Growth, Toxin Production and Allelopathic Effects of Pseudo-nitzschia multiseries under Iron-Enriched Conditions


Abstract
:1. Introduction
2. Results
2.1. Experiment 1: Pseudo-nitzschia multiseries
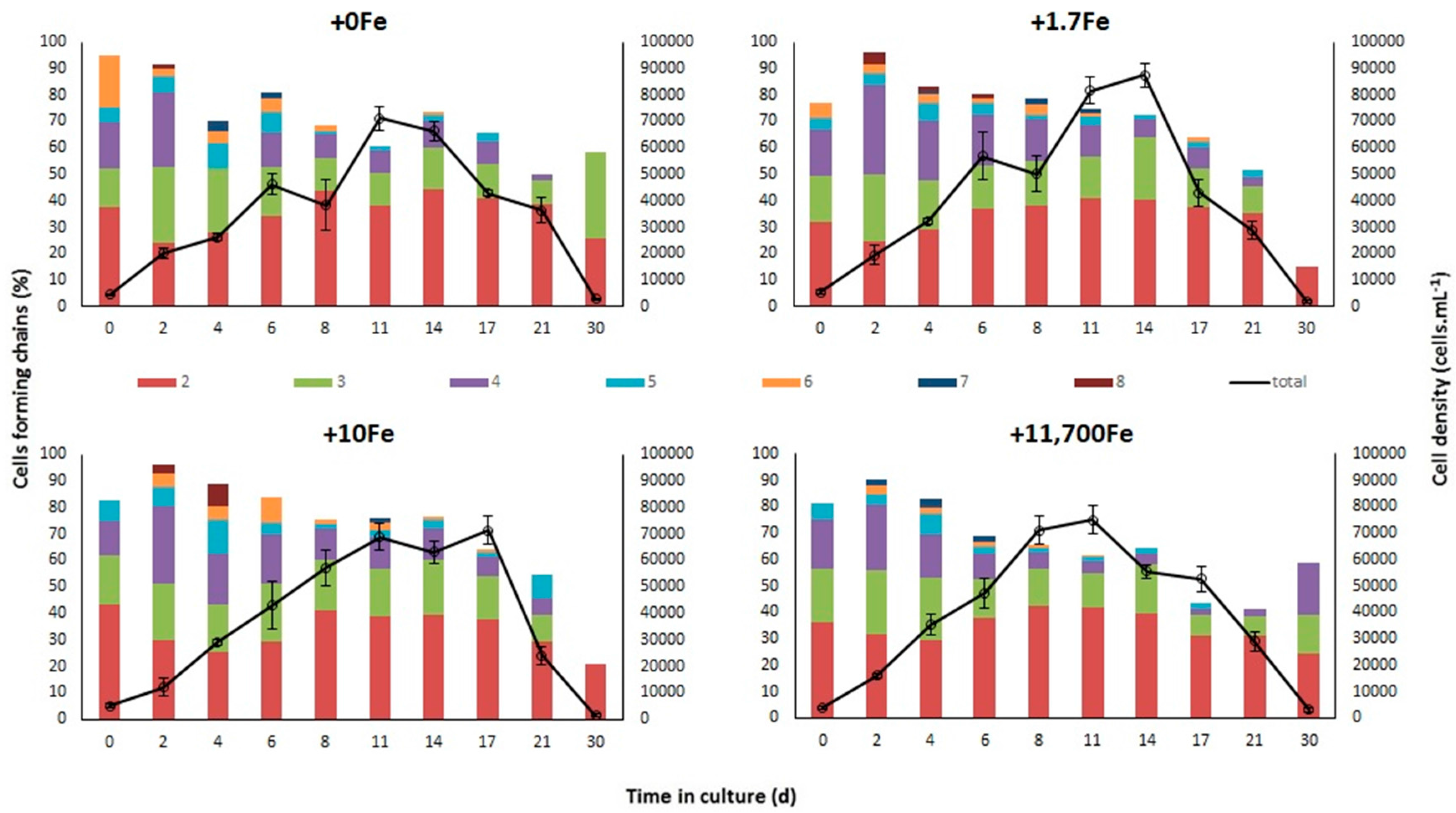
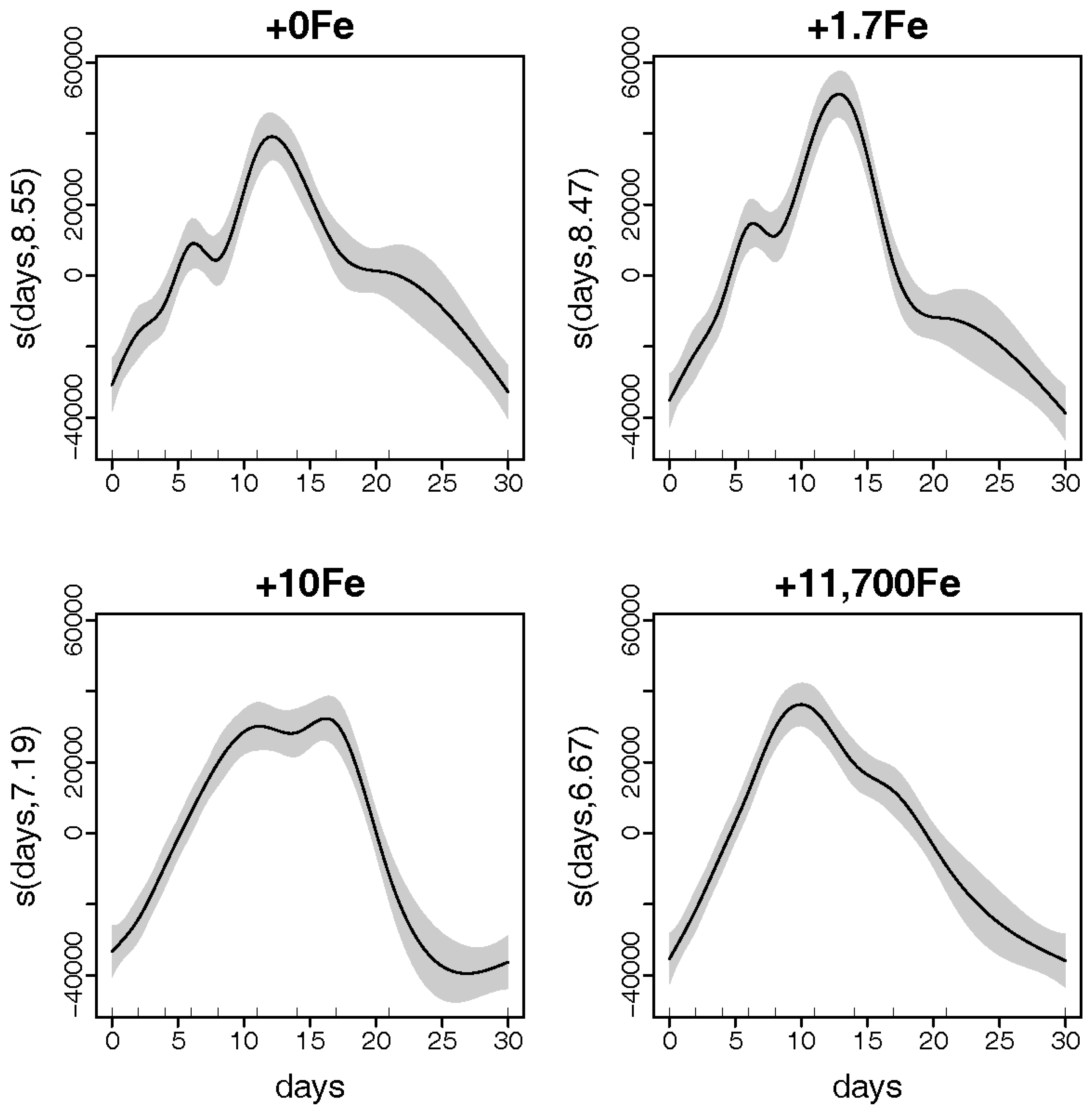
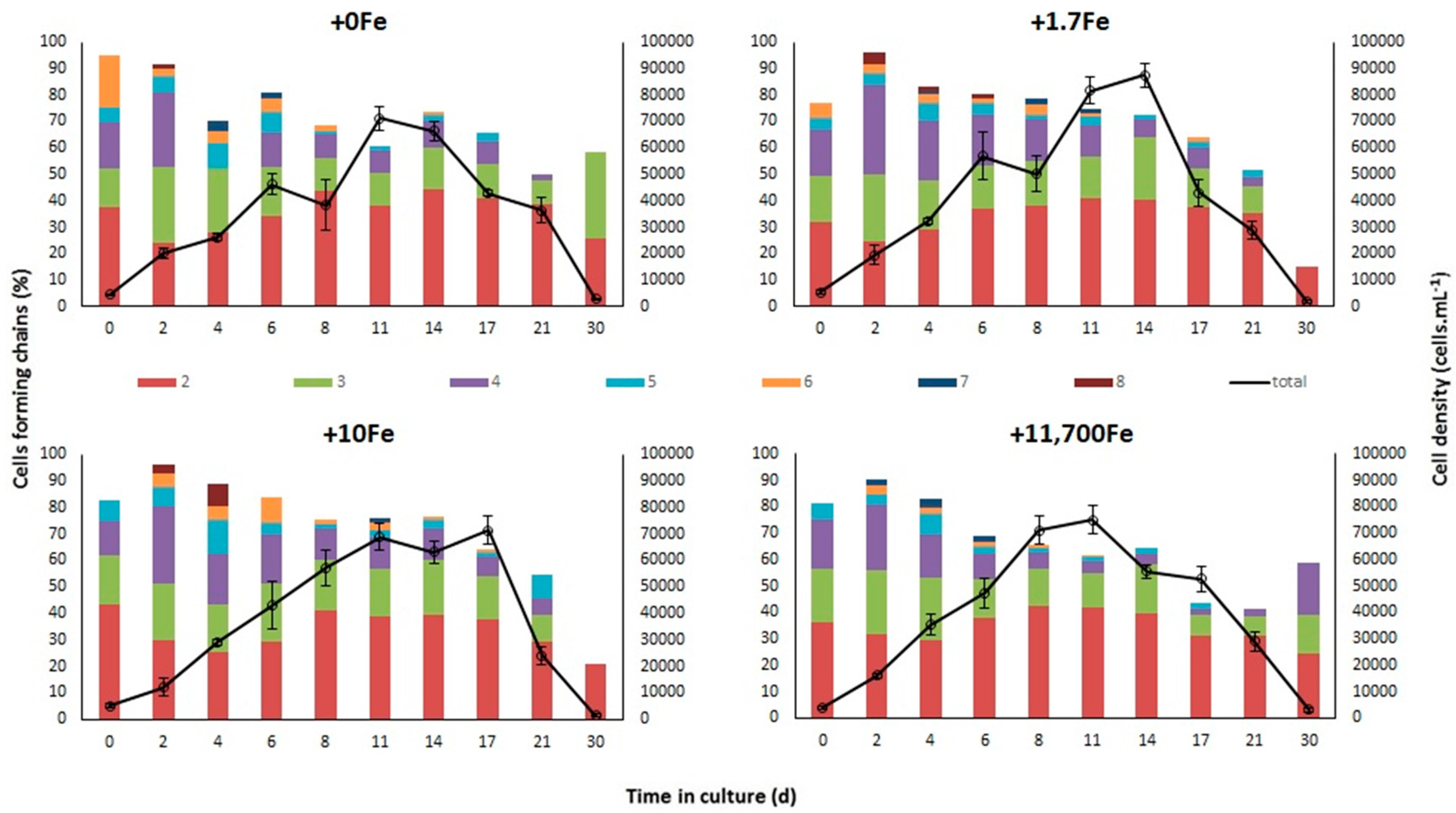
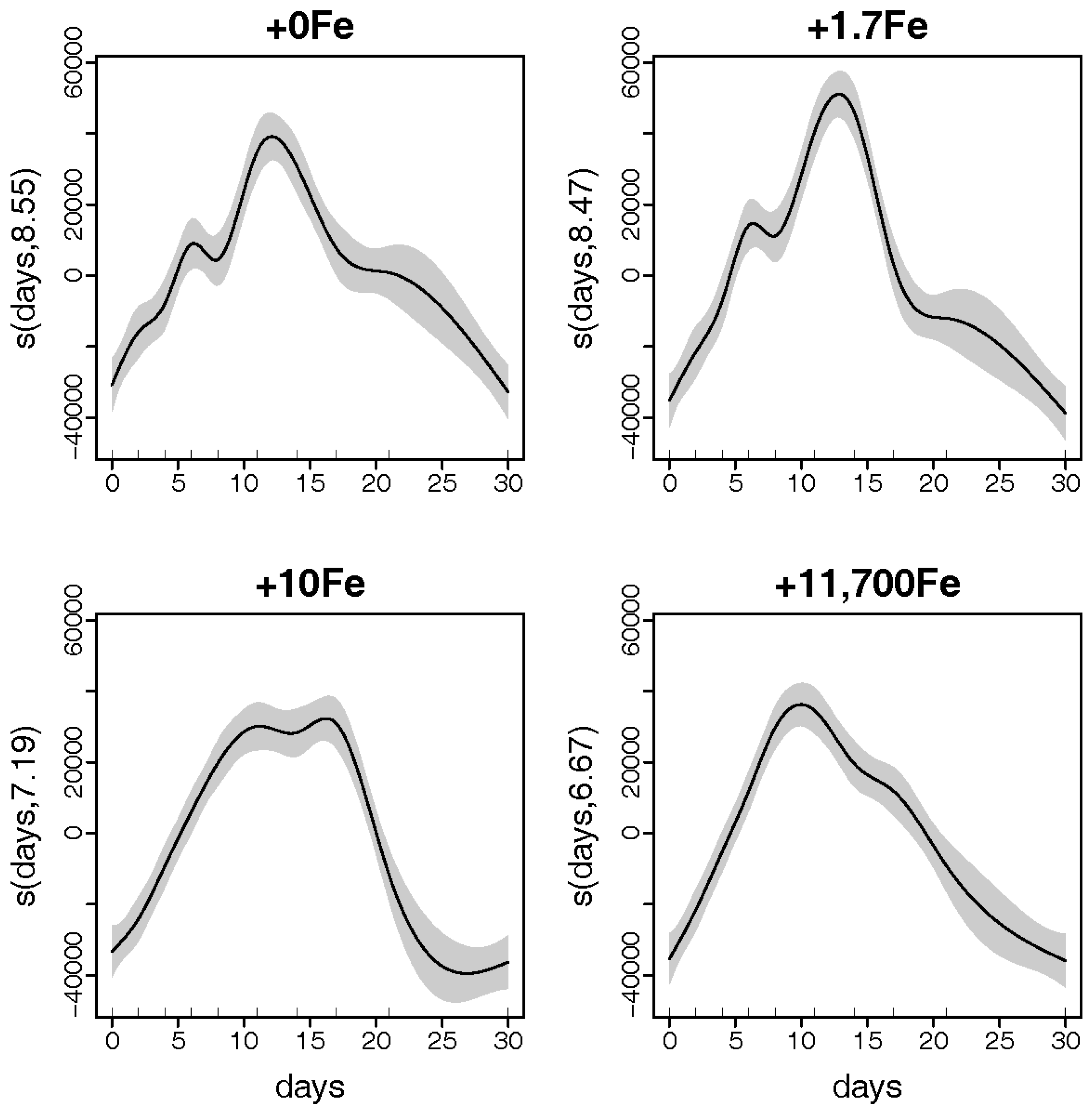
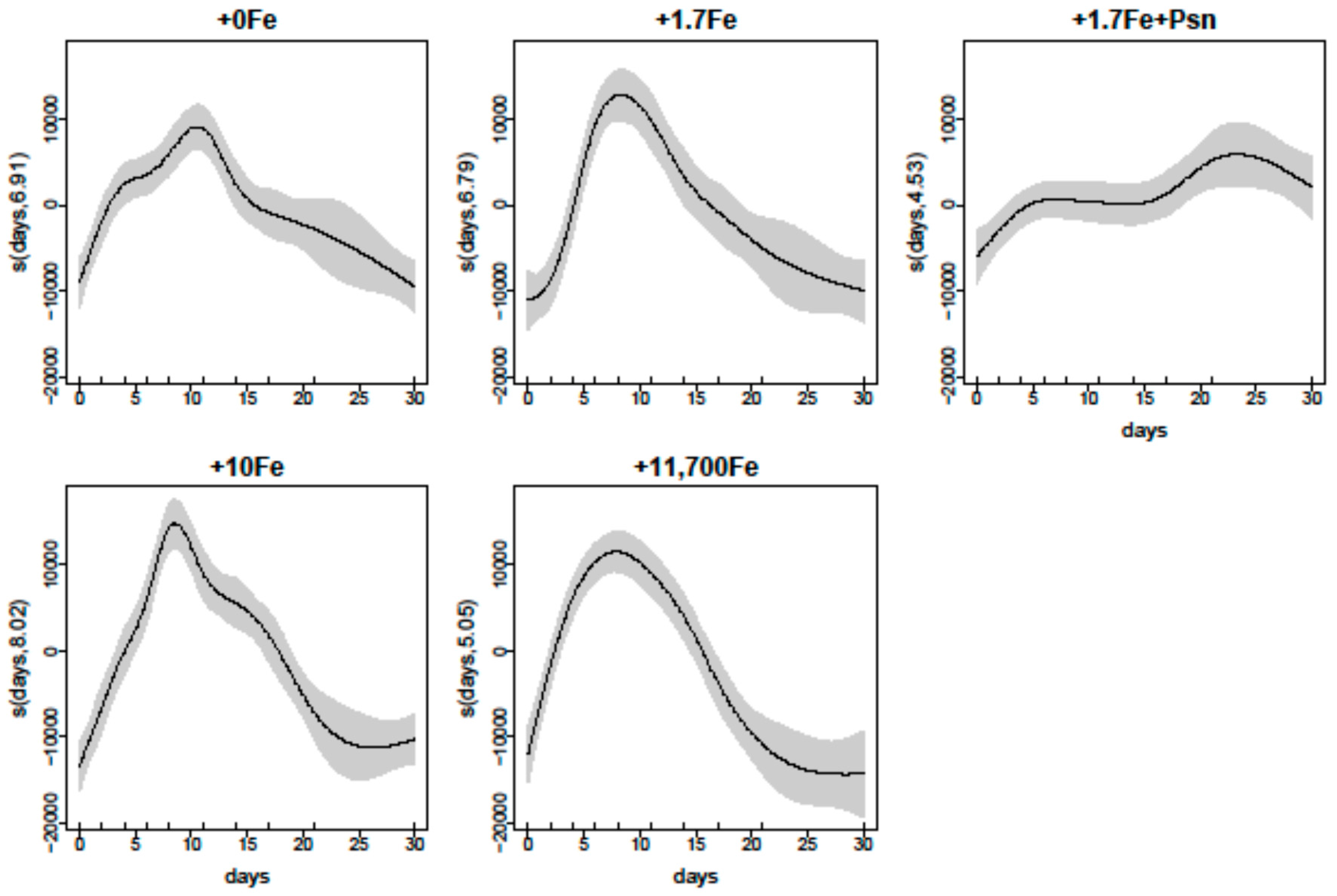
2.1.1. Growth
2.1.2. Domoic Acid
2.2. Experiment 2: Bacillaria sp.
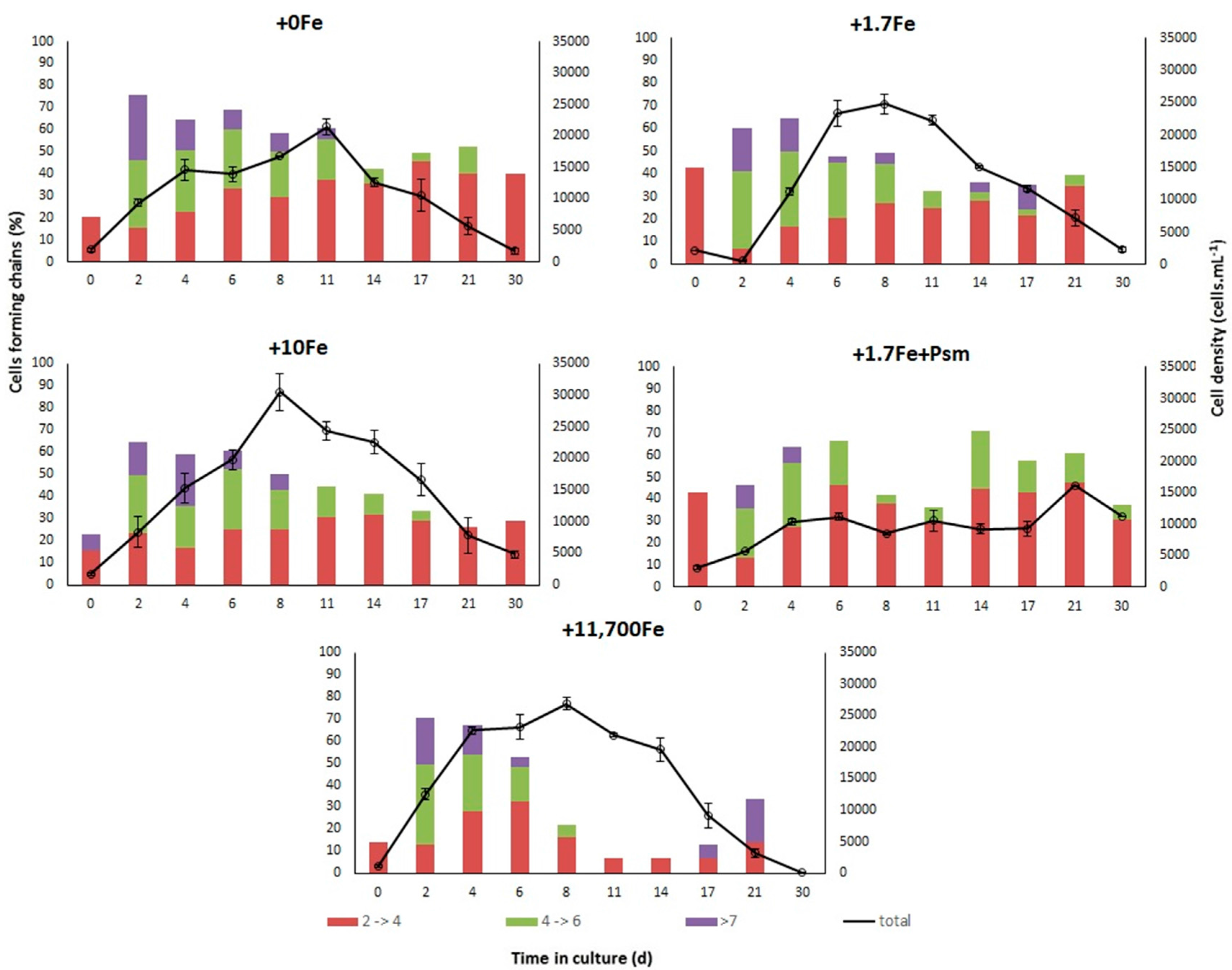
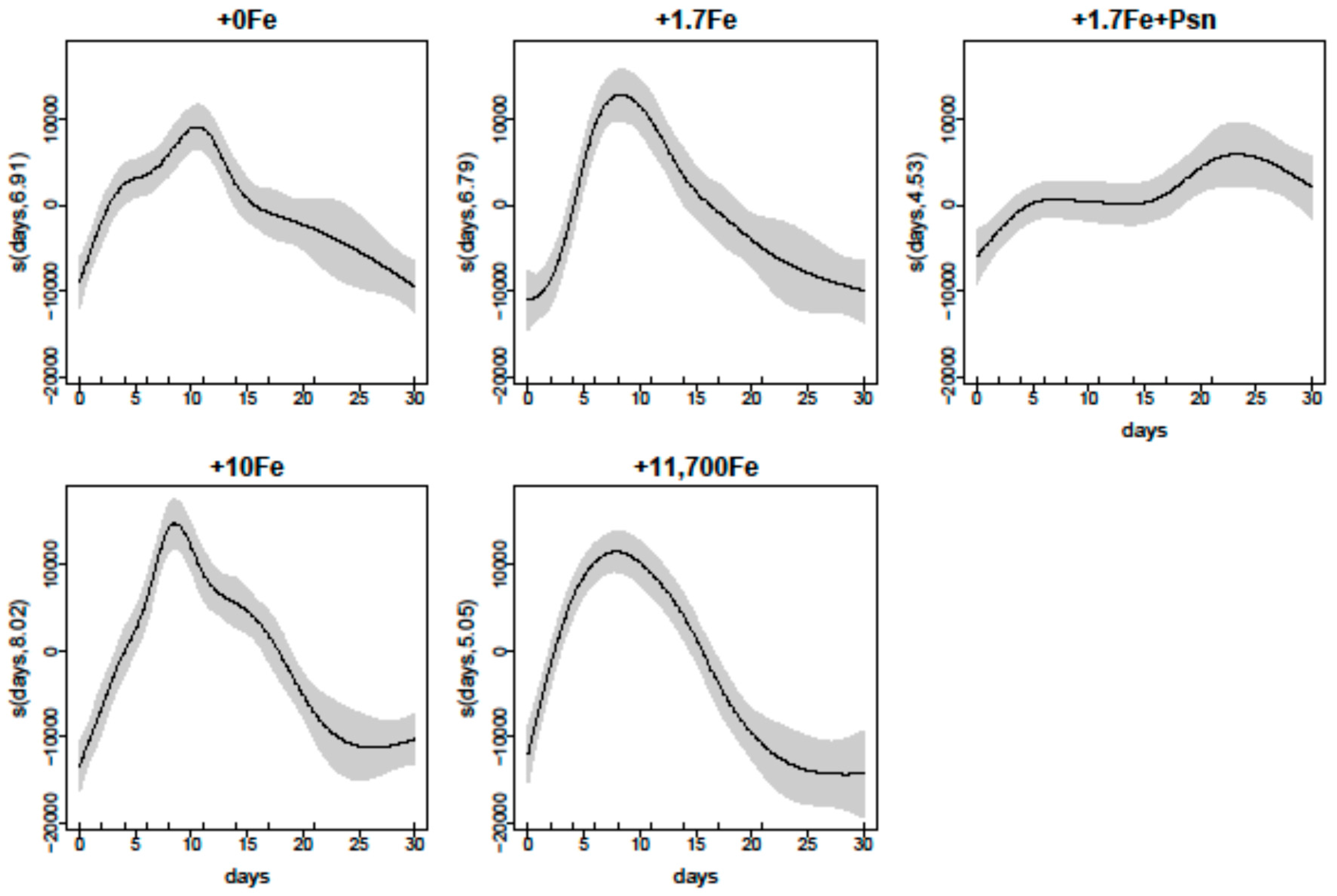
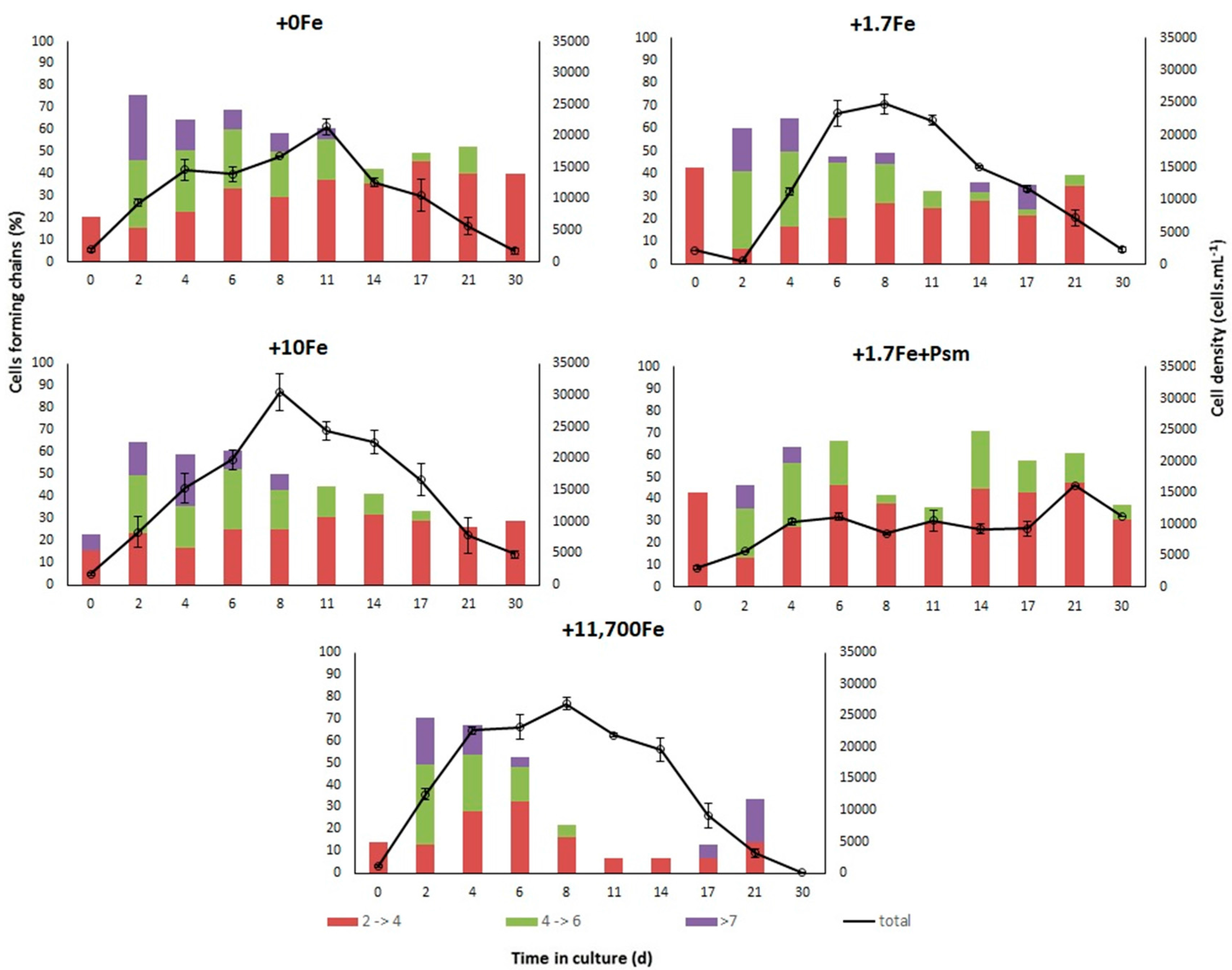
2.2.1. Growth
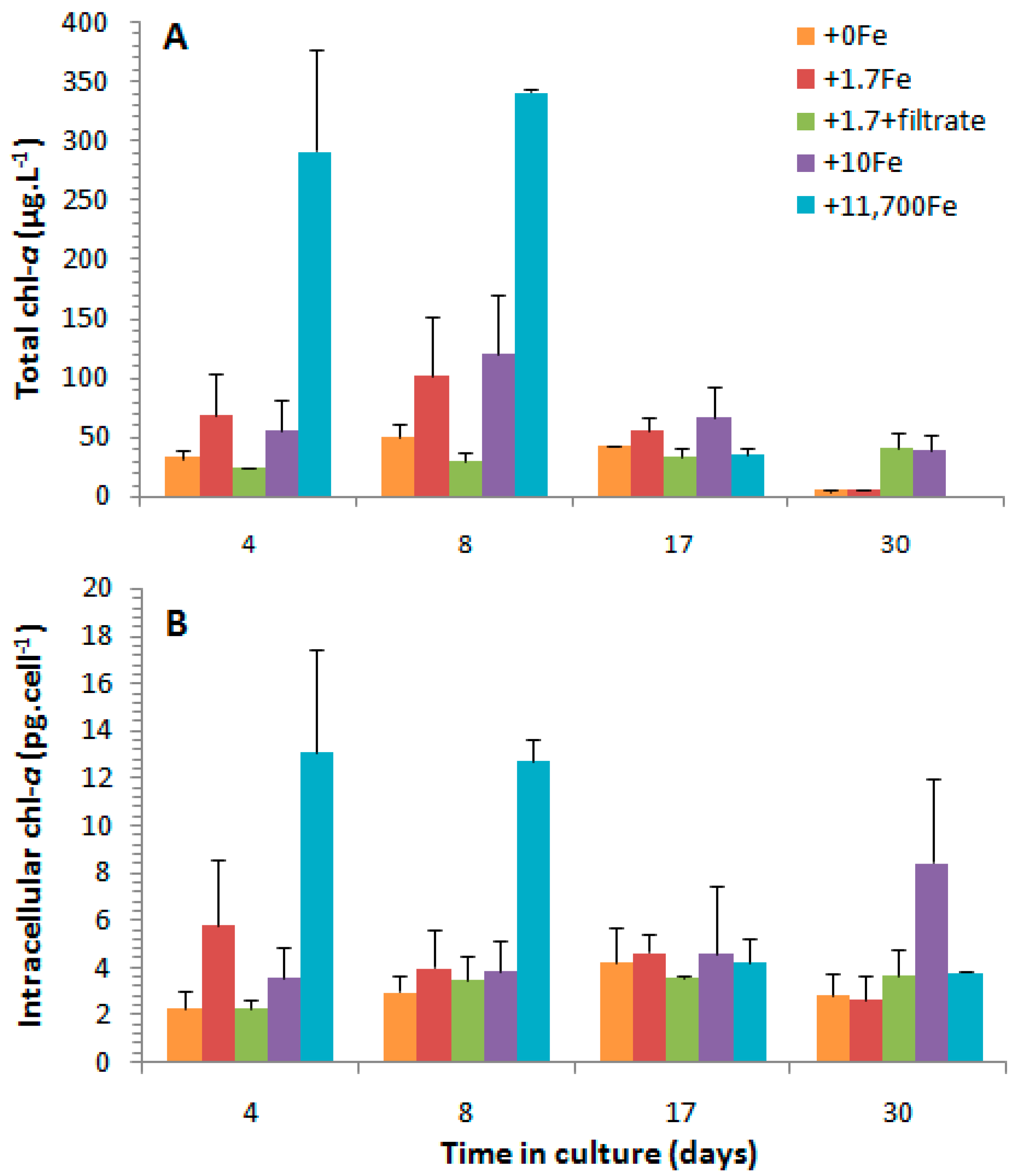
