
Chitosan Oligosaccharide Reduces Propofol Requirements and Propofol-Related Side Effects

Abstract
:1. Introduction
2. Results
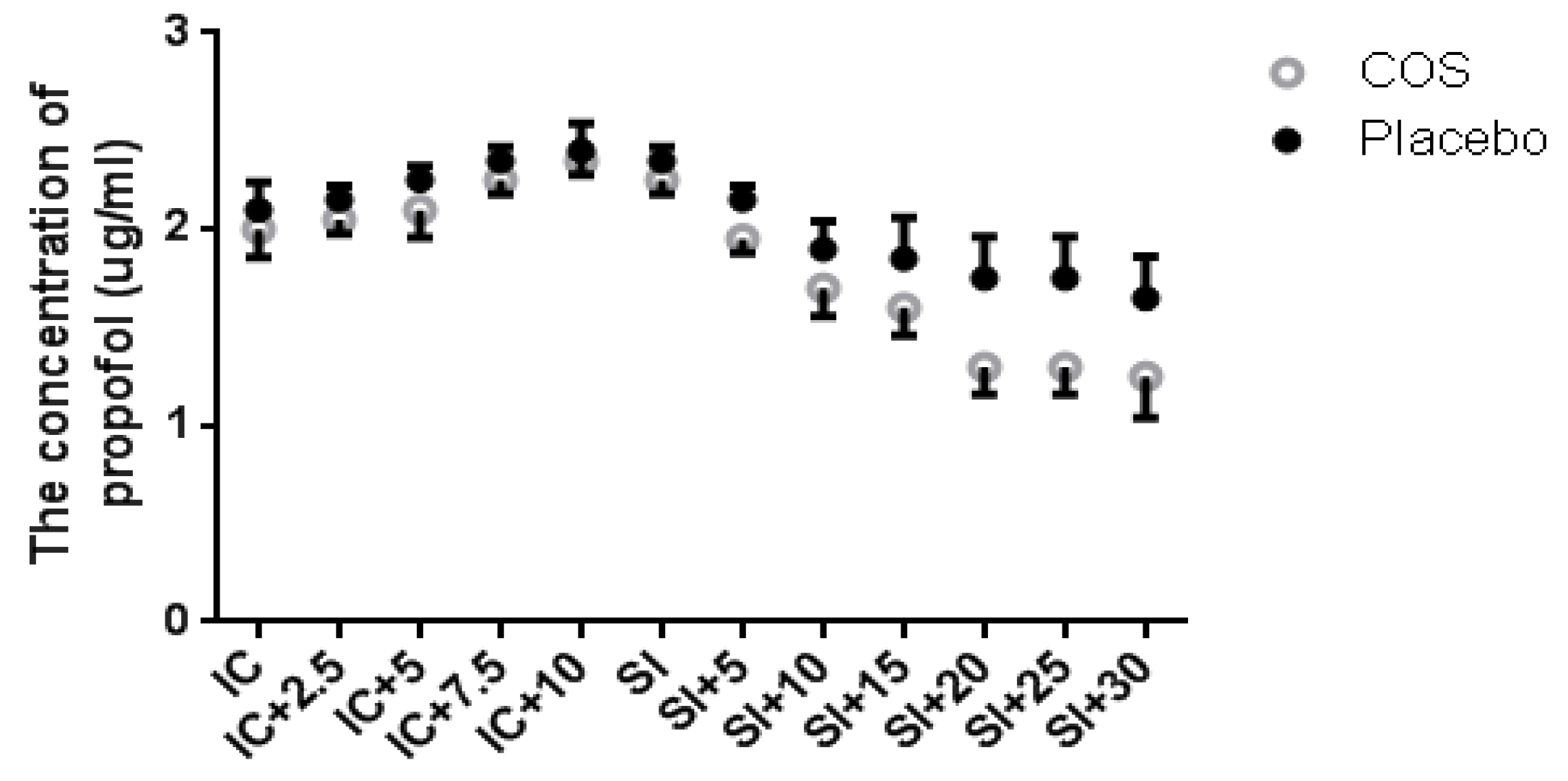
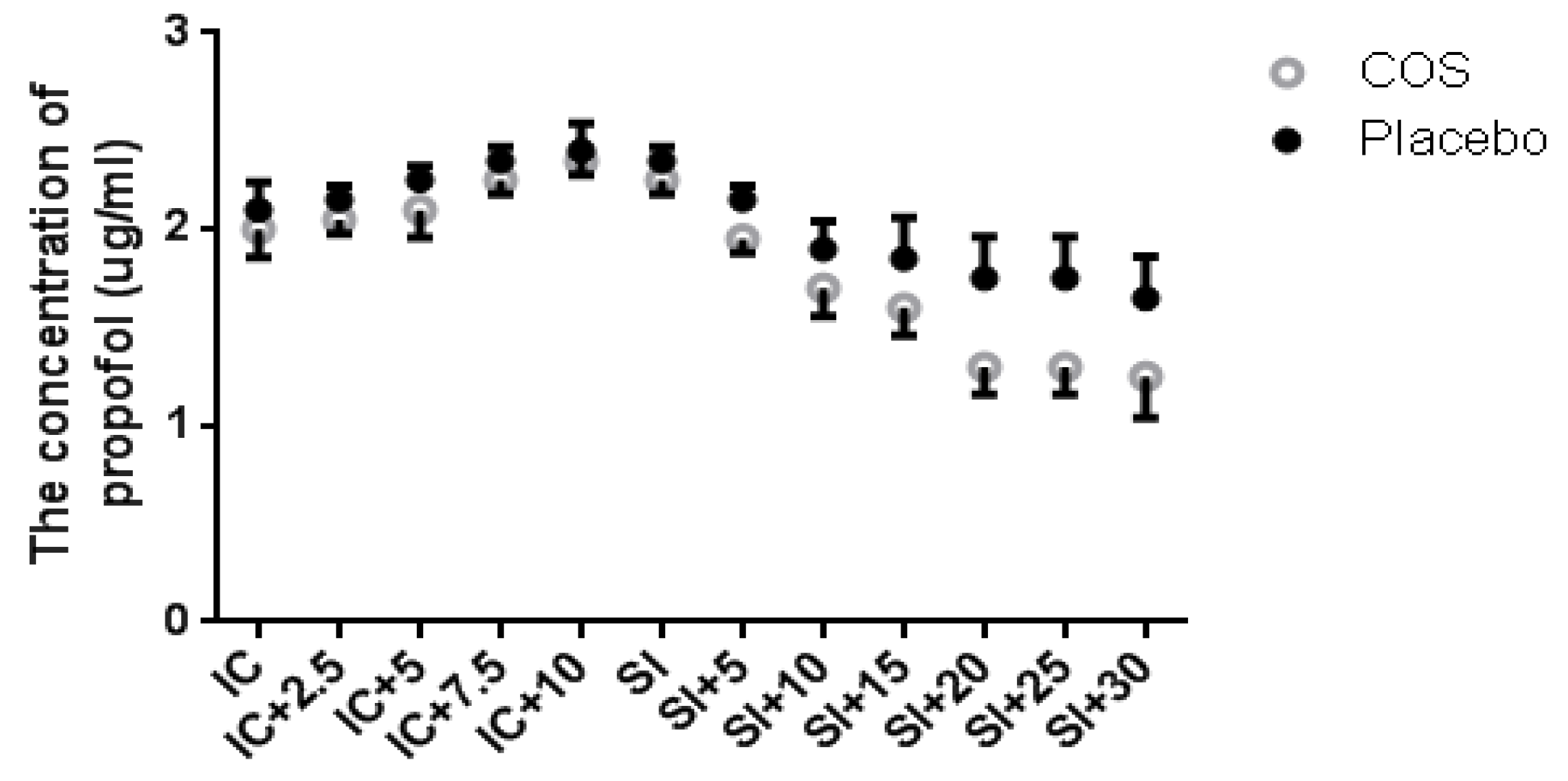
2.1. Chitosan Oligosaccharide (COS) Pretreatment Reduces Propofol Dose during Anesthesia
2.2. The Incidence of Propofol-Induced Injection Pain in the Subjects Undergoing Surgery
2.3. COS Pretreatment Reduces the Side Effects of Propofol
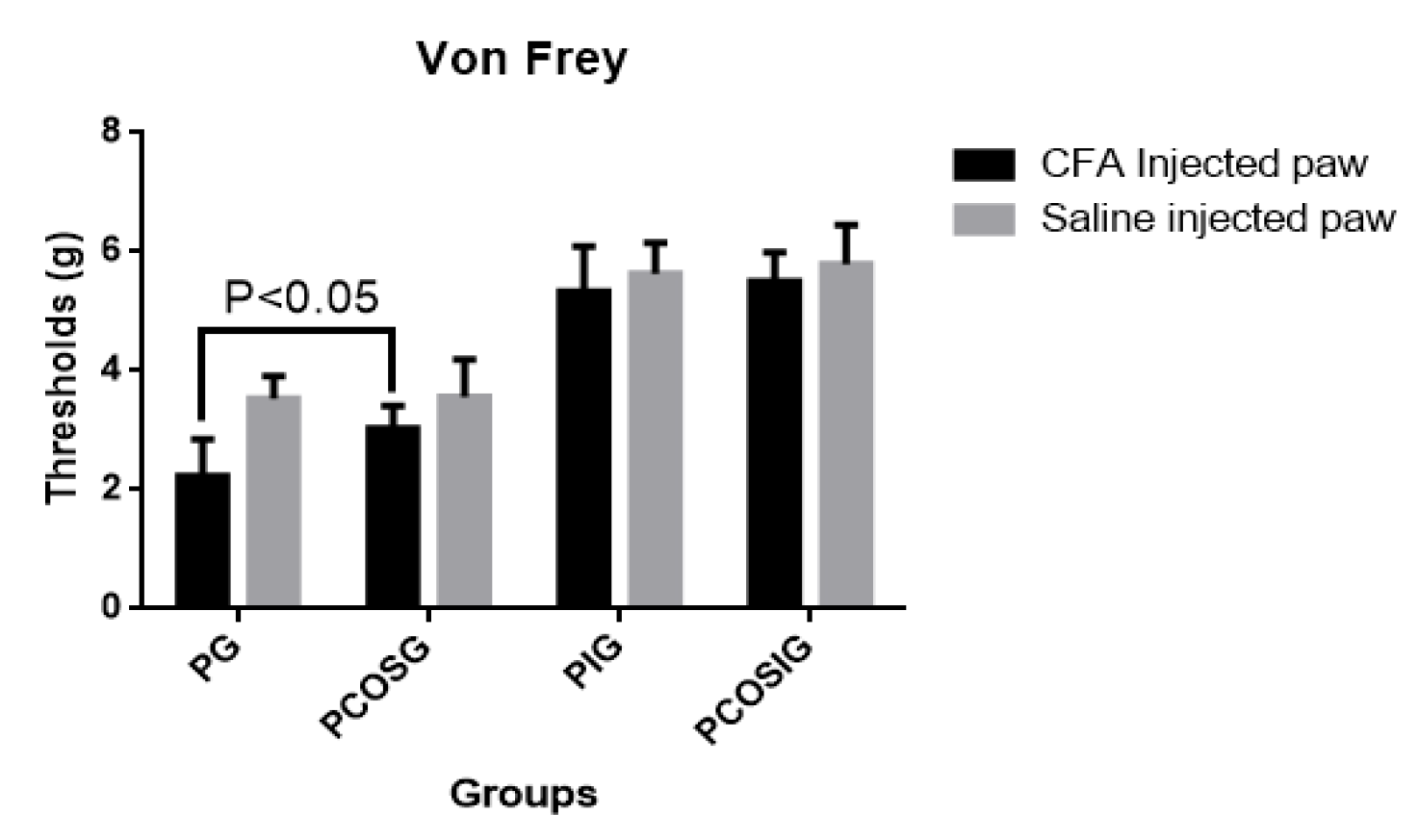
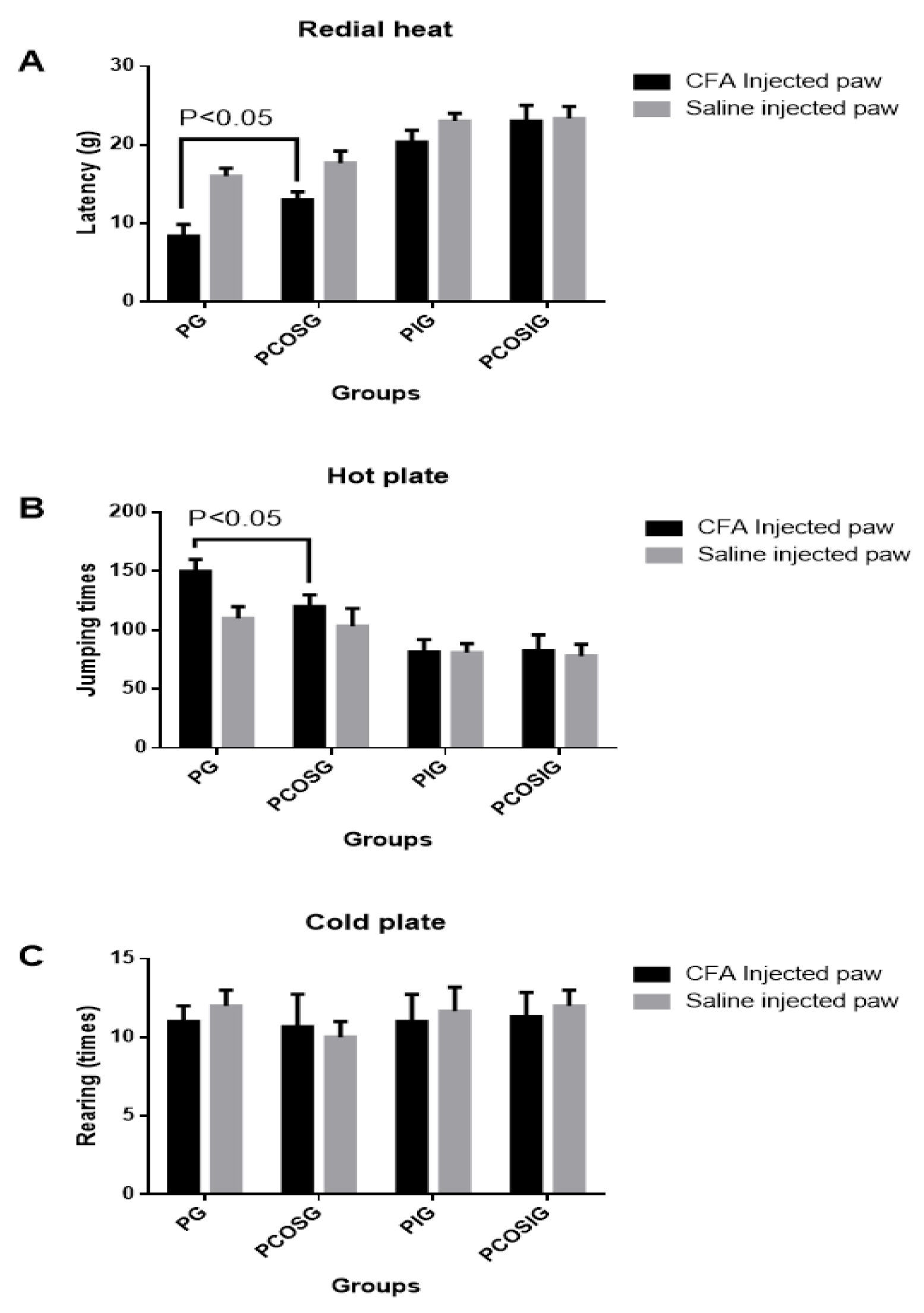
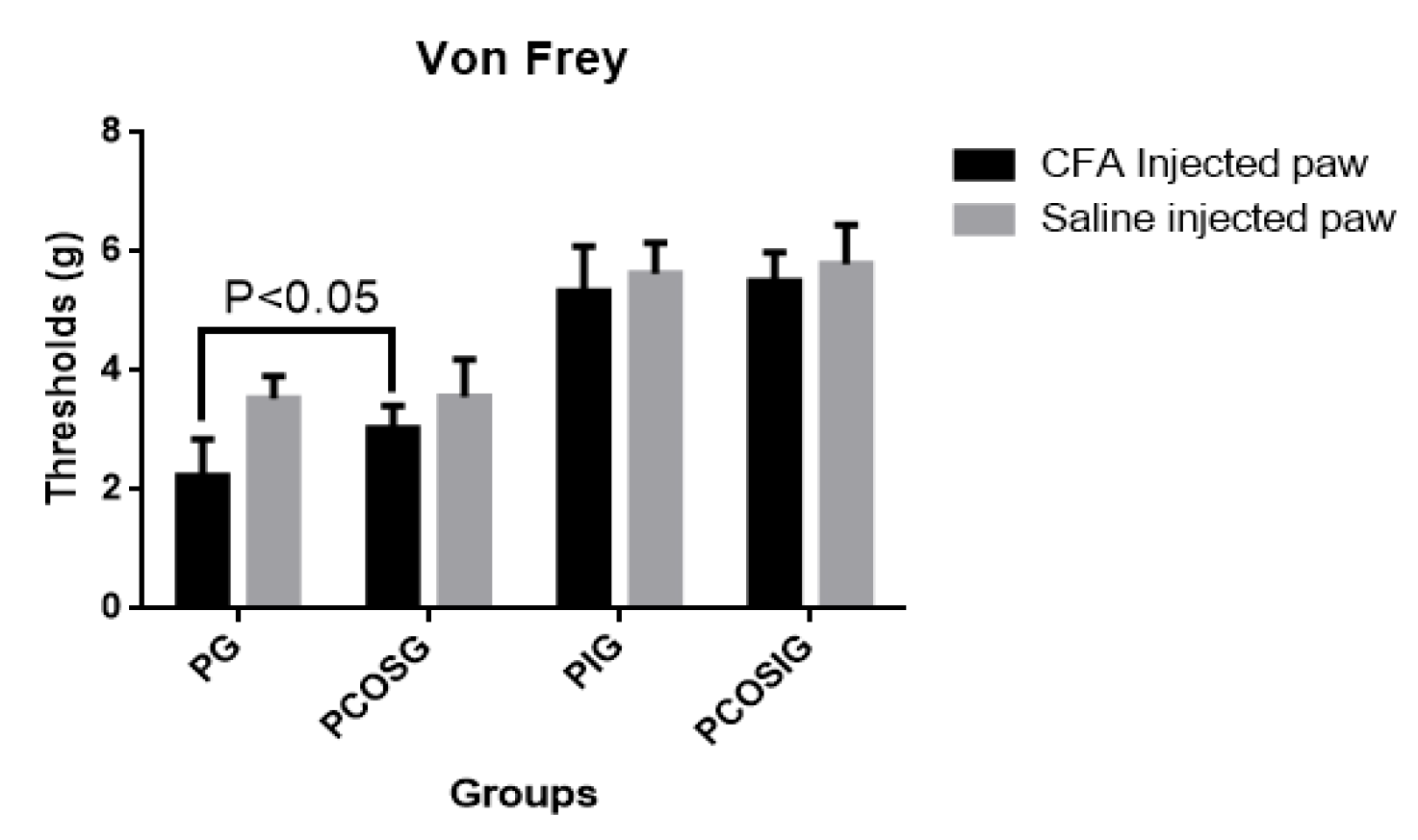
2.4. Analysis of Mechanic Hyperalgesia
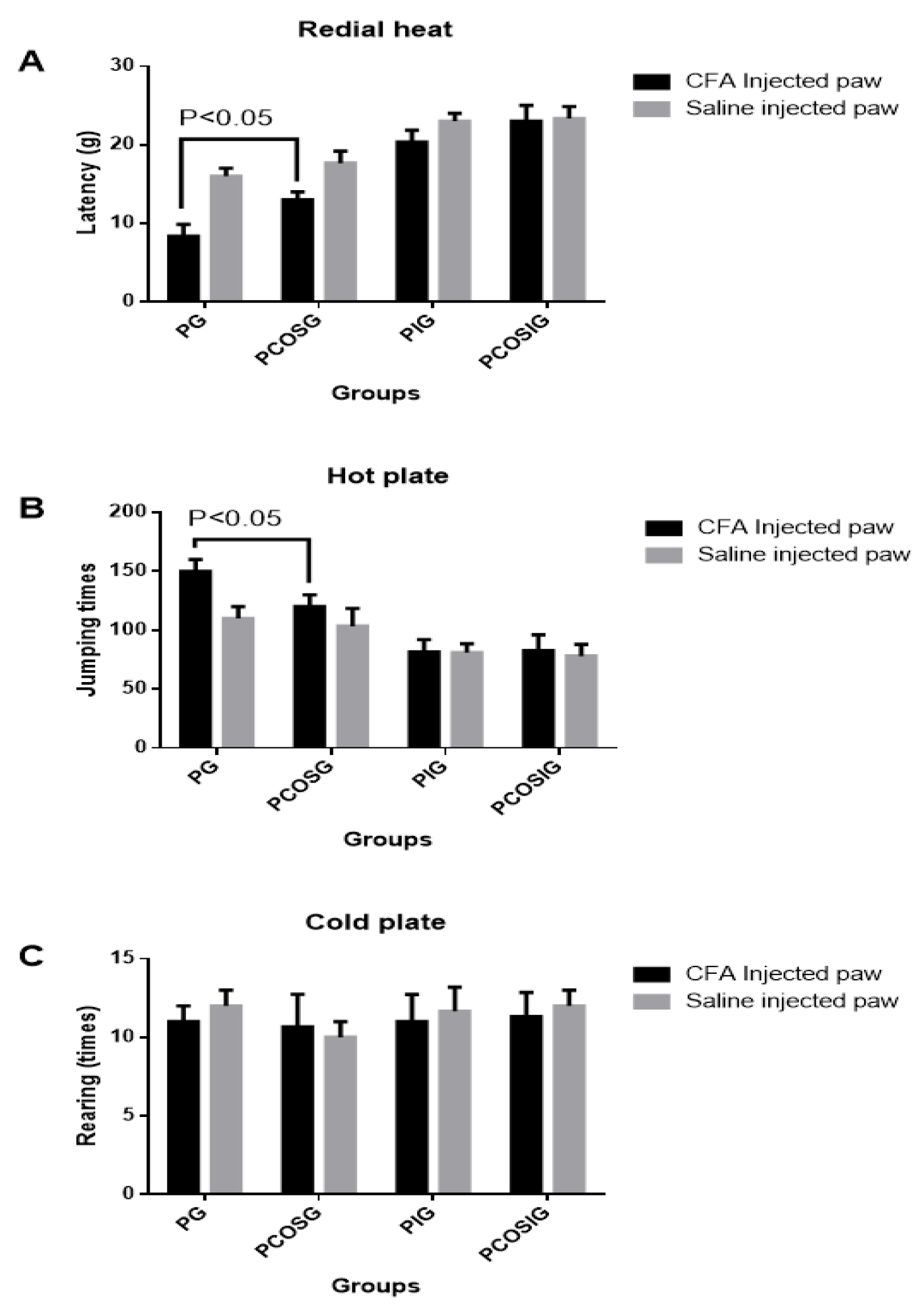
2.5. Analysis of Thermal Hyperalgesia
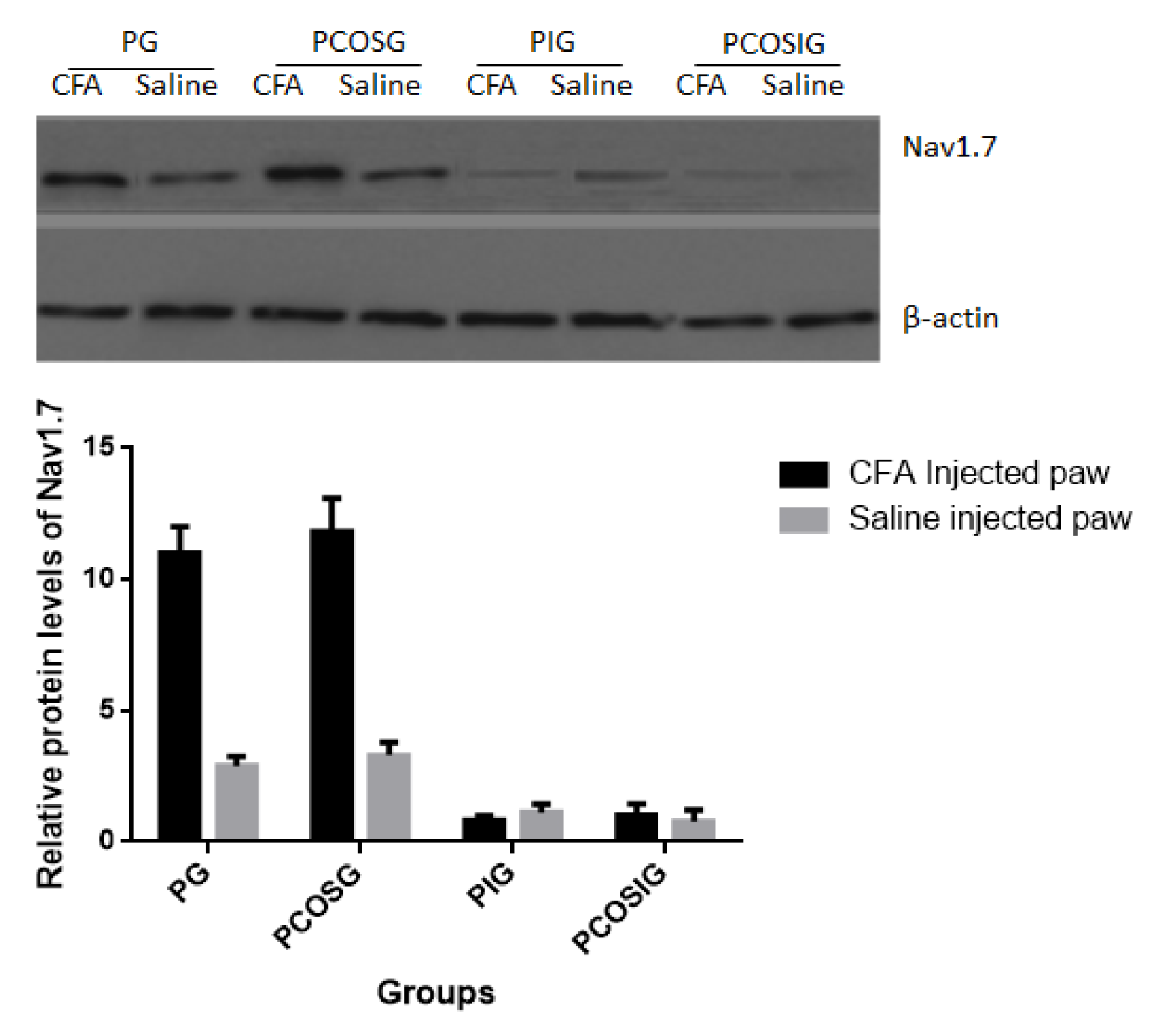
2.6. The Protein Level of Voltage-Gated Sodium Channels (Nav)1.7 in Dorsal Root Ganglia (DRG) Neurons
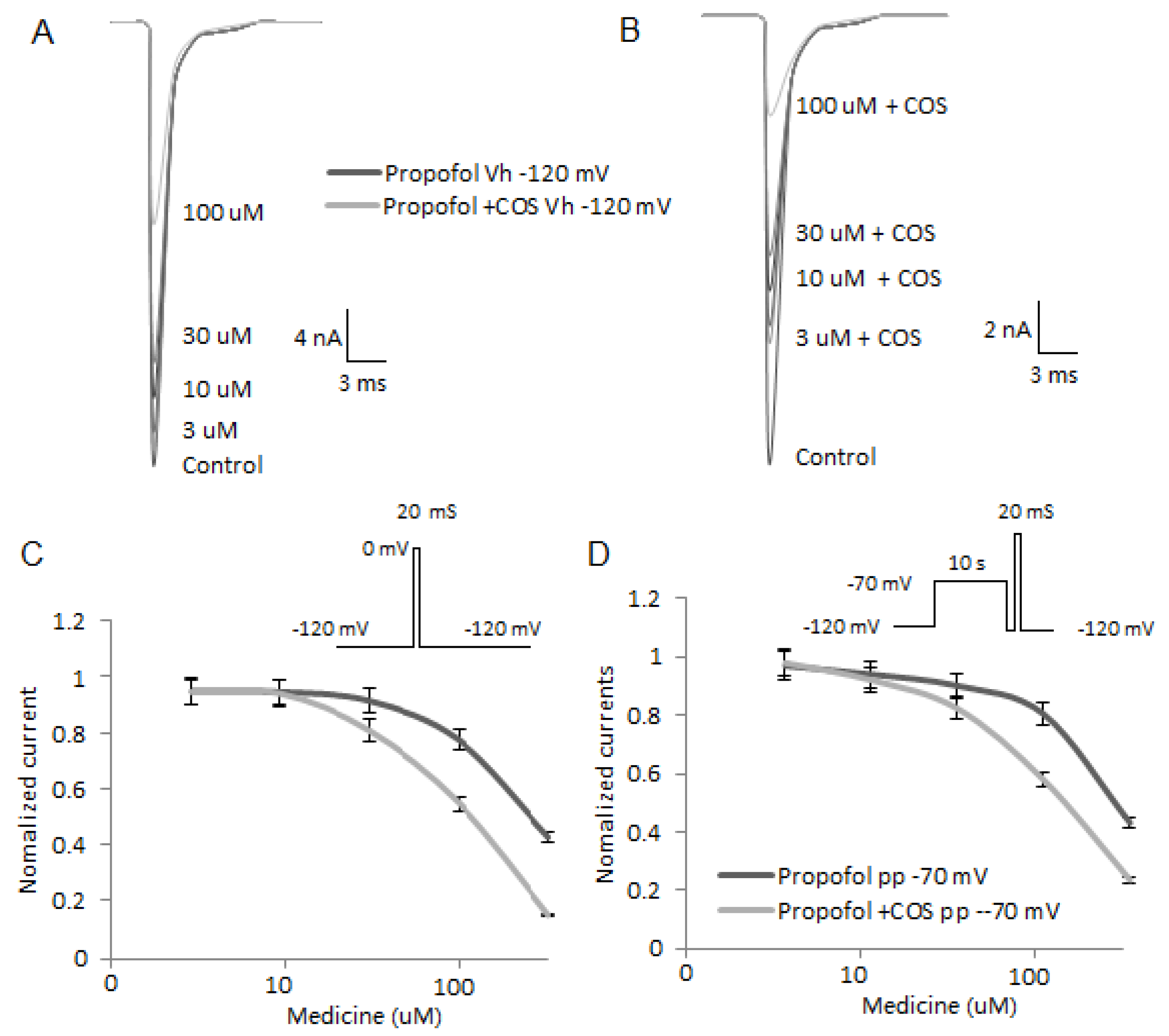
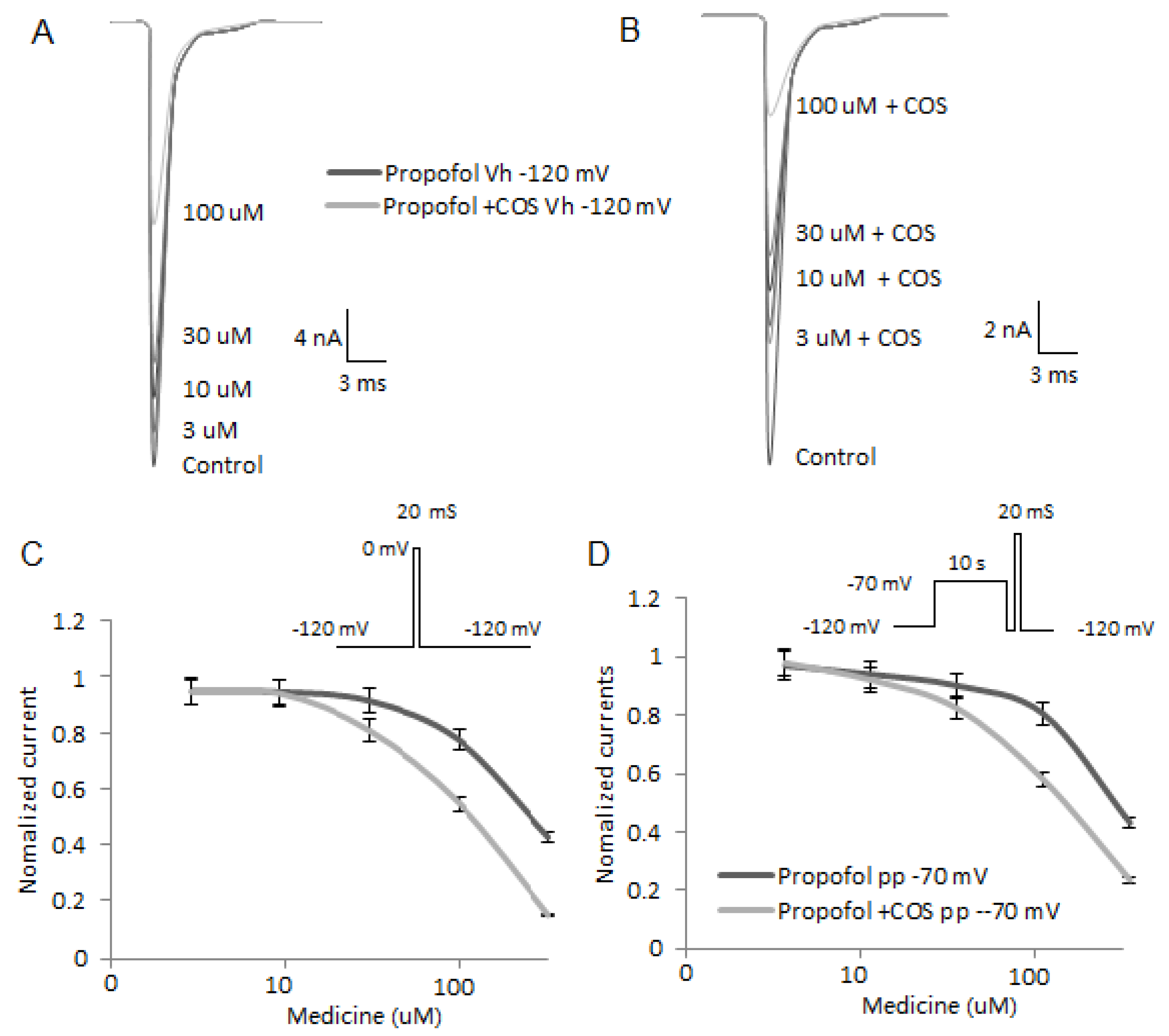
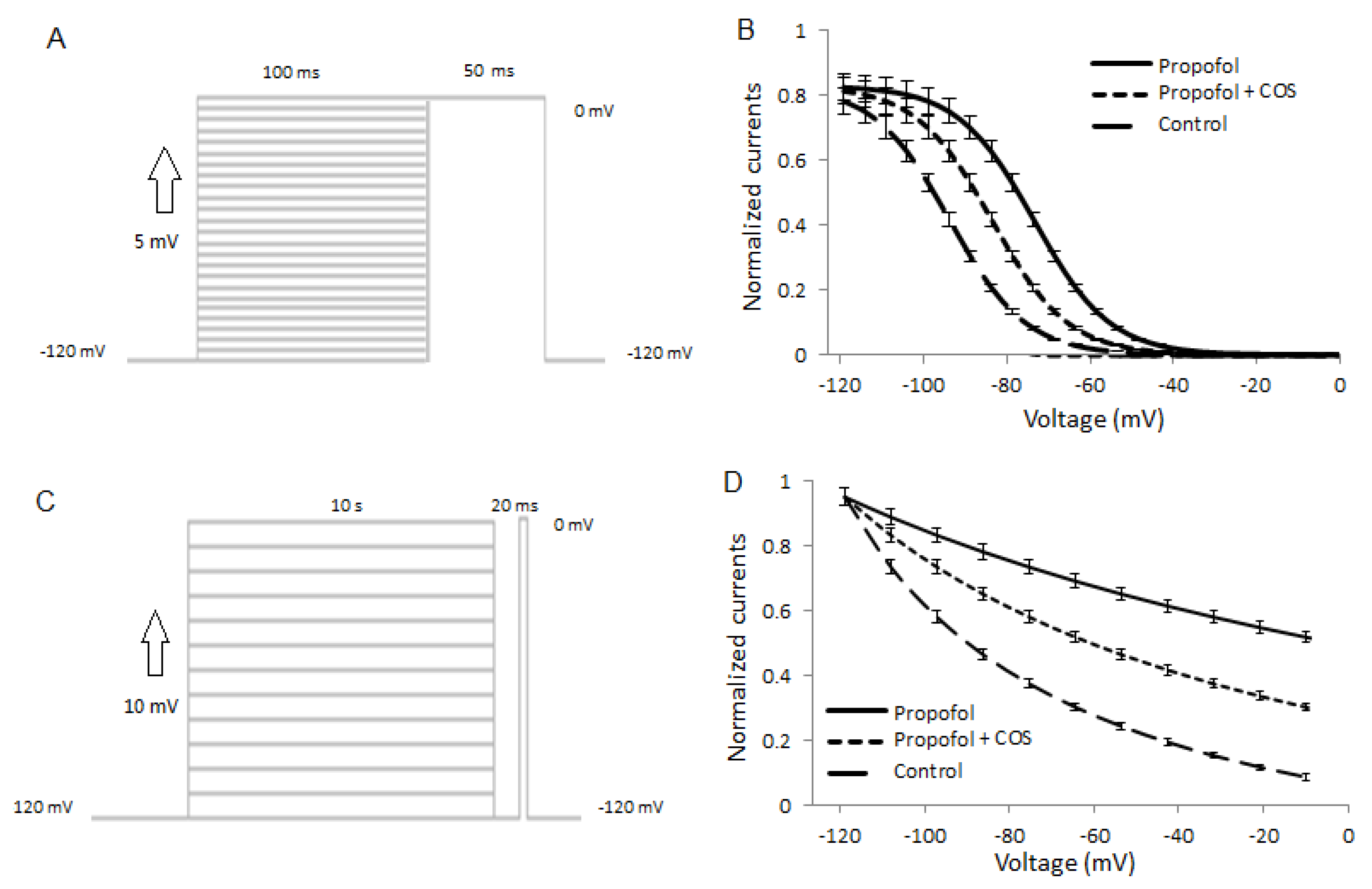
2.7. COS Reduces the Activity of Nav1.7
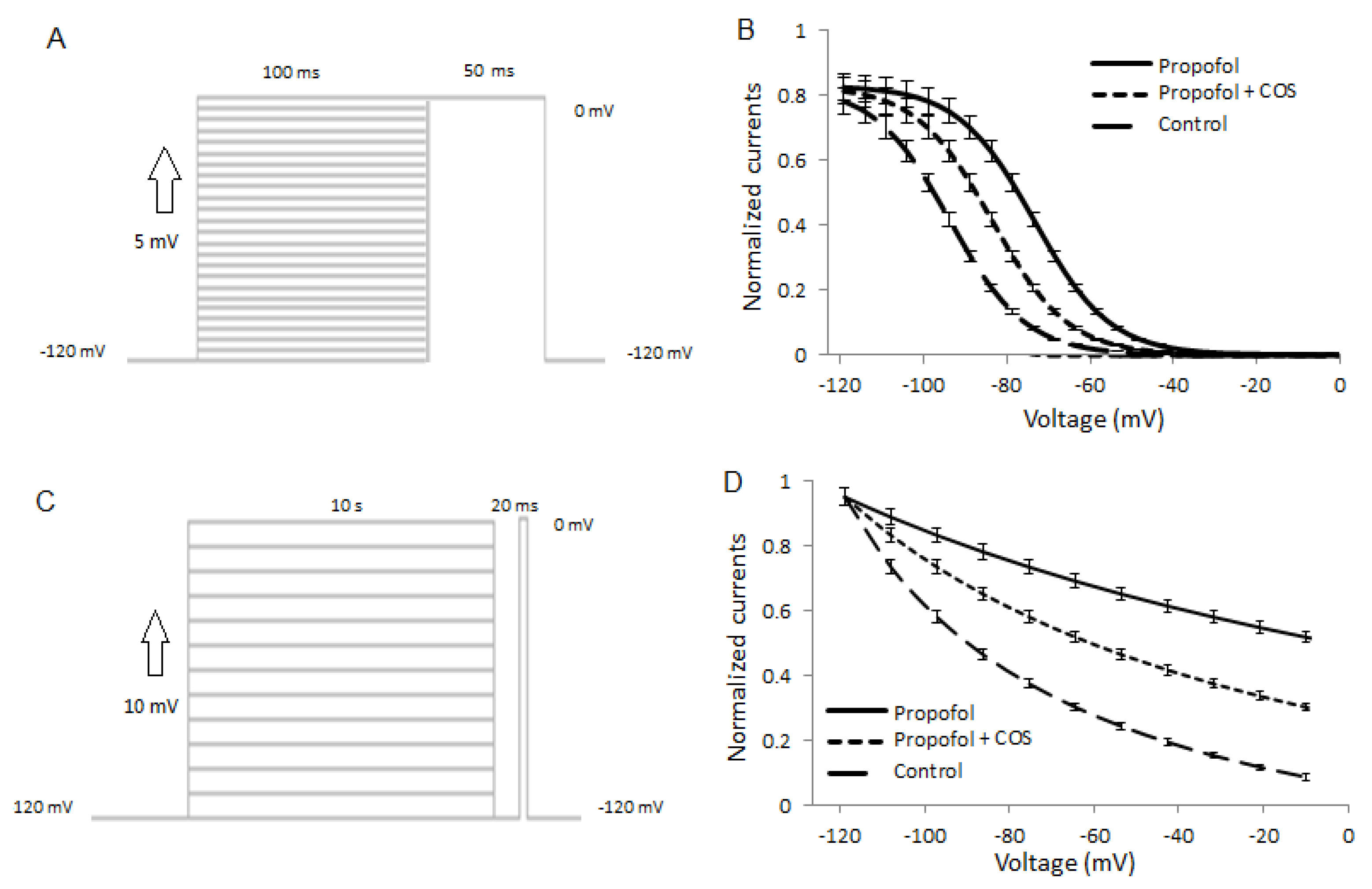
2.8. COS Also Promotes Propofol-Produced Stabilization of Fast and Slow Inactivation
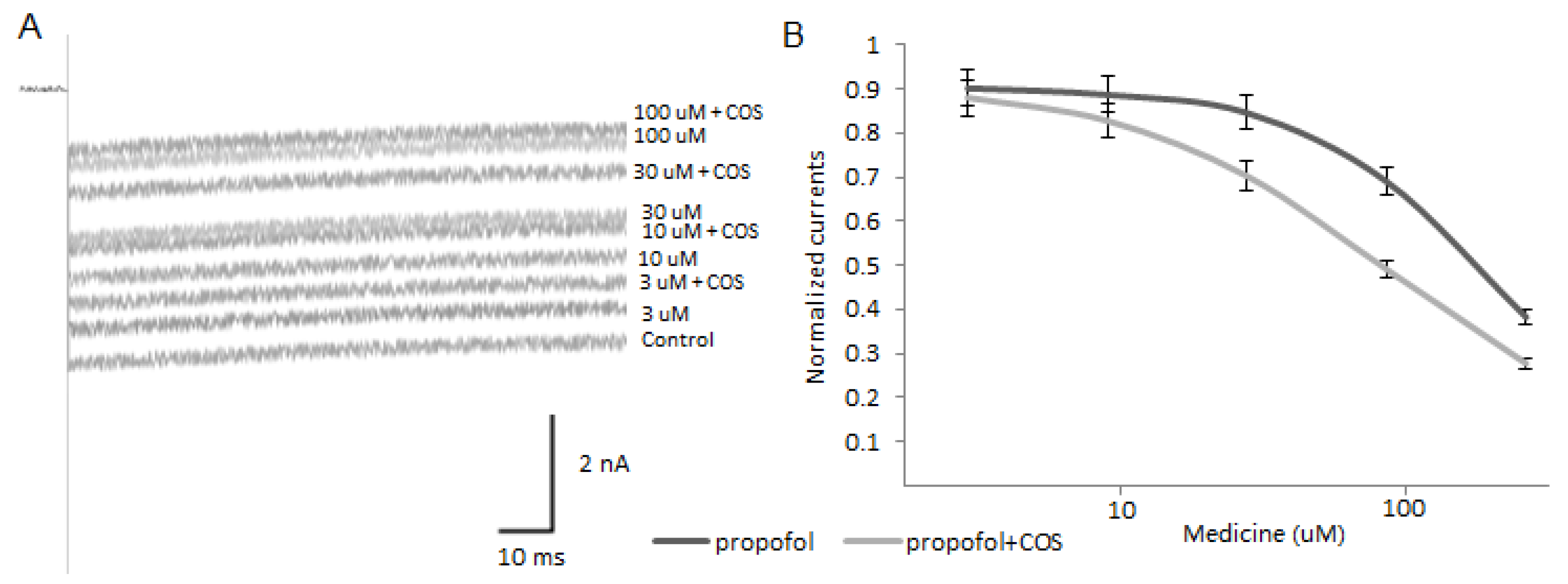
2.9. COS Promotes Propofol Blocking Veratridine-Induced Persistent Sodium Current of Nav1.7
3. Discussion
4. Materials and Methods
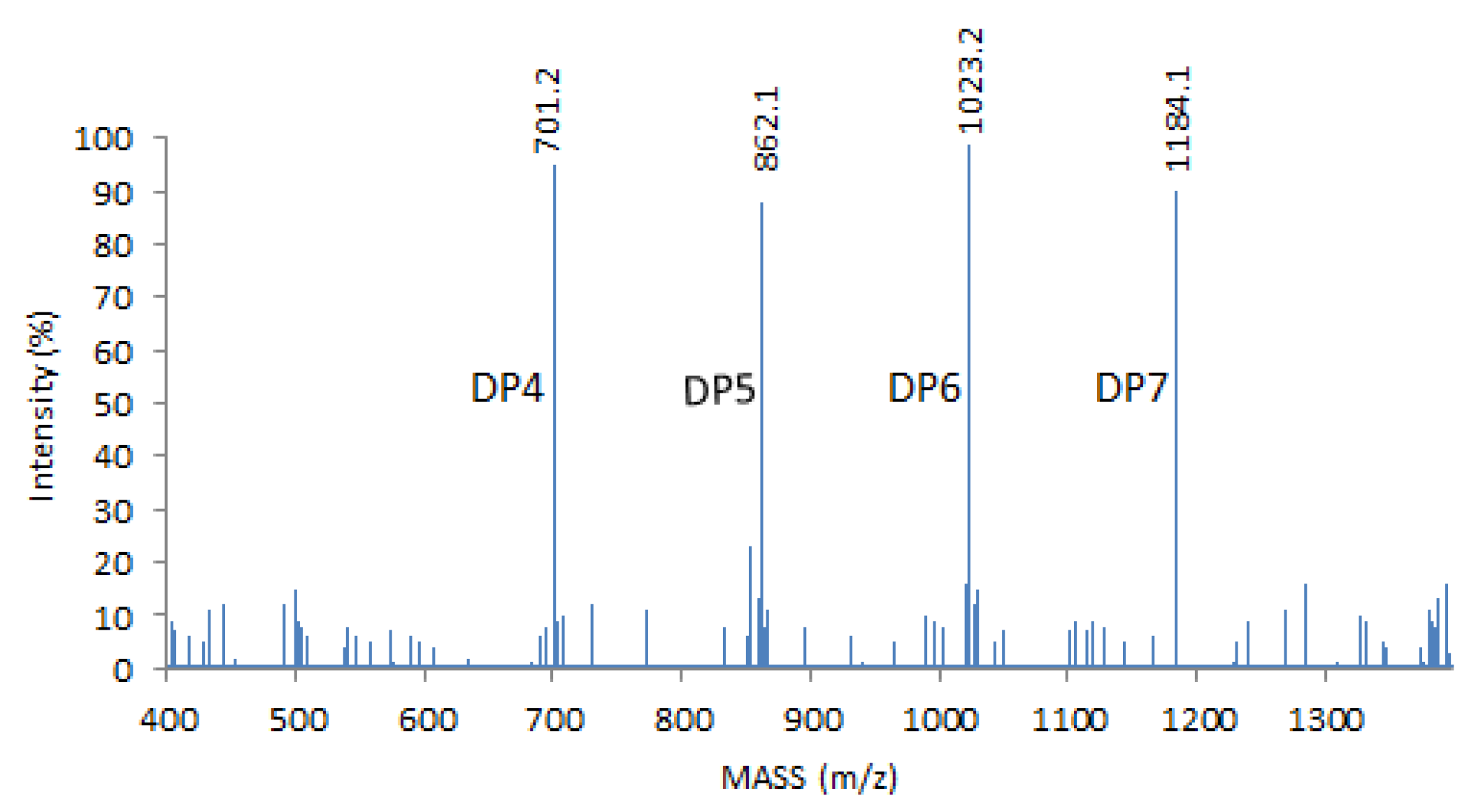
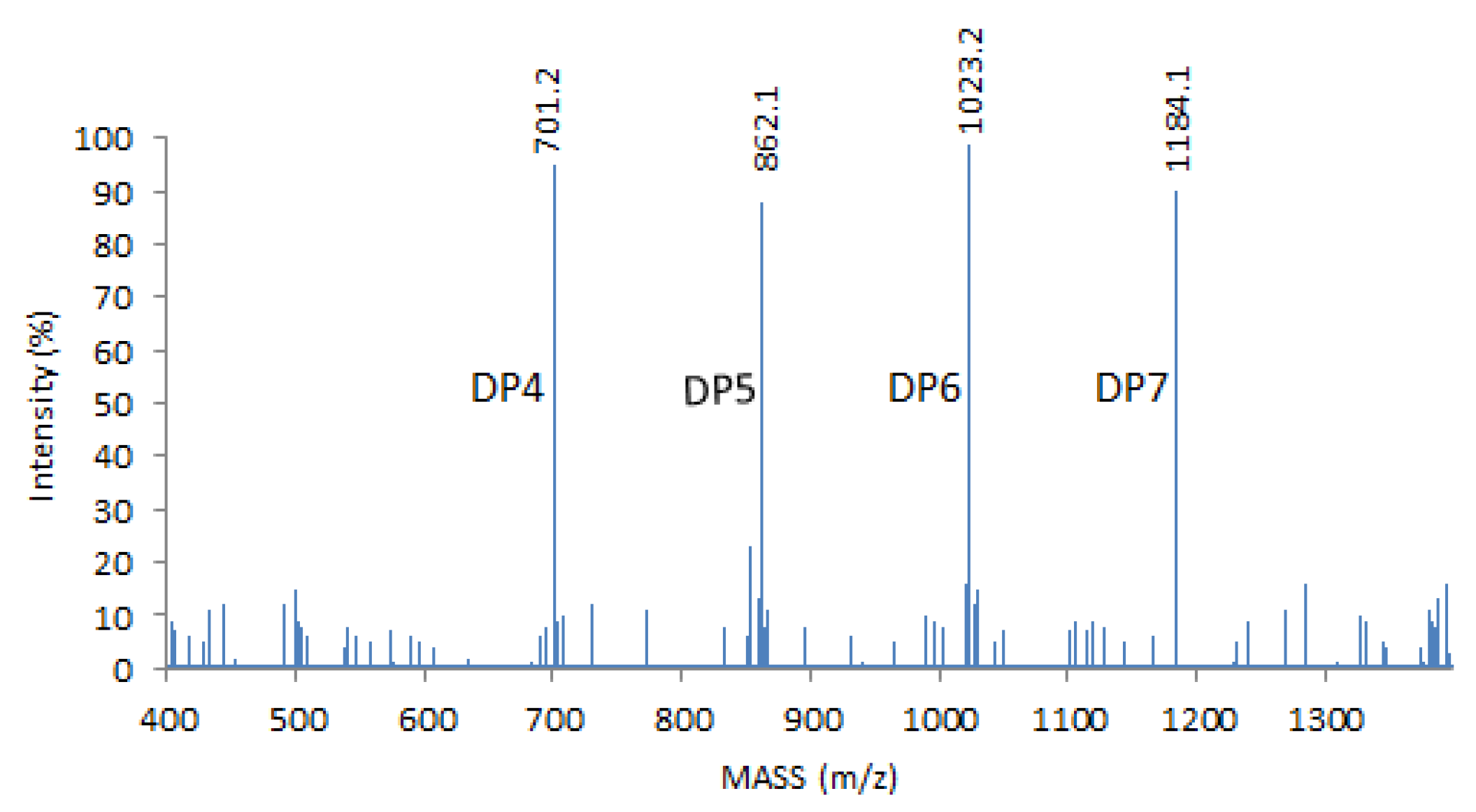
4.1. COS Preparation and MALDI–TOF (Matrix-Assisted Laser-Desorption Ionization–Time-of-Flight) MS Analysis
4.2. Participants
4.3. Patient Grouping
4.4. Animals
4.5. Nav1.7 Gene Silencing
4.6. Animal Grouping
4.7. Animal Behavior of Mechanical and Thermal Hyperalgesia
4.8. Western Blot
4.9. Electrophysiology Analysis of Nav1.7
4.10. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Sapate, M.; Andurkar, U.; Markandeya, M.; Gore, R.; Thatte, W. To study the effect of injection dexmedetomidine for prevention of pain due to propofol injection and to compare it with injection lignocaine. Braz. J. Anesthesiol. 2015, 65, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Marik, P.E. Propofol: Therapeutic indications and side-effects. Curr. Pharm. Des. 2004, 10, 3639–3649. [Google Scholar] [CrossRef] [PubMed]
- Mays, N. Reducing unwarranted variations in healthcare in the English NHS. BMJ 2011, 342, d1849. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, S.E.; Chung, S.; Lee, H.J.; Jeong, S. Impact of time interval between remifentanil and propofol on propofol injection pain. J. Clin. Anesth. 2016, 34, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Madan, H.K.; Singh, R.; Sodhi, G.S. Comparsion of Intravenous Lignocaine, Tramadol and Keterolac for Attenuation of Propofol Injection Pain. J. Clin. Diagn. Res. 2016, 10, UC05–UC08. [Google Scholar] [CrossRef] [PubMed]
