
Enzymatic Processes in Marine Biotechnology


Abstract
:
1. Introduction
2. Literature Search
3. Biorefinery
4. Food Applications
5. Fine Chemistry and Lab Techniques
6. Sediments and Bioremediation
7. Others
8. Conclusions
Acknowledgments
Conflicts of Interest
References
- Census of Marine Life. Available online: http://www.coml.org (accessed on 23 March 2017).
- Trincone, A. Increasing knowledge: The grand challenge in marine biotechnology. Front. Mar. Sci. 2014, 1. [Google Scholar] [CrossRef]
- Trincone, A. Potential biocatalysts originating from sea environments. J. Mol. Catal. B Enzym. 2010, 66, 241–256. [Google Scholar] [CrossRef]
- Trincone, A. Marine Biocatalysts: Enzymatic features and applications. Mar. Drugs 2011, 9, 478–499. [Google Scholar] [CrossRef] [PubMed]
- Lima, R.N.; Porto, A.L.M. Recent advances in marine enzymes for biotechnological processes. In Marine Enzymes Biotechnology: Production and Industrial Applications, Part I—Production of Enzymes; Kim, S.-K., Toldra, F., Eds.; Academic Press Elsevier: New York, NY, USA, 2016; pp. 153–192. [Google Scholar]
- Cherubini, F. The biorefinery concept: Using biomass instead of oil for producing energy and chemicals. Energy Convers. Manag. 2010, 51, 1412–1421. [Google Scholar] [CrossRef]
- Bhalla, A.; Bansal, N.; Kumar, S.; Bischoff, K.; Sani, R. Improved lignocellulose conversion to biofuels with thermophilic bacteria and thermostable enzymes. Bioresour. Technol. 2013, 128, 751–759. [Google Scholar] [CrossRef] [PubMed]
- Menon, V.; Rao, M. Trends in bioconversion of lignocellulose: Biofuels, platform chemicals & biorefinery concept. Prog. Energy Combust. Sci. 2012, 38, 522–550. [Google Scholar]
- Trivedi, N.; Reddy, C.; Radulovich, R.; Jha, B. Solid state fermentation (SSF)-derived cellulase for saccharification of the green seaweed Ulva for bioethanol production. Algal Res. 2015, 9, 48–54. [Google Scholar] [CrossRef]
- Satheeja Santhi, V.; Bhagat, A.; Saranya, S.; Govindarajan, G.; Jebakumar, S. Seaweed (Eucheuma cottonii) associated microorganisms, a versatile enzyme source for the lignocellulosic biomass processing. Int. Biodeterior. Biodegrad. 2014, 96, 144–151. [Google Scholar] [CrossRef]
- Annamalai, N.; Rajeswari, M.; Balasubramanian, T. Enzymatic saccharification of pretreated rice straw by cellulase produced from Bacillus carboniphilus CAS 3 utilizing lignocellulosic wastes through statistical optimization. Biomass Bioenergy 2014, 68, 151–160. [Google Scholar] [CrossRef]
- Harshvardhan, K.; Mishra, A.; Jha, B. Purification and characterization of cellulase from a marine Bacillus sp. H1666: A potential agent for single step saccharification of seaweed biomass. J. Mol. Catal. B Enzym. 2013, 93, 51–56. [Google Scholar] [CrossRef]
- Kim, B.; Lee, B.; Lee, Y.; Jin, I.; Chung, C.; Lee, J. Purification and characterization of carboxymethylcellulase isolated from a marine bacterium, Bacillus subtilis subsp. subtilis A-53. Enzyme Microb. Technol. 2009, 44, 411–416. [Google Scholar] [CrossRef]
- Kang, D.; Hyeon, J.; You, S.; Kim, S.; Han, S. Efficient enzymatic degradation process for hydrolysis activity of the carrageenan from red algae in marine biomass. J. Biotechnol. 2014, 192, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.B.; Park, J.; Huh, I.Y.; Hong, S.-K.; Chang, Y.K. Agarose hydrolysis by two-stage enzymatic process and bioethanol production from the hydrolysate. Process Biochem. 2016, 51, 759–764. [Google Scholar] [CrossRef]
- Kim, H.; Lee, S.; Kim, K.; Choi, I. The complete enzymatic saccharification of agarose and its application to simultaneous saccharification and fermentation of agarose for ethanol production. Bioresour. Technol. 2012, 107, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Byreddy, A.; Barrow, C.; Puri, M. Bead milling for lipid recovery from thraustochytrid cells and selective hydrolysis of Schizochytrium DT3 oil using lipase. Bioresour. Technol. 2016, 200, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Y.; Bai, M.-D.; Chang, J.-S. Improving microalgal oil collecting efficiency by pretreating the microalgal cell wall with destructive bacteria. Biochem. Eng. J. 2013, 81, 170–176. [Google Scholar] [CrossRef]
- Chen, J.; Liu, X.; Wei, D.; Chen, G. High yields of fatty acid and neutral lipid production from cassava bagasse hydrolysate (CBH) by heterotrophic Chlorella protothecoides. Bioresour. Technol. 2015, 191, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, J.; Gerken, H.; Zhang, C.; Hu, Q.; Li, Y. Highly-efficient enzymatic conversion of crude algal oils into biodiesel. Bioresour. Technol. 2014, 172, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.; Caprioni, R.; Nogueira, M.; Menduiña, A.; Ramos, P.; Pérez-Martín, R. Valorisation of effluents obtained from chemical and enzymatic chitin production of Illex argentinus pen by-products as nutrient supplements for various bacterial fermentations. Biochem. Eng. J. 2015, 116, 34–44. [Google Scholar] [CrossRef]
- Prieto, M.; Prieto, I.; Vázquez, J.; Ferreira, I. An environmental management industrial solution for the treatment and reuse of mussel wastewaters. Sci. Total Environ. 2015, 538, 117–128. [Google Scholar] [CrossRef] [PubMed]
- Mathimani, T.; Uma, L.; Prabaharan, D. Homogeneous acid catalysed transesterification of marine microalga Chlorella sp. BDUG 91771 lipid–an efficient biodiesel yield and its characterization. Renew. Energy 2015, 81, 523–533. [Google Scholar] [CrossRef]
- Li, Y.; Cui, J.; Zhang, G.; Liu, Z.; Guan, H.; Hwang, H.; Aker, W.; Wang, P. Optimization study on the hydrogen peroxide pretreatment and production of bioethanol from seaweed Ulva prolifera biomass. Bioresour. Technol. 2016, 214, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Healy, M.; Romo, C.; Bustos, R. Bioconversion of marine crustacean shell waste. Resour. Conserv. Recycl. 1994, 11, 139–147. [Google Scholar] [CrossRef]
- Kim, H.M.; Oh, C.H.; Bae, H.-J. Comparison of red microalagae (Porphyridium cruentum) culture conditions for bioethanol production. Biores. Technol. 2017, 233, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Ravanal, M.; Pezoa-Conte, R.; von Schoultz, S.; Hemming, J.; Salazar, O.; Anugwom, I.; Jogunola, O.; Mäki-Arvela, P.; Willför, S.; Mikkola, J.; et al. Comparison of different types of pretreatment and enzymatic saccharification of Macrocystis pyrifera for the production of biofuel. Algal Res. 2016, 13, 141–147. [Google Scholar] [CrossRef]
- Kwon, O.; Kim, D.; Kim, S.; Jeong, G. Production of sugars from macro-algae Gracilaria verrucosa using combined process of citric acid-catalyzed pretreatment and enzymatic hydrolysis. Algal Res. 2016, 13, 293–297. [Google Scholar] [CrossRef]
- Tan, I.; Lee, K. Comparison of different process strategies for bioethanol production from Eucheuma cottonii: An economic study. Bioresour. Technol. 2016, 199, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Hansen, J.; Bjerre, A. Integrated bioethanol and protein production from brown seaweed Laminaria digitata. Bioresour. Technol. 2015, 197, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Bohutskyi, P.; Chow, S.; Ketter, B.; Betenbaugh, M.; Bouwer, E. Prospects for methane production and nutrient recycling from lipid extracted residues and whole Nannochloropsis salina using anaerobic digestion. Appl. Energy 2015, 154, 718–731. [Google Scholar] [CrossRef]
- Mirsiaghi, M.; Reardon, K. Conversion of lipid-extracted Nannochloropsis salina biomass into fermentable sugars. Algal Res. 2015, 8, 145–152. [Google Scholar] [CrossRef]
- Abd-Rahim, F.; Wasoh, H.; Zakaria, M.; Ariff, A.; Kapri, R.; Ramli, N.; Siew-Ling, L. Production of high yield sugars from Kappaphycus alvarezii using combined methods of chemical and enzymatic hydrolysis. Food Hydrocoll. 2014, 42, 309–315. [Google Scholar] [CrossRef]
- Kim, D.; Lee, S.; Jeong, G. Production of reducing sugar from Enteromorpha intestinalis by hydrothermal and enzymatic hydrolysis. Bioresour. Technol. 2014, 161, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Wu, J.; Liao, Y.; Wang, M.; Shih, I. Sequential acid and enzymatic hydrolysis in situ and bioethanol production from Gracilaria biomass. Bioresour. Technol. 2014, 156, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Choi, I.; Kim, H.; Wi, S.; Bae, H. Bioethanol production from the nutrient stress-induced microalga Chlorella vulgaris by enzymatic hydrolysis and immobilized yeast fermentation. Bioresour. Technol. 2014, 153, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Gupta, R.; Kumar, G.; Sahoo, D.; Kuhad, R. Bioethanol production from Gracilaria verrucosa, a red alga, in a biorefinery approach. Bioresour. Technol. 2013, 135, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, P.; Barcelos, C.; da Costa, A.; Pereira, N. Production of ethanol 3G from Kappaphycus alvarezii: Evaluation of different process strategies. Bioresour. Technol. 2013, 134, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Pilavtepe, M.; Sargin, S.; Celiktas, M.; Yesil-Celiktas, O. An integrated process for conversion of Zostera marina residues to bioethanol. J. Supercrit. Fluids 2012, 68, 117–122. [Google Scholar] [CrossRef]
