al. Short interfering RNAs can induce unexpected and divergent changes in the levels of untargeted proteins in mammalian cells. Proc. Nat. Acad. Sci. USA 2004, 101, 1892–1897. [Google Scholar] [CrossRef] [PubMed]
- Slanchev, K.; Stebler, J.; Goudarzi, M.; Cojocaru, V.; Weidinger, G.; Raz, E. Control of Dead end localization and activity—Implications for the function of the protein in antagonizing miRNA function. Mech. Dev. 2009, 126, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Raz, E. The function and regulation of vasa-like genes in germ-cell development. Gen. Biol. 2000, 1, 1017.1–1017.6. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, N.; Collier, B.; Maratou, K.; Bingham, V.; Speed, R.M.; Taggart, M.; Semple, C.A.; Gray, N.K.; Cooke, H.J. Dazl binds in vivo to specific transcripts and can regulate the pre-meiotic translation of Mvh in germ cells. Hum. Mol. Genet. 2005, 14, 3899–3909. [Google Scholar] [CrossRef] [PubMed]
- Kato, Y.; Katsuki, T.; Kokubo, H.; Masuda, A.; Saga, Y. Dazl is a target RNA suppressed by mammalian NANOS2 in sexually differentiating male germ cells. Nat. Commun. 2016, 7, 11272. [Google Scholar] [CrossRef] [PubMed]
- PIPs: Human protein-protein interactions prediction database. Available online: http://www.compbio.dundee.ac.uk/www-pips/evidence.jsp?Prot=IPI00395811&Prot1=21244&Prot2=10958&Interactions=Prot&LR=0.025&modules=on (accessed on 2 May 2017).
- Perry, W.G. Note on the spawning of blue and channel catfish in brackish water ponds. Prog. Fish Cultur. 1973, 35, 164–166. [Google Scholar] [CrossRef]
- Perera, D.A. Studies for Improvement of Reproductive Biotechnology for Production of Channel Catfish (Ictalurus punctatus) Female X Blue Catfish (Ictalurus furcatus) Male Hybrid Embryos. Ph.D. Thesis, Auburn University, Auburn, AL, USA, 2012. [Google Scholar]
- Su, B.F.; Perera, D.A.; Zohar, Y.; Abraham, E.; Stubblefield, J.; Fobes, M.; Beam, R.; Argue, B.; Ligeon, C.; Padi, J.; et al. Relative effectiveness of carp pituitary extract, luteininzing hormone releasing hormone analog (LHRHa) injections and LHRHa implants for producing hybrid catfish fry. Aquaculture 2013, 372–375, 133–136. [Google Scholar] [CrossRef]
- Chatakondi, N.G.; Yant, D.R.; Dunham, R.A. Final Report-Spawning and Hatchery Technology to Improve Hybrid Catfish Fry Production; SBIR project No. 2002-00384; U.S. Department of Agriculture (USDA): Washington, DC, USA, 2005.
- Armstrong, J.B.; Duhon, S.T.; Malacinski, G.M. Raising the Axolotl in Captivity. In Developmental Biology of the Axolotl; Armstrong, J.B., Malacinski, G.M., Eds.; Oxford University Press: New York, NY, USA, 1989; pp. 220–227. [Google Scholar]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constructs | Untreated | Treated | ||||
|---|---|---|---|---|---|---|
| Spawned | Total | % | Spawned | Total | % | |
| ADSSN1 | 1 | 4 | 25 | - | - | - |
| ADSSN2 * | 2 | 8 | 25 | 4 | 4 | 100 |
| ADSScDNA | 8 | 13 | 62 | 8 | 8 | 100 |
| ADSSDND | 6 | 8 | 75 | 5 | 5 | 100 |
| RMN1 | 7 | 9 | 78 | 8 | 8 | 100 |
| RMN2 | 6 | 8 | 75 | 8 | 10 | 80 |
| RMcDNA | 2 | 3 | 67 | 3 | 3 | 100 |
| RMDND | 2 | 5 | 40 | 2 | 3 | 67 |
| Total ** | 34 | 58 | 59 | 38 | 41 | 93 |
| Constructs | Untreated | Treated | Ratio of T/U | ||||
