
Enzymatic Pre-Treatment Increases the Protein Bioaccessibility and Extractability in Dulse (Palmaria palmata)


Abstract
:1. Introduction
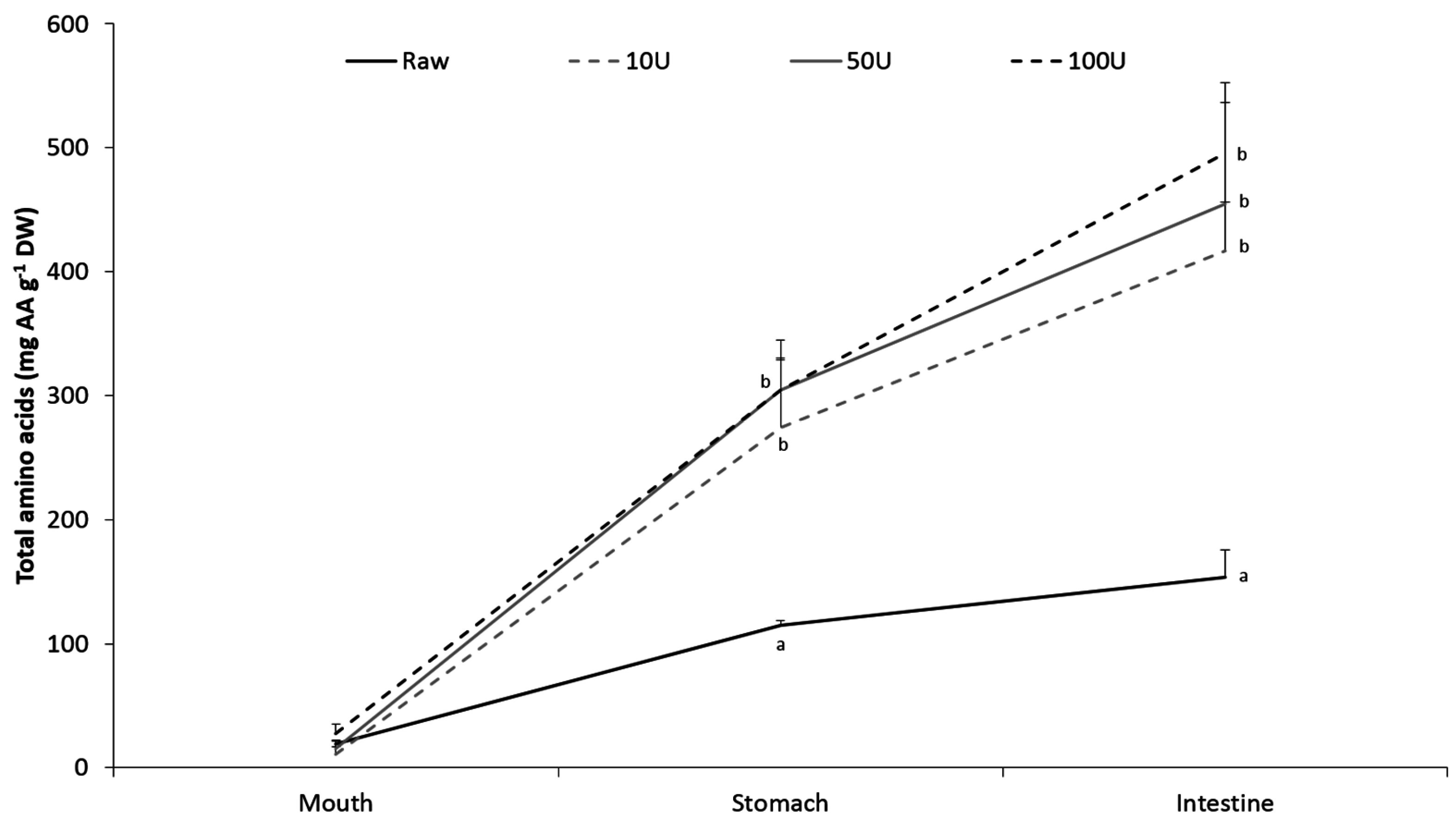
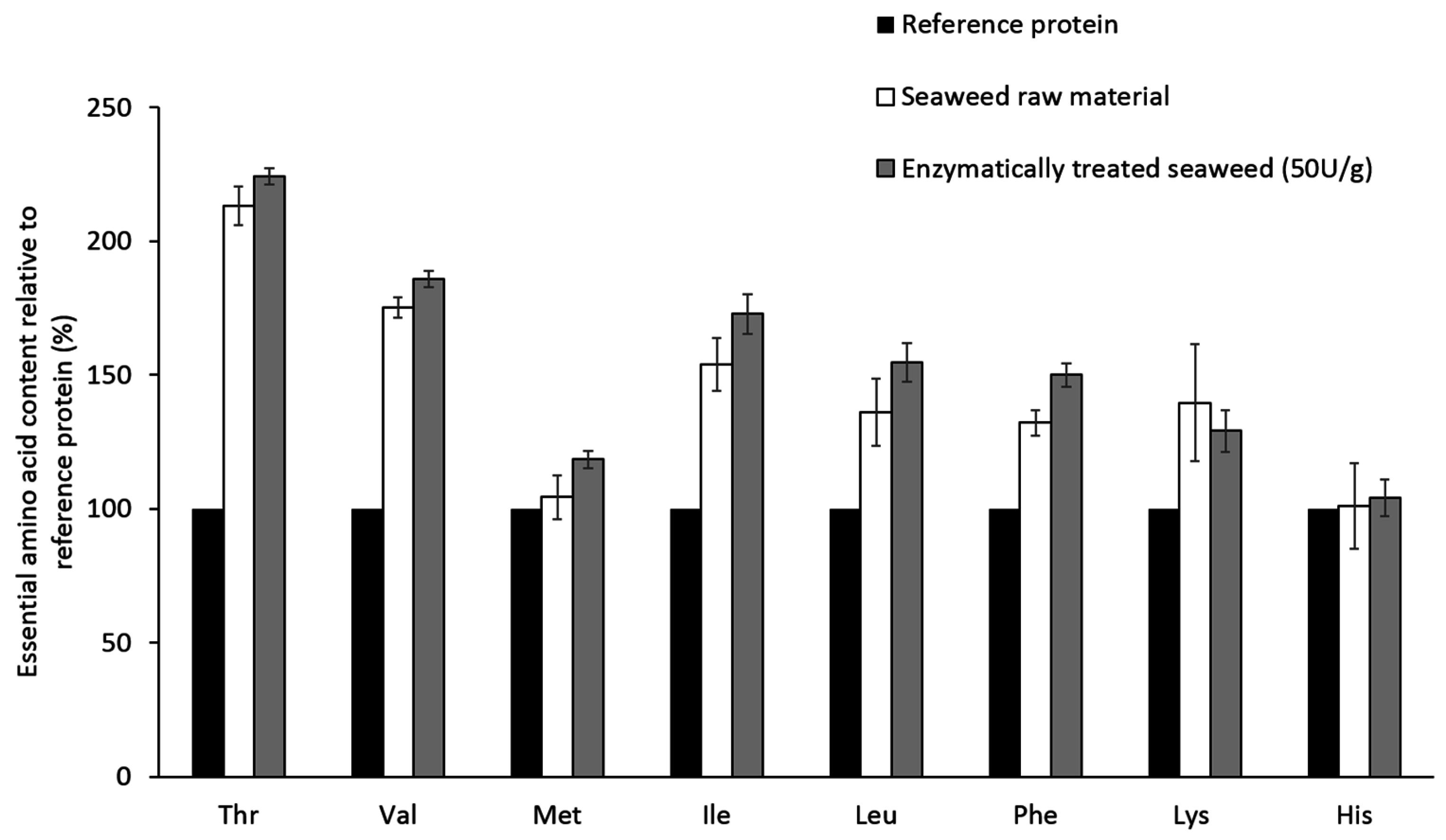
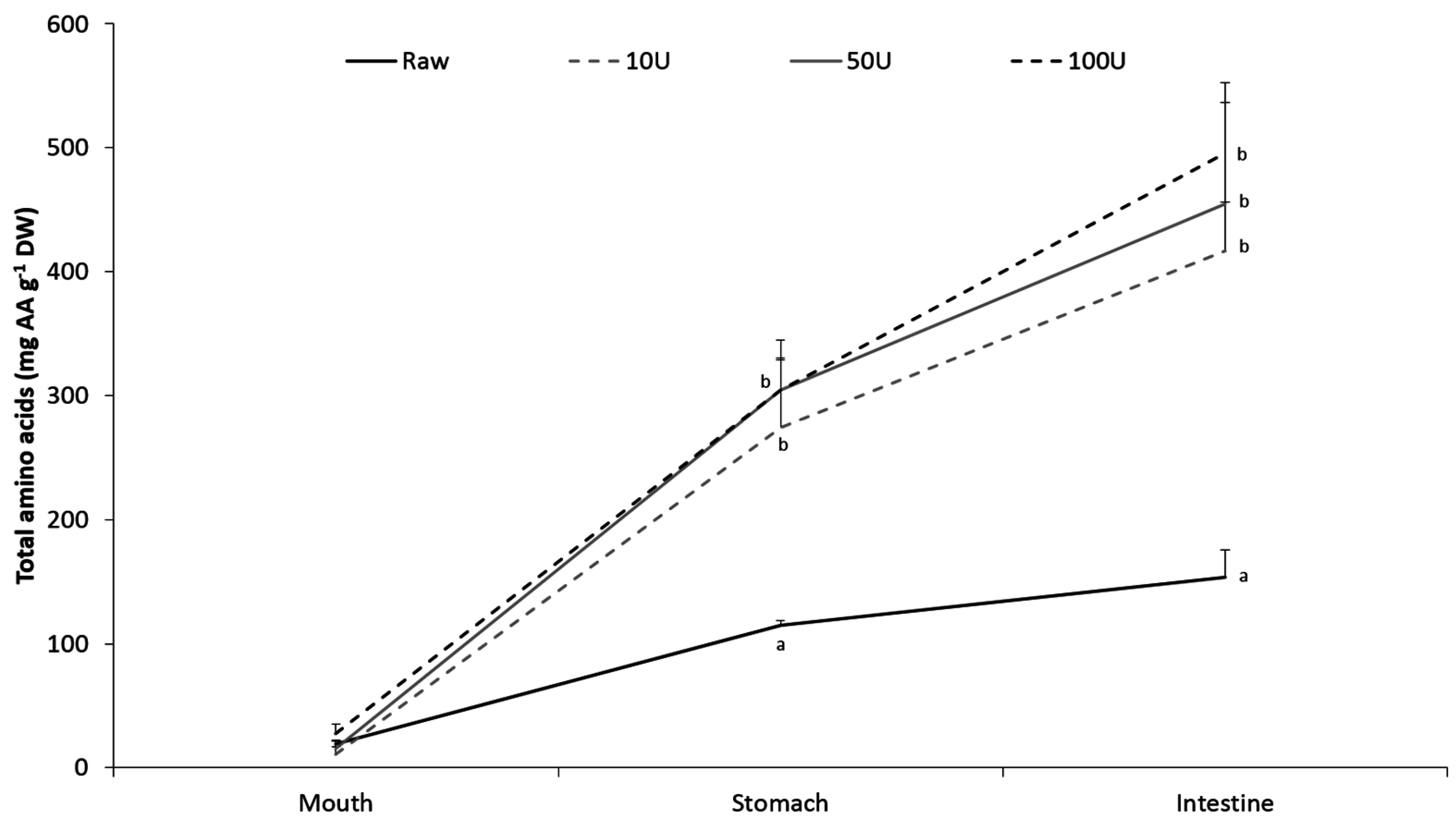
2. Results
3. Discussion
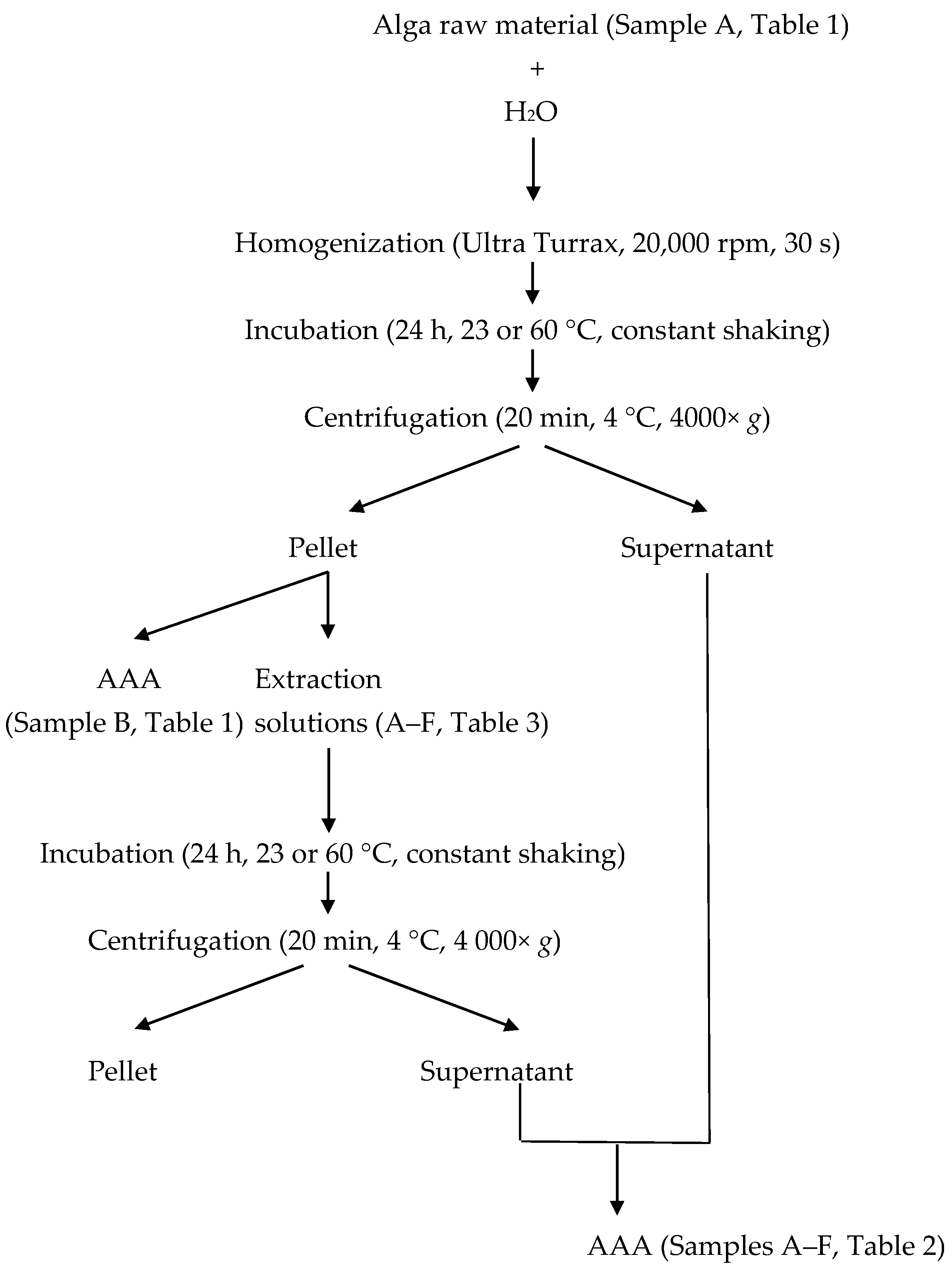
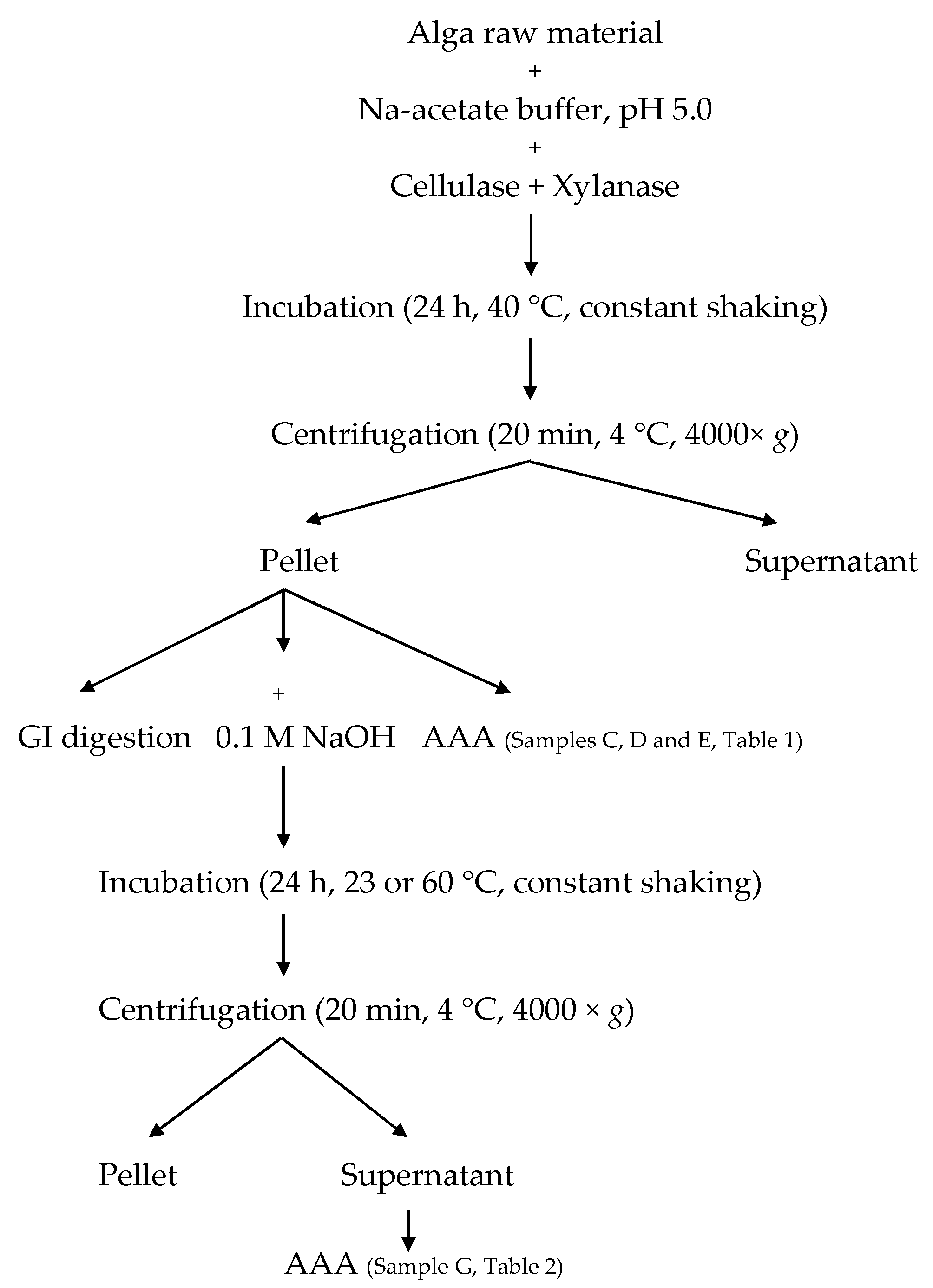
4. Experimental Section
4.1. Raw Material
4.2. Water Content
4.3. Protein Extraction
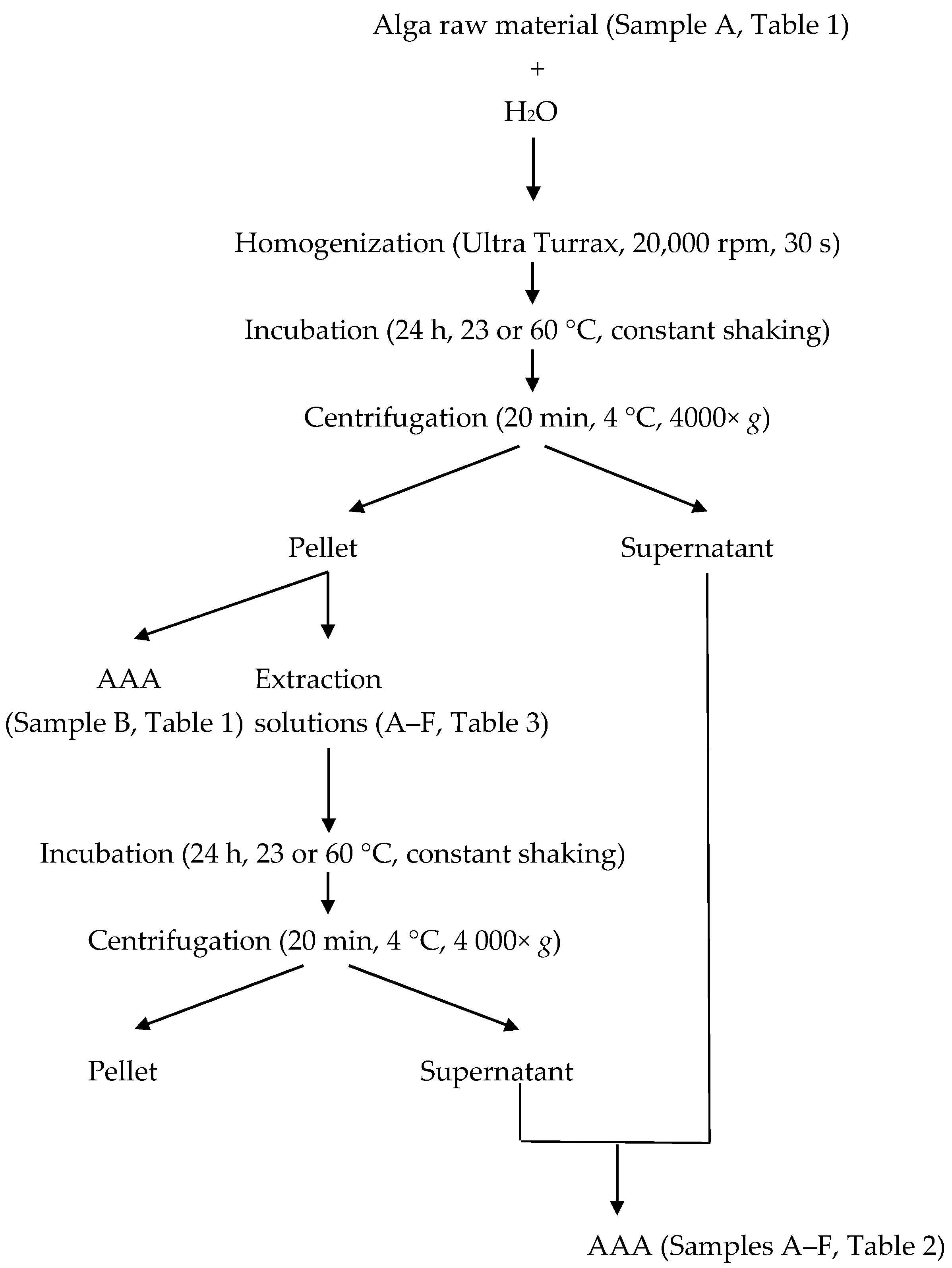
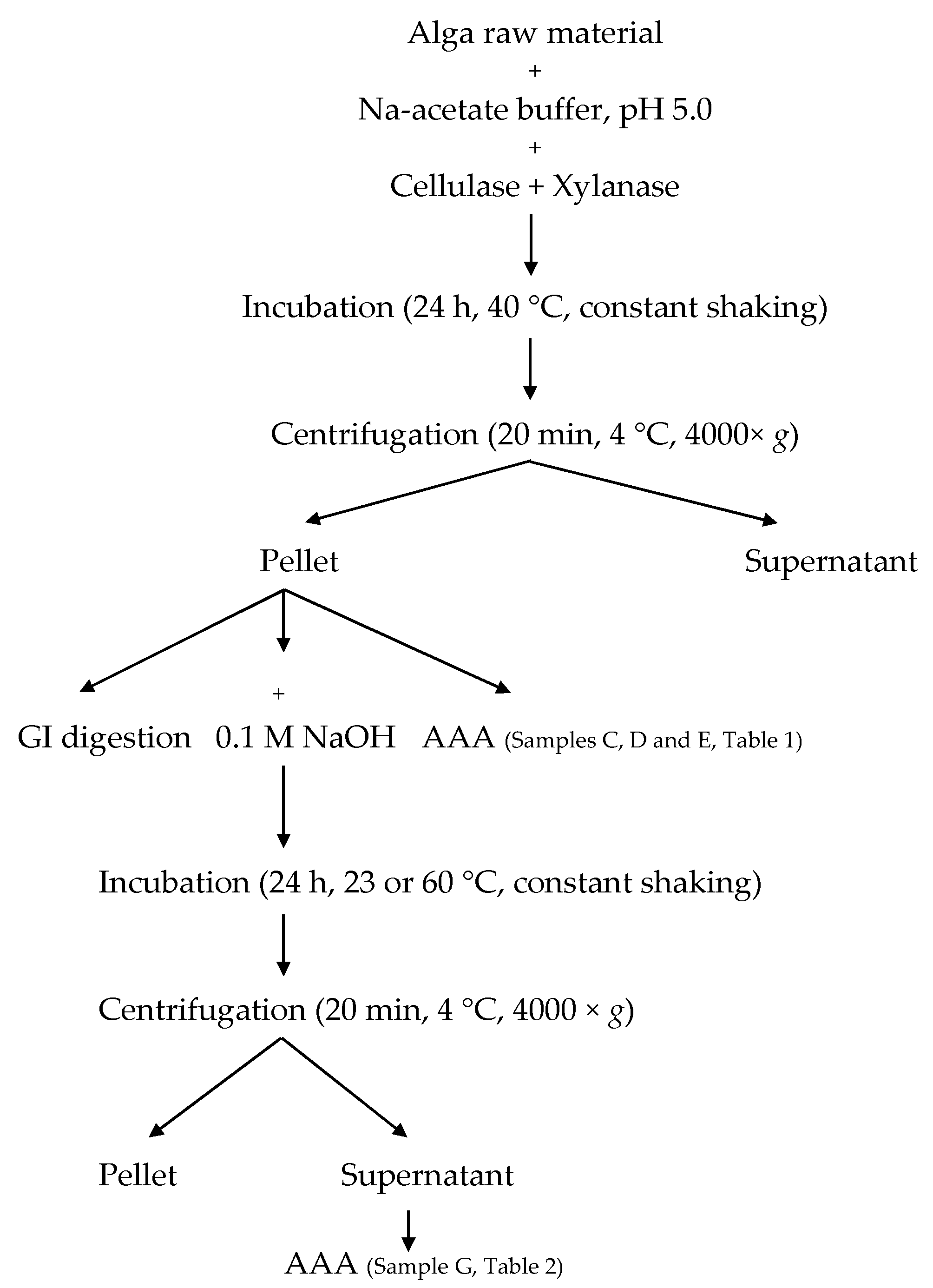