- Ge, L.; Wang, P.; Mou, H. Study on saccharification techniques of seaweed wastes for the transformation of ethanol. Renew. Energy 2011, 36, 84–89. [Google Scholar] [CrossRef]
- Kim, S.; Hong, C.; Jeon, S.; Shin, H. High-yield production of biosugars from Gracilaria verrucosa by acid and enzymatic hydrolysis processes. Bioresour. Technol. 2015, 196, 634–641. [Google Scholar] [CrossRef] [PubMed]
- Le Guillard, C.; Dumay, J.; Donnay-Moreno, C.; Bruzac, S.; Ragon, J.; Fleurence, J.; Bergé, J. Ultrasound-assisted extraction of R-phycoerythrin from Grateloupia turuturu with and without enzyme addition. Algal Res. 2015, 12, 522–528. [Google Scholar] [CrossRef]
- Ladisch, M.; Lin, K.; Voloch, M.; Tsao, G. Process considerations in the enzymatic hydrolysis of biomass. Enzyme Microb. Technol. 1983, 5, 82–102. [Google Scholar] [CrossRef]
- Carrillo-Reyes, J.; Barragán-Trinidad, M.; Buitrón, G. Biological pretreatments of microalgal biomass for gaseous biofuel production and the potential use of rumen microorganisms: A review. Algal Res. 2016, 18, 341–351. [Google Scholar] [CrossRef]
- Eijsink, V.; Vaaje-Kolstad, G.; Vårum, K.; Horn, S. Towards new enzymes for biofuels: Lessons from chitinase research. Trends Biotechnol. 2008, 26, 228–235. [Google Scholar] [CrossRef] [PubMed]
- Guldhe, A.; Singh, B.; Mutanda, T.; Permaul, K.; Bux, F. Advances in synthesis of biodiesel via enzyme catalysis: Novel and sustainable approaches. Renew. Sustain. Energy Rev. 2015, 41, 1447–1464. [Google Scholar] [CrossRef]
- Wang, L.; Chi, Z.; Wang, X.; Liu, Z.; Li, J. Diversity of lipase-producing yeasts from marine environments and oil hydrolysis by their crude enzymes. Ann. Microbiol. 2007, 57, 495–501. [Google Scholar] [CrossRef]
- Ledesma-Amaro, R.; Nicaud, J. Metabolic engineering for expanding the substrate range of Yarrowia lipolytica. Trends Biotechnol. 2016, 34, 798–809. [Google Scholar] [CrossRef] [PubMed]
- Suganya, T.; Varman, M.; Masjuki, H.H.; Renganathan, S. Macroalgae and microalgae as a potential source for commercial applications along with biofuels production: A biorefinery approach. Renew. Sust. Energy Rev. 2016, 55, 909–941. [Google Scholar] [CrossRef]
- Demuez, M.; González-Fernández, C.; Ballesteros, M. Algicidal microorganisms and secreted algicides: New tools to induce microalgal cell disruption. Biotechnol. Adv. 2015, 33, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.; Singhania, R.; Pandey, A. Novel enzymatic processes applied to the food industry. Curr. Opin. Food Sci. 2016, 7, 64–72. [Google Scholar] [CrossRef]
- Shahidi, F.; Janak Kamil, Y. Enzymes from fish and aquatic invertebrates and their application in the food industry. Trends Food Sci. Technol. 2001, 12, 435–464. [Google Scholar] [CrossRef]
- Carroad, P.; Wilke, C. Enzymes and microorganisms in food industry waste processing and conversion to useful products: A review of the literature. Resour. Recover. Conserv. 1978, 3, 165–178. [Google Scholar] [CrossRef]
- Cudennec, B.; Caradec, T.; Catiau, L.; Ravallec, R. Upgrading of sea by-products: Potential nutraceutical applications. In Marine Medicinal Foods Implications and Applications—Animals and Microbes; Kim, S.-K., Ed.; Academic Press: New York, NY, USA, 2012; Volume 65, pp. 479–494. [Google Scholar]
- Grienke, U.; Silke, J.; Tasdemir, D. Bioactive compounds from marine mussels and their effects on human health. Food Chem. 2014, 142, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Undeland, I. Oxidative stability of seafood. In Oxidative Stability and Shelf Life of Foods Containing Oils and Fats; Hu, M., Jacobsen, C., Eds.; AOCS Press: Champaign, IL, USA, 2016; pp. 391–460. [Google Scholar]
- Adachi, K.; Hirata, T.; Nagai, K.; Sakaguchi, M. Hemocyanin a most likely inducer of black spots in Kuruma prawn Penaeus japonicus during storage. J. Food Sci. 2008, 66, 1130–1136. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Dinesh kumar, B.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef] [PubMed]
- Guerard, F. Enzymatic methods for marine by-products recovery. In Maximising the Value of Marine By-Products Woodhead Publishing Series in Food Science, Technology and Nutrition; Shahidi, F., Ed.; Woodhead Publishing: Cambridge, UK, 2007; pp. 107–143. [Google Scholar]
- Sila, A.; Bougatef, A. Antioxidant peptides from marine by-products: Isolation, identification and application in food systems. A review. J. Funct. Foods 2016, 21, 10–26. [Google Scholar] [CrossRef]
- Fleurence, J. Seaweeds as food. In Seaweed in Health and Disease Prevention; Fleurence, J., Levine, I., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 149–167. [Google Scholar]
- Bhaskar, N.; Sudepa, E.; Rashimi, H.; Tamilselvi, A. Partial purification and characterization of protease of Bacillus proteolyticus CFR3001 isolated from fish processing waste and its antibacterial activities. Bioresour. Technol. 2007, 98, 2758–2764. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, D.A.S.B.; Minozzo, M.G.; Licodiedoff, S.; Waszczynskyj, N. Physicochemical and sensory characterization of refined and deodorized tuna (Thunnus albacares) by-product oil obtained by enzymatic hydrolysis. Food Chem. 2016, 207, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zhang, B.; Chen, B.; Jing, L.; Zhu, Z.; Kazemi, K. Modeling and optimization of Newfoundland shrimp waste hydrolysis for microbial growth using response surface methodology and artificial neural networks. Mar. Pollut. Bull. 2016, 109, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Sayari, N.; Sila, A.; Abdelmalek, B.E.; Abdallah, R.B.; Ellouz-Chaabouni, S.; Bougatef, A.; Balti, R. Chitin and chitosan from the Norway lobster by-products: Antimicrobial and anti-proliferative activities. Int. J. Biol. Macromol. 2016, 87, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Charoensiddhi, S.; Conlon, M.A.; Vuaran, M.S.; Franco, C.M.M.; Zhang, W. Impact of extraction processes on prebiotic potential of the brown seaweed Ecklonia radiata by in vitro human gut bacteria fermentation. J. Funct. Foods 2016, 24, 221–230. [Google Scholar] [CrossRef]
- Tonon, R.V.; dos Santos, B.A.; Couto, C.C.; Mellinger-Silva, C.; Brígida, A.I.S.; Cabral, L.M.C. Coupling of ultrafiltration and enzymatic hydrolysis aiming at valorizing shrimp wastewater. Food Chem. 2016, 198, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, J.A.; Blanco, M.; Fraguas, J.; Pastrana, L.; Pérez-Martín, R. Optimisation of the extraction and purification of chondroitin sulphate from head by-products of Prionace glauca by environmental friendly processes. Food Chem. 2016, 198, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Grant, B.; Picchi, N.; Davie, A.; Leclercq, E.; Migaud, H. Removal of the adhesive gum layer surrounding naturally fertilised ballan wrasse (Labrus bergylta) eggs. Aquaculture 2016, 456, 44–49. [Google Scholar] [CrossRef]
- Solaesa, Á.G.; Sanz, M.T.; Falkeborg, M.; Beltrán, S.; Guo, Z. Production and concentration of monoacylglycerols rich in omega-3 polyunsaturated fatty acids by enzymatic glycerolysis and molecular distillation. Food Chem. 2016, 190, 960–967. [Google Scholar] [CrossRef] [PubMed]
- Lassoued, I.; Mora, L.; Nasri, R.; Aydi, M.; Toldrá, F.; Aristoy, M.-C.; Barkia, A.; Nasri, M. Characterization, antioxidative and ACE inhibitory properties of hydrolysates obtained from thornback ray (Raja clavata) muscle. J. Proteom. 2015, 128, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Gringer, N.; Hosseini, S.V.; Svendsen, T.; Undeland, I.; Christensen, M.L.; Baron, C.P. Recovery of biomolecules from marinated herring (Clupea harengus) brine using ultrafiltration through ceramic membranes. LWT Food Sci. Technol. 2015, 63, 423–429. [Google Scholar] [CrossRef]
- Opheim, M.; Šližytė, R.; Sterten, H.; Provan, F.; Larssen, E.; Kjos, N.P. Hydrolysis of Atlantic salmon (Salmo salar) rest raw materials—Effect of raw material and processing on composition, nutritional value, and potential bioactive peptides in the hydrolysates. Process Biochem. 2015, 50, 1247–1257. [Google Scholar] [CrossRef]
- Younes, I.; Nasri, R.; Bkhairia, I.; Jellouli, K.; Nasri, M. New proteases extracted from red scorpionfish (Scorpaena scrofa) viscera: Characterization and application as a detergent additive and for shrimp waste deproteinization. Food Bioprod. Process. 2015, 94, 453–462. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Hemalatha, R.; Jyothirmayi, T.; Diwan, P.V.; Bhaskarachary, K.; Vajreswari, A.; Ramesh Kumar, R.; Dinesh Kumar, B. Chemical composition and immunomodulatory effects of enzymatic protein hydrolysates from common carp (Cyprinus carpio) egg. Nutrition 2015, 31, 388–398. [Google Scholar] [CrossRef] [PubMed]
- Laohakunjit, N.; Selamassakul, O.; Kerdchoechuen, O. Seafood-like flavour obtained from the enzymatic hydrolysis of the protein by-products of seaweed (Gracilaria sp.). Food Chem. 2014, 158, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Šližytė, R.; Carvajal, A.K.; Mozuraityte, R.; Aursand, M.; Storrø, I. Nutritionally rich marine proteins from fresh herring by-products for human consumption. Process Biochem. 2014, 49, 1205–1215. [Google Scholar] [CrossRef]
