
Biotransformation and Detoxification of Xylidine Orange Dye Using Immobilized Cells of Marine-Derived Lysinibacillus sphaericus D3

Abstract
:1. Introduction
2. Results and Discussion
2.1. Xylidine Orange Dye and Decolorizing Bacteria
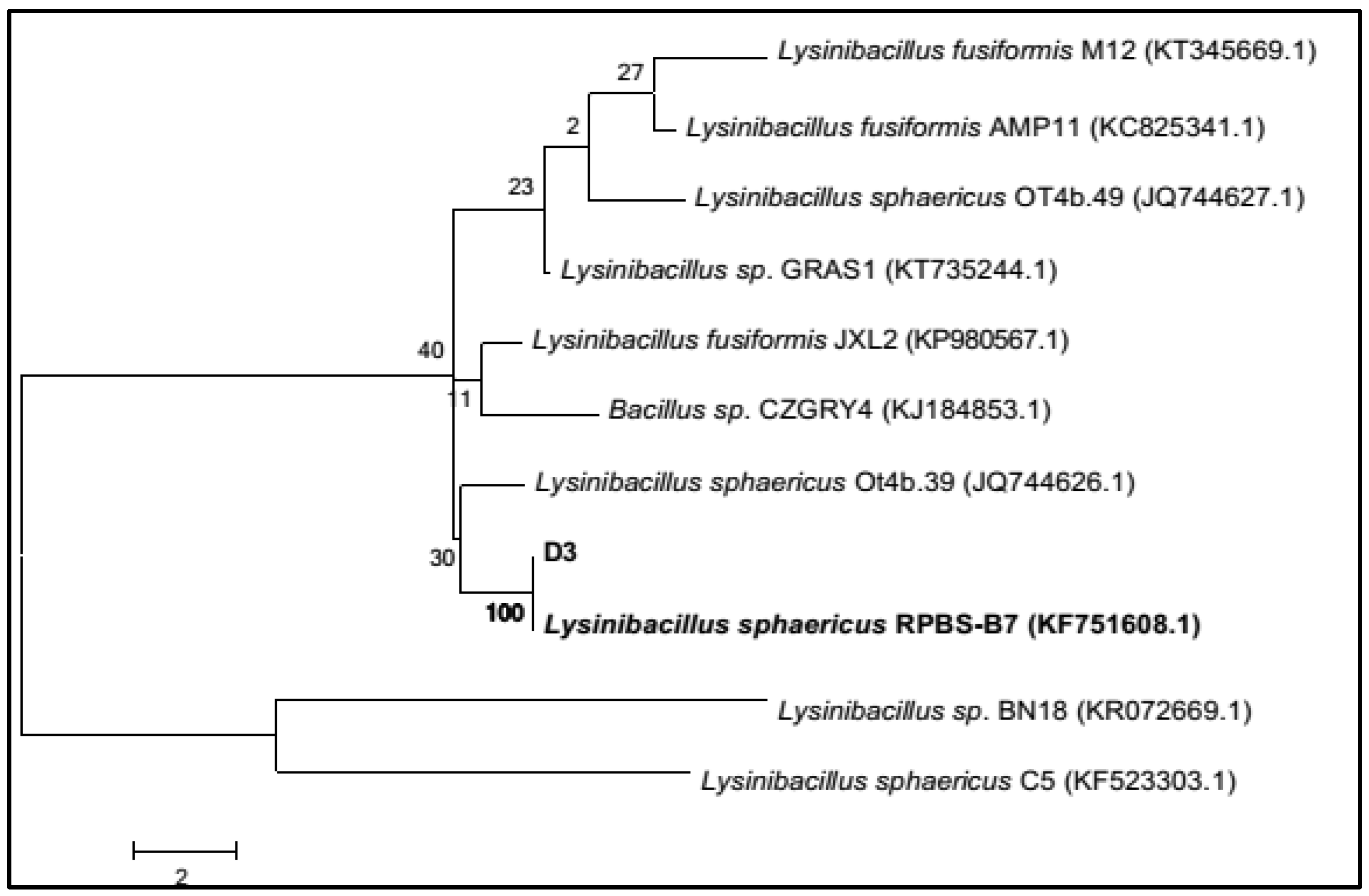
2.2. Identification of Dye Degrading Strain D3 by Polymerase Chain Reaction (PCR) Analysis
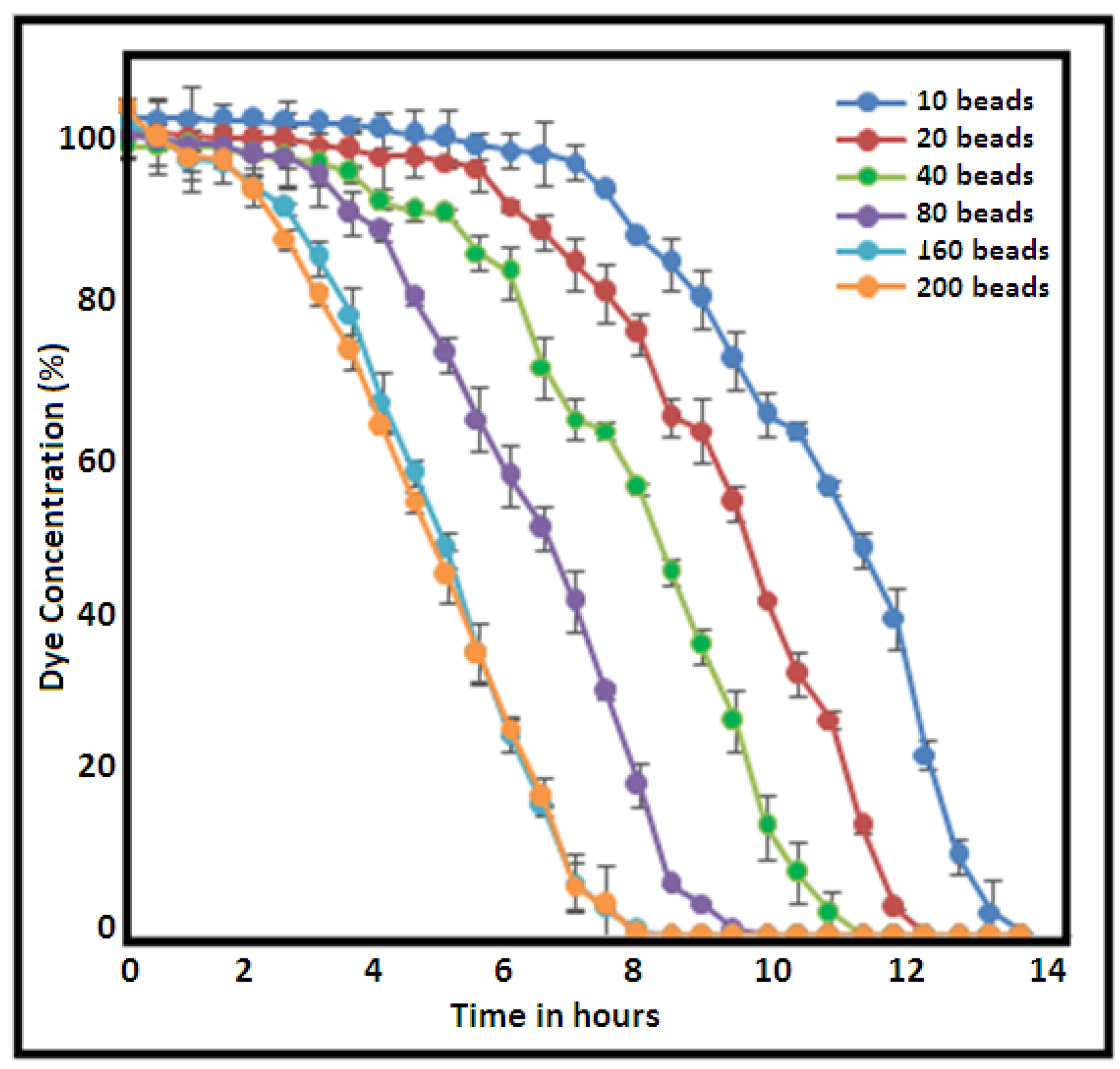
2.3. Physico-Chemical Parameters for Xylidine Orange Decolorization
2.4. Toxicological Evaluation of the Dye before and after Treatment
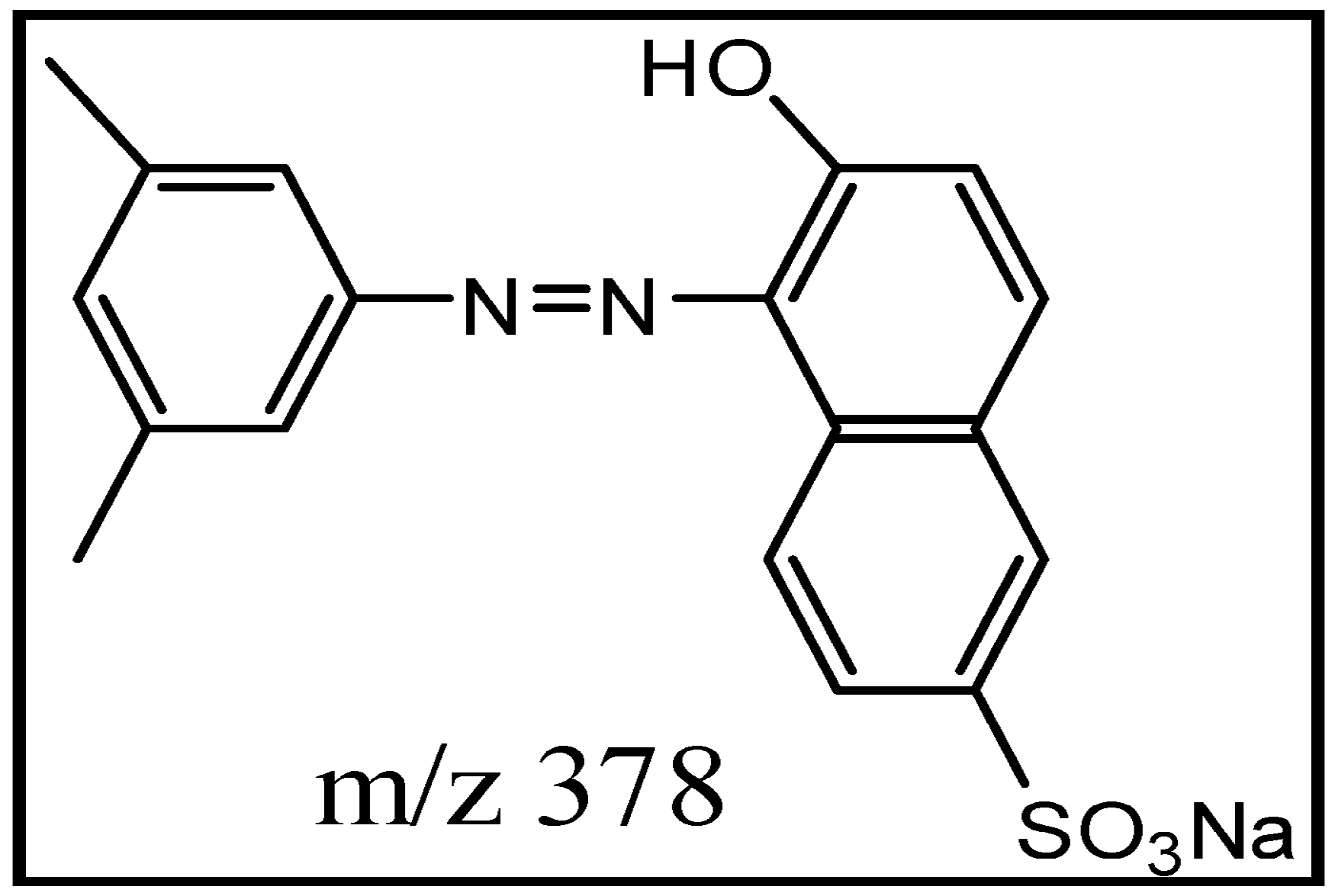
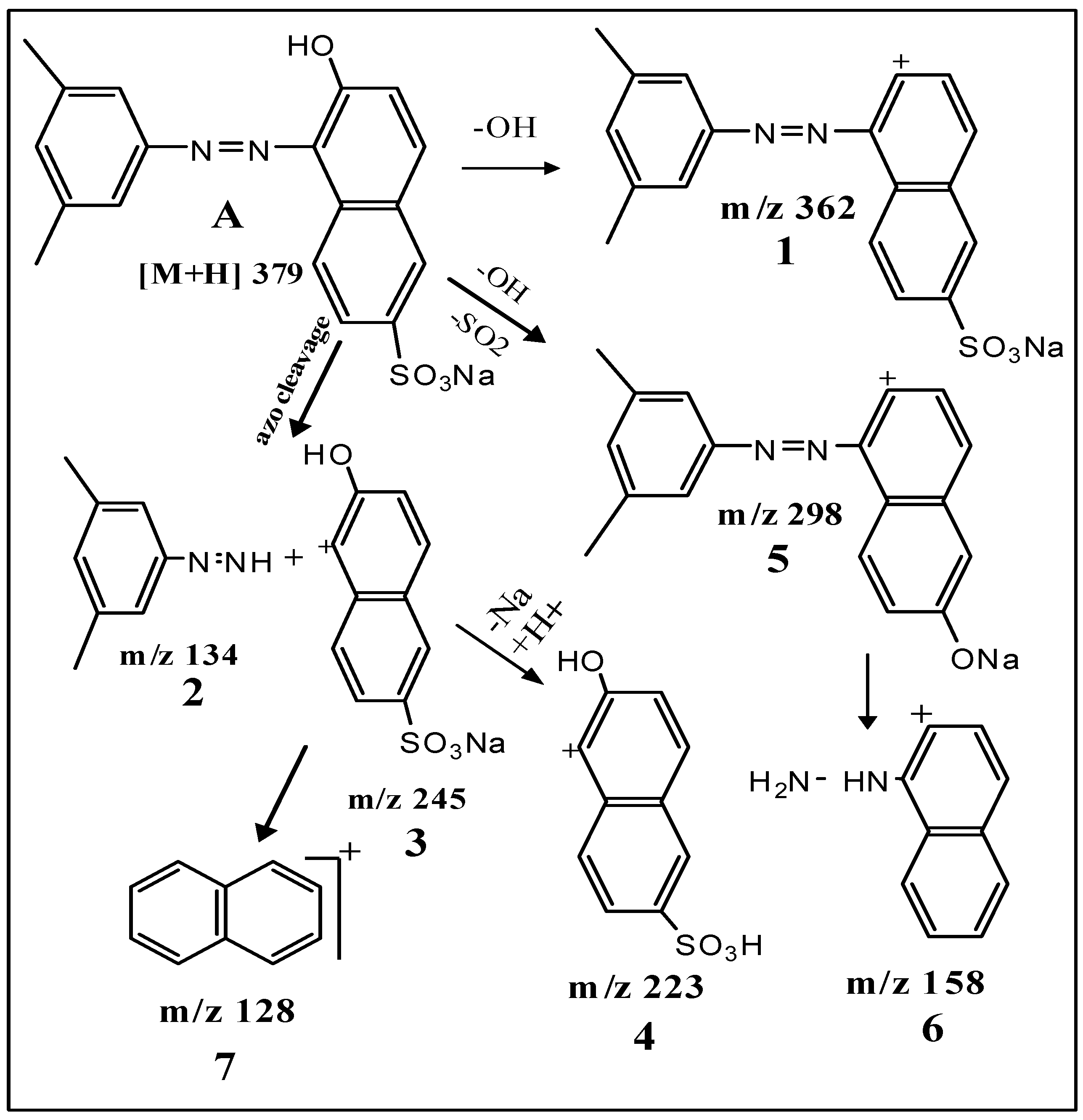
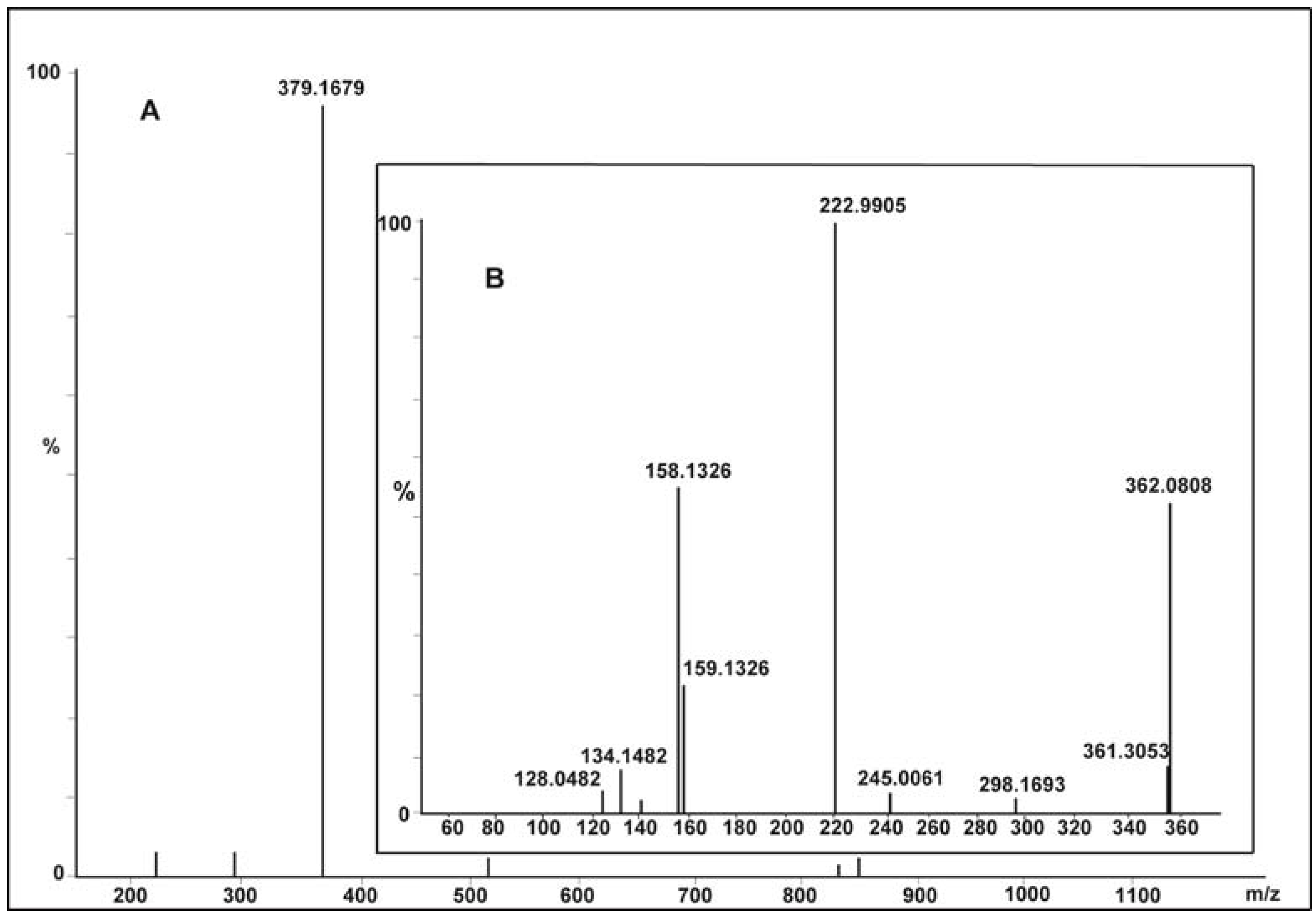
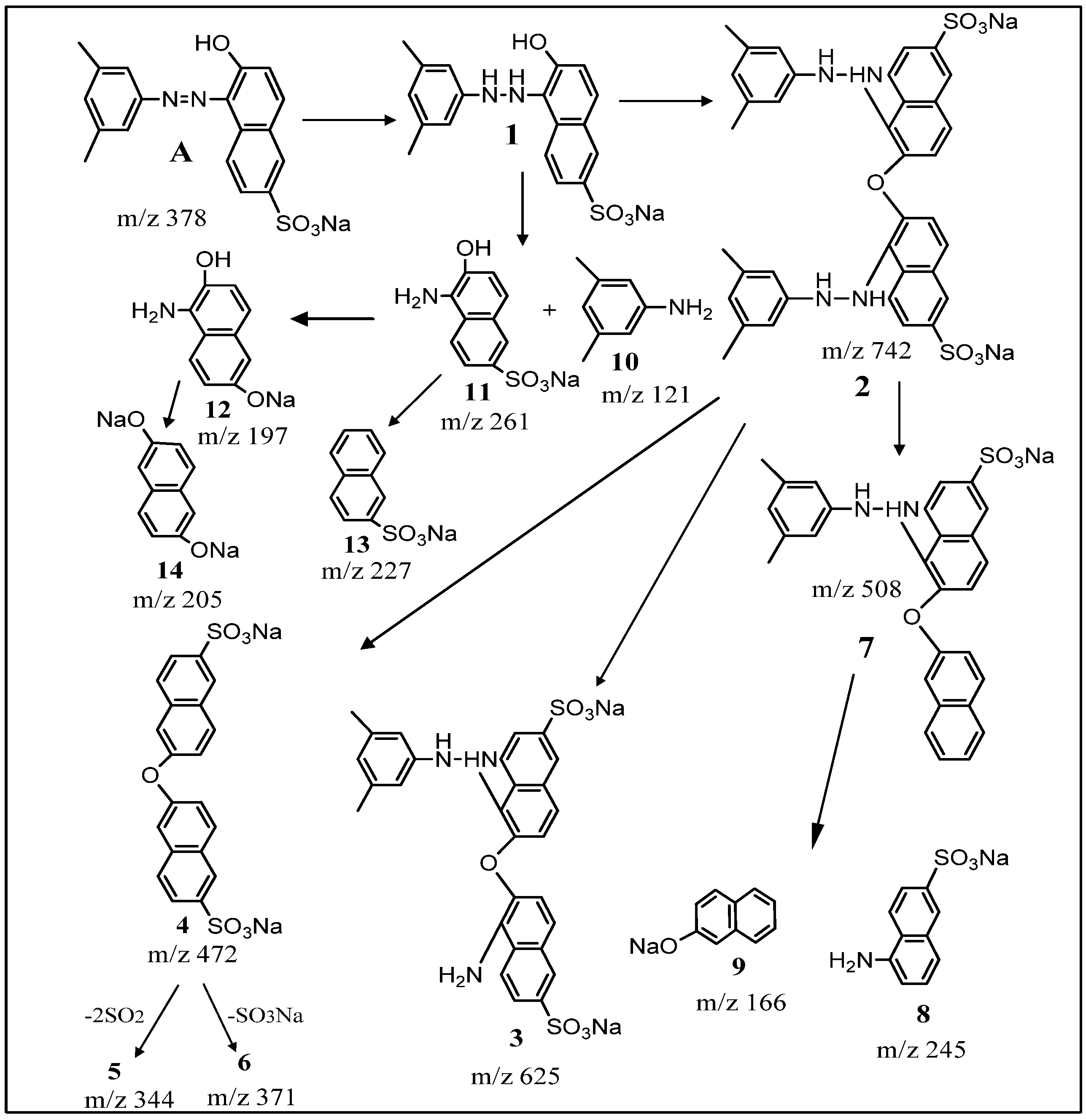
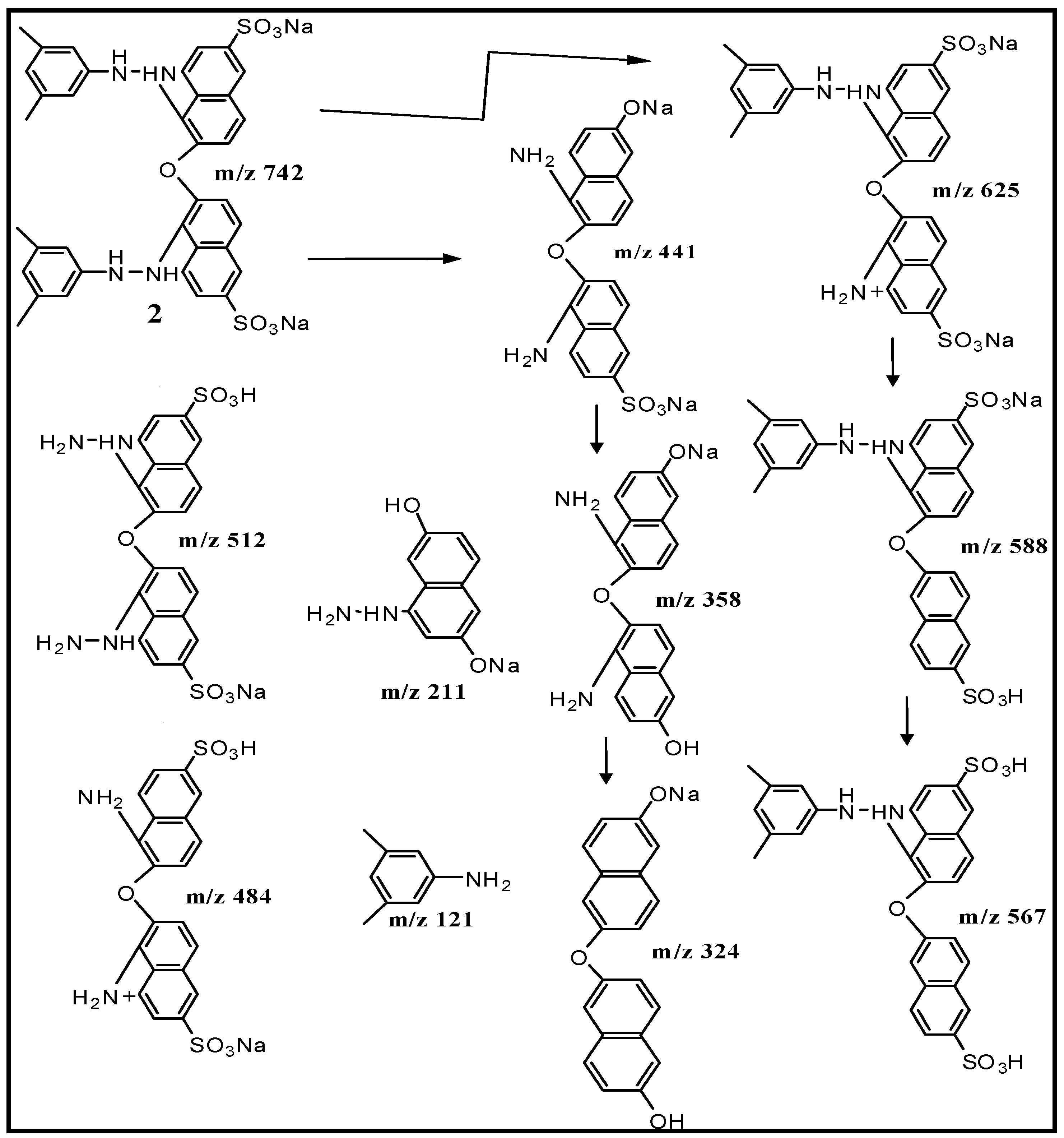
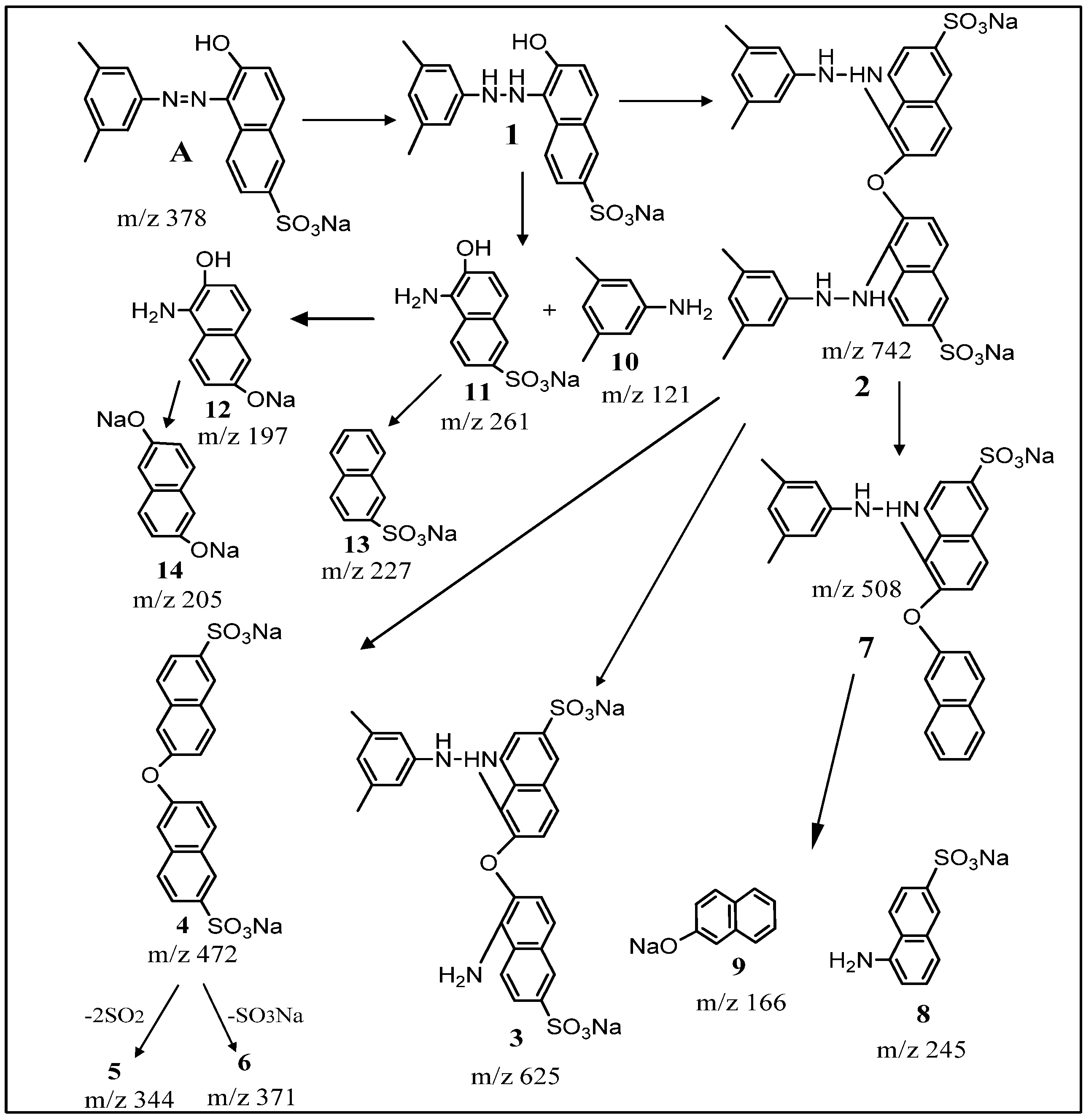
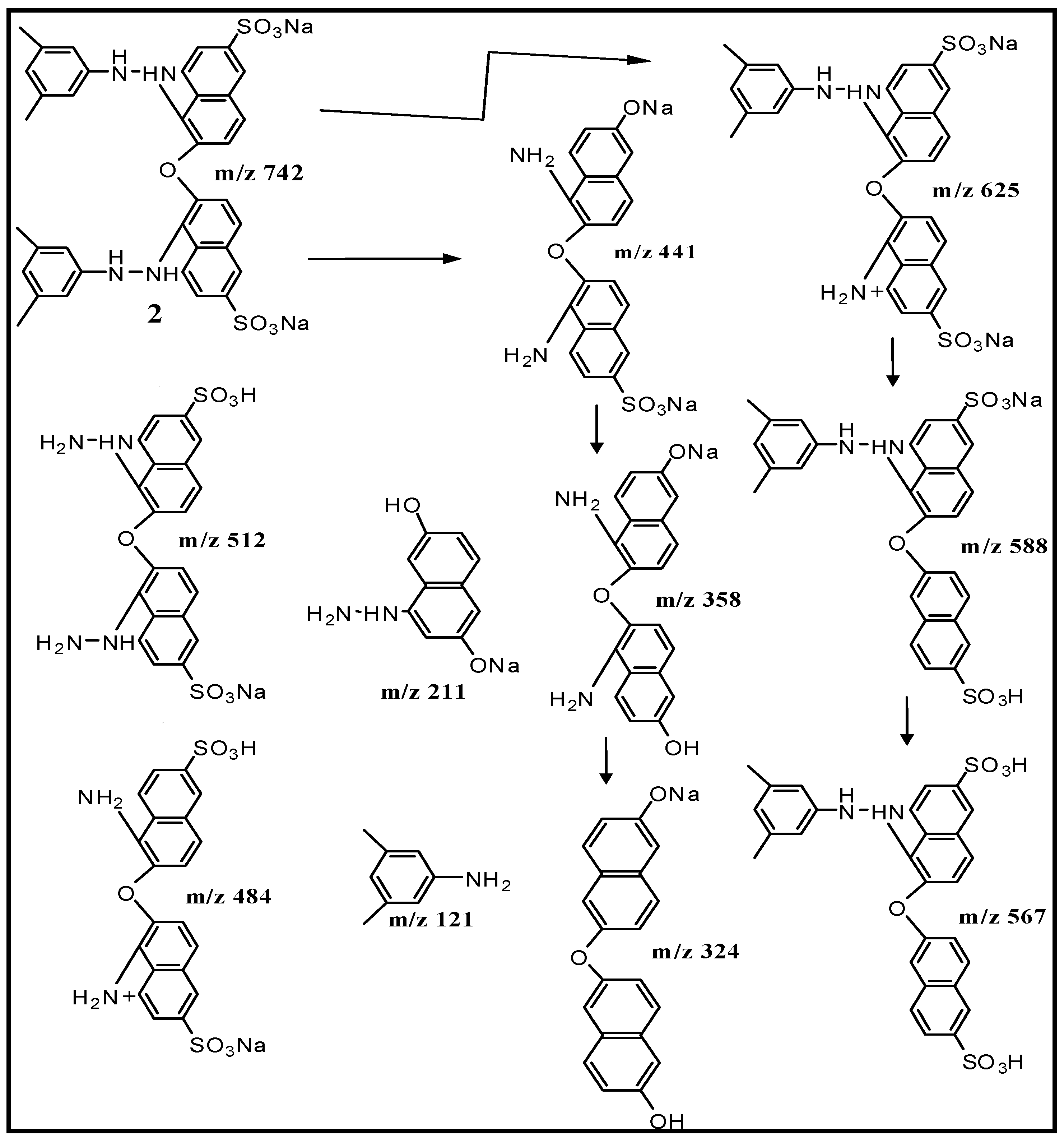
2.5. Spectral Analysis and Identification of Biotransformed Products
3. Materials and Methods
3.1. Dye and Reagents Used in the Study
3.2. Analytical Techniques/Instrumentation
3.3. Isolation of Sponge-Associated Bacteria Capable of Dye Decolorization