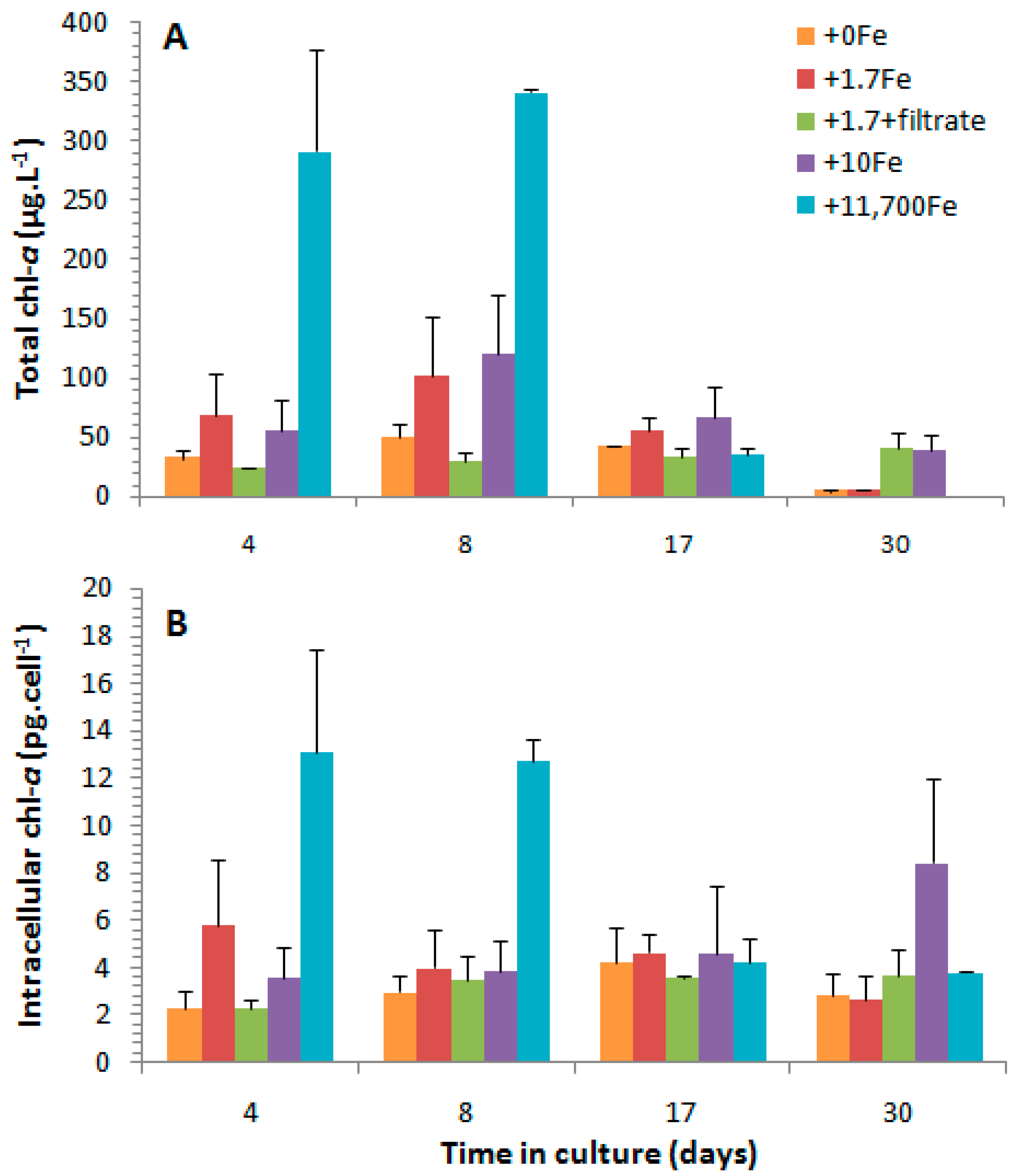
2.2.2. Chlorophyll-a
3. Discussion
3.1. Effects of Fe-Enrichment on the Growth of P. multiseries and Bacillaria sp.
3.2. Effects of Fe-Enrichment on Toxin Production by P. multiseries
3.3. Effects of the Cell Contents from P. multiseries Cultures on Bacillaria sp.
4. Material and Methods
4.1. Culture Establishment and Acclimation
4.2. Growth and Domoic Acid Production Experiments
4.3. Chemical Analysis
4.3.1. Domoic Acid
4.3.2. Chlorophyll-a
4.4. Data Treatment and Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bates, S.S.; Bird, C.J.; de