- Halldorsdottir, S.M.; Sveinsdottir, H.; Freysdottir, J.; Kristinsson, H.G. Oxidative processes during enzymatic hydrolysis of cod protein and their influence on antioxidant and immunomodulating ability. Food Chem. 2014, 142, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Beaulieu, L.; Thibodeau, J.; Bonnet, C.; Bryl, P.; Carbonneau, M.-É. Detection of antibacterial activity in an enzymatic hydrolysate fraction obtained from processing of Atlantic rock crab (Cancer irroratus) by-products. Pharmanutrition 2013, 1, 149–157. [Google Scholar] [CrossRef]
- Amado, I.R.; Vázquez, J.A.; González, M.P.; Murado, M.A. Production of antihypertensive and antioxidant activities by enzymatic hydrolysis of protein concentrates recovered by ultrafiltration from cuttlefish processing wastewaters. Biochem. Eng. J. 2013, 76, 43–54. [Google Scholar] [CrossRef]
- Saidi, S.; Deratani, A.; Ben Amar, R.; Belleville, M.-P. Fractionation of a tuna dark muscle hydrolysate by a two-step membrane process. Sep. Purif. Technol. 2013, 108, 28–36. [Google Scholar] [CrossRef]
- Castro-Ceseña, A.B.; del Pilar Sánchez-Saavedra, M.; Márquez-Rocha, F.J. Characterisation and partial purification of proteolytic enzymes from sardine by-products to obtain concentrated hydrolysates. Food Chem. 2012, 135, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Cian, R.E.; Martínez-Augustin, O.; Drago, S.R. Bioactive properties of peptides obtained by enzymatic hydrolysis from protein byproducts of Porphyra columbina. Food Res. Int. 2012, 49, 364–372. [Google Scholar] [CrossRef]
- Rubio-Rodríguez, N.; de Diego, S.M.; Beltrán, S.; Jaime, I.; Sanz, M.T.; Rovira, J. Supercritical fluid extraction of fish oil from fish by-products: A comparison with other extraction methods. J. Food Eng. 2012, 109, 238–248. [Google Scholar] [CrossRef]
- Beaulieu, L.; Thibodeau, J.; Bryl, P.; Carbonneau, M.-É. Characterization of enzymatic hydrolyzed snow crab (Chionoecetes opilio) by-product fractions: A source of high-valued biomolecules. Bioresour. Technol. 2009, 100, 3332–3342. [Google Scholar] [CrossRef] [PubMed]
- Huo, J.; Zhao, Z. Study on enzymatic hydrolysis of Gadus morrhua skin collagen and molecular weight distribution of hydrolysates. Agric. Sci. China 2009, 8, 723–729. [Google Scholar] [CrossRef]
- Vázquez, J.A.; Murado, M.A. Enzymatic hydrolysates from food wastewater as a source of peptones for lactic acid bacteria productions. Enzyme Microb. Technol. 2008, 43, 66–72. [Google Scholar] [CrossRef]
- Barcia, I.; Sánchez-Purriños, M.L.; Novo, M.; Novás, A.; Maroto, J.F.; Barcia, R. Optimisation of Dosidicus gigas mantle proteolysis at industrial scale. Food Chem. 2008, 107, 869–875. [Google Scholar] [CrossRef]
- Guerard, F.; Sumaya-Martinez, M.T.; Laroque, D.; Chabeaud, A.; Dufossé, L. Optimization of free radical scavenging activity by response surface methodology in the hydrolysis of shrimp processing discards. Process Biochem. 2007, 42, 1486–1491. [Google Scholar] [CrossRef]
- Šližytė, R.; Rustad, T.; Storrø, I. Enzymatic hydrolysis of cod (Gadus morhua) by-products. Process Biochem. 2005, 40, 3680–3692. [Google Scholar] [CrossRef]
- Daukšas, E.; Falch, E.; Šližytė, R.; Rustad, T. Composition of fatty acids and lipid classes in bulk products generated during enzymic hydrolysis of cod (Gadus morhua) by-products. Process Biochem. 2005, 40, 2659–2670. [Google Scholar] [CrossRef]
- Lignot, B.; Lahogue, V.; Bourseau, P. Enzymatic extraction of chondroitin sulfate from skate cartilage and concentration-desalting by ultrafiltration. J. Biotechnol. 2003, 103, 281–284. [Google Scholar] [CrossRef]
- Guerard, F.; Guimas, L.; Binet, A. Production of tuna waste hydrolysates by a commercial neutral protease preparation. J. Mol. Catal. B Enzym. 2002, 19–20, 489–498. [Google Scholar] [CrossRef]
- Guérard, F.; Dufossé, L.; De La Broise, D.; Binet, A. Enzymatic hydrolysis of proteins from yellowfin tuna (Thunnus albacares) wastes using alcalase. J. Mol. Catal. B Enzym. 2001, 11, 1051–1059. [Google Scholar] [CrossRef]
- Charoensiddhi, S.; Lorbeer, A.J.; Lahnstein, J.; Bulone, V.; Franco, C.M.M.; Zhang, W. Enzyme-assisted extraction of carbohydrates from the brown alga Ecklonia radiata: Effect of enzyme type, pH and buffer on sugar yield and molecular weight profiles. Process Biochem. 2016, 10, 1503–1510. [Google Scholar] [CrossRef]
- Younes, I.; Hajji, S.; Frachet, V.; Rinaudo, M.; Jellouli, K.; Nasri, M. Chitin extraction from shrimp shell using enzymatic treatment. Antitumor, antioxidant and antimicrobial activities of chitosan. Int. J. Biol. Macromol. 2014, 69, 489–498. [Google Scholar] [CrossRef] [PubMed]
- Sovik, S.L.; Rustad, T. Effect of season and fishing ground on the activity of lipases in byproducts from cod (Gadus morhua). LWT Food Sci. Technol. 2005, 38, 867–876. [Google Scholar] [CrossRef]
- Fernandes, P. Enzymes in fish and seafood processing. Front. Bioeng. Biotechnol. 2016, 4, 59. [Google Scholar] [CrossRef] [PubMed]
- Blamey, J.M.; Fischer, F.; Meyer, H.P.; Sarmiento, F.; Zinn, M. Enzymatic biocatalysis in chemical transformations: A promising and emerging field in green chemistry practice. In Biotechnology of Microbial Enzymes; Brahmachari, G., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 347–403. [Google Scholar]
- Doukyu, N.; Ogino, H. Organic solvent-tolerant enzymes. Biochem. Eng. J. 2010, 48, 270–282. [Google Scholar] [CrossRef]
- Hardouin, K.; Bedoux, G.; Burlot, A.S.; Nyvall-Collén, P.; Bourgougnon, N. Enzymatic recovery of metabolites from seaweeds: Potential applications. In Advances in Botanical Research; Nathalie Bourgougnon, N., Ed.; Academic Press: San Diego, CA, USA, 2014; Volume 71, pp. 279–320. [Google Scholar]
- Aljawish, A.; Chevalot, I.; Jasniewski, J.; Scher, J.; Muniglia, L. Enzymatic synthesis of chitosan derivatives and their potential applications. J. Mol. Catal. B Enzym. 2015, 112, 25–39. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, R.; Xiao, A.; Li, L.; Jiang, Z.; Chen, F.; Ni, H. Characterization of an alkaline β-agarase from Stenotrophomonas sp. NTa and the enzymatic hydrolysates. Int. J. Biol. Macromol. 2016, 86, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Fu, X.; Yan, Q.; Guo, Y.; Liu, Z.; Jiang, Z. Cloning, expression, purification and application of a novel chitinase from a thermophilic marine bacterium Paenibacillus barengoltzi. Food Chem. 2016, 192, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- García-Fraga, B.; da Silva, A.; López-Seijas, J.; Sieiro, C. A novel family 19 chitinase from the marine-derived Pseudoalteromonas tunicata CCUG 44952T: Heterologous expression, characterization and antifungal activity. Biochem. Eng. J. 2015, 93, 84–93. [Google Scholar] [CrossRef]
- Kermanshahi-pour, A.; Sommer, T.; Anastas, P.; Zimmerman, J. Enzymatic and acid hydrolysis of Tetraselmis suecica for polysaccharide characterization. Bioresour. Technol. 2014, 173, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Jana, S.; Gandhi, A.; Sen, K.; Zhiang, W.; Kokare, C. Gellan gum microspheres containing a novel α-amylase from marine Nocardiopsis sp. strain B2 for immobilization. Int. J. Biol. Macromol. 2014, 70, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Menshova, R.; Ermakova, S.; Anastyuk, S.; Isakov, V.; Dubrovskaya, Y.; Kusaykin, M.; Um, B.; Zvyagintseva, T. Structure, enzymatic transformation and anticancer activity of branched high molecular weight laminaran from brown alga Eisenia bicyclis. Carbohydr. Polym. 2014, 99, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, A.; Nishiyama, N.; Ohshima, M.; Maniwa, S.; Kuwamura, S.; Shiraishi, M.; Yuasa, K. Comprehensive enzymatic analysis of the amylolytic system in the digestive fluid of the sea hare, Aplysia kurodai: Unique properties of two α-amylases and two α-glucosidases. FEBS Open Bio 2014, 4, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Trincone, A.; Pagnotta, E.; Tramice, A. Enzymatic routes for the production of mono- and di-glucosylated derivatives of hydroxytyrosol. Bioresour. Technol. 2012, 115, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Shchipunov, Y.; Burtseva, Y.; Karpenko, T.; Shevchenko, N.; Zvyagintseva, T. Highly efficient immobilization of endo-1,3-β-d-glucanases (laminarinases) from marine mollusks in novel hybrid polysaccharide-silica nanocomposites with regulated composition. J. Mol. Catal. B Enzym. 2006, 40, 16–23. [Google Scholar] [CrossRef]
- Ilankovan, P.; Hein, S.; Ng, C.; Trung, T.; Stevens, W. Production of N-acetyl chitobiose from various chitin substrates using commercial enzymes. Carbohydr. Polym. 2006, 63, 245–250. [Google Scholar] [CrossRef]
- Shchipunov, Y.; Karpenko, T.; Bakunina, I.; Burtseva, Y.; Zvyagintseva, T. A new precursor for the immobilization of enzymes inside sol–gel-derived hybrid silica nanocomposites containing polysaccharides. J. Biochem. Biophys. Methods 2004, 58, 25–38. [Google Scholar] [CrossRef]
