
Liquid Chromatography-Mass Spectrometry-Based Rapid Secondary-Metabolite Profiling of Marine Pseudoalteromonas sp. M2

 ,
, 
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Screening and Identification of SW1-1 Strain

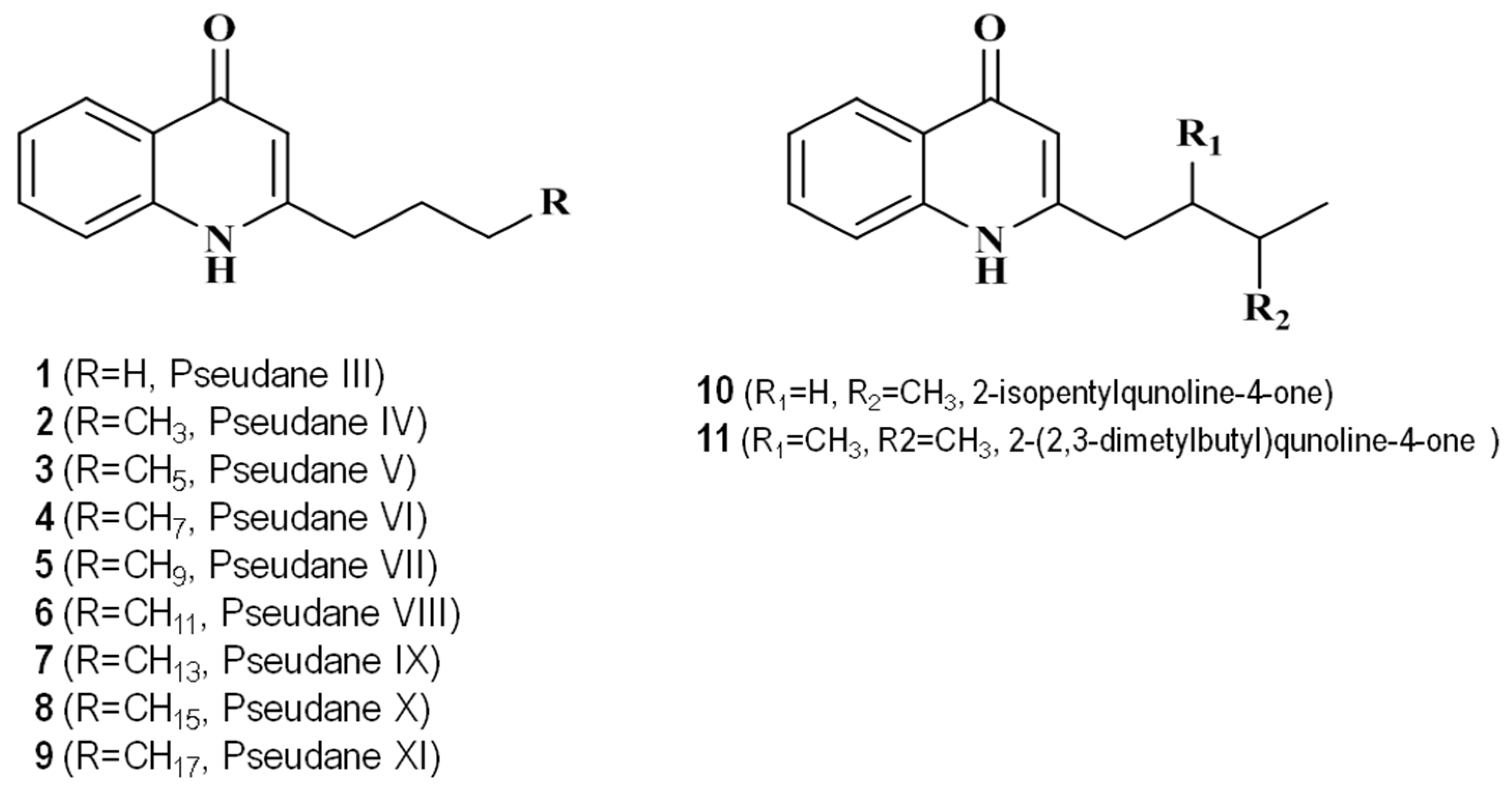