- Berberian, P.; Obimba, C.; Glickman-Simon, R.; Sethi, T. Herbs for Low-Back Pain, Acupuncture for Psychological Distress, Osteopathic Manipulative Therapy for Chronic Migraine, Honey Dressings for Burns, Vegetarian Diet and Risk of Colorectal Cancer. Explore 2015, 11, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Schroder, S.; Beckmann, K.; Franconi, G.; Meyer-Hamme, G.; Friedemann, T.; Greten, H.J.; Rostock, M.; Efferth, T. Can medical herbs stimulate regeneration or neuroprotection and treat neuropathic pain in chemotherapy-induced peripheral neuropathy? Evid. Based Complement. Altern. Med. 2013, 2013, 423713. [Google Scholar] [CrossRef] [PubMed]
- Tatsumi, S.; Mabuchi, T.; Abe, T.; Xu, L.; Minami, T.; Ito, S. Analgesic effect of extracts of Chinese medicinal herbs Moutan cortex and Coicis semen on neuropathic pain in mice. Neurosci. Lett. 2004, 370, 130–134. [Google Scholar] [CrossRef] [PubMed]
- Euasobhon, P.; Dej-Arkom, S.; Siriussawakul, A.; Muangman, S.; Sriraj, W.; Pattanittum, P.; Lumbiganon, P. Lidocaine for reducing propofol-induced pain on induction of anaesthesia in adults. Cochrane Database Syst. Rev. 2016. [Google Scholar] [CrossRef]
- Joo, J.D.; In, J.H.; Kim, D.W.; Jung, H.S.; Kang, J.H.; Yeom, J.H.; Choi, J.W. The comparison of sedation quality, side effect and recovery profiles on different dosage of remifentanil patient-controlled sedation during breast biopsy surgery. Korean J. Anesthesiol. 2012, 63, 431–435. [Google Scholar] [CrossRef] [PubMed]
- McCleskey, P.E.; Patel, S.M.; Mansalis, K.A.; Elam, A.L.; Kinsley, T.R. Serum lidocaine levels and cutaneous side effects after application of 23% lidocaine 7% tetracaine ointment to the face. Dermatol. Surg. 2013, 39, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Romanazzi, G.; Feliziani, E.; Banos, S.B.; Sivakumar, D. Shelf life extension of fresh fruit and vegetables by chitosan treatment. Crit. Rev. Food Sci. Nutr. 2017, 57, 579–601. [Google Scholar] [CrossRef] [PubMed]