- Shimoda, K. Efficient preparation of β-(1→6)-(GlcNAc)2 by enzymatic conversion of chitin and chito-oligosaccharides. Carbohydr. Polym. 1996, 29, 149–154. [Google Scholar] [CrossRef]
- Inoue, A.; Nishiyama, R.; Ojima, T. The alginate lyases FlAlyA, FlAlyB, FlAlyC, and FlAlex from Flavobacterium sp. UMI-01 have distinct roles in the complete degradation of alginate. Algal Res. 2016, 19, 355–362. [Google Scholar] [CrossRef]
- Coste, O.; Malta, E.; López, J.; Fernández-Díaz, C. Production of sulfated oligosaccharides from the seaweed Ulva sp. using a new ulvan-degrading enzymatic bacterial crude extract. Algal Res. 2015, 10, 224–231. [Google Scholar] [CrossRef]
- Yang, M.; Mao, X.; Liu, N.; Qiu, Y.; Xue, C. Purification and characterization of two agarases from Agarivorans albus OAY02. Process Biochem. 2014, 49, 905–912. [Google Scholar] [CrossRef]
- Tramice, A.; Pagnotta, E.; Romano, I.; Gambacorta, A.; Trincone, A. Transglycosylation reactions using glycosyl hydrolases from Thermotoga neapolitana, a marine hydrogen-producing bacterium. J. Mol. Catal. B Enzym. 2007, 47, 21–27. [Google Scholar] [CrossRef]
- Wu, S. Degradation of κ-carrageenan by hydrolysis with commercial α-amylase. Carbohydr. Polym. 2012, 89, 394–396. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Du, J.; Huang, Q.; Shi, Y.; Chen, Q. Inhibitory kinetics of bromacetic acid on β-N-acetyl-d-glucosaminidase from prawn (Penaeus vannamei). Int. J. Biol. Macromol. 2007, 41, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Martinou, A.; Kafetzopoulos, D.; Bouriotis, V. Chitin deacetylation by enzymatic means: Monitoring of deacetylation processes. Carbohydr. Res. 1995, 273, 235–242. [Google Scholar] [CrossRef]
- Shin, S.; Sim, J.; Kishimura, H.; Chun, B. Characteristics of menhaden oil ethanolysis by immobilized lipase in supercritical carbon dioxide. J. Ind. Eng. Chem. 2012, 18, 546–550. [Google Scholar] [CrossRef]
- Kavitha, V.; Radhakrishnan, N.; Madhavacharyulu, E.; Sailakshmi, G.; Sekaran, G.; Reddy, B.; Rajkumar, G.; Gnanamani, A. Biopolymer from microbial assisted in situ hydrolysis of triglycerides and dimerization of fatty acids. Bioresour. Technol. 2010, 101, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhang, C.; Hong, P.; Ji, H. Lipase-catalysed acylglycerol synthesis of glycerol and n−3 PUFA from tuna oil: Optimisation of process parameters. Food Chem. 2007, 103, 1009–1015. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Sun, A.; Hu, Y. Characterization of a novel marine microbial esterase and its use to make D-methyl lactate. Chin. J. Catal. 2016, 37, 1396–1402. [Google Scholar] [CrossRef]
- Alsufyani, T.; Engelen, A.; Diekmann, O.; Kuegler, S.; Wichard, T. Prevalence and mechanism of polyunsaturated aldehydes production in the green tide forming macroalgal genus Ulva (Ulvales, Chlorophyta). Chem. Phys. Lipids 2014, 183, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Rocha, L.; de Souza, A.; Rodrigues Filho, U.; Campana Filho, S.; Sette, L.; Porto, A. Immobilization of marine fungi on silica gel, silica xerogel and chitosan for biocatalytic reduction of ketones. J. Mol. Catal. B Enzym. 2012, 84, 160–165. [Google Scholar] [CrossRef]
- Mertens, R.; Greiner, L.; van den Ban, E.; Haaker, H.; Liese, A. Practical applications of hydrogenase I from Pyrococcus furiosus for NADPH generation and regeneration. J. Mol. Catal. B Enzym. 2003, 24–25, 39–52. [Google Scholar] [CrossRef]
- Boonprab, K.; Matsui, K.; Akakabe, Y.; Yotsukura, N.; Kajiwara, T. Hydroperoxy-arachidonic acid mediated n-hexanal and (Z)-3- and (E)-2-nonenal formation in Laminaria angustata. Phytochemistry 2003, 63, 669–678. [Google Scholar] [CrossRef]
- Rontani, J.; Beker, B.; Volkman, J. Regiospecific oxygenation of alkenones in the benthic haptophyte Anand HAP 17. Phytochemistry 2004, 65, 3269–3278. [Google Scholar] [CrossRef] [PubMed]
- Kerr, R.; Rodriguez, L.; Keliman, J. A chemoenzymatic synthesis of 9(11)-secosteroids using an enzyme extract of the marine gorgonian Pseudopterogorgia americana. Tetrahedron Lett. 1996, 37, 8301–8304. [Google Scholar] [CrossRef]
- Wischang, D.; Radlow, M.; Schulz, H.; Vilter, H.; Viehweger, L.; Altmeyer, M.; Kegler, C.; Herrmann, J.; Müller, R.; Gaillard, F.; et al. Molecular cloning, structure, and reactivity of the second bromoperoxidase from Ascophyllum nodosum. Bioorg. Chem. 2012, 44, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Saborowski, R.; Sahling, G.; del Toro, M.A.N.; Walter, I.; Garcı́a-Carreño, F. Stability and effects of organic solvents on endopeptidases from the gastric fluid of the marine crab Cancer pagurus. J. Mol. Catal. B Enzym. 2004, 30, 109–118. [Google Scholar] [CrossRef]
- Sana, B.; Ghosh, D.; Saha, M.; Mukherjee, J. Purification and characterization of a salt, solvent, detergent and bleach tolerant protease from a new gamma-Proteobacterium isolated from the marine environment of the Sundarbans. Process Biochem. 2006, 41, 208–215. [Google Scholar] [CrossRef]
- Wang, Q.; Dordick, J.S.; Linhardt, R.J. Synthesis and application of carbohydrate-containing polymers. Chem. Mater. 2002, 14, 3232–3244. [Google Scholar] [CrossRef]
- Wang, L.; Chi, Z.; Yue, L.; Chi, Z.; Zhang, D. Occurrence and diversity of Candida genus in marine environments. J. Ocean Univ. China 2008, 7, 416–420. [Google Scholar] [CrossRef]
- Andreou, A.; Brodhun, F.; Feussner, I. Biosynthesis of oxylipins in non-mammals. Prog. Lipid Res. 2009, 48, 148–170. [Google Scholar] [CrossRef] [PubMed]
- Johnston, A.; Green, R.; Todd, J. Enzymatic breakage of dimethylsulfoniopropionate—A signature molecule for life at sea. Curr. Opin. Chem. Biol. 2016, 31, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Van der Oost, R.; Beyer, J.; Vermeulen, N. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol. 2003, 13, 57–149. [Google Scholar] [CrossRef]
- Apraiz, I. Identification of proteomic signatures of exposure to marine pollutants in mussels (Mytilus edulis). Mol. Cell. Proteom. 2006, 5, 1274–1285. [Google Scholar] [CrossRef] [PubMed]
- Hühnerfuss, H. Chromatographic enantiomer separation of chiral xenobiotics and their metabolites–A versatile tool for process studies in marine and terrestrial ecosystems. Chemosphere 2000, 40, 913–919. [Google Scholar] [CrossRef]
- Demarche, P.; Junghanns, C.; Nair, R.; Agathos, S. Harnessing the power of enzymes for environmental stewardship. Biotechnol. Adv. 2012, 30, 933–953. [Google Scholar] [CrossRef] [PubMed]
- Daffonchio, D.; Ferrer, M.; Mapelli, F.; Cherif, A.; Lafraya, Á.; Malkawi, H.; Yakimov, M.; Abdel-Fattah, Y.; Blaghen, M.; Golyshin, P.; et al. Bioremediation of Southern Mediterranean oil polluted sites comes of age. New Biotechnol. 2013, 30, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Pan, H.; Xu, J.; Xu, W.; Liu, L. Acclimation of a marine microbial consortium for efficient Mn(II) oxidation and manganese containing particle production. J. Hazard. Mater. 2016, 304, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Rontani, J.-F.; Bonin, P.; Vaultier, F.; Guasco, S.; Volkman, J.K. Anaerobic bacterial degradation of pristenes and phytenes in marine sediments does not lead to pristane and phytane during early diagenesis. Org. Geochem. 2013, 58, 43–55. [Google Scholar] [CrossRef]
- Châtel, A.; Hamer, B.; Talarmin, H.; Dorange, G.; Schröder, H.C.; Müller, W.E.G. Activation of MAP kinase signaling pathway in the mussel Mytilus galloprovincialis as biomarker of environmental pollution. Aquat. Toxicol. 2010, 96, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Cheung, K.H.; Gu, J.-D. Reduction of chromate (CrO42−) by an enrichment consortium and an isolate of marine sulfate-reducing bacteria. Chemosphere 2003, 52, 1523–1529. [Google Scholar] [CrossRef]
- Chang, C.-C.; Rahmawaty, A.; Chang, Z.-W. Molecular and immunological responses of the giant freshwater prawn, Macrobrachium rosenbergii, to the organophosphorus insecticide, trichlorfon. Aquat. Toxicol. 2013, 130–131, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.E. The toxicology and metabolism of chlorothalonil in fish. III. Metabolism, enzymatics and detoxication in Salmo spp. and Galaxias spp. Aquat. Toxicol. 1985, 7, 277–299. [Google Scholar] [CrossRef]
- Agatova, A.I.; Andreeva, N.M.; Kucheryavenko, A.V.; Torgunova, N.I. Transformation of organic matter in areas inhabited by natural and artificially cultured populations of marine invertebrates in the bay of Pos’et (sea of Japan). Aquaculture 1986, 53, 49–66. [Google Scholar] [CrossRef]
- Pfaffenberger, B.; Hühnerfuss, H.; Kallenborn, R.; Köhler-Günther, A.; König, W.A.; Krüner, G. Chromatographic separation of the enantiomers of marine pollutants. Part 6: Comparison of the enantioselective degradation of α-hexachlorocyclohexane in marine biota and water. Chemosphere 1992, 25, 719–725. [Google Scholar] [CrossRef]
- Regoli, F.; Principato, G. Glutathione, glutathione-dependent and antioxidant enzymes in mussel, Mytilus galloprovincialis, exposed to metals under field and laboratory conditions: Implications for the use of biochemical biomarkers. Aquat. Toxicol. 1995, 31, 143–164. [Google Scholar] [CrossRef]