|---|---|---|---|---|---|---|---|
| Spawned | Total | % | Spawned | Total | % | ||
| ADSS * | 17 | 33 | 52 | 17 | 17 | 100 | 1.94 |
| Rm | 17 | 25 | 68 | 21 | 24 | 88 | 1.29 |
| N1 | 8 | 13 | 62 | 8 | 8 | 100 | 1.61 |
| N2 * | 8 | 16 | 50 | 12 | 14 | 86 | 1.72 |
| cDNA | 10 | 16 | 63 | 11 | 11 | 100 | 1.59 |
| DND | 8 | 13 | 62 | 7 | 8 | 82 | 1.32 |
| Regulator Gene | Nanos | Vasa | Dead End | |
|---|---|---|---|---|
| Responder Gene | ||||
| Nanos | Vasa is required for promoting translation of nanos | Dead end protects nanos from being degraded by miR-430 in germ cells | ||
| Vasa | Nanos suppresses dazl, dazl regulates vasa expression | Dead end potentially regulates dazl, dazl regulates vasa expression | ||
| Dead end | Nanos potentially regulates dead end, mechanism unknown | No known regulation found | ||
| Primers | Sequence (5′–3′) |
|---|---|
| Forward primer for insulator | ACACTGTCCTTGGTATCAGCA |
| Reverse primer for insulator | ACACCTCCTTGATCCTGTGC |
| Forward primer for ocean pout terminator | TGCCCAAATTTCCTGCCTGA |
| Reverse primer for ocean pout terminator | TGCCTGTGAGGGTGACAAAA |
| Forward primer for adss2 promoter | CCGCCAAAATACGACTG |
| Reverse primer for adss2 promoter | TAAGCGAGGGAACAACG |
| Forward primer for racemase promoter | TACAACACCCACAATCCC |
| Reverse primer for racemase promoter | CACCGTAACAAAGAAACTCC |
| Forward primer for N1 | ATGACTCGCGATGGTGTTTT |
| Reverse primer for N1 | CATACTGGGGTGTTTGCTGA |
| Forward primer for N2 | CACTCGCTAGAGTGGGGTTC |
| Reverse primer for N2 | TCATACTGGGGTGTTTGCTG |
| Forward primer for cDNA | GAACCCCACTCTCACACTCG |
| Reverse primer for cDNA | AGACCCGCACCTGTAACATG |
| Forward primer for DND | TAGCGCCGCTCTACGAGT |
| Reverse primer for DND | TCATACTGGGGTGTTTGCTG |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Su, B.; Qin, G.; Ye, Z.; Alsaqufi, A.; Perera, D.A.; Shang, M.; Odin, R.; Vo, K.; Drescher, D.; et al. Salt Sensitive Tet-Off-Like Systems to Knockdown Primordial Germ Cell Genes for Repressible Transgenic Sterilization in Channel Catfish, Ictalurus punctatus. Mar. Drugs 2017, 15, 155. https://doi.org/10.3390/md15060155
Li H, Su B, Qin G, Ye Z, Alsaqufi A, Perera DA, Shang M, Odin R, Vo K, Drescher D, et al. Salt Sensitive Tet-Off-Like Systems to Knockdown Primordial Germ Cell Genes for Repressible Transgenic Sterilization in Channel Catfish, Ictalurus punctatus. Marine Drugs. 2017; 15(6):155. https://doi.org/10.3390/md15060155
Chicago/Turabian StyleLi, Hanbo, Baofeng Su, Guyu Qin, Zhi Ye, Ahmed Alsaqufi, Dayan A. Perera, Mei Shang, Ramjie Odin, Khoi Vo, David Drescher, and et al. 2017. "Salt Sensitive Tet-Off-Like Systems to Knockdown Primordial Germ Cell Genes for Repressible Transgenic Sterilization in Channel Catfish, Ictalurus punctatus" Marine Drugs 15, no. 6: 155. https://doi.org/10.3390/md15060155
APA StyleLi, H., Su, B., Qin, G., Ye, Z., Alsaqufi, A., Perera, D. A., Shang, M., Odin, R., Vo, K., Drescher, D., Robinson, D., Zhang, D., Abass, N., & Dunham, R. A. (2017). Salt Sensitive Tet-Off-Like Systems to Knockdown Primordial Germ Cell Genes for Repressible Transgenic Sterilization in Channel Catfish, Ictalurus punctatus. Marine Drugs, 15(6), 155. https://doi.org/10.3390/md15060155