Freitas, A.S.W.; Foxall, R.; Gilgan, M.; Hanic, L.A.; Johnson, G.R.; McCulloch, A.W.; Odense, P.; Pocklington, R.; et al. Pennate diatom Niztschia pungens as the primary source of domoic acid, a toxin in shellfish from eastern Prince Edward Island. Can. J. Fish. Aquat. Sci. 1989, 46, 1203–1215. [Google Scholar] [CrossRef]
- Wright, J.L.C.; Boyd, R.K.; De Freitas, A.S.W.; Falk, M.; Foxall, R.A.; Jamieson, W.D.; Laycock, M.V.; McCulloch, A.W.; McInnes, A.G.; Odense, P.; et al. Identification of domoic acid, a neuroexcitatory amino acid, in toxic mussels from eastern Prince Edward Island. Can. J. Chem. 1989, 67, 481–490. [Google Scholar] [CrossRef]
- Scholin, C.A.; Gulland, F.; Doucette, G.J.; Benson, S.; Busman, M.; Chavez, F.P.; Cordaro, J.; DeLong, R.; De Vogelaere, A.; Harvey, J.; et al. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 403, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, K.A.; Silver, M.W.; Coale, S.L.; Tjeerdema, R.S. Domoic acid in planktivorous fish in relation to toxic Pseudo-nitzschia cell densities. Mar. Biol. 2002, 140, 625–631. [Google Scholar]
- McCabe, R.M.; Hickey, B.M.; Kudela, R.M.; Lefebvre, K.A.; Adams, N.G.; Bill, B.D.; Gulland, F.M.D.; Thomson, R.E.; Cochlan, W.P.; Trainer, V.L. An unprecedented coastwide toxic algal bloom linked to anomalous ocean conditions. Geophys. Res. Lett. 2016, 43, 10366–10376. [Google Scholar] [CrossRef] [PubMed]