- Warshawsky, D.; Cody, T.; Radike, M.; Reilman, R.; Schumann, B.; LaDow, K.; Schneider, J. Biotransformation of benzo[a]pyrene and other polycyclic aromatic hydrocarbons and heterocyclic analogs by several green algae and other algal species under gold and white light. Chem. Biol. Interact. 1995, 97, 131–148. [Google Scholar] [CrossRef]
- Hahlbeck, E.; Arndt, C.; Schiedek, D. Sulphide detoxification in Hediste diversicolor and Marenzelleria viridis, two dominant polychaete worms within the shallow coastal waters of the southern Baltic Sea. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2000, 125, 457–471. [Google Scholar] [CrossRef]
- Goto, N.; Mitamura, O.; Terai, H. Biodegradation of photosynthetically produced extracellular organic carbon from intertidal benthic algae. J. Exp. Mar. Biol. Ecol. 2001, 257, 73–86. [Google Scholar] [CrossRef]
- Company, R.; Serafim, A.; Bebianno, M.J.; Cosson, R.; Shillito, B.; Fiala-Médioni, A. Effect of cadmium, copper and mercury on antioxidant enzyme activities and lipid peroxidation in the gills of the hydrothermal vent mussel Bathymodiolus azoricus. Mar. Environ. Res. 2004, 58, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Gallizia, I.; Vezzulli, L.; Fabiano, M. Oxygen supply for biostimulation of enzymatic activity in organic-rich marine ecosystems. Soil Biol. Biochem. 2004, 36, 1645–1652. [Google Scholar] [CrossRef]
- Barata, C.; Carlos Navarro, J.; Varo, I.; Carmen Riva, M.; Arun, S.; Porte, C. Changes in antioxidant enzyme activities, fatty acid composition and lipid peroxidation in Daphnia magna during the aging process. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 140, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Buet, A.; Banas, D.; Vollaire, Y.; Coulet, E.; Roche, H. Biomarker responses in European eel (Anguilla anguilla) exposed to persistent organic pollutants. A field study in the Vaccarès lagoon (Camargue, France). Chemosphere 2006, 65, 1846–1858. [Google Scholar] [PubMed]
- Lima, I.; Moreira, S.M.; Osten, J.R.-V.; Soares, A.M.V.M.; Guilhermino, L. Biochemical responses of the marine mussel Mytilus galloprovincialis to petrochemical environmental contamination along the north-western coast of Portugal. Chemosphere 2007, 66, 1230–1242. [Google Scholar] [CrossRef] [PubMed]
- Kankaanpää, H.; Leiniö, S.; Olin, M.; Sjövall, O.; Meriluoto, J.; Lehtonen, K.K. Accumulation and depuration of cyanobacterial toxin nodularin and biomarker responses in the mussel Mytilus edulis. Chemosphere 2007, 68, 1210–1217. [Google Scholar] [CrossRef] [PubMed]
- Correia, A.D.; Gonçalves, R.; Scholze, M.; Ferreira, M.; Henriques, M.A.R. Biochemical and behavioral responses in gilthead seabream (Sparus aurata) to phenanthrene. J. Exp. Mar. Biol. Ecol. 2007, 347, 109–122. [Google Scholar] [CrossRef]
- Koenig, S.; Solé, M. Natural variability of hepatic biomarkers in Mediterranean deep-sea organisms. Mar. Environ. Res. 2012, 79, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.D.; Diab, A.M.; Gubbins, M.; Collins, C.; Matejusova, I.; Kerr, R.; Chipman, J.K.; Kuiper, R.; Vethaak, A.D.; George, S.G. Transcriptomic responses of European flounder (Platichthys flesus) liver to a brominated flame retardant mixture. Aquat. Toxicol. 2013, 142–143, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Arnosti, C. Contrasting patterns of peptidase activities in seawater and sediments: An example from arctic fjords of Svalbard. Mar. Chem. 2015, 168, 151–156. [Google Scholar] [CrossRef]
- Si, Y.-X.; Gu, X.-X.; Cai, Y.; Yin, S.-J.; Yang, J.-M.; Park, Y.-D.; Lee, J.; Qian, G.-Y. Molecular dynamics simulation integrating study for Cr3+-binding to arginine kinase. Process Biochem. 2015, 50, 1363–1371. [Google Scholar] [CrossRef]
- Da Fonseca, F.S.A.; Angolini, C.F.F.; Arruda, M.A.Z.; Junior, C.A.L.; Santos, C.A.; Saraiva, A.M.; Pilau, E.; Souza, A.P.; Laborda, P.R.; de Oliveira, P.F.L.; et al. Identification of oxidoreductases from the petroleum Bacillus safensis strain. Biotechnol. Rep. 2015, 8, 152–159. [Google Scholar] [CrossRef]
- Pi, Y.; Meng, L.; Bao, M.; Sun, P.; Lu, J. Degradation of crude oil and relationship with bacteria and enzymatic activities in laboratory testing. Int. Biodeter. Biodegrad. 2016, 106, 106–116. [Google Scholar] [CrossRef]
- Nicemol, J.; Niladevi, K.N.; Anisha, G.S.; Prema, P. Hydrolysis of pectin: An enzymatic approach and its application in banana fiber processing. Microbiol. Res. 2008, 163, 538–544. [Google Scholar]
- Fines, B.C.; Holt, G.J. Chitinase and apparent digestibility of chitin in the digestive tract of juvenile cobia. Rachycentron canadum. Aquaculture 2010, 303, 34–39. [Google Scholar] [CrossRef]
- Ziervogel, K.; Arnosti, C. Enzyme activities in the Delaware Estuary affected by elevated suspended sediment load. Estuar. Coast. Shelf Sci. 2009, 84, 253–258. [Google Scholar] [CrossRef]
- Richir, J.; Velimirov, B.; Poulicek, M.; Gobert, S. Use of semi-quantitative kit methods to study the heterotrophic bacterial community of Posidonia oceanica meadows: Limits and possible applications. Estuar. Coast. Shelf Sci. 2012, 109, 20–29. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. Carbonic Anhydrases from Extremophiles and Their Biotechnological Applications. In Carbonic Anhydrases as Biocatalysts; Supuran, C.T., De Simone, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 311–324. [Google Scholar]
- Migliardini, F.; De Luca, V.; Carginale, V.; Rossi, M.; Corbo, P.; Supuran, C.T.; Capasso, C. Biomimetic CO2 capture using a highly thermostable bacterial alpha-carbonic anhydrase immobilized on a polyurethane foam. J. Enzyme Inhib. Med. Chem. 2013, 29, 146–150. [Google Scholar] [CrossRef] [PubMed]
- Lionetto, M.G.; Caricato, R.; Giordano, M.E.; Erroi, E.; Schettino, T. Carbonic anhydrase as pollution biomarker: An ancient enzyme with a new use. Int. J. Environ. Res. Public Health 2012, 9, 3965–3977. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, A.; Scozzafava, S.; Parkkila, L.; Puccetti, G.; de Simone, G.; Supuran, C.T. Investigations of the esterase, phosphatase, and sulfatase activities of the cytosolic mammalian carbonic anhydrase isoforms I, II, and XIII with 4-nitrophenyl esters as substrates. Bioorg. Med. Chem. Lett. 2008, 18, 2267–2271. [Google Scholar] [CrossRef] [PubMed]
- Trincone, A. Biocatalytic processes using marine biocatalysts: Ten cases in point. Curr. Org. Chem. 2013, 17, 1058–1066. [Google Scholar] [CrossRef]
- Dawfik, D.S.; van der Donk, W.A. Editorial overvies: Biocatalysis and biotransformation: Esoteric, niche enzymology. Curr. Opin. Chem. Biol. 2016, 31, v–vii. [Google Scholar] [CrossRef]
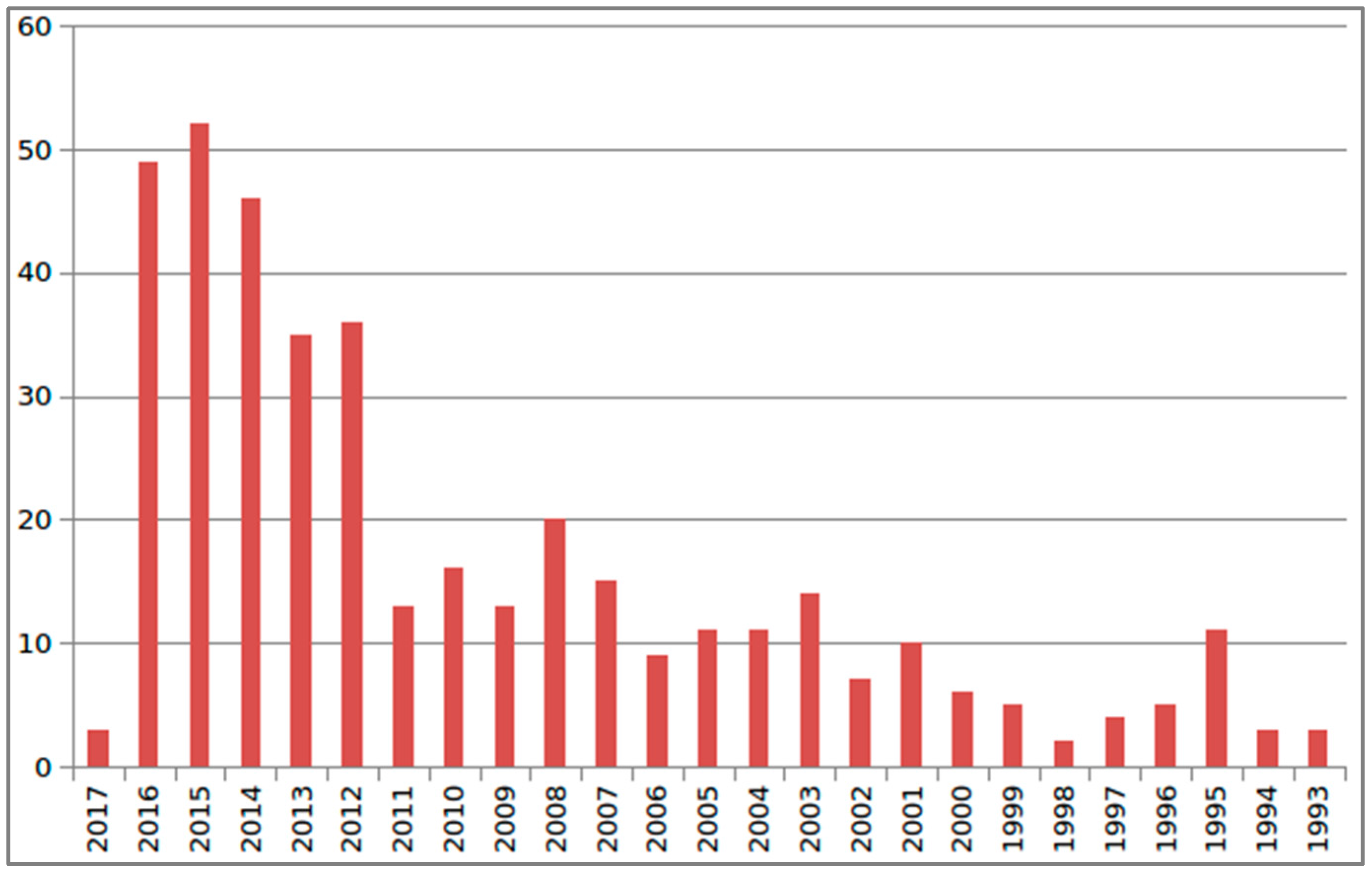

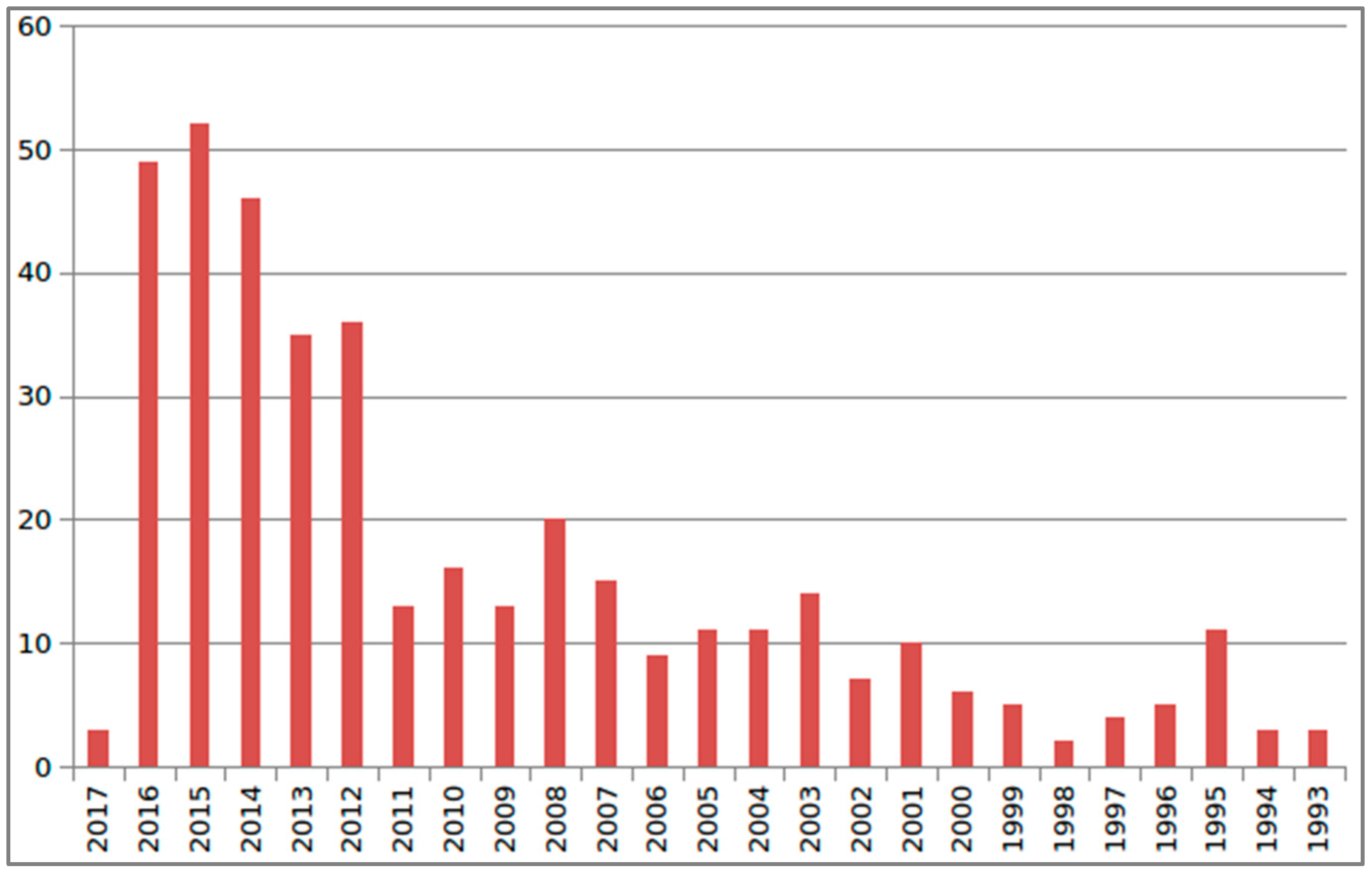
{kind=link}
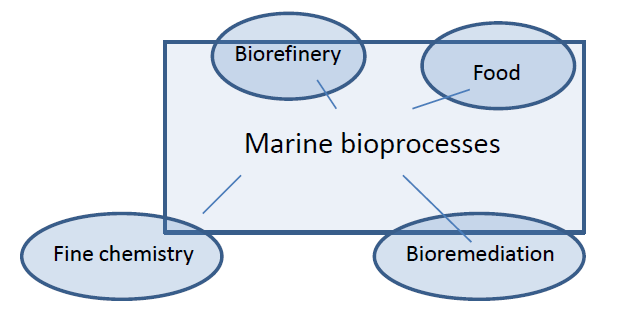
{kind=link}
| Entry | Reference | Biocatalyst(s) | Biomass | Notes |
|---|---|---|---|---|
| Cellulases and other important carbohydrate active hydrolases | ||||
| 1/2015 | [9] | Cellulase of marine fungus Cladosporium sphaerospermum | Seaweed biomass Ulva fasciata | Cellulases found are active and stable in different ionic liquids |
| 2/2014 | [10] | Lignocellulose depolymerizing multi-enzyme complex: lignin peroxidase, xylanase and cellulases | 13 microbial marine isolates from seaweed were studied. They belong to the genera Brachybacterium, Brevibacterium, Halomonas, Kokuria, Micrococcus, Nocardiopsis, Pseudomonas and Streptomyces | |
| 3/2014 | [11] | Cellulase from a marine bacterium, Bacillus carboniphilus | Optimization study of saccharification using marine microbial cellulase | |
| 4/2013 | [12] | Cellulase from a marine Bacillus sp. H1666 | Ulva lactuca macro algae is studied for cellulase treatment | Isolated enzyme has saccharification applicability on Ulva lactuca |
| 5/2009 | [13] | Cellulase isolated from a marine bacterium, Bacillus subtilis subsp. subtilis A-53 | ||
| 6/2014 | [14] | κ-carrageenase CgkA and λ-carrageenase CglA from Pseudoalteromonas carrageenovora | Carrageenan from red algae | Improvement of the process of degradation by the study of functional carrageenolytic complex |
| 7/2016 | [15] | Endo-type β-agarase AgaG1, screened from Alteromonas sp. GNUM1; exo-type β-agarase DagB from S. coelicolor A3 and a α-neoagarobiose hydrolase from Alcanivorax sp. | Agarose | Enzymatic agarose hydrolysis process without acid pretreatment |
| 8/2012 | [16] | Mimicked the natural agarolytic pathway using three microbial agarases (Aga16B, Aga50D and DagA) and NABH | Recalcitrant agar polysaccharide | |
| Feedstock oils to biodiesel | ||||
| 9/2016 | [17] | - | Marine microalgae | Optimization study for disruption of thraustochytrid cell using bead mill for maximising lipid extraction yields and hydrolysis of oil extracted studied using commercial lipases |
| 10/2013 | [18] | Bacterial isolate Flammeovirga yaeyamensis | Oil-rich microalga (Chlorella vulgaris ESP-1) | Cell-wall destruction analyzed by SEM micrographs was associated with the activity of hydrolytic enzymes |
| 11/2015 | [19] | Chlorella protothecoides | Study of fermentations developed to produce lipid by heterotrophic C. protothecoides using cassava bagasse as the low-cost feedstock | |
