
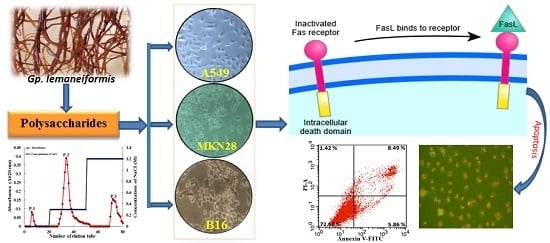
Characterization and Potential Antitumor Activity of Polysaccharide from Gracilariopsis lemaneiformis

Abstract
:
1. Introduction
2. Results and Discussion
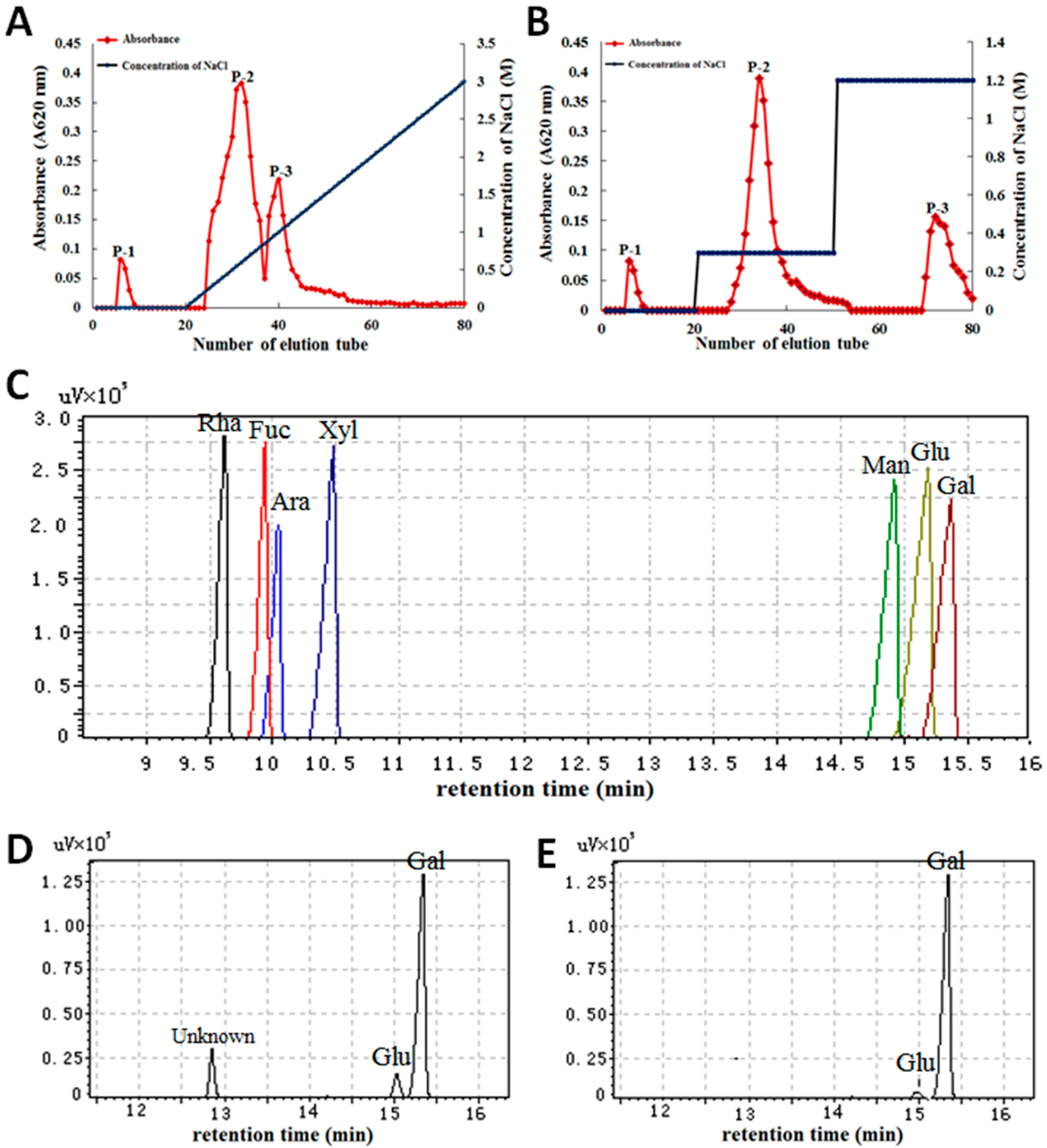
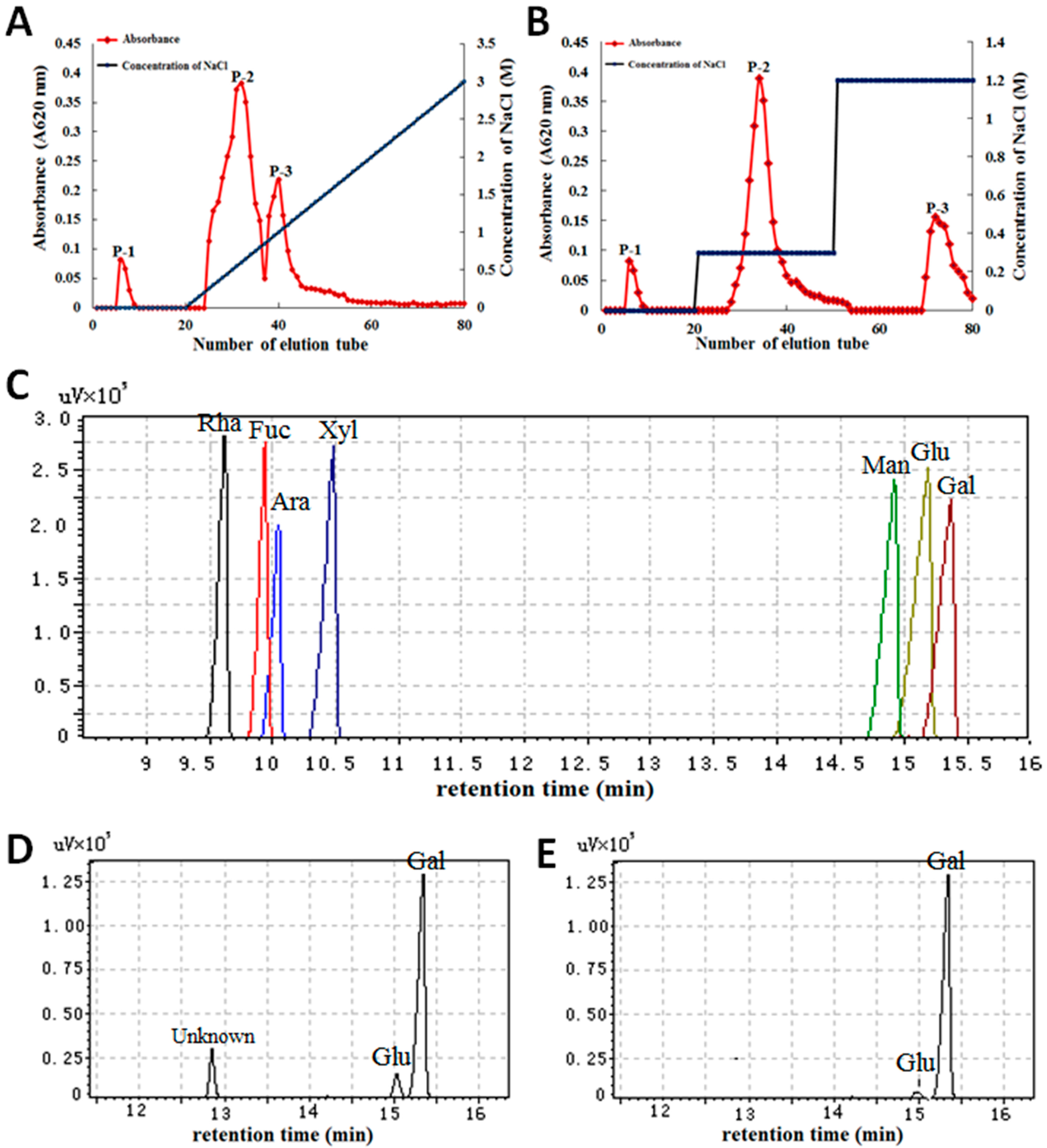
2.1. Characterization of Polysaccharides from Gp. lemaneiformis
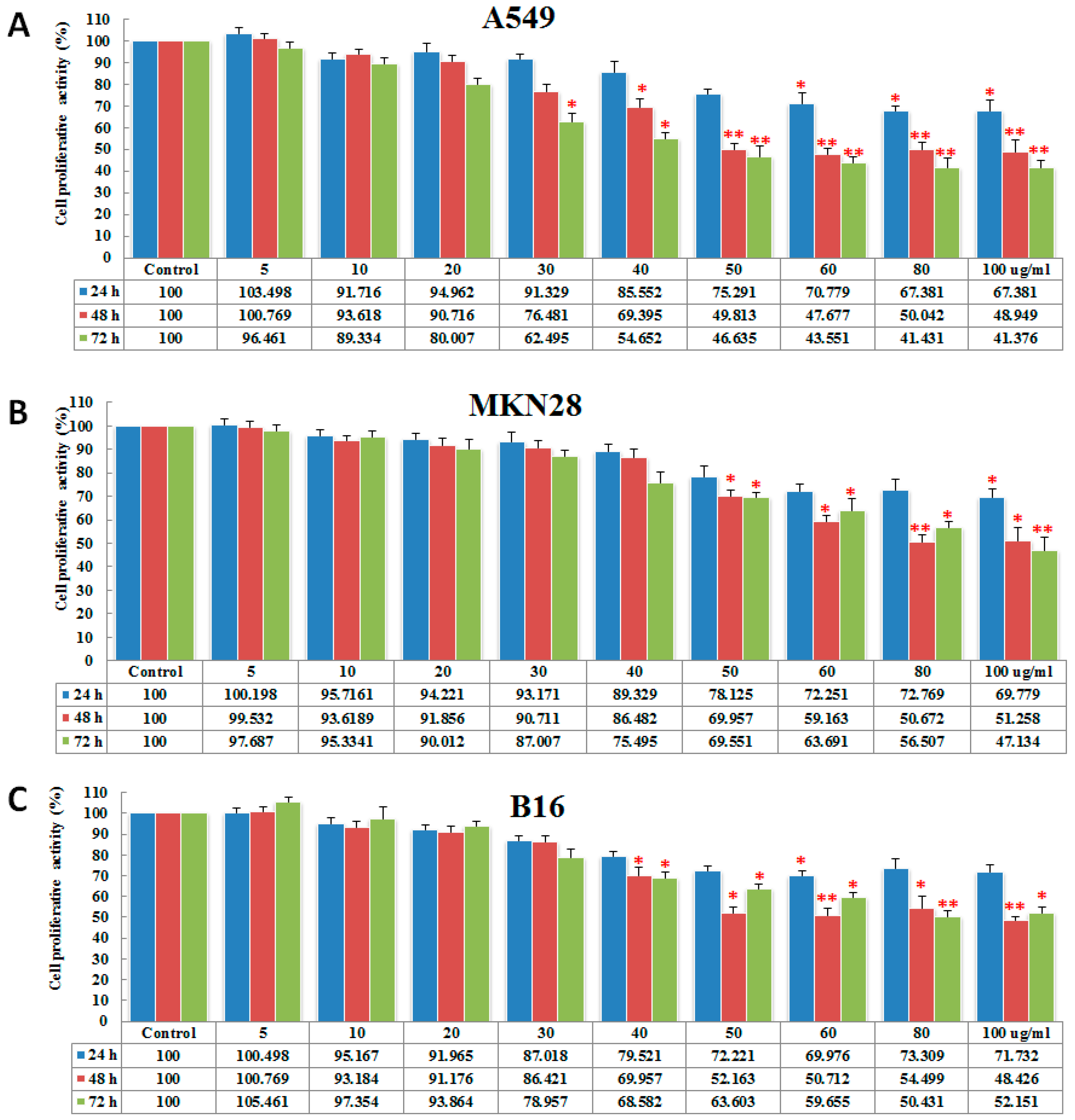
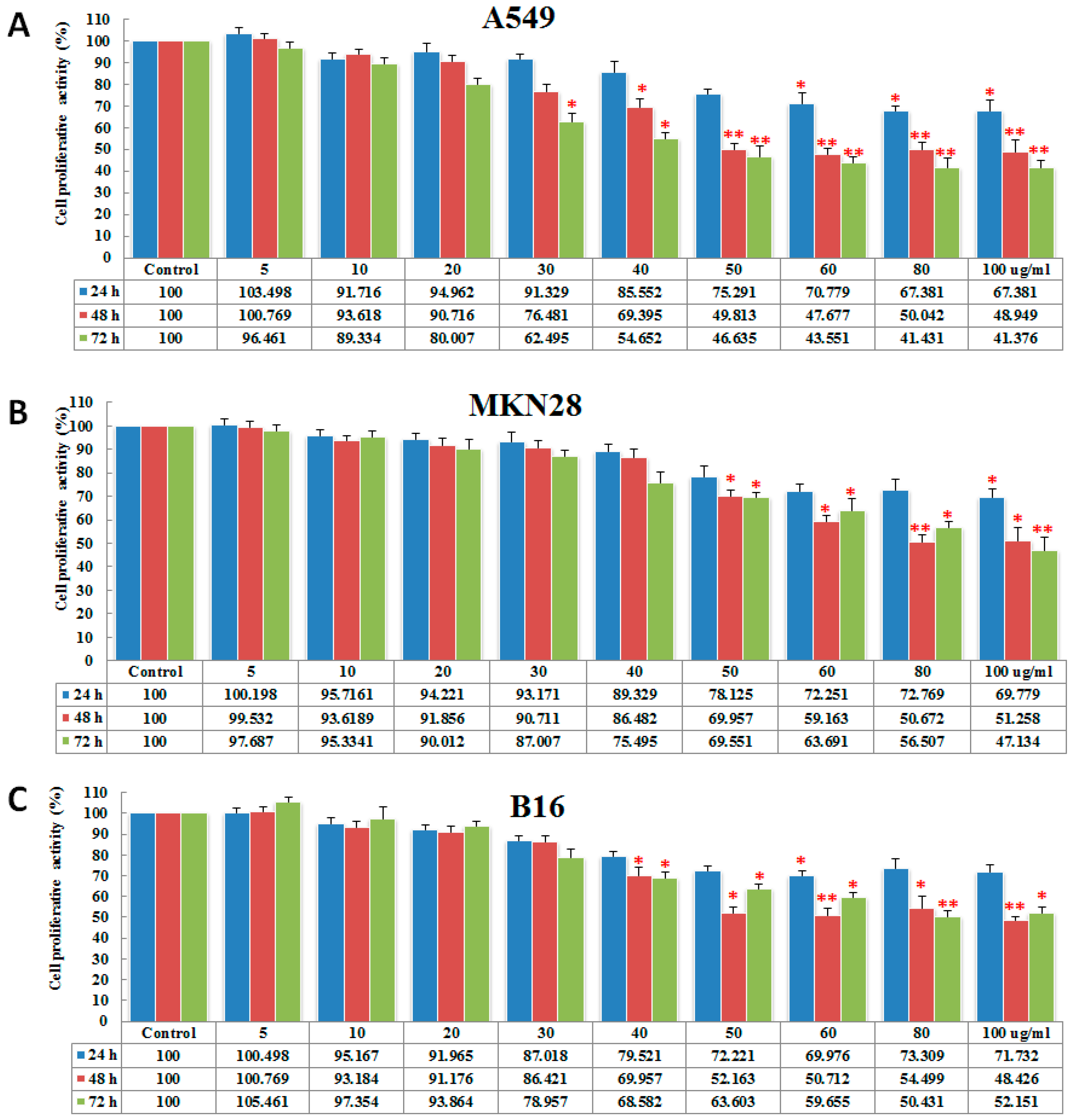
2.2. PGL Inhibits Cell Proliferation
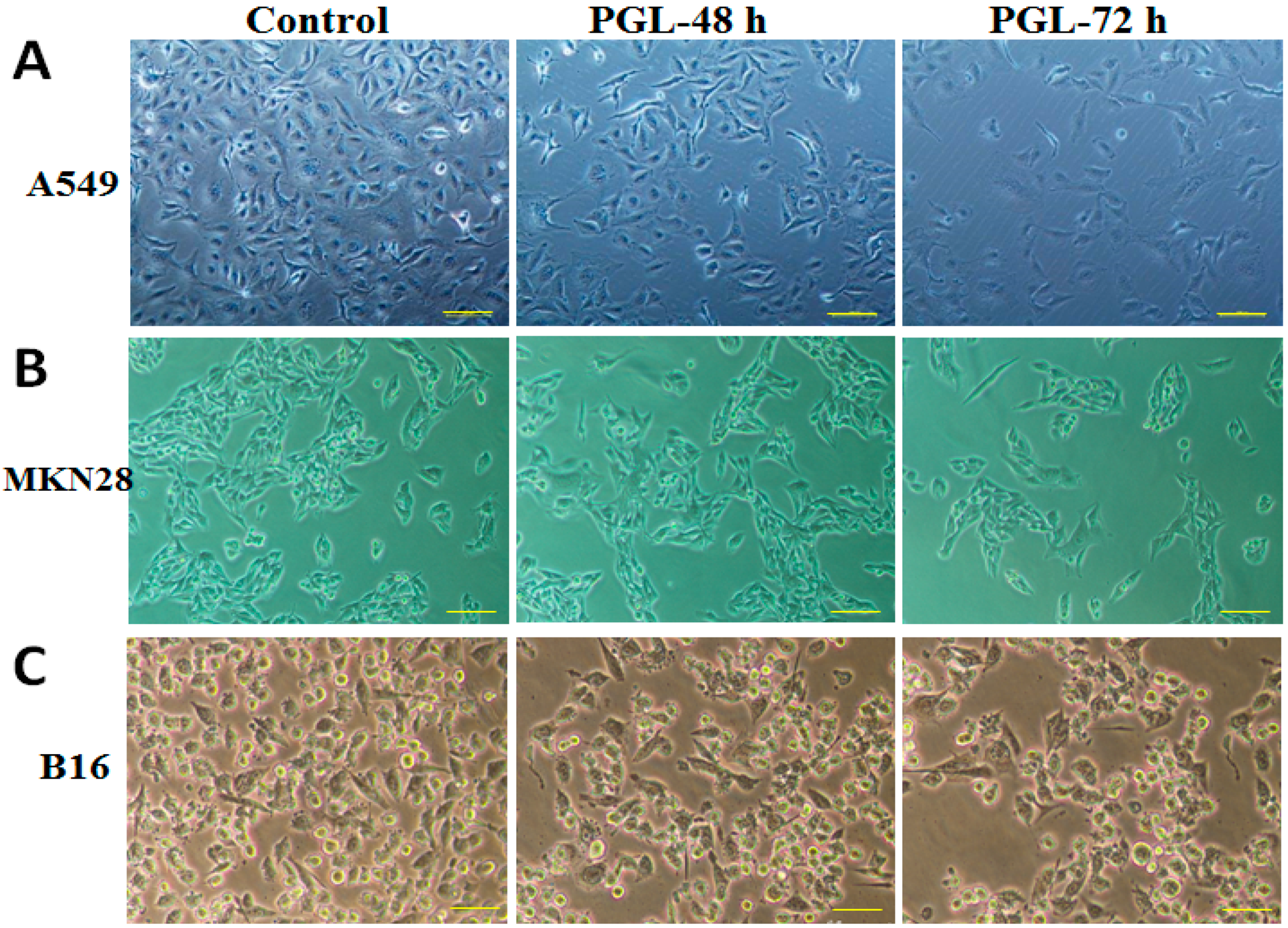
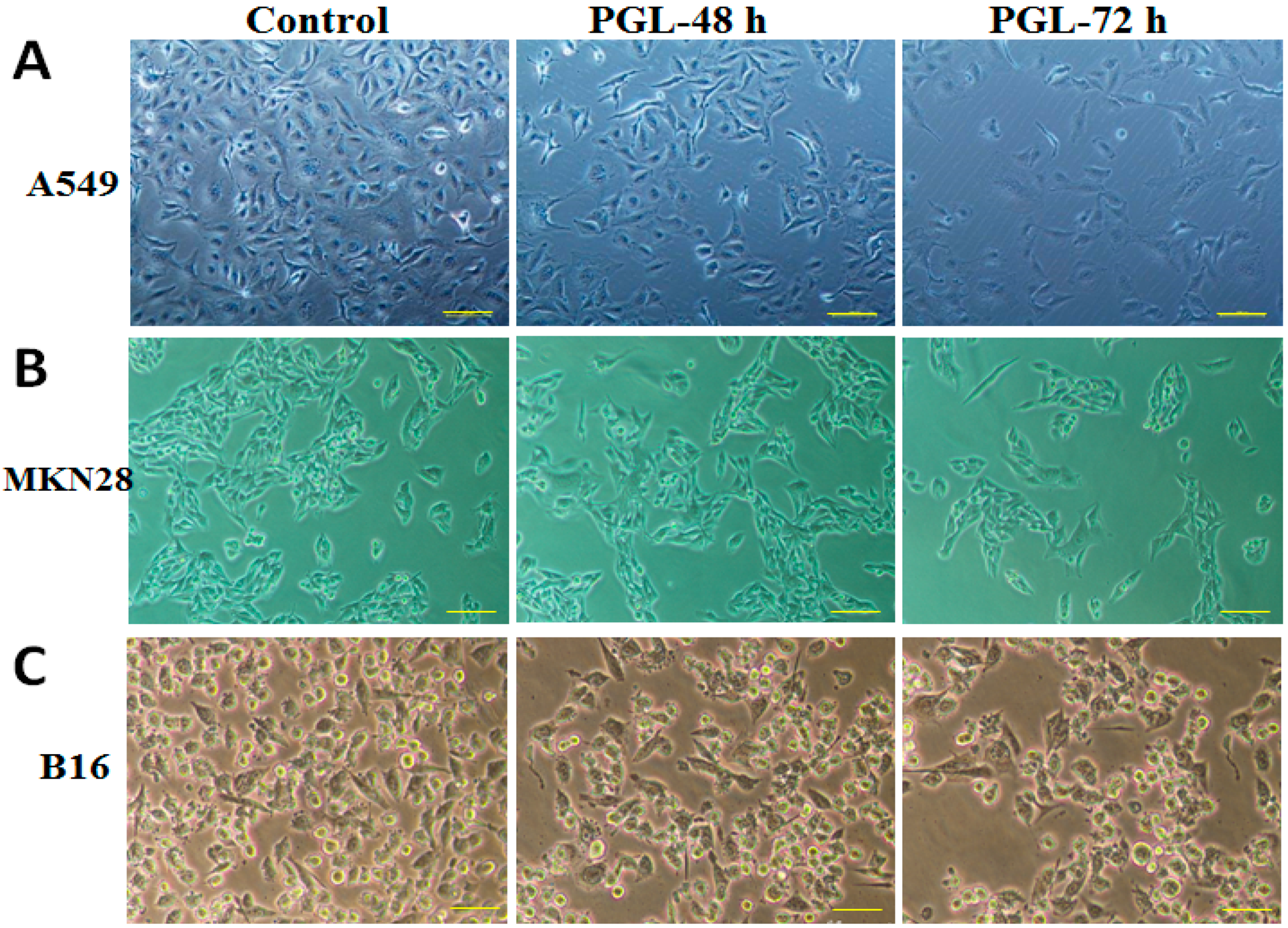
2.3. PGL Changes Cell Morphology and Reduces Cell Number
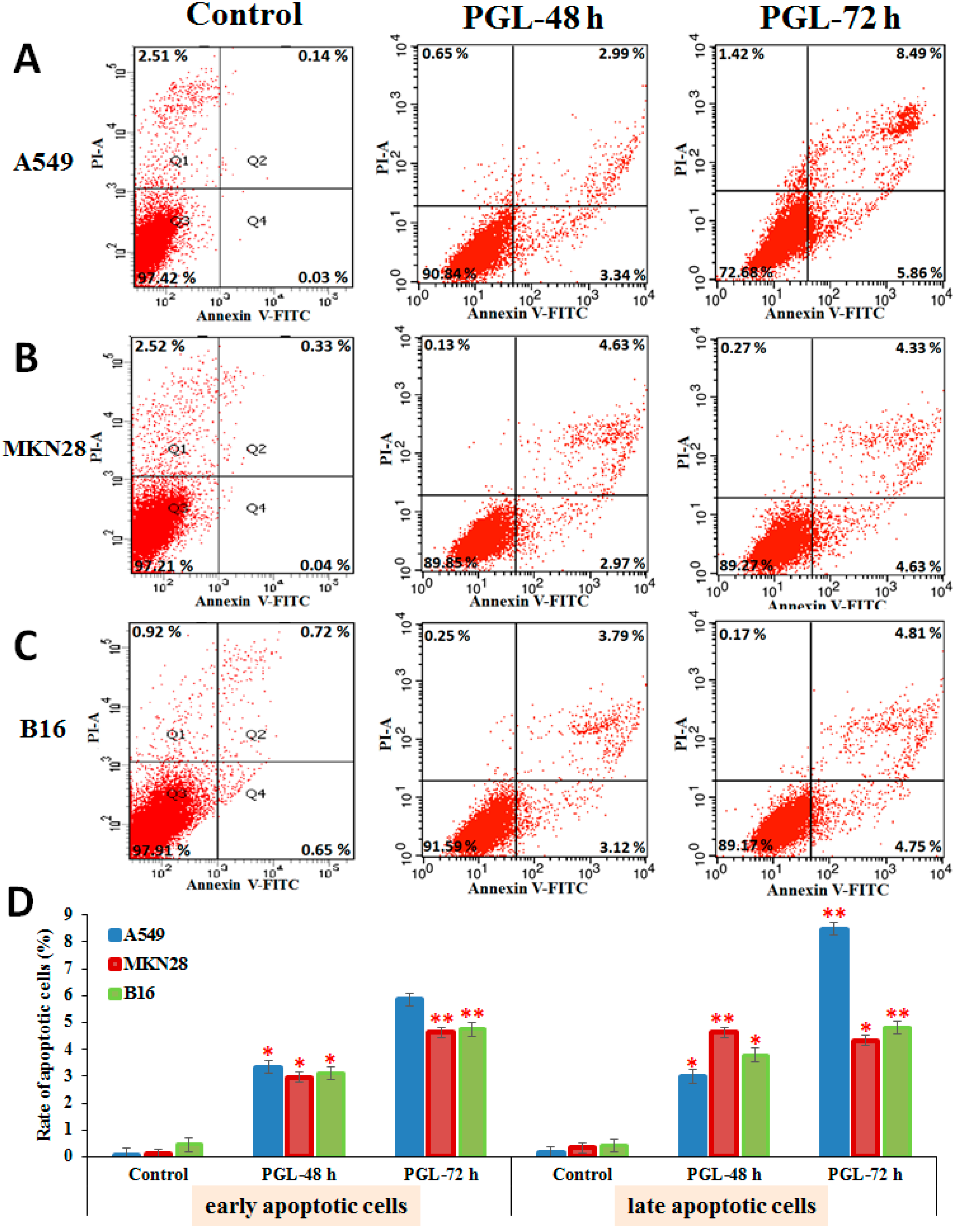
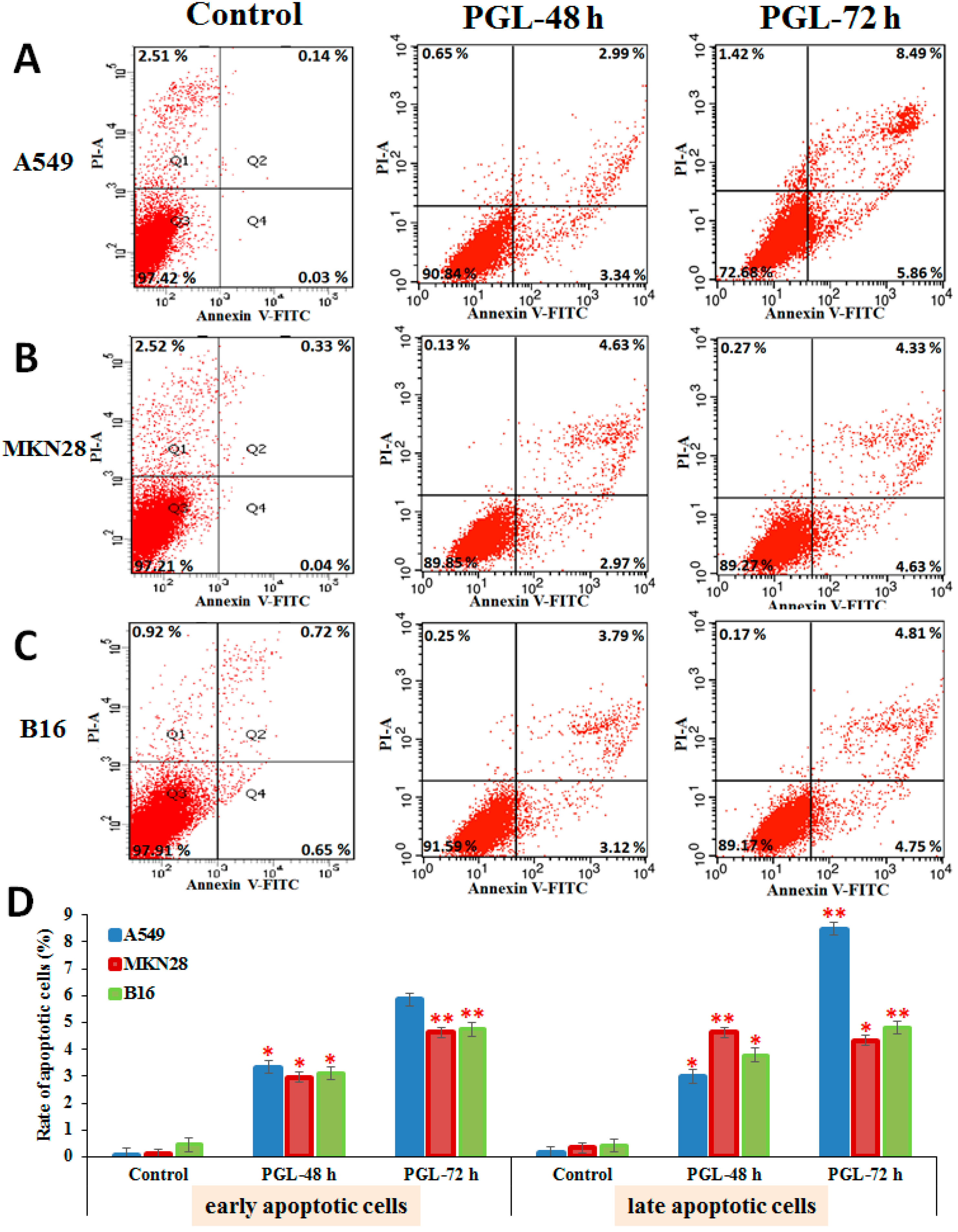
2.4. PGL Induces Apoptosis
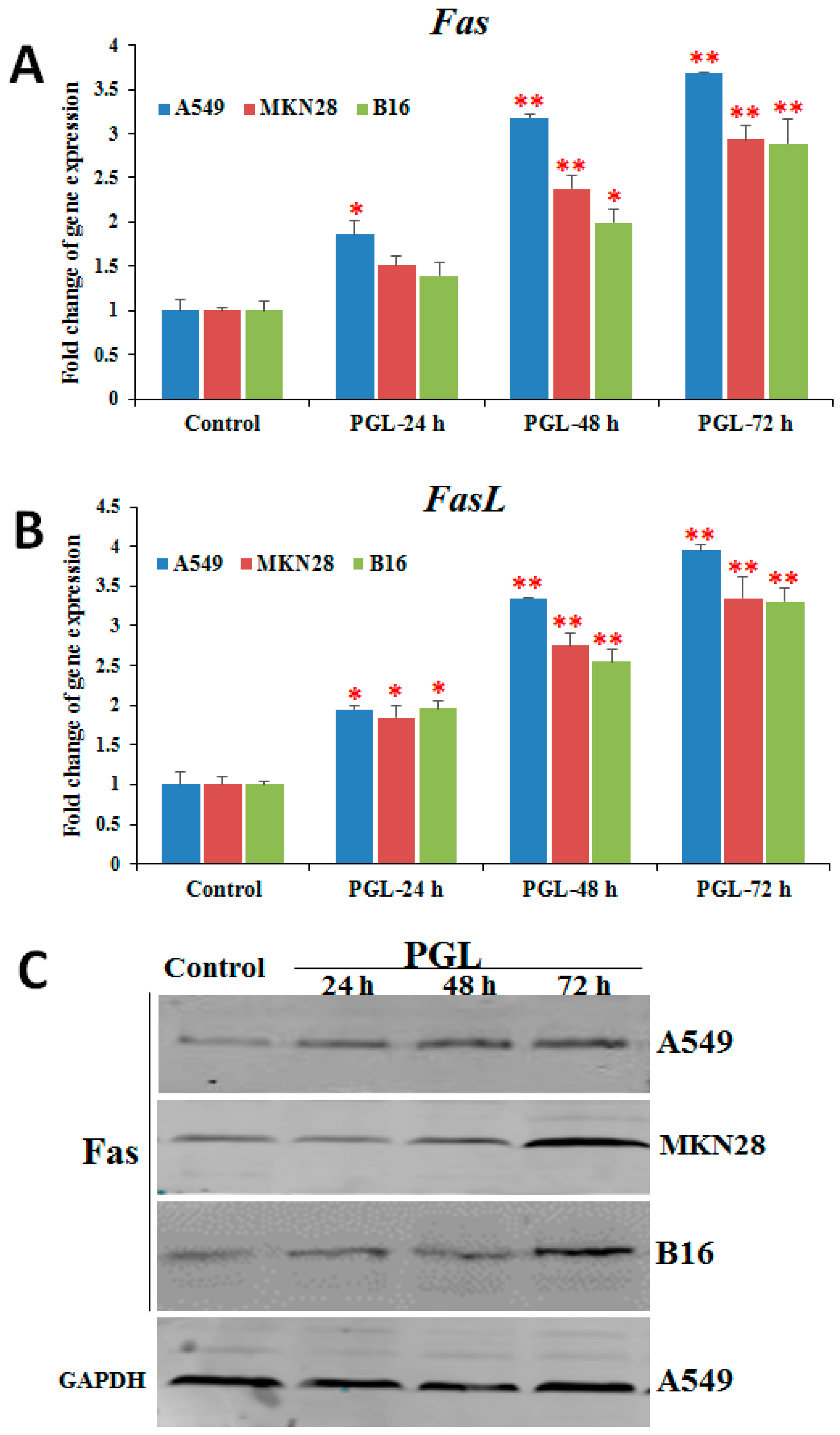
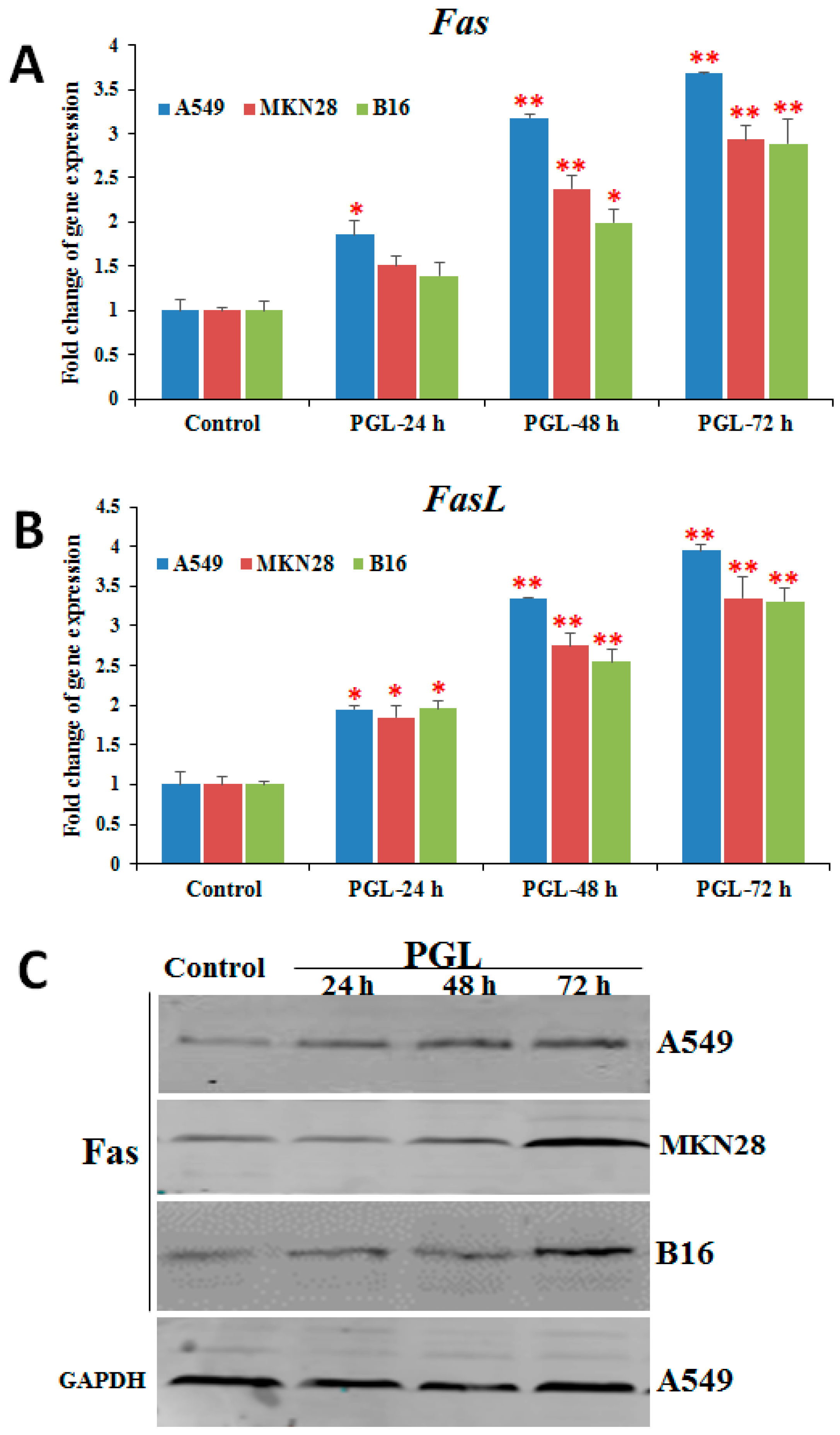
2.5. PGL Activates Fas/FasL Expression
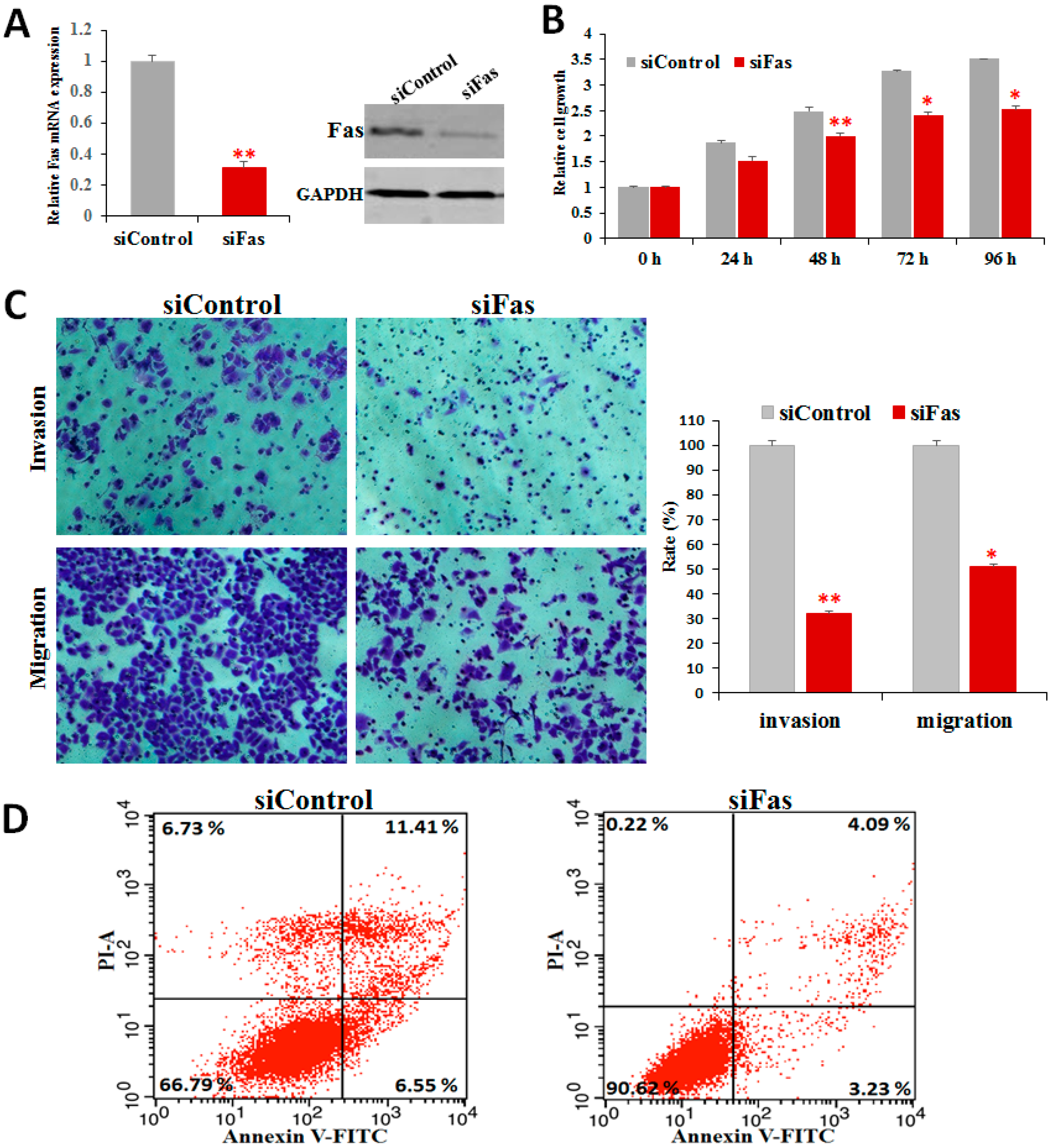
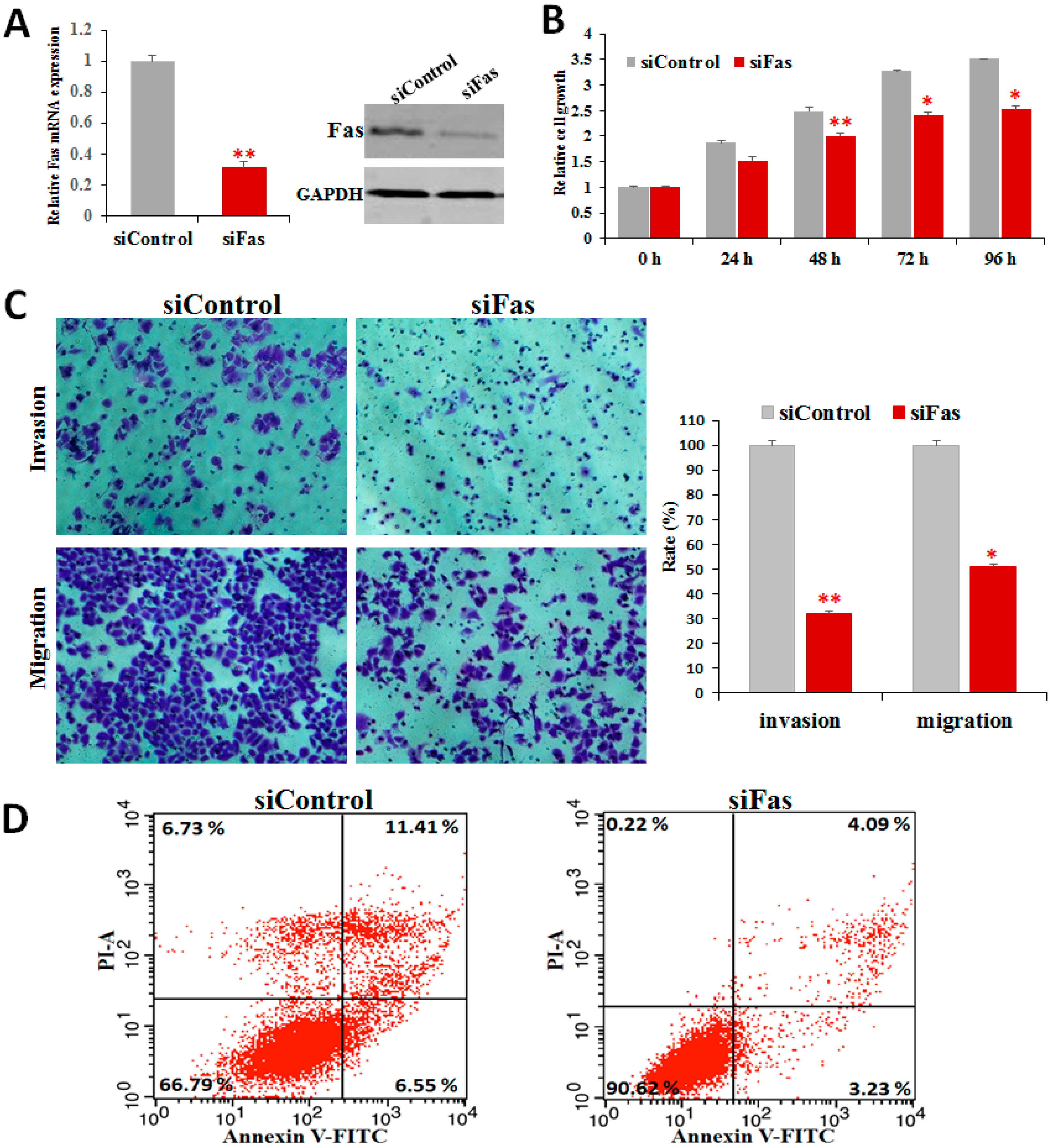
2.6. Fas Knockdown Inhibits Cell Proliferation, Invasion, and Migration and Reduces Apoptosis
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Characterization of Polysaccharides from Gp. lemaneiformis
4.2.1. Extraction of Crude Polysaccharides
4.2.2. Isolation and Purification of Polysaccharides
4.2.3. Determination of Molecular Weight