3.4. Immobilization of Marine-Derived Bacterium
3.5. 16S Ribosomal Ribonucleic Acid (rRNA) Gene Sequencing of the Dye Degrading Strain D3
3.6. Optimization of Culture Media for Dye Decolourization
3.7. Extraction of Dye-Degraded Metabolites
3.8. Toxicity Evaluation of the Dye and the Biodegraded Product
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Axelesson, J.; Nilsson, U.; Terrazas, E.; Aliaga, T.A.; Welander, U. Decolorization of the textile dyes Reactive Red 2 and Reactive Blue 4 using Bjerkandera sp. Strain BOL 13 in a continuous rotating biological contactor reactor. Enzyme Microb. Technol. 2006, 39, 32. [Google Scholar] [CrossRef]
- Zee, F.P. Anaerobic Azo Dye Reduction. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2002. [Google Scholar]
- Schievano, E.; Menegazzo, I.; Marotta, E. Structure elucidation of the dye Acid red 131: Complete 1H, 13C and 15N NMR data assignment. Magn. Reson. Chem. 2011, 49, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Resmi, C.; Senan, T.; Abraham, E. Bioremediation of textile azo dyes by aerobic bacterial consortium. Biodegradation 2004, 15, 275–280. [Google Scholar]
- Kulandaivel, S.; Kaleeswari, P.; Mohanapriya, P. Decolorization and adsorption of dyes by consortium of bacteria with agriculture waste. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 865–882. [Google Scholar]
- Weisburger, J.H. Comments on the history and importance of aromatic and heterocyclic amines in public health. Mutat. Res. 2002, 506, 9–20. [Google Scholar] [CrossRef]
- Robinson, T.; McMullan, G.; Marchant, R.; Nigam, P. Remediation of dyes in textile effluents: A critical review on current treatment technologies with a proposed alternative. Bioresour. Technol. 2001, 77, 247–255. [Google Scholar] [CrossRef]
- Prigione, V.; Tigini, V.; Pezella, C.; Anastasi, A.; Sannia, G.; Varese, G.C. Decolourisation and detoxification of textile effluents by fungal biosorption. Water Res. 2008, 42, 2911–2920. [Google Scholar] [CrossRef] [PubMed]
- Prigione, V.; Varese, G.C.; Casieri, L.; Filipello Marchisio, V. Biosorption of simulated dyes effluents by inac-tivated fungal biomasses. Bioresour. Technol. 2008, 99, 3559–3567. [Google Scholar] [CrossRef] [PubMed]