4.4. Enzymatic Pre-Treatment
4.5. In Vitro Gastrointestinal Digestion
4.6. Amino Acid Analysis
4.7. Statistics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Béné, C.; Barange, M.; Subasinghe, R.; Pinstrup-Andersen, P.; Merino, G.; Hemre, G.I.; Williams, M. Feeding 9 billion by 2050-Putting fish back on the menu. Food Secur. 2015, 7, 261–274. [Google Scholar] [CrossRef]
- Food and Agricultural Organization (FAO). The State of World Fisheries and Aquaculture 2014—Opportunities and Challenges; Food and Agricultural Organization of the United Nations: Rome, Italy, 2014; p. 223.
- Hurd, C.L.; Harrison, P.J.; Bischof, K.; Lobban, C.S. Nutrients. In Seaweed Ecology and Physiology; Hurd, C.L., Harrison, P.J., Bischof, K., Lobban, C.S., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 238–293. [Google Scholar]
- Kloareg, B.; Quatrano, R.S. Structure of the cell walls of marine algae and ecophysiological functions of the matrix polysaccharides. Oceanogr. Mar. Biol. 1988, 26, 259–315. [Google Scholar]
- Joubert, Y.; Fleurence, J. Simultaneous extraction of proteins and DNA by an enzymatic treatment of the cell wall of Palmaria palmata (Rhodophyta). J. Appl. Phycol. 2008, 20, 55–61. [Google Scholar] [CrossRef]
- Maehre, H.K.; Malde, M.K.; Eilertsen, K.E.; Elvevoll, E.O. Characterization of protein, lipid and mineral contents in common Norwegian seaweeds and evaluation of their potential as food and feed. J. Sci. Food Agric. 2014, 94, 3281–3290. [Google Scholar] [CrossRef] [PubMed]
- Barbarino, E.; Lourenço, S.O. An evaluation of methods for extraction and quantification of protein from marine macro- and microalgae. J. Appl. Phycol. 2005, 17, 447–460. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Extraction of protein from the macroalga Palmaria palmata. LWT Food Sci. Technol. 2013, 51, 375–382. [Google Scholar] [CrossRef]
- Galland-Irmouli, A.V.; Fleurence, J.; Lamghari, R.; Lucon, M.; Rouxel, C.; Barbaroux, O.; Bronowicki, J.P.; Villaume, C.; Gueant, J.L. Nutritional value of proteins from edible seaweed Palmaria palmata (Dulse). J. Nutr. Biochem. 1999, 10, 353–359. [Google Scholar] [CrossRef]
- Maehre, H.K.; Edvinsen, G.K.; Eilertsen, K.E.; Elvevoll, E.O. Heat treatment increases the protein bioaccessibility in the red seaweed dulse (Palmaria palmata), but not in the brown seaweed winged kelp (Alaria esculenta). J. Appl. Phycol. 2016, 28, 581–590. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO); World Health Organization (WHO); United Nations University (UNU). Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation; World Health Organization: Geneva, Switzerland, 2007; p. 265.