4.2.4. Chemical Properties of Monosaccharides
4.3. Assays for Antitumor Properties
4.3.1. Cell Culture and PGL Treatment
4.3.2. Cell Viability Analysis
4.3.3. Observation of Cell Morphology Changes
4.3.4. Apoptosis Analysis
4.3.5. Fas/FasL Expression Analysis
4.3.6. Western Blot Analysis
4.3.7. siRNA and Transfection
4.3.8. Cell Invasion and Migration Assays
4.4. Statistical Analyses
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Jesus Raposo, M.F.; Bernardo de Morais, A.M.; Santos Costa de Morais, R.M. Marine polysaccharides from algae with potential biomedical applications. Mar. Drugs 2015, 13, 2967–3028. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-B.; Hwang, C.J.; Song, S.-Y.; Jung, Y.Y.; Yun, H.-M.; Sok, C.H.; Sung, H.C.; Yi, J.-M.; Park, D.H.; Ham, Y.W. Anti-cancer effect of tectochrysin in nsclc cells through overexpression of death receptor and inactivation of stat3. Cancer Lett. 2014, 353, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Zorofchian Moghadamtousi, S.; Karimian, H.; Khanabdali, R.; Razavi, M.; Firoozinia, M.; Zandi, K.; Abdul Kadir, H. Anticancer and antitumor potential of fucoidan and fucoxanthin, two main metabolites isolated from brown algae. Sci. World J. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Usoltseva, R.V.; Anastyuk, S.D.; Shevchenko, N.M.; Zvyagintseva, T.N.; Ermakova, S.P. The comparison of structure and anticancer activity in vitro of polysaccharides from brown algae Alaria marginata and A. angusta. Carbohydr. Polym. 2016, 153, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.J.; Costa, R.R.; Mano, J.F. Marine origin polysaccharides in drug delivery systems. Mar. Drugs 2016, 14, 34. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-K.; Thomas, N.V.; Li, X. Anticancer compounds from marine macroalgae and their application as medicinal foods. In Advances in Food and Nutrition Research, Vol 64: Marine Medicinal Foods: Implications and Applications, Macro and Microalgae; Kim, S.K., Ed.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2011; Volume 64, pp. 213–224. [Google Scholar]
- Wang, Z.H.; Wang, X.N.; Ke, C.L. Bioaccumulation of trace metals by the live macroalga Gracilaria lemaneiformis. J. Appl. Phycol. 2014, 26, 1889–1897. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, X.; Qian, H.; Chi, S.; Liu, C.; Liu, T. Complete sequences of the mitochondrial DNA of the wild Gracilariopsis lemaneiformis and two mutagenic cultivated breeds (Gracilariaceae, Rhodophyta). PLoS ONE 2012, 7, e40241. [Google Scholar] [CrossRef] [PubMed]