- Rue, E.; Bruland, K. Domoic acid binds iron and copper: A possible role for the toxin produced by the marine diatom Pseudo-nitzschia. Mar. Chem. 2001, 76, 127–134. [Google Scholar] [CrossRef]
- Kumar, K.P.; Kumar, S.; Nair, G.A. Risk assessment of the amnesic shellfish poison, domoic acid, on animals and humans. J. Environ. Biol. 2009, 30, 319–325. [Google Scholar] [PubMed]
- SERGAS. Primeira Notificación de Intoxicación po la Biotoxina ASP en Galicia. Boletín Epidemiolóxico de Galicia. 1 August 2014, Volume 3. Número 16. Available online: https://www.sergas.es/Saude-publica/Documents/1590/Venres_Epidemioloxico_vol3_n16_20140801.pdf (accessed on 24 October 2017).
- Hasle, G.R. Are most of the domoic acid-producing species of the diatom genus Pseudo-nitzschia cosmopolites? Harmful Algae 2002, 1, 137–146. [Google Scholar] [CrossRef]
- Wells, M.L.; Trick, C.G.; Cochlan, W.P.; Hughes, M.P.; Trainer, V.L. Domoic acid: The synergy of iron, copper, and the toxicity of diatoms. Limnol. Oceanogr. 2005, 50, 1908–1917. [Google Scholar] [CrossRef]
- Lundholm, N. Bacillariophyceae. IOC-UNESCO Taxonomic Reference List of Harmful Micro Algae. 2017. Available online: http://www.marinespecies.org/hab (accessed on 25 August 2017).