| 12/2014 | [20] | Nannochloropsis oceanica | Crude algal oils were extracted from the oleaginous microalga Nannochloropsis oceanica | |
| Marine feedstock valorization | ||||
| 13/2015 | [21] | Three enzymes used: alcalase, neutrase and esperase | Effluents obtained from chemical and enzymatic chitin production of Illex argentinus pen byproducts | Study for production of lactic acid bacteria, marine probiotic bacteria and two common gram (+) bacteria using effluents as substrates |
| 14/2015 | [22] | Chemical treatment | Mussel processing wastewaters | Laboratory optimization to transform mussel processing wastewater into a growth culture medium to produce microbial biomass. The lab-scale processes studied were upscaled to a pre-industrial level using a 70-L fermenter |
| 15/2015 | [23] | Sulfuric acid was seen as the best catalyst with a lipid conversion efficiency of 44.9% | Marine microalga Chlorella sp. BDUG 91771 | - |
| 16/2016 | [24] | Pretreatment with hydrogen peroxide | Seaweed Ulva prolifera | Optimization study |
| 17/1994 | [25] | Commercial bacterial inoculum (Stabisil) | Crustacean shell waste from the world’s fishing industry | Optimization study for recovery of protein, pigment and chitin from waste shell of prawn |
| 18/2017 | [26] | Cellulase and pectinase | Porphyridium cruentum, red microalagae | Evaluation of bioethanol production in response culture conditions of to Porphyridium cruemtum. Enzymatic hydrolysis resulted in high glucose conversion yields for both seawater and freshwater conditions |
| 19/2016 | [27] | Commercial cellulases and alginases | Brown algae Macrocystis pyrifera | Study of various pretreatments |
| 20/2016 | [28] | Mixture of commercial enzymes: Viscozyme® L, Cellic® CTec2, Cellic® HTec2 | Macro-algae Gracilaria verrucosa | |
| 21/2016 | [29] | Chemical and enzymatic process with commercial cellulase and β-glucosidase | Red macroalgae Eucheuma cottonii | |
| 22/2015 | [30] | Celluclast and commercial alginate lyase (EC4.2.2.3) from Sphingobacterium spiritivorum were used | Brown seaweed Laminaria digitata | |
| 23/2015 | [31] | Commercial cellulase, xylanase and β-glucosidase | Nannochloropsis salina | Anaerobic digestion study |
| 24/2015 | [32] | Commercially available enzymes (pectinase) and enzyme mixtures (Accellerase 1500, Accellerase XC, and Accellerase XY) with multiple enzyme activities (exoglucanase, endoglucanase, hemi-cellulase, and β-glucosidase were used) | Nannochloropsis salina | Study of conversion of lipid-extracted biomass into fermentable sugars |
| 25/2014 | [33] | Commercial cellulase | Red algae Kappaphycus alvarezii | Optimization study |
| 26/2014 | [34] | Commercial Viscozyme L and Cellic CTec2 | Marine green macro-algae Enteromorpha intestinalis | Study of hydrotermal method |
| 27/2014 | [35] | Free and immobilized yeast | Red alga Gracilaria sp. | A study for bioethanol production using hydrolisate of Gracilaria |
| 28/2014 | [36] | Yeast fermentation | Microalga Chlorella vulgaris | Pectinase enzyme was used for disrupting microalgal cells |
| 29/2013 | [37] | Saccharomyces fermentation | Red alga Gracilaria verrucosa | A study for the combined production of agar and bioethanol; the pulp was used after agar extraction |
| 30/2013 | [38] | Saccharomyces fermentation | Red algae Kappaphycus alvarezii | 105 L of ethanol per ton of seaweed were obtained after a dilute acid pretreatment |
| 31/2012 | [39] | Yeast fermentation | Aquatic plant Zostera marina | Study of the potential of this plant as a source of bioactives and sugars for bioethanol production |
| 32/2011 | [40] | Saccharomyces fermentation | Byproduct from the alginate extraction process | Interesting study for exploitation of seaweed waste from alginate production |
| 33/2015 | [41] | - | Red alga Gracilaria verrucosa | Optimization study of this suitable feedstock for biosugar production |
| 34/2015 | [42] | β-1,3-glucanase, cellulase and β-glucosidase were studied | Red seaweed Grateloupia turuturu | Enzyme-assisted extraction of R-phycoerythrin with ultrasound technology |
| Entry | Reference | Biocatalyst(s) | Biomass | Notes |
|---|---|---|---|---|
| 1/2007 | [62] | Antibacterial alkaline protease | Fish processing waste | Action exerted by cell lysis of pathogenic bacteria |
| 2/2016 | [63] | Alcalase for oil extraction | Thunnus albacares byproducts (heads) | Study for deodorization of fish oil |
| 3/2016 | [64] | Commercial alcalase | Shrimp waste | Response surface methodology study to grow hydrocarbon-degrading bacteria Bacillus subtilis |
| 4/2016 | [65] | Enzymatic deproteinization by commercial enzyme savinase | Norway lobster (Nephrops norvegicus) processing byproducts | Chitin and chitosan production |
| 5/2016 | [66] | Commercial enzymes used for the preparation of seaweed: Celluclast and Alcalase | Brown seaweed Ecklonia radiata | Study of extraction methods of the alga and potential in vitro prebiotic effect |
| 6/2016 | [67] | Commercial alcalase | Wastewater generated during shrimp cooking | Study for the production of enzymic hydrolysates with antioxidant capacity and production of essential amino acids |
| 7/2016 | [68] | Commercial alcalase | Head byproducts of Prionace glauca | Production of chondroitin sulphate from blue shark waste was studied after cartilage hydrolysis with alcalase |
| 8/2016 | [69] | Alcalase | Adhesive gum layer surrounding naturally fertilised ballan wrasse (Labrus bergylta) eggs | A study for the biological control (by cleaner fish Labrus bergylta) of sea lice in farming Salmo salar |
| 9/2016 | [70] | Commercial lipase B from C. antarctica (Lipozyme 435, immobilized lipase) | Sardine oil | Sardine oil was evaluated by glycerolysis using commercial lipase to produce monoacyl glycerols rich in omega-3 polyunsaturated fatty acids |
| 10/2015 | [71] | Proteases from Bacillus subtilis A26 (TRMH-A26), Raja clavata crude alkaline protease extract, alcalase and neutrase | Thornback ray (Raja clavata) muscle | Study of bioactivity of extracts after proteolytic hydrolysis with different enzyme preparations |
| 11/2015 | [72] | - | Brines marinated herring (Clupea harengus) | Brines from marinated herring processing used for recovery of useful material |
| 12/2015 | [73] | Commercial proteases | Atlantic salmon (Salmo salar) rest raw materials | Study of the production of different hydrolysates using commercial enzymes for the valorization of viscera-containing raw material from Atlantic salmon |
| 13/2015 | [74] | Marine proteases | Red scorpionfish (Scorpaena scrofa) viscera | Alkaline proteases of marine origin suggested for detergent formulations and deproteinization of shrimp shells |
| 14/2015 | [75] | Commercial alcalase, pepsin and trypsin | Common carp (Cyprinus carpio) egg | Hydrolysates improve the immune system with differential influences on the immune function. Interesting study for several applications in the health food, pharmaceutical, and nutraceutical industries |
| 15/2014 | [76] | Hydrolysis by bromelain | Protein byproducts of seaweed (Gracilaria sp.) | Set up of a flavouring agent with umami taste and seaweed odour |
| 16/2014 | [77] | Different commercial proteases | Fresh herring byproducts | Enzymatic hydrolysis to produce fish protein hydrolysates and separate oil |
| 17/2014 | [78] | Hydrolysis by commercial proteases | Cod (Gadus morhua) fillets | Study of influences of oxidative processes during protein hydrolysis using cod |
| 18/2013 | [79] | Proteolytic processing with commercial proteases | Fractions obtained from processing of Atlantic rock crab (Cancer irroratus) byproducts | Small peptides with biological activity recovered |
| 19/2013 | [80] | Commercial alcalase | Protein concentrates recovered from cuttlefish processing wastewater | Selective ultrafiltration methods under study for concentrating active components with antihypertensive and antioxidant activities |
| 20/2013 | [81] | Commercial alcalase | Tuna dark muscle | Basic study for fractionation of protein hydrolysates with ultrafiltration and nanofiltration |
| 21/2012 | [82] | Proteases and lipases from marine waste | Byproducts of Monterey sardine (Sardinops sagax caerulea) processing | Actions of enzymes from sardine byproduct (viscera and byproduct concentrate extracts) produced 3-fold greater hydrolysis than with the commercial enzyme |
| 22/2012 | [83] | Trypsin and alcalase | Waste byproducts of red seaweed Porphyra columbina | Study on protein water extracts wasted during traditional phycollloids extraction procedure from P. columbina. Interesting immunosuppressive effects and antihypertensive and antioxidant activities found |
| 23/2012 | [84] | Commercial alcalase | Fish byproducts | Comparison of methods including enzymatic extraction |
| 24/2009 | [85] | Proteolytic commercial enzyme mix | Snow crab (Chionoecetes opilio) byproduct fractions | Pilot scale enzymatic hydrolysis to entire snow crab byproducts followed by fractionation operations in order to recover enriched fractions of proteins, lipids and chitin |