- Swiatkiewicz, S.; Swiatkiewicz, M.; Arczewska-Wlosek, A.; Jozefiak, D. Chitosan and its oligosaccharide derivatives (chito-oligosaccharides) as feed supplements in poultry and swine nutrition. J. Anim. Physiol. Anim. Nutr. (Berl.) 2015, 99, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.J.; Park, J.K.; Park, Y.I. Anti-inflammatory effects of low-molecular weight chitosan oligosaccharides in IgE–antigen complex-stimulated RBL-2H3 cells and asthma model mice. Int. Immunopharmacol. 2012, 12, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Ma, Y.; Wang, C.; Liu, H.; Li, Q.; Fei, M. Synthesis, anti-oxidant activity, and biodegradability of a novel recombinant polysaccharide derived from chitosan and lactose. Carbohydr. Polym. 2015, 118, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, I.; Mengíbar, M.; Harris, R.; Paños, I.; Miralles, B.; Acosta, N.; Galed, G.; Heras, Á. Functional characterization of chitin and chitosan. Curr. Chem. Biol. 2009, 3, 203–230. [Google Scholar] [CrossRef]
- Lai, J.; Porreca, F.; Hunter, J.C.; Gold, M.S. Voltage-gated sodium channels and hyperalgesia. Annu. Rev. Pharmacol. Toxicol. 2004, 44, 371–397. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.S.; Stevens, E.B.; Pinnock, R.D.; Dixon, A.K.; Lee, K. Developmental expression of the novel voltage-gated sodium channel auxiliary subunit β3, in rat CNS. J. Physiol. 2001, 534, 763–776. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, W.; Faull, R.; Waldvogel, H.; Plumpton, C.; Burbidge, S.; Emson, P.; Clare, J. Localization of the type VI voltage-gated sodium channel protein in human CNS. Neuroreport 1999, 10, 3703–3709. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Liu, D.; Chhoa, M.; Li, C.M.; Luo, Y.; Zhang, M.; Lehto, S.G.; Immke, D.C.; Moyer, B.D. Voltage-gated sodium channel function and expression in injured and uninjured rat dorsal root ganglia neurons. Int. J. Neurosci. 2015, 126, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Rabert, D.K.; Koch, B.D.; Ilnicka, M.; Obernolte, R.A.; Naylor, S.L.; Herman, R.C.; Eglen, R.M.; Hunter, J.C.; Sangameswaran, L. A tetrodotoxin-resistant voltage-gated sodium channel from human dorsal root ganglia, hPN3/SCN10A. Pain 1998, 78, 107–114. [Google Scholar] [CrossRef]
- Cohen, C.J. Targeting voltage-gated sodium channels for treating neuropathic and inflammatory pain. Curr. Pharm. Biotechnol. 2011, 12, 1715–1719. [Google Scholar] [CrossRef] [PubMed]
- Suh, H.R.; Chung, H.J.; Park, E.H.; Moon, S.W.; Park, S.J.; Park, C.W.; Kim, Y.I.; Han, H.C. The effects of Chamaecyparis obtusa essential oil on pain-related behavior and expression of pro-inflammatory cytokines in carrageenan-induced arthritis in rats. Biosci. Biotechnol. Biochem. 2015, 80, 203–209. [Google Scholar] [PubMed]