- Kalyani, D.C.; Telke, A.A.; Dhanve, R.S.; Jadhav, J.P. Ecofriendly biodegradation and detoxification of Reactive Red 2 textile dye by newly isolated Pseudomonas sp. SUK1. J. Hazard. Mater. 2009, 163, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Wong, P.K.; Yuen, P.Y. Decolorization and biodegradation of Methyl Red by K. pneumonia RS13. Water Res. 1996, 30, 1736–1744. [Google Scholar] [CrossRef]
- Dhanve, R.S.; Shedbalkar, U.U.; Jadhav, J.P. Biodegradation of diazo reactive dye navy blue HE2R (Reactive blue 172) by an isolated Exiguobacterium sp. RD3. Biotechnol. Bioprocess Eng. 2008, 13, 53–60. [Google Scholar] [CrossRef]
- Mutafov, S.; Avramova, T.; Stefanova, L.; Angelova, B. Decolorization of acid orange 7 by bacteria of different tinactorial type: A comparative study. World J. Microbiol. Biotechnol. 2007, 23, 417–422. [Google Scholar] [CrossRef]
- Telke, A.; Kalyani, D.; Jadhav, J.; Govindwar, S. Kinetics and Mechanism of Reactive Red 141 Degradation by a Bacterial Isolate Rhizobiumradiobacter MTCC 8161. Acta Chim. Slov. 2008, 55, 320–329. [Google Scholar]