| 25/2009 | [86] | Commercial alcalase preparations | Gadus morrhua skin collagen | Optimization of parameters for the hydrolysis |
| 26/2008 | [87] | Three types of enzymes used: papain, trypsin and pepsin | Wastewater from the industrial processing of octopus | Marine peptones as promising alternatives to expensive commercial medium for growth of lactic acid bacteria |
| 27/2008 | [88] | Commerical proteases | Dosidicus gigas mantle | Tenderization of mantle for commercial use as substitute of Illex argentinus |
| 28/2007 | [89] | Alcalase | Shrimp processing discards | Isolation and characterisation of a natural antioxidant from shrimp waste |
| 29/2005 | [90] | Alcalase, Lecitase® a carboxylic ester hydrolase with inherent activity towards both phospholipid and triacylglycerol structures | Cod (Gadus morhua) byproducts | Study for protein and oil extractions |
| 30/2005 | [91] | Flavourzyme, a fungal protease/peptidase complex produced by Aspergillus oryzae, and Neutrase | Cod (Gadus morhua) byproducts | Composition of products generated by hydrolyses of byproducts of cod processing for optimization and design on desired product |
| 31/2003 | [92] | Crude papain was selected to perform the enzymatic extraction | Skate cartilage | Study for a low-cost process for glycosaminoglycan extraction from skate cartilage |
| 32/2002 | [93] | Umamizyme (commercial endo-peptidase activity from a strain of A. oryzae) | Tuna waste | Study for evaluation of activity of Umamizyme in comparison to other fungal enzymes |
| 33/2001 | [94] | Alcalase | Yellowfin tuna (Thunnus albacares) waste | Study of hydrolysis of tuna stomach proteins |
| 34/2016 | [95] | Six commercial enzyme mixtures and individual enzymes were used: Viscozyme® L, Celluclast® 1.5 L, Ultraflo® L and the three proteases Alacalase® 2.4 L FG, Neutrase® 0.8 L and Flavourzyme® 1000 L. | Brown alga Ecklonia radiata | Study of enzyme-assisted extraction of carbohydrates for the design and optimization of processes to obtain oligo- and polysaccharides |
| 35/2014 | [96] | Proteolytic preparations from Bacillus mojavensis A21, Bacillus subtilis A26, Bacillus licheniformis NH1, B. licheniformis MP1, Vibrio metschnikovii J, Aspergillus clavatus ES1 and crude alkaline protease extracts from Sardinelle (Sardinella aurita), Goby (Zosterisessor ophiocephalus) and Grey triggerfish (Balistes capriscus) prepared and characterized by the group | Shrimp processing byproducts | Enzymatic deproteinization for extraction of chitin |
| Entry | Reference | Biocatalyst(s) | Product(s) | Feedstocks | Notes |
|---|---|---|---|---|---|
| Carbohydrate active hydrolases | |||||
| 1/2016 | [103] | Alkaline β-agarase from marine bacterium Stenotrophomonas sp. NTa | From agarose as substrate neoagarobiose, neoagarotetraose and neoagarohexaose are the predominant products | - | First evidence of extracellular agarolytic activity in Stenotrophomonas, the enzyme exhibited stability across a wide pH range and resistance against some inhibitors, detergents and denaturants |
| 2/2016 | [104] | Cloned novel chitinase from a marine bacterium Paenicibacillus barengoltzii functionally expressed in E. coli | The chitinase hydrolyzed colloidal chitin to yield mainly N-acetyl chitobiose | Chitin (from crab shells) | Production of 21.6 mg·mL−1 of N-acetyl chitobiose from colloidal chitin with the highest conversion yield of 89.5% (w/w) |
| 3/2015 | [105] | Chitinase from the marine-derived Pseudoalteromonas tunicata CCUG 44952T | Active also on chromogenic substrate pNP-(GlcNAc) but not on pNP-(GlcNAc)2 and pNP-(GlcNAc)3 | Colloidal and crystalline chitin | The recombinant enzyme exhibited antifungal activity against phytopathogenic and human pathogenic fungi, (biofungicide) |
| 4/2014 | [106] | Commercial pectinase or acidic hydrolysis | 3-deoxy-d-manno-oct-2-ulosonic acid (Kdo): a sugar that is difficult to obtain by chemical synthesis and that has applications in medicinal chemistry | Marine microalgae, Tetraselmis suecica | Evaluation of T. suecica as feedstock for a KDO production |
| 5/2014 | [107] | α-amylase from marine Nocardiopsis sp. strain B2 | - | - | Study for immobilization of a marine α-amylase by ionotropic gelation technique using gellan gum (GG) |
| 6/2014 | [108] | Endo- and exo-glucanases from marine sources: endo-1,3-β-d-glucanase (LIV) from Pseudocardium sacchalinensis and the exo-1,3-β-d-glucanase from Chaetomium indicum | Different fractions of oligosaccharides | Laminaran from brown alga Eisenia bicyclis | Study for anticancer activity of the native laminaran and products of its enzymatic hydrolysis |
| 7/2014 | [109] | Amylolytic system in the digestive fluid of the sea hare, Aplysia kurodai | Maltotriose, maltose, and glucose | Sea lettuce (Ulva pertusa) | Enzymatic analysis of the amylolytic system in the digestive fluid of the sea hare Aplysia kurodai and efficient production of glucose from sea lettuce |
| 8/2012 | [110] | α-glucosidase from Aplysia fasciata | Glucosylated anti-oxidant derivatives of hydroxytyrosol | - | Biocatalytic production of mono- and disaccharide derivatives at final concentrations of 9.35 and 10.8 g/L of reaction |
| 9/2006 | [111] | Endo-1,3-β-d-glucanases (laminarinases) from marine mollusks Spisula sacchalinensis and Chlamys albidus | Biologically active 1,3;1,6-β-d-glucan, called translam | Hydrolysis of laminaran | Study of immobilization |
| 10/2006 | [112] | Commercial enzymes | N-acetyl chitobiose | Various chitin substrates α-chitin from shrimp waste | Experimental conditions studied to achive 10% N-acetyl chitobiose |
| 11/2004 | [113] | 1→3-β-d-glucanase LIV from marine mollusk Spisula sacchalinensis and α-d-galactosidase from marine bacterium Pseudoalteromonas sp. KMM 701 | Oligo- and polysaccharide derivatives possessing immunostimulating, antiviral, anticancer and/or radioprotective activity | Laminaran from the brown seaweeds Laminaria cichorioides | Immobilization study |
| 12/1996 | [114] | Chitin degrading enzymes from sea water bacterium strain identified as Alteromonas | β-(1→6)-(GlcNAc)2 | Chitin and chito-oligosaccharides | High transglycosylation activity of the enzyme preparation was also confirmed |
| 13/2016 | [115] | Endolytic alginate lyases | 4-deoxy-l-erythro-5-hexoseulose uronic acid | Alginate and alginate oligosaccharides | In depth study of degradation process from alginate to unsaturated monosaccharides |
| 14/2015 | [116] | Ulvan-degrading bacterial β-lyase from a new Alteromonas species | Sulfated oligosaccharides from the seaweed Ulva | Ulvan | Fractions of molecular weight down to a 5 kDa of oligosaccharides mix are obtained |
| 15/2014 | [117] | Extracellular β-agarases from Agarivorans albus OAY2 | Neoagarobiose NA2, neoagarotetraose NA4 and neoagarohexaose NA6 | Report about enzyme purification and oligosaccharides preparation | |
| 16/2007 | [118] | Glycosyl hydrolases in crude extracts from extremophilic marine bacterium Thermotoga neapolitana (DSM 4359) | (β-1,4)-xylooligosaccharides of 1-hexanol, 9-fluorene methanol, 1,4-butanediol and geraniol | - | Transglycosylation reactions by xylose, galactose, fucose, glucose and mannose enzymatic transfers |
| 17/2012 | [119] | Commercial α-amylase | Carrageenan-derived oligosaccharide | Hydrolysis of κ-carrageenan | |
| 18/2007 | [120] | β-N-acetyl-d-glucosaminidase from prawn Penaeus vannamei | Mechanistic and inhibition studies | ||
| Ester hydrolysis | |||||
| 19/1995 | [121] | Fungal deacetylase | Hexa-N-deacetylchitohexaose | Natural or artificial chitin substrates as well as N-acetylchito-oligosaccharides | Enzymatic deacetylation: methodological study |
| 20/2012 | [122] | Commercial immobilized lipase, lipozyme from Thermomyces lanuginosa | Diglycerides and monoglycerides containing polyunsaturated fatty acids | Menhaden oil | Enzymatic ethanolysis of menhaden oil |
| 21/2010 | [123] | Organism isolated from marine sediments | Fatty acid-based biopolymer | Triglycerides of sunflower, soybean, olive, sesame and peanut as substrates | Hydrolysis of triglycerides and dimerization of fatty acid to anhydrides and subsequent formation of a Fatty acid based biopolymer (FAbBP) |
| 22/2007 | [124] | Commercial enzymes | Acylglycerol synthesis | N-3 PUFA from tuna oil | - |
| 23/2016 | [125] | Novel marine microbial esterase PHE14 | Asymmetric synthesis of d-methyl lactate by enzymatic kinetic resolution | Racemic methyl lactate commercially available | Esterase PHE14 exhibited very good tolerance to most organic solvents, surfactants and metal ions |
| Oxidoreductases | |||||
| 24/2014 | [126] | Lipoxygenase/hydroperoxide lyase | Polyunsaturated aldehydes: 2,4,7-decatrienal and 2,4-decadienal | Macroalgal genus Ulva (Ulvales, Chlorophyta) | Ulva mutabilis is selected as cultivable for production |
| 25/2012 | [127] | Marine fungi Aspergillus sclerotiorum CBMAI 849 and Penicillium citrinum CBMAI 1186 | Reduction of 1-(4-methoxyphenyl)-ethanone to its stereochemical pure alcohol (ee > 99%, yield = 95%) | Immobilization study | |