- Yang, Y.; Li, Y.X.; Wang, H.L.; Jin, S.J.; Zhou, R.; Qiao, H.Q.; Du, J.; Wu, J.; Zhao, C.J.; Niu, Y.; et al. Oxysophocarpine Ameliorates Carrageenan-induced Inflammatory Pain via Inhibiting Expressions of Prostaglandin E2 and Cytokines in Mice. Planta Med. 2015, 81, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.L.; He, X.F.; Shen, Y.F.; Yin, X.H.; Du, J.Y.; Liang, Y.I.; Fang, J.Q. Analgesic roles of peripheral intrinsic met-enkephalin and dynorphin A in long-lasting inflammatory pain induced by complete Freund’s adjuvant in rats. Exp. Ther. Med. 2015, 9, 2344–2348. [Google Scholar] [CrossRef] [PubMed]
- Qian, B.; Li, F.; Zhao, L.X.; Dong, Y.L.; Gao, Y.J.; Zhang, Z.J. Ligustilide Ameliorates Inflammatory Pain and Inhibits TLR4 Upregulation in Spinal Astrocytes Following Complete Freund’s Adjuvant Peripheral Injection. Cell. Mol. Neurobiol. 2015, 36, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Strickland, I.T.; Martindale, J.C.; Woodhams, P.L.; Reeve, A.J.; Chessell, I.P.; McQueen, D.S. Changes in the expression of NaV1.7, NaV1.8 and NaV1.9 in a distinct population of dorsal root ganglia innervating the rat knee joint in a model of chronic inflammatory joint pain. Eur. J. Pain 2008, 12, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Leo, S.; D’Hooge, R.; Meert, T. Exploring the role of nociceptor-specific sodium channels in pain transmission using Nav1.8 and Nav1.9 knockout mice. Behav. Brain Res. 2010, 208, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Nassar, M.A.; Stirling, L.C.; Forlani, G.; Baker, M.D.; Matthews, E.A.; Dickenson, A.H.; Wood, J.N. Nociceptor-specific gene deletion reveals a major role for Nav1.7 (PN1) in acute and inflammatory pain. Proc. Natl. Acad. Sci. USA 2004, 101, 12706–12711. [Google Scholar] [CrossRef] [PubMed]
- Shields, S.D.; Cheng, X.; Üçeyler, N.; Sommer, C.; Dib-Hajj, S.D.; Waxman, S.G. Sodium channel Nav1.7 is essential for lowering heat pain threshold after burn injury. J. Neurosci. 2012, 32, 10819–10832. [Google Scholar] [CrossRef] [PubMed]
- De Rooij, A.M.; Gosso, M.F.; Alsina-Sanchis, E.; Marinus, J.; Hilten, J.J.V.; Maagdenberg, A.M.V.D. No mutations in the voltage-gated NaV1.7 sodium channel alpha1 subunit gene SCN9A in familial complex regional pain syndrome. Eur. J. Neurol. 2010, 17, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Diss, J.K.; Calissano, M.; Gascoyne, D.; Djamgoz, M.B.; Latchman, D.S. Identification and characterization of the promoter region of the Nav1.7 voltage-gated sodium channel gene (SCN9A). Mol. Cell. Neurosci. 2008, 37, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.J.; Reimann, F.; Nicholas, A.K.; Thornton, G.; Roberts, E.; Springell, K.; Karbani, G.; Jafri, H.; Mannan, J.; Raashid, Y.; et al. An SCN9A channelopathy causes congenital inability to experience pain. Nature 2006, 444, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Cao, J.; Ren, X.; Zang, W. Nav1.7 protein and mRNA expression in the dorsal root ganglia of rats with chronic neuropathic pain. Neural Regen. Res. 2012, 7, 1540–1544. [Google Scholar] [PubMed]