- Bella Devassy, T.; Goyal, D.; Khanna, S. Decolourization of textile azo dyes by aerobic bacterial consortium. Int. Biodeterior. Biodegrad. 2009, 63, 462–469. [Google Scholar]
- Cassidy, M.B.; Lee, H.; Trevors, J.T. Environmental application of immobilized microbial cells: A review. J. Ind. Microbiol. 1996, 16, 79–101. [Google Scholar] [CrossRef]
- Chang, J.S.; Lin, Y.C. Fed-Batch Bioreactor Strategies for Microbial Decolorization of Azo Dye using a Pseudomonasluteola Strain. Biotechnol. Prog. 2000, 16, 979–985. [Google Scholar] [CrossRef] [PubMed]
- Kodam, K.M.; Soojhawon, I.; Lokhande, P.D.; Gawai, K.R. Microbial decolorization of reactive azo dyes under aerobic conditions. World J. Microbiol. Biotechnol. 2005, 21, 367–370. [Google Scholar] [CrossRef]
- Santos-Gandelman, J.F.; Giambiagi-deMarval, M.; Oelemann, W.M.; Laport, M.S. Biotechnological potential of Sponge-associated bacteria. Curr. Pharm. Biotechnol. 2014, 15, 143–145. [Google Scholar] [CrossRef] [PubMed]
- Lakshmi, P.P.; Soundharya, B.; Amarendra, V.; Dhevendaran, K.; Santhosh, R.S. Isolation and characterization of Lysinibacillus sphaericus DSLS5 from marine sponge. Perspect. Biodivers. India 2014, 1, 229–236. [Google Scholar]