- Lelong, A.; Hégaret, H.; Soudant, P.; Bates, S.S. Pseudo-nitzschia (Bacillariophyceae) species, domoic acid and amnesic shellfish poisoning: Revisiting previous paradigms. Phycologia 2012, 51, 168–216. [Google Scholar] [CrossRef]
- Trainer, V.L.; Bates, S.S.; Lundholm, N.; Thessen, A.E.; Cochlan, W.P.; Adams, N.G.; Trick, C.G. Pseudo-nitzschia physiological ecology, phylogeny, toxicity, monitoring and impacts on ecosystem health. Harmful Algae 2012, 14, 271–300. [Google Scholar] [CrossRef]
- Mafra, L.L., Jr.; Fernandes, L.F.; Proença, L.A.O. Harmful algae and toxins in Paranaguá Bay, Brazil: Bases for monitoring. Braz. J. Oceanogr. 2006, 54, 107–121. [Google Scholar] [CrossRef]
- Bates, S.S.; Richard, J. Domoic acid production and cell division by Pseudo-nitzschia multiseries in relation to a light: Dark cycle in silicate-limited chemostat culture. Can. Tech. Rep. Fish. Aquat. Sci. 1996, 0, 140–143. [Google Scholar]
- Fehling, J.; Davidson, K.; Bates, S.S. Growth dynamics of non-toxic Pseudo-nitzschia delicatissima and toxic P. seriata (Bacillariophyceae) under simulated spring and summer photoperiods. Harmful Algae 2005, 4, 763–769. [Google Scholar] [CrossRef]
- Lundholm, N.; Hansen, P.J.; Kotaki, Y. Effect of pH on growth and domoic acid production by potentially toxic diatoms of the genera Pseudo-nitzschia and Nitzschia. Mar. Ecol. Prog. Ser. 2004, 273, 1–15. [Google Scholar] [CrossRef]
- Doucette, G.J.; King, K.L.; Thessen, A.E.; Dortch, Q. The effect of salinity on domoic acid production by the diatom Pseudo-nitzschia multiseries. Nova Hedwig. 2008, 133, 31–46. [Google Scholar]
- Hansen, L.R.; Soylu, S.Í.; Kotaki, Y.; Moestrup, Ø.; Lundholm, N. Toxin production and temperature-induced morphological variation of the diatom Pseudo-nitzschia seriata from the Arctic. Harmful Algae 2011, 10, 689–696. [Google Scholar] [CrossRef]
- Bates, S.; de Freitas, A.S.W.; Milley, J.E.; Pocklington, R.; Quilliam, M.; Smith, J.C.; Worms, J. Controls on domoic acid production by the diatom Nitzschia punges f. multiseries in culture: Nutrients and irradiance. Can. J. Fish. Aquat. Sci. 1991, 48, 1136–1144. [Google Scholar] [CrossRef]
- Pan, Y.; Subba Rao, D.V.; Mann, K.H.; Brown, R.G.; Pocklington, R. Effets of silicate limitation on production of domoic acid, a neurotoxin, by the diatom Pseudo-nitschia multiseries. I. Batch culture studies. Mar. Ecol. Prog. Ser. 1996, 131, 225–233. [Google Scholar] [CrossRef]
- Martin-Jézéquel, V.; Calu, G.; Candela, L.; Amzil, Z.; Jauffrais, T.; Séchet, V.; Weigel, P. Effects of organic and inorganic nitrogen on the growth and production of domoic acid by Pseudo-nitzschia multiseries and P. australis (Bacillariophyceae) in culture. Mar. Drugs 2015, 13, 7067–7086. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.; Kudela, R.; Birch, J.; Blum, M.; Bowers, H.; Chavez, F.; Doucette, G.; Hayashi, K.; Marin, R., III; Mikulski, C.; et al. Causality of an extreme harmful algal bloom in Monterey Bay, California during the 2014–2016 northeast Pacific warm anomaly. Geophys. Res. Lett. 2017, 44, 5571–5579. [Google Scholar] [CrossRef]
- Bates, S.S.; Léger, C.; Satchwell, M.; Boyer, G.L. The effects of iron on Domoic Acid production by Pseudo-nitzchia multiseries. In Harmful Algal Blooms 2000, Proceedings of the 9th International Conference on Harmful Algal Blooms, Hobart, Tasmania, 7–11 February 2000; Hallegraeff, G.A., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; IOC/UNESCO: Paris, France, 2001; pp. 320–323. [Google Scholar]
- Maldonado, M.T.; Hughes, M.P.; Rue, E.L.; Wells, M.L. The effect of Fe and Cu on growth and domoic acid production by Pseudo-nitzschia multiseries and Pseudo-nitzschia australis. Limnol. Oceanogr. 2002, 47, 515–526. [Google Scholar] [CrossRef]
- Fuentes, M.S.; Wikfors, G.H. Control of domoic acid toxin expression in Pseudo-nitzschia multiseries by copper and silica: Relevance to mussel aquaculture in New England (USA). Mar. Environ. Res. 2013, 83, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Withfield, M. Interactions between phytoplankton and trace metals in the ocean. Adv. Mar. Biol. 2001, 41, 3–128. [Google Scholar]
- Wells, M.L.; Prince, N.M.; Bruland, K.W. Iron chemistry in seawater and its relationships to phytoplankton: A workshop report. Mar. Chem. 1994, 48, 157–182. [Google Scholar] [CrossRef]
- Endo, H.; Youshimura, T.; Kataoka, T.; Suzuki, K. Effects of CO2 and iron avalilability on phytoplankton and eubacterial community compositions in the northwest subarctic Pacific. J. Exp. Mar. Ecol. 2013, 439, 160–175. [Google Scholar] [CrossRef]
- Albrecht-Gary, A.M.; Crumbliss, A.L. Coordination chemistry of siderophores: Thermodynamics and kinetics of iron chelation and release. Met. Ions Biol Syst. 1998, 35, 239–327. [Google Scholar] [PubMed]
- Russo, A.D.P.G.; Souza, M.S.; Mendes, C.R.B.; Jesus, B.; Tavano, V.M.; Garcia, A.E. Photophysiological effects of Fe concentration on diatom-dominated phytoplankton assemblages in the Antarctic Peninsula region. J. Exp. Mar. Biol. Ecol. 2015, 466, 49–58. [Google Scholar] [CrossRef]
- Sarthou, G.; Timmermans, K.R.; Blain, S.; Tréguer, P. Growth physiology and fate of diatoms in the ocean: A review. J. Sea Res. 2005, 53, 25–42. [Google Scholar] [CrossRef]
- Marchetti, A.; Maldonado, M.T.; Lane, E.S.; Harrison, P.J. Iron requirements of the pennate diatom Pseudo-nitzschia: Comparison of oceanic (high-nitrate, low-chlorophyll waters) and coastal species. Limnol. Oceanogr. 2006, 51, 2092–2101. [Google Scholar] [CrossRef]
- Marchetti, A.; Parker, M.S.; Moccia, L.P.; Lin, E.O.; Arrieta, A.L.; Ribalet, F.; Murphy, M.E.P.; Maldonado, M.T.; Armbrust, E.V. Ferritin is used for iron storage in bloom-forming marine pennate diatoms. Nature 2009, 457, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Bates, S.S.; Garrison, D.L.; Horner, R.A. Bloom Dynamics and Physiology of Domoic acid Producing Pseudo-nitzschia species. In Physiological Ecology of Harmful Algal Blooms; Springer: Heidelberg, Germany, 1998; pp. 267–292. [Google Scholar]
- Pan, Y.; Bates, S.S.; Cembella, A.D. Environmental stress and domoic acid production by Pseudo-nitzschia: A physiological perspective. Nat. Toxins 1998, 6, 127–135. [Google Scholar] [CrossRef]
- Bouillon, R.C.; Kieber, R.J.; Skrabal, S.A.; Wright, J.L.C. Photochemistry and identification of photodegradation products of the marine toxin domoic acid. Mar. Chem. 2008, 110, 18–27. [Google Scholar] [CrossRef]
- Lundholm, N.; Hansen, P.J.; Kotaki, Y. Lack of allelopathic effects of the domoic acid-producing marine diatom Pseudo-nitzschia multiseries. Mar. Ecol. Prog. Ser. 2005, 288, 21–33. [Google Scholar] [CrossRef]
- Xu, N.; Tang, Y.Z.; Qin, J.; Duan, S.; Gobler, C.J. Ability of the marine diatoms Pseudo-nitzschia multiseries and P. pungens to inhibit the growth of co-occurring phytoplankton via allelopathy. Aquat. Microb. Ecol. 2015, 74, 29–41. [Google Scholar] [CrossRef]
- Granéli, E.; Hanses, P.J. Allelopathy in Harmful Algae: A Mechanism to Compete for Resources? In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2007; Volume 88, pp. 189–201. ISBN 978-3-540-32209-2. [Google Scholar]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chaley, H.H., Eds.; Plenum Press: New York, NY, USA, 1975; pp. 29–60. [Google Scholar]
- Price, N.M.; Harrison, G.I.; Hering, J.G.; Hudson, R.J.; Nirel, P.M.V.; Palenik, B.; Morel, F.M.M. Preparation and Chemistry of the Artificial Algal Culture Medium Aquil. Biol. Oceanogr. 1989, 6, 443–461. [Google Scholar]
- Mafra, L.L.; Léger, C.; Bates, S.S.; Quilliam, M.A. Analysis of trace levels of domoic acid in seawater and plankton by liquid chromatography without derivatization, using UV or mass spectrometry detection. J. Chromatogr. A 2009, 1216, 6003–6011. [Google Scholar] [CrossRef] [PubMed]
- Arar, E.J.; Collins, G.B. Method 445.0 In Vitro Determination of Chlorophyll a and Pheophytin-a in Marine and Freshwater Algae by Fluorescence; U.S. Environmental Protection Agency, National Expossure Research Laboratory: Cincinnati, OH, USA, 1997.