- Minett, M.S.; Pereira, V.; Sikandar, S.; Matsuyama, A.; Lolignier, S.; Kanellopoulos, A.H.; Mancini, F.; Iannetti, G.D.; Bogdanov, Y.D.; Santana-Varela, S.; et al. Endogenous opioids contribute to insensitivity to pain in humans and mice lacking sodium channel Nav1.7. Nat. Commun. 2015, 6, 8967. [Google Scholar] [CrossRef] [PubMed]
- Gandini, R.; Merolla, S.; Chegai, F.; Del Giudice, C.; Stefanini, M.; Pampana, E. Foot Embolization During Limb Salvage Procedures in Critical Limb Ischemia Patients Successfully Managed With Mechanical Thromboaspiration: A Technical Note. J. Endovasc. Ther. 2015, 22, 558–563. [Google Scholar] [CrossRef] [PubMed]
- SudheesháKumar, P. Flexible, micro-porous chitosan–gelatin hydrogel/nanofibrin composite bandages for treating burn wounds. RSC Adv. 2014, 4, 65081–65087. [Google Scholar]
- Capasso, R.; Rosa, T.; Tsou, D.Y.; Nekhendzy, V.; Drover, D.; Collins, J.; Zaghi, S.; Camacho, M. Variable Findings for Drug-Induced Sleep Endoscopy in Obstructive Sleep Apnea with Propofol versus Dexmedetomidine. Otolaryngol. Head Neck Surg. 2016, 154, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Au, A.K.; Steinberg, D.; Thom, C.; Shirazi, M.; Papanagnou, D.; Ku, B.S.; Fields, J.M. Ultrasound measurement of inferior vena cava collapse predicts propofol-induced hypotension. Am. J. Emerg. Med. 2016, 34, 1125–1128. [Google Scholar] [CrossRef] [PubMed]
- Bang, Y.S.; Kim, Y.U.; Oh, D.; Shin, E.Y.; Park, S.K. A randomized, double-blind trial evaluating the efficacy of palonosetron with total intravenous anesthesia using propofol and remifentanil for the prevention of postoperative nausea and vomiting after gynecologic surgery. J. Anesth. 2016, 30, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Bataille, A.; Letourneulx, J.F.; Charmeau, A.; Lemedioni, P.; Leger, P.; Chazot, T.; Guen, M.L.; Diemunsch, P.; Fischler, M.; Liu, N. Impact of prophylactic combination of dexamethasone-ondansetron on postoperative nausea and vomiting in obese adult patients undergoing laparoscopic sleeve gastrectomy during closed-loop propofol-remifentanil anaesthesia: A randomised double-blind placebo study. Eur. J. Anaesthesiol. 2016, 33, 898–905. [Google Scholar] [PubMed]