- Rajeswari, K.; Subashkumar, R.; Vijayaraman, K. Degradation of Textile dyes by isolated Lysinibacillus sphaericus strain RSV-1 and Stenotrophomonas maltophilia strain RSV-2 and Toxicity assessment of degraded product. J. Environ. Anal. Toxicol. 2014, 4, 222–226. [Google Scholar]
- Misal, S.A.; Lingojwar, D.P.; Lokhande, M.N.; Lokhande, P.D.; Gawai, K.R. Enzymatic transformation of nitro-aromatic compounds by a flavin-free NADH azoreductase from Lysinibacillus sphaericus. Biotechnol. Lett. 2014, 36, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Zhou, J.; Wang, D.; Tian, C.; Wang, P.; Salah Uddin, M.; Yu, H. Biocatalyst effects of immobilized anthraquinone on the anaerobic reduction of azo dyes by salt-tolerant bacteria. Water Res. 2007, 41, 426–432. [Google Scholar] [CrossRef] [PubMed]
- Djefal-Kerrar, A.; Abdoun, K.O.; Chouikrat, R.; El-Kahina, R.G.; Khodja, A.J.; Mahlous, M. Study of bacterium fixation stability on Gamma radiation synthesized carriers to improve degradation. J. Bioremed. Biodegrad. 2011, 2, 117–124. [Google Scholar]
- Solis, M.; Solis, A.; Perez, H.I.; Manjarrez, N.; Flores, M. Microbial decolorization of azo dyes: A review. Proc. Biochem. 2012, 47, 1723–1748. [Google Scholar] [CrossRef]
- Ali, H. Biodegradation of synthetic dyes—A review. Water Air Soil Pollut. 2010, 213, 251–273. [Google Scholar] [CrossRef]
- Devi, P.; Wahidullah, S.; Rodrigues, C.; D’Souza, L. The sponge-associated bacterium Bacillus licheniformis SAB1: A source of antimicrobial compounds. Mar. Drugs 2010, 8, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Bashan, Y. Alginate beads as synthetic inoculant carriers for the slow release of bacteria that affect plant growth. Appl. Environ. Microbiol. 1986, 51, 1089–1098. [Google Scholar] [PubMed]
- Devi, P.; Gomes, S.; Ribeiro, M.; Deshpande, S.A.; Singh, S.K.; Souza, L.D. Exploring rhizosphere bacteria of Eichornia crassipes for metal tolerance and biological activity. In Proceedings of the International Conference on Advances in Biotechnology (BioTech), Kanpur, India, 13–15 March 2015; pp. 2251–2489.