- Khanavi, M.; Nabavi, M.; Sadati, N.; Ardekani, M.S.; Sohrabipour, J.; Nabavi, S.M.B.; Ghaeli, P.; Ostad, S.N. Cytotoxic activity of some marine brown algae against cancer cell lines. Biol. Res. 2010, 43, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wang, Z.; Wang, Y.; Chu, J.; Zhuang, Y.; Zhang, S. Structural elucidation and antioxidant activity of a polysaccharide from mycelia fermentation of Hirsutella sinensis isolated from Ophiocordyceps sinensis. J. Bioprocess. Biotech. 2014, 4, 183. [Google Scholar]
- Yang, X.; Liu, M.; Qi, B.; Li, L.; Deng, J.; Hu, X.; Wu, Y.; Hao, S. Extraction, purification and partial characterizations of polysaccharides from Gracilaria lemaneiformis. Adv. Mater. Res. 2014, 881–883, 776–780. [Google Scholar]
- Kang, Y.; Lu, J.; Xu, X.; Sun, X.; Xu, N.; Zhao, X. Effect of cell proliferation activity and morphology in lung cancer cells by polysaccharide of Gracilaria lemaneiformis. J. Fish. China 2015, 39, 1405–1411. [Google Scholar]
- Kang, Y.; Li, H.; Wu, J.; Xu, X.; Sun, X.; Zhao, X.; Xu, N. Transcriptome profiling reveals the antitumor mechanism of polysaccharide from marine algae Gracilariopsis lemaneiformis. PLoS ONE 2016, 11, e0158279. [Google Scholar] [CrossRef] [PubMed]