- Cho, S.Y.; Jeong, C.W.; Jeong, C.Y.; Lee, H.G. Efficacy of the combination of cold propofol and pretreatment with remifentail on propofol injection pain. Korean J. Anesthesiol. 2010, 59, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Y.; Li, B.; Zhang, W. Remifentail infusion for paediatric bronchoscopic foreign body removal: Comparison of sevoflurane with propofol for anaesthesia supplementation for bronchoscope insertion. Anaesth. Intensive Care 2010, 38, 905–910. [Google Scholar] [PubMed]
- Rivara, M.; Zuliani, V. Novel sodium channel antagonists in the treatment of neuropathic pain. Expert Opin. Investig. Drugs 2015, 25, 215–226. [Google Scholar] [CrossRef] [PubMed]
- Hockley, J.R.; Winchester, W.J.; Bulmer, D.C. The voltage-gated sodium channel Na 1.9 in visceral pain. Neurogastroenterol. Motil. 2016, 28, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Mackenzie, F.E.; Parker, A.; Parkinson, N.J.; Oliver, P.L.; Brooker, D.; Underhill, P.; Lukashkina, V.A.; Lukashkin, A.N.; Holmes, C.; Brown, S.D. Analysis of the mouse mutant Cloth-ears shows a role for the voltage-gated sodium channel Scn8a in peripheral neural hearing loss. Genes Brain Behav. 2009, 8, 699–713. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, C.E.; Baracat, J.S.; Arantes, E.C.; de Nucci, G.; Antunes, E. Effects of β-adrenoceptor antagonists in the neural nitric oxide release induced by electrical field stimulation and sodium channel activators in the rabbit corpus cavernosum. Eur. J. Pharmacol. 2005, 519, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.W.; Ho, G.D.; Tulshian, D.; Bercovici, A.; Tan, Z.; Hanisak, J.; Brumfield, S.; Matasi, J.; Sun, X.; Sakwa, S.A.; et al. Bioavailable pyrrolo-benzo-1,4-diazines as Nav 1.7 sodium channel blockers for the treatment of pain. Bioorg. Med. Chem. Lett. 2014, 24, 4958–4962. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-P.; Chen, H.-N.; Su, H.-L.; Hsieh, C.-L.; Chen, W.-H.; Lai, Z.-R.; Lin, Y.-W. Electroacupuncture reduces carrageenan-and CFA-induced inflammatory pain accompanied by changing the expression of Nav1.7 and Nav1.8, rather than Nav1.9, in mice dorsal root ganglia. Evid. Based Complement. Altern. Med. 2013. [Google Scholar] [CrossRef] [PubMed]