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Condition Causing Decolorization | Optimum Conditions |
|---|---|---|
| Free/encapsulated cells | Free as well as encapsulated | Encapsulated in alginate beads |
| Medium: liquid/solid | Both types | Liquid Nutrient Agar (NA) medium |
| pH | 5–11 | 7.2 |
| Temp | 26–34 °C | 28 °C |
| Saline/fresh water | 0–25 g/L | 16 g/L |
| Medium strength | Half/full strength | Full strength |
| Shaking/stationary | Both conditions | Shaker at 120 rpm |
| Reuse of beads | Upto threeuses | First use |
| Number of beads | Optional(3–35 beads/100 mL) | 27 beads/100 mL |
| Use of unlyophilized/lyophilized beads | Both produced results (8–10 h) Lyophilized beads stored for 1–3 months | Cells aged one week |
| Marine Bacterium (Gram Characteristics) | Dye (Untreated) | Treated Dye |
|---|---|---|
| Bacillus sp. LTW29 [G+] | - | - |
| Bacillus aerophilus strain IHB B15224 [G+] | + (2 mm) | - |
| Bacillus altitudinis strain BCX-57 [G+] | + (3 mm) | - |
| Bacillus sp. L219 [G+] | - | - |
| Bacillus pocheonensis BCX-16 [G+] | + (1 mm) | - |
| Bacillus flexus strain TAX4 [G+] | - | - |
| Oceanobacillus oncorhynchi strain AM-71 [G+] | - | - |
| Novosphingobium barchamii strain LL02 [G-] | - | - |
| Vibrio natriegens strain F7S194 [G-] | - | - |
| Halomonas sp. WHB35 [G-] | - | - |
| Pseudovibrio sp. 010K 105-5-5 [G-] | - | - |
| Paracoccusyeei strain G1212 [G-] | - | - |
| Product | Mass/Charge (m/z) | Collision Energy (V) | m/z (% Relative Intensity) |
|---|---|---|---|
| Dye alone (A) | [M + H]+ 379 | 3.34 × 107 | 379, 362, 298, 223, 158, 156, 128 |
| Dimer (2) | [M + H]+ 743 | 4.69 × 105 | 742, 724, 625, 588, 567, 512, 484, 441 (100%), 358, 324, 211, 169, 120 |
| (4) | [M + H]+ 472 | 1.29 × 107 | 472, 227 |
| (5) | [M + H]+ 344 | 3.60 × 107 | 344, 211, 212 |
| (7) | [M + H]+ 508 | 2.38 × 107 | 508, 439, 245, 246 |
| (8) | [M + H]+ 245 | 1.10 × 107 | 245, 217, 172, 154, 120 (100%), 98, 86, 70 |
| (9) | [M + H]+ 166 | 1.25 × 108 | 166, 167 |
| (13) | [M + H]+ 227 | 1.56 × 107 | 227, 197 |
| (14) | [M + H]+ 205 | 1.69 × 107 | 205, 206 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Devi, P.; Wahidullah, S.; Sheikh, F.; Pereira, R.; Narkhede, N.; Amonkar, D.; Tilvi, S.; Meena, R.M. Biotransformation and Detoxification of Xylidine Orange Dye Using Immobilized Cells of Marine-Derived Lysinibacillus sphaericus D3. Mar. Drugs 2017, 15, 30. https://doi.org/10.3390/md15020030
Devi P, Wahidullah S, Sheikh F, Pereira R, Narkhede N, Amonkar D, Tilvi S, Meena RM. Biotransformation and Detoxification of Xylidine Orange Dye Using Immobilized Cells of Marine-Derived Lysinibacillus sphaericus D3. Marine Drugs. 2017; 15(2):30. https://doi.org/10.3390/md15020030
Chicago/Turabian StyleDevi, Prabha, Solimabi Wahidullah, Farhan Sheikh, Rochelle Pereira, Niteen Narkhede, Divya Amonkar, Supriya Tilvi, and Ram Murthy Meena. 2017. "Biotransformation and Detoxification of Xylidine Orange Dye Using Immobilized Cells of Marine-Derived Lysinibacillus sphaericus D3" Marine Drugs 15, no. 2: 30. https://doi.org/10.3390/md15020030
APA StyleDevi, P., Wahidullah, S., Sheikh, F., Pereira, R., Narkhede, N., Amonkar, D., Tilvi, S., & Meena, R. M. (2017). Biotransformation and Detoxification of Xylidine Orange Dye Using Immobilized Cells of Marine-Derived Lysinibacillus sphaericus D3. Marine Drugs, 15(2), 30. https://doi.org/10.3390/md15020030