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Sandrini-Neto, L.; Camargo, M.G. GAD: An R Package for ANOVA Designs from General Principles. R Package Version 1.1.1. 2012. Available online: http://CRAN.R-project.org/package=GAD (accessed on 10 August 2017).
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G. Mixed Effects Models and Extension in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | P. multiseries | Bacillaria sp. | ||
|---|---|---|---|---|
| µ0–8 | µ | µ0–8 | µ | |
| +0Fe | 0.25 (±0.05) | 0.74 (±0.05) | 0.28 (±0.02) | 0.81 (±0.05) |
| +1.7Fe | 0.28 (±0.02) | 0.65 (±0.06) | 0.30 (±0.01) | 0.91 (±0.04) |
| +1.7Fe+Psm | N/A * | N/A | 0.13 (±0.02) | 0.36 (±0.03) |
| +10Fe | 0.31 (±0.01) | 0.54 (±0.03) | 0.36 (±0.02) | 0.80 (±0.16) |
| +11,700Fe | 0.36 (±0.02) | 0.71 (±0.09) | 0.41 (±0.01) | 1.25 (±0.02) |
| Days | Treatments | DA Concentration | ||
|---|---|---|---|---|
| Intracellular (pg·Cell−1) | Extracellular (pg·Cell−1) | Total (µg·mL−1) | ||
| 4 | +0Fe | 0.04 (0.001) | 0.44 (0.07) | 0.01 (0.001) |
| +1.7Fe | 0.03 (0.001) | 0.54 (0.09) | 0.02 (0.002) | |
| +10Fe | 0.04 (0.005) | 0.72 (0.17) | 0.02 (0.008) | |
| +11,700Fe | 0.04 (0.005) | 0.74 (0.20) | 0.03 (0.007) | |
| 8 | +0Fe | 0.06 (0.01) | 1.85 (1.02) | 0.05 (0.02) |
| +1.7Fe | 0.06 (0.02) | 0.81 (0.49) | 0.03 (0.001) | |
| +10Fe | 0.03 (0.006) | 0.74 (0.18) | 0.04 (0.01) | |
| +11,700Fe | 0.20 (0.02) | 0.55 (0.08) | 0.04 (0.008) | |
| 17 | +0Fe | 0.26 (0.06) | 1.95 (1.11) | 0.10 (0.05) |
| +1.7Fe | 0.27 (0.07) | 25.8 (16.7) | 1.15 (0.65) | |
| +10Fe | 0.14 (0.03) | 2.15 (1.05) | 0.18 (0.08) | |
| +11,700Fe | 0.45 (0.11) | 91.3 (38.5) | 4.75 (1.88) | |
| 30 | +0Fe | 0.84 (0.31) | 18.6 (11.5) | 0.66 (0.34) |
| +1.7Fe | 1.21 (0.37) | 4.56 (1.95) | 0.20 (0.08) | |
| +10Fe | 1.29 (0.42) | 7.24 (5.50) | 0.21 (0.14) | |
| +11,700Fe | 2.07 (1.25) | 6.83 (2.51) | 0.28 (0.11) | |
| Low-Fe Addition (0–1.7 nmol·L−1) | High-Fe Addition (120–11,700 nmol·L−1) | |||||
|---|---|---|---|---|---|---|
| Exponential phase | P | D | T | P | D | T |
| Maldonado et al. (2002) [25] | 0.07 | 1.8 | 0.03 | 0.21 | ||
| Present study | 0.06 | 0.81–1.8 | 0.20 | 0.55 | ||
| Early stationary phase | ||||||
| Bates et al. (2001) [24] | 4.0 | 14–18 | ||||
| Present study | 0.26 | 1.95–25.8 | 0.45 | 91.3 | ||
| Declining/late stationary phase | ||||||
| Bates et al. (2001) [24] | 5.0 | 22–46 | ||||
| Present study | 0.84–1.21 | 4.56–18.6 | 2.07 | 6.83 | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobrinho, B.F.; De Camargo, L.M.; Sandrini-Neto, L.; Kleemann, C.R.; Machado, E.d.C.; Mafra, L.L. Growth, Toxin Production and Allelopathic Effects of Pseudo-nitzschia multiseries under Iron-Enriched Conditions. Mar. Drugs 2017, 15, 331. https://doi.org/10.3390/md15100331
Sobrinho BF, De Camargo LM, Sandrini-Neto L, Kleemann CR, Machado EdC, Mafra LL. Growth, Toxin Production and Allelopathic Effects of Pseudo-nitzschia multiseries under Iron-Enriched Conditions. Marine Drugs. 2017; 15(10):331. https://doi.org/10.3390/md15100331
Chicago/Turabian StyleSobrinho, Bruna Fernanda, Luana Mocelin De Camargo, Leonardo Sandrini-Neto, Cristian Rafael Kleemann, Eunice da Costa Machado, and Luiz Laureno Mafra. 2017. "Growth, Toxin Production and Allelopathic Effects of Pseudo-nitzschia multiseries under Iron-Enriched Conditions" Marine Drugs 15, no. 10: 331. https://doi.org/10.3390/md15100331
APA StyleSobrinho, B. F., De Camargo, L. M., Sandrini-Neto, L., Kleemann, C. R., Machado, E. d. C., & Mafra, L. L. (2017). Growth, Toxin Production and Allelopathic Effects of Pseudo-nitzschia multiseries under Iron-Enriched Conditions. Marine Drugs, 15(10), 331. https://doi.org/10.3390/md15100331