- Popper, Z.A.; Michel, G.; Herve, C.; Domozych, D.S.; Willats, W.G.T.; Tuohy, M.G.; Kloareg, B.; Stengel, D.B. Evolution and diversity of plant cell walls: From algae to flowering plants. Annu. Rev. Plant Biol. 2011, 62, 567–588. [Google Scholar] [CrossRef] [PubMed]
- Karsten, U. Seaweed acclimation to salinity and desiccation stress. In Seaweed Biology: Novel Insights into Ecophysiology, Ecology and Utilization; Wiencke, C., Bischof, K., Eds.; Springer: Heidelberg, Germany, 2012; pp. 87–107. [Google Scholar]
- Fleurence, J.; LeCoeur, C.; Mabeau, S.; Maurice, M.; Landrein, A. Comparison of different extractive procedures for proteins from the edible seaweeds Ulva rigida and Ulva rotundata. J. Appl. Phycol. 1995, 7, 577–582. [Google Scholar] [CrossRef]
- Horwitz, W. Official Methods of Analysis of AOAC International; AOAC International: Gaithersburg, MD, USA, 2004. [Google Scholar]
- Versantvoort, C.H.M.; Oomen, A.G.; Van de Kamp, E.; Rompelberg, C.J.M. Sips AJAM Applicability of an in vitro digestion model in assessing the bioaccessibility of mycotoxins from food. Food Chem. Toxicol. 2005, 43, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.; Stein, W.H. Chromatographic determination of amino acids by the use of automatic recording system. Methods Enzymol. 1963, 6, 819–831. [Google Scholar]
- Maehre, H.K.; Hamre, K.; Elvevoll, E.O. Nutrient evaluation of rotifers and zooplankton: Feed for marine fish larvae. Aquac. Nutr. 2013, 19, 301–311. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. | B. | C. | D. | E. | |
|---|---|---|---|---|---|
| Raw Material | After Homogenization | After Enzyme Pre-Treatment (10 U) | After Enzyme Pre-Treatment (50 U) | After Enzyme Pre-Treatment (100 U) | |
| Essential Amino Acids (EAA) | |||||
| Threonine | 8.9 ± 0.7 a | 16.8 ± 3.3 b | 25.7 ± 2.9 c | 30.4 ± 2.6 c | 27.1 ± 2.2 c |
| Valine | 12.4 ± 0.9 a | 22.6 ± 3.6 b | 36.9 ± 5.0 c | 42.9 ± 4.0 c | 38.5 ± 2.5 c |
| Methionine | 4.0 ± 0.4 a | 9.0 ± 1.7 b | 13.1 ± 2.2 b,c | 14.7 ± 1.3 c | 13.2 ± 0.8 c |
| Isoleucine | 8.2 ± 0.7 a | 15.8 ± 3.0 b | 26.2 ± 4.1 c | 30.0 ± 3.2 c | 26.8 ± 2.7 c |
| Leucine | 14.3 ± 1.8 a | 27.7 ± 4.7 b | 46.1 ± 6.3 c | 53.2 ± 6.0 c | 47.1 ± 4.8 c |
| Phenylalanine | 8.7 ± 0.6 a | 16.6 ± 2.9 b | 26.5 ± 3.0 c | 31.4 ± 2.4 c | 28.4 ± 1.6 c |
| Lysine | 11.0 ± 2.0 a | 19.5 ± 3.6 b | 28.8 ± 4.3 c | 33.7 ± 2.1 c | 29.5 ± 3.4 c |
| Histidine | 2.6 ± 0.5 a | 4.7 ± 0.7 b | 7.0 ± 0.8 c | 8.7 ± 0.5 c | 7.9 ± 0.8 c |
| Non-Essential Amino Acids (NEAA) | |||||
| Aspartic acid * | 21.9 ± 1.2 a | 32.1 ± 5.5 a | 50.9 ± 6.2 b | 59.2 ± 5.7 b | 52.8 ± 4.4 b |
| Serine | 10.5 ± 0.8 a | 20.5 ± 3.9 b | 31.8 ± 4.3 c | 37.7 ± 3.9 c | 33.2 ± 3.0 c |
| Glutamic acid * | 20.4 ± 1.8 a | 27.7 ± 5.6 a | 43.1 ± 5.3 b | 50.3 ± 5.2 b | 44.1 ± 3.1 b |