- Liang, L.; Fan, L.; Tao, B.; Yaster, M.; Tao, Y.-X. Protein kinase B/Akt is required for complete Freund’s adjuvant-induced upregulation of Nav1.7 and Nav1.8 in primary sensory neurons. J. Pain 2013, 14, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, M.; Mata, M.; Fink, D.J. Continuous delta-opioid receptor activation reduces neuronal voltage-gated sodium channel (NaV1.7) levels through activation of protein kinase C in painful diabetic neuropathy. J. Neurosci. 2008, 28, 6652–6658. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Pei, Q.; Cao, L.; Xu, L.; Zhang, B.; Liu, S. Propofol increases micro-opioid receptor expression in SH-SY5Y human neuroblastoma cells. Mol. Med. Rep. 2012, 6, 1333–1336. [Google Scholar] [PubMed]
- Richards, M.J.; Skues, M.A.; Jarvis, A.P.; Prys-Roberts, C. Total i.v. anaesthesia with propofol and alfentanil: Dose requirements for propofol and the effect of premedication with clonidine. Br. J. Anaesth. 1990, 65, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Altmayer, P.; Buch, U.; Buch, H.P. Propofol binding to human blood proteins. Arzneimittelforschung 1995, 45, 1053–1056. [Google Scholar] [PubMed]
- Ludbrook, G.L.; Upton, R.N.; Grant, C.; Gray, E.C. Brain and blood concentrations of propofol after rapid intravenous injection in sheep, and their relationships to cerebral effects. Anaesth. Intensive Care 1996, 24, 445–452. [Google Scholar] [PubMed]
- Raoof, A.A.; Obbergh, L.J.V.; de Goyet, J.V.; Verbeeck, R.K. Extrahepatic glucuronidation of propofol in man: Possible contribution of gut wall and kidney. Eur. J. Clin. Pharmacol. 1996, 50, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Shafer, A.; Doze, V.A.; Shafer, S.L.; White, P.F. Pharmacokinetics and pharmacodynamics of propofol infusions during general anesthesia. Anesthesiology 1988, 69, 348–356. [Google Scholar] [CrossRef] [PubMed]
- Kotani, Y.; Shimazawa, M.; Yoshimura, S.; Iwama, T.; Hara, H. The experimental and clinical pharmacology of propofol, an anesthetic agent with neuroprotective properties. CNS Neurosci. Ther. 2008, 14, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Younes, I.; Rinaudo, M. Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [PubMed]
- Becher, R.D.; Peitzman, A.B.; Sperry, J.L.; Gallaher, J.R.; Neff, L.P.; Sun, Y.; Miller, P.R.; Chang, M.C. Damage control operations in non-trauma patients: Defining criteria for the staged rapid source control laparotomy in emergency general surgery. World J. Emerg. Surg. 2016, 11, 10. [Google Scholar] [CrossRef] [PubMed]
- Yousef, M.; Pichyangkura, R.; Soodvilai, S.; Chatsudthipong, V.; Muanprasat, C. Chitosan oligosaccharide as potential therapy of inflammatory bowel disease: Therapeutic efficacy and possible mechanisms of action. Pharmacol. Res. 2012, 66, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.Y.; Wang, G.K. A mutation in segment I-S6 alters slow inactivation of sodium channels. Biophys. J. 1997, 72, 1633–1640. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grade, n (%) | CG (n = 47) | PG (n = 47) | Chi-Square Statistic | p Values |