- Villa-Morales, M.; Fernández-Piqueras, J. Targeting the fas/fasl signaling pathway in cancer therapy. Expert Opin. Ther. Targets 2012, 16, 85–101. [Google Scholar] [CrossRef] [PubMed]
- Svandova, E.B.; Vesela, B.; Lesot, H.; Poliard, A.; Matalova, E. Expression of fas, fasl, caspase-8 and other factors of the extrinsic apoptotic pathway during the onset of interdigital tissue elimination. Histochem. Cell Biol. 2016. [Google Scholar] [CrossRef]
- Yu, J.; Wang, X.; Chen, M.; Zhang, Y.; Long, Z. Analysis on nutritional components and polysaccharide composition of Gracilaria lemaneiformis from chaoshan coast. Food Sci. 2006, 27, 93–97. [Google Scholar]
- Zong, A.; Cao, H.; Wang, F. Anticancer polysaccharides from natural resources: A review of recent research. Carbohydr. Polym. 2012, 90, 1395–1410. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, S.; Ermakova, S.; Zvyagintseva, T.; Stonik, V. Anticancer and cancer preventive properties of marine polysaccharides: Some results and prospects. Mar. Drugs 2013, 11, 4876–4901. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.; Tanabe, K.; Uchida, Y.; Emi, M.; Inoue, H.; Toge, T. Current status of the molecular mechanisms of anticancer drug-induced apoptosis - the contribution of molecular-level analysis to cancer chemotherapy. Cancer Chemother. Pharmacol. 