| Proline | 9.1 ± 0.4 a | 14.0 ± 3.1 a | 23.8 ± 2.2 b | 27.7 ± 3.8 b | 25.3 ± 2.4 b |
| Glycine | 12.1 ± 0.8 a | 20.7 ± 3.9 b | 32.4 ± 3.3 c | 37.6 ± 3.5 c | 34.6 ± 2.0 c |
| Alanine | 16.4 ± 1.4 a | 28.7 ± 5.2 b | 44.7 ± 7.0 c | 50.5 ± 5.1 c | 43.7 ± 3.1 c |
| Cysteine | 1.4 ± 0.4 a | 3.0 ± 0.8 a | 4.4 ± 1.7 b | 7.1 ± 1.2 b | 7.1 ± 1.4 b |
| Tyrosine | 6.9 ± 0.9 a | 13.3 ± 2.8 a | 23.6 ± 3.2 a,b | 29.2 ± 2.7 b | 26.2 ± 2.7 b |
| Arginine | 11.5 ± 1.1 a | 22.5 ± 4.4 b | 34.7 ± 4.9 c | 41.6 ± 2.3 c | 35.8 ± 3.6 c |
| Sum | 180.5 ± 12.3 a | 312.0 ± 54.2 b | 495.2 ± 59.5 c | 586.1 ± 53.5 c | 521.2 ± 40.7 c |
| Relative amount EAA (%) | 38.9 ± 0.6 a | 42.6 ± 0.9 b | 42.5 ± 1.2 b | 41.8 ± 0.3 b | 41.9 ± 0.5 b |
| Extraction Temperature | |||||
|---|---|---|---|---|---|
| 23 °C | 60 °C | ||||
| Solvent | Amount Extracted Amino Acids (mg·g−1·DW) | Extraction Yield (%) | Amount Extracted Amino Acids (mg·g−1·DW) | Extraction Yield (%) | |
| A | 0.01 M NaOH | 55.8 ± 10.2 b | 17.9 | 59.9 ± 7.2 B | 19.2 |
| B | 0.05 M NaOH | 80.6 ± 9.5 b,c | 25.8 | 118.1 ± 25.2 B,C,* | 37.9 |
| C | 0.1 M NaOH | 90.1 ± 7.9 c | 28.9 | 122.0 ± 10.5 C,* | 39.1 |
| D | 3.5% NaCl | 18.3 ± 4.7 a | 5.9 | 26.6 ± 7.0 A | 8.5 |
| E | 70% Ethanol | 23.5 ± 4.6 a | 7.5 | 27.3 ± 4.6 A | 8.8 |
| F | 0.1 M NaOH in 3.5% NaCl | 58.8 ± 13.3 b | 18.8 | 114.6 ± 19.2 C,* | 36.7 |
| G | 0.1 M NaOH following enzymatic pre-treatment | 409.2 ± 46.0 d | 69.8 | 442.8 ± 86.5 D | 75.6 |
| Extract | Extraction Solvent | Type of Extracted Protein |
|---|---|---|
| All | Water | Albumins |
| A | 0.01 M NaOH | Glutelins |
| B | 0.05 M NaOH | Glutelins |
| C | 0.1 M NaOH | Glutelins |
| D | 3.5% NaCl | Globulins |
| E | 70% Ethanol | Prolamines |
| F | 0.1 M NaOH in 3.5% NaCl | Combination |
| G | Enzymes + 0.1 M NaOH | Combination |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mæhre, H.K.; Jensen, I.-J.; Eilertsen, K.-E. Enzymatic Pre-Treatment Increases the Protein Bioaccessibility and Extractability in Dulse (Palmaria palmata). Mar. Drugs 2016, 14, 196. https://doi.org/10.3390/md14110196
Mæhre HK, Jensen I-J, Eilertsen K-E. Enzymatic Pre-Treatment Increases the Protein Bioaccessibility and Extractability in Dulse (Palmaria palmata). Marine Drugs. 2016; 14(11):196. https://doi.org/10.3390/md14110196
Chicago/Turabian StyleMæhre, Hanne K., Ida-Johanne Jensen, and Karl-Erik Eilertsen. 2016. "Enzymatic Pre-Treatment Increases the Protein Bioaccessibility and Extractability in Dulse (Palmaria palmata)" Marine Drugs 14, no. 11: 196. https://doi.org/10.3390/md14110196
APA StyleMæhre, H. K., Jensen, I.-J., & Eilertsen, K.-E. (2016). Enzymatic Pre-Treatment Increases the Protein Bioaccessibility and Extractability in Dulse (Palmaria palmata). Marine Drugs, 14(11), 196. https://doi.org/10.3390/md14110196