|---|---|---|---|---|
| No pain | 36 (76.6) | 6 (12.77) | 39.25 | 0.000 |
| Mild pain | 4 (8.51) | 12 (25.53) | ||
| Moderate pain | 5 (10.64) | 16 (34.04) | ||
| Severe pain | 2 (4.26) | 13 (27.66) | ||
| Total pain | 11 (23.4) | 41 (87.23) | 38.736 | 0.000 |
| Side Effects | CG (n = 47) | PG (n = 47) | Chi-Square Statistic | p Values |
|---|---|---|---|---|
| Apnea | 2 (4.26) | 8 (17.02) | 2.798 | 0.094 |
| Bradypnea (breaths < 6/min) | 1 (2.13) | 6 (12.77) | 2.470 | 0.116 |
| Obstructive respiration | 0 (0) | 5 (10.64) | 4.451 | 0.035 |
| Tachycardia (HR > 30% above BL) | 1 (2.13) | 8 (17.02) | 4.424 | 0.035 |
| Hypertension(MAP > 30%above BL) | 0 (0) | 6 (12.77) | 2.470 | 0.116 |
| Bradycardia (HR > 30% under BL) | 0 (2.13) | 5 (10.64) | 4.451 | 0.035 |
| Hypotension (MAP > 30% under BL) | 0 (0) | 7 (14.89) | 5.557 | 0.018 |
| Burning and stinging | 2 (4.26) | 10 (21.28) | 6.114 | 0.013 |
| Baseline Characters | CG (n = 47) | PG (n = 47) | T Value/Chi-Square Statistic | p Values |
|---|---|---|---|---|
| Age | 38.9 ± 15.6 | 40.1 ± 16.8 | 0.141 | 0.235 |
| Gender, male (%) | 32 (68.09) | 34 (72.34) | 0.203 | 0.652 |
| BMI | 22.7 ± 4.8 | 23.9 ± 6.5 | 0.037 | 0.326 |
| Smoking, n (%) | 28 (59.57) | 26 (55.32) | 0.174 | 0.677 |
| Drinking, n (%) | 29 (61.7) | 25 (53.19) | 0.696 | 0.404 |
| Spouse, n (%) | 44 (93.62) | 46 (97.87) | 0.261 | 0.409 |
| ASA | ||||
| I | 35 (74.47) | 37 (78.72) | 0.237 | 0.626 |
| II | 12 (25.53) | 10 (21.28) | ||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Yang, X.; Song, X.; Ma, H.; Zhang, P. Chitosan Oligosaccharide Reduces Propofol Requirements and Propofol-Related Side Effects. Mar. Drugs 2016, 14, 234. https://doi.org/10.3390/md14120234
Li Z, Yang X, Song X, Ma H, Zhang P. Chitosan Oligosaccharide Reduces Propofol Requirements and Propofol-Related Side Effects. Marine Drugs. 2016; 14(12):234. https://doi.org/10.3390/md14120234
Chicago/Turabian StyleLi, Zhiwen, Xige Yang, Xuesong Song, Haichun Ma, and Ping Zhang. 2016. "Chitosan Oligosaccharide Reduces Propofol Requirements and Propofol-Related Side Effects" Marine Drugs 14, no. 12: 234. https://doi.org/10.3390/md14120234
APA StyleLi, Z., Yang, X., Song, X., Ma, H., & Zhang, P. (2016). Chitosan Oligosaccharide Reduces Propofol Requirements and Propofol-Related Side Effects. Marine Drugs, 14(12), 234. https://doi.org/10.3390/md14120234