| 26/2003 | [128] | Hydrogenase | Enzymatic production and regeneration of NADPH | 6.2 g·L−1 NADPH produced with a total turnover number (ttn: mol produced NADPH/mol consumed enzyme) of 10,000 | |
| 27/2003 | [129] | Lipoxygenase–hydroperoxide lyase pathway | C6 and C9 unsaturated aldehydes | Brown alga Laminaria angustata | Study of biosynthetic pathway |
| 28/2004 | [130] | Cultures of the haptophyte microalga Chrysotila lamellosa | Alkanediones | Regiospecific oxygenation of alkenones | Biogenetic study |
| 29/1996 | [131] | Enzymatic extract of the marine gorgonian Pseudopterogorgia americana | 9(11)-secosteroids | Cholesterol, stigmasterol and progesterone | Claimed as the first chemoenzymatic preparation of a natural product using the enzymatic machinery of a marine invertebrate |
| 30/2012 | [132] | Bromoperoxidase of brown alga Ascophyllum nodosum | 4-bromopyrrole-2-carboxylate | Bromination of methyl pyrrole-2-carboxylate in bromoperoxidase II-catalyzed oxidation | Bromoperoxidase II mimics biosynthesis of methyl 4-bromopyrrole-2-carboxylate, a natural product isolated from the marine sponge Axinella tenuidigitata |
| Proteolytic activities | |||||
| 31/2004 | [133] | Proteolytic enzymes | Products of proteolysis | Gastric fluid of the marine crab, Cancer pagurus | Influence of metal ions and organic solvents other than pH and temperature are analyzed, including long-term stability over a period of several months |
| 32/2006 | [134] | Alkaline serine protease | Marine gamma-Proteobacterium | Activity in presence of up to 30% NaCl. Water miscible and immiscible organic solvents like ethylene glycol, ethanol, butanol, acetone, DMSO, xylene and perchloroethylene enhance as well as stabilize the enzyme activity | |
| Entry | Reference | Biocatalyst(s)/Organism(s) | Application(s) | Notes |
|---|---|---|---|---|
| 1/2016 | [144] | Marine microbial community for manganese oxidation | - | Several new genera associated with Mn(II) oxidation were found in metal-contaminated marine sediments and are seen as a solution for metal bioremediation |
| 2/2013 | [145] | Marine sedimentary bacterial communities | Anaerobic degradation of mixtures of isomeric pristenes and phytenes | Several bacterial products of transformation confirm the key role played by hydration in the metabolism of alkenes |
| 3/2010 | [146] | MAP kinase signaling pathway | - | Different pollutants generated different patterns of induction of the biomarker MAPK phosphorylation |
| 4/2003 | [147] | Isolate (isolate TKW) of sulfate-reducing bacteria | Reduction of chromate (CrO42−) | Soluble hexavalent chromium (Cr6+) enzymatically transformed into less toxic and insoluble trivalent chromium (Cr3+) with potential in bioremediation of sediments contaminated by metals |
| 5/2013 | [148] | Giant freshwater prawn Macrobrachium rosenbergii | Study for potential biomarkers of exposure to organophosphorus pollutants: molecular and immunological responses | Investigation on the effects of the pesticide trichlorfon used in aquaculture, on molecular and enzymatic processes related to the response of the giant freshwater prawn, Macrobrachium rosenbergii |
| 6/1985 | [149] | Five fish species: Salmo gairdneri, S. trutta, Galaxias maculatus, G. truttaceus and G. auratus | Study of detoxication enzyme activities for a fungicide, chlorothalonil | Metabolism of chlorothalonil |
| 7/1986 | [150] | - | Role of enzymatic processes in the metabolism of organic matter | Proteolysis and aerobic oxidation of organic material |
| 8/1992 | [151] | - | Study for distinction between enzymatic and non-enzymatic degradation pathways in marine ecosystems | Enzymatic degradation pathways for α-hexachlorocyclohexane |
| 9/1995 | [152] | Antioxidant enzymes in mussel, Mytilus galloprovincialis | Use of mussels as bioindicators in monitoring heavy metal pollution | Adaptation as a compensatory mechanism in chronically polluted organisms was found |
| 10/1995 | [153] | Dioxygenase | Dioxygenase pathway with subsequent conjugation and excretion | Metabolism of benzo[a]pyrene in a freshwater green alga, Selenastrum capricornutum |
| 11/2000 | [154] | Polychaete worms | Sulphide detoxification by polychaete worms Marenzelleria viridis (Verrill 1873) and Hediste diversicolor (O.F. Müller) | Detoxification end-product is thiosulphate |
| 12/2001 | [155] | - | - | Biodegradation of extracellular organic carbon by bacteria in sediments |
| 13/2004 | [156] | Antioxidant enzyme activities and lipid peroxidation in the gills of the hydrothermal vent mussel Bathymodiolus azoricus | Study of enzymatic defences (superoxide dismutase (SOD), catalase (CAT), total glutathione peroxidase (Total GPx) and selenium-dependent glutathione peroxidase (Se–GPx) and lipid peroxidation against metals | Assessment of physiological adaptation to continuous metal exposure in natural environment |
| 14/2004 | [157] | Marine glycosidases | Biostimulation of enzymatic activities (glycosidases) by oxygen supply | Enzymatic activity increased when oxygenation was increased and the supply of oxygen into the sediment enhanced enzymatic degradation rates |
| 15/2005 | [158] | Antioxidant enzymes in a model organism Daphnia magna | Study of age-related biochemical changes in aquatic organism | General evaluation of importance of oxidative stress in aging |
| 16/2006 | [159] | European eel (Anguilla anguilla) exposed to persistent organic pollutants | Detection of early warning responses to pollutant exposition | Metabolic responses including detoxification mechanisms (biotransformation, antioxidant process) in European eel (Anguilla anguilla) exposed to persistent organic pollutants |
| 17/2007 | [160] | Catalase, superoxide dismutase, glutathione peroxidase, glutathione reductase, glutathione S-transferases in wild populations of mussels (Mytilus galloprovincialis) | Study of biochemical response to to petrochemical environmental contamination | Environmental monitoring programmes to get data that could be used as a baseline reference during oil accidents |
| 18/2007 | [161] | Acetylcholinesterase, catalase, and glutathione-S-transferase (GST) of blue mussels (Mytilus edulis) | Study of specific reaction to exposure to nodularin | Acetylcholinesterase activity, catalase (CAT) activity and glutathione-S-transferase (GST) in blue mussels (Mytilus edulis) exposed to an extract made of natural cyanobacterial mixture containing toxic cyanobacterium Nodularia spumigena |
| 19/2007 | [162] | Induction of biotransformation enzymes in Sparus aurata | Relationship between specific molecular processes (induction of enzymes) and the behavioral performance of fish is of great interest in understanding the impact of PAHs at increasing levels of biological complexity | The study investigates biochemical response to phenantrene in Sparus aurata |
| 20/2012 | [163] | Tow deep-sea fish species, namely Alepocephalus rostratus and Lepidion lepidion and the decapod crustacean Aristeus antennatus | - | Study of hepatic biomarkers (ethoxyresorufin-O-deethylase, EROD, pentoxyresorufin-O-deethylase PROD, catalase CAT, carboxylesterase CbE, glutathione-S-transferase GST, total glutathione peroxidase GPX and glutathione reductase demonstrating seasonal variation despite constant temperature and salinity |
| 21/2013 | [164] | Platichthys flesus | Transcriptomic study | Assessment of hepatic transcriptional differences between fish exposed to mixture of brominated diphenyl ethers and controls |
| 22/2015 | [165] | Peptidase activities | A study of enzymes using labeled substrate in diverse regions of the ocean | Enzymatic capabilities differ in Pelagic–benthic environments, affecting the processing of marine organic matter |
| 23/2015 | [166] | Arginine kinase of the crustacean Exopalaemon carinicauda | Bioinformatic study. Insights on the role of Cr3+ on enyzme with respect to inhibition and aggregation with structural disruption | An investigation of the effect of Cr3+ on enzymes of seawater organisms providing information on the physiological role of metal pollution in marine environments |
| 24/2015 | [167] | Oxidoreductases and catalases in Bacillus safensis | Isolation and enzyme identification study | The organism is responsible for degradation of the petroleum aromatic fractions |
| 25/2016 | [168] | Dehydrogenase activity or peroxidase activity | A surface methodology study | Degradation of crude oil fitted linearly with increasing biomass and enzyme activities with growth |
© 2017 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trincone, A. Enzymatic Processes in Marine Biotechnology. Mar. Drugs 2017, 15, 93. https://doi.org/10.3390/md15040093
Trincone A. Enzymatic Processes in Marine Biotechnology. Marine Drugs. 2017; 15(4):93. https://doi.org/10.3390/md15040093
Chicago/Turabian StyleTrincone, Antonio. 2017. "Enzymatic Processes in Marine Biotechnology" Marine Drugs 15, no. 4: 93. https://doi.org/10.3390/md15040093
APA StyleTrincone, A. (2017). Enzymatic Processes in Marine Biotechnology. Marine Drugs, 15(4), 93. https://doi.org/10.3390/md15040093