2002, 50, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, Z.; Shukla, Y. Death receptors: Targets for cancer therapy. Exp. Cell Res. 2010, 316, 887–899. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Liu, R.; Si, C.; Zhou, F.; Wang, Y.; Ding, L.; Jing, C.; Liu, A.; Zhang, Y. Structural analysis and anti-tumor activity comparison of polysaccharides from Astragalus. Carbohydr. Polym. 2011, 85, 895–902. [Google Scholar] [CrossRef]
- Hao, L.; Sheng, Z.; Lu, J.; Tao, R.; Jia, S. Characterization and antioxidant activities of extracellular and intracellular polysaccharides from Fomitopsis pinicola. Carbohydr. Polym. 2016, 141, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Yang, W.; Lou, Q.; Wang, Y.; Yao, Z. The impact of yield of polysaccharides and antioxygenic property extracted with technical conditions from Gracilaria lamaneiformis. Food Sci. Technol. 2008, 50, 162–164. [Google Scholar]
- Liu, G. Chemical compositions, a-glucosidase and a-amylase inhibitory activities of crude polysaccharides from the endodermis of shaddock (Citrus maxima). Arch. Biol. Sci. 2012, 64, 71–76. [Google Scholar] [CrossRef]
- Meng, L.; Sun, S.; Li, R.; Shen, Z.; Wang, P.; Jiang, X. Antioxidant activity of polysaccharides produced by Hirsutella sp. and relation with their chemical characteristics. Carbohydr. Polym. 2015, 117, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Yang, L.; Chen, M.; Yu, J.; Zhang, S.; Ju, Y. The hypoglycemic effect of a polysaccharide from Gracilaria lemaneiformis and its degradation products in diabetic mice. Food Funct. 2015, 6, 2542–2549. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Zhang, X.; Kang, Y.; Wu, J.; Chen, J.; Li, H.; Guo, Y.; Liu, B.; Shao, Z.; Zhao, X. Genome-Wide transcriptional profiling analysis reveals annexin a6 as a novel ezh2 target gene involving gastric cellular proliferation. Mol. Biosyst. 2015, 11, 1980–1986. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Tan, S.; Kang, Y.; Wu, J.; Ooi, H.; Chen, J.; Shen, T.; Qi, Y.; Zhang, X.; Guo, Y. Genome-Wide profiling of polyadenylation sites reveals a link between selective polyadenylation and cancer metastasis. Hum. Mol. Genet. 2015, 24, 3410–3417. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Crude PGL | P-1 | P-2 | P-3 |
|---|---|---|---|---|
| 3,6-anhydro-l-galactose (%) | 41.20 ± 0.09 | 58.30 ± 0.04 | 47.60 ± 0.06 | 38.61 ± 0.05 |
| d-galactose (%) | 57.38 ± 0.04 | 29.53 ± 0.03 | 51.27 ± 0.05 | 59.76 ± 0.02 |
| Sulfate ester (%) | 9.24 ± 0.01 | 8.15 ± 0.02 | 9.16 ± 0.03 | 8.14 ± 0.02 |
| Protein contents (%) | 19.49 ± 0.08 | 0.35 ± 0.03 | 0.26 ± 0.02 | 0.42 ± 0.03 |
| Molecular weight (kDa) | 123.06 | 14.29 | 64.78 | 57.02 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.; Wang, Z.-J.; Xie, D.; Sun, X.; Yang, W.; Zhao, X.; Xu, N. Characterization and Potential Antitumor Activity of Polysaccharide from Gracilariopsis lemaneiformis. Mar. Drugs 2017, 15, 100. https://doi.org/10.3390/md15040100
Kang Y, Wang Z-J, Xie D, Sun X, Yang W, Zhao X, Xu N. Characterization and Potential Antitumor Activity of Polysaccharide from Gracilariopsis lemaneiformis. Marine Drugs. 2017; 15(4):100. https://doi.org/10.3390/md15040100
Chicago/Turabian StyleKang, Yani, Zhi-Jiang Wang, Dongsheng Xie, Xue Sun, Wenge Yang, Xiaodong Zhao, and Nianjun Xu. 2017. "Characterization and Potential Antitumor Activity of Polysaccharide from Gracilariopsis lemaneiformis" Marine Drugs 15, no. 4: 100. https://doi.org/10.3390/md15040100
APA StyleKang, Y., Wang, Z.-J., Xie, D., Sun, X., Yang, W., Zhao, X., & Xu, N. (2017). Characterization and Potential Antitumor Activity of Polysaccharide from Gracilariopsis lemaneiformis. Marine Drugs, 15(4), 100. https://doi.org/10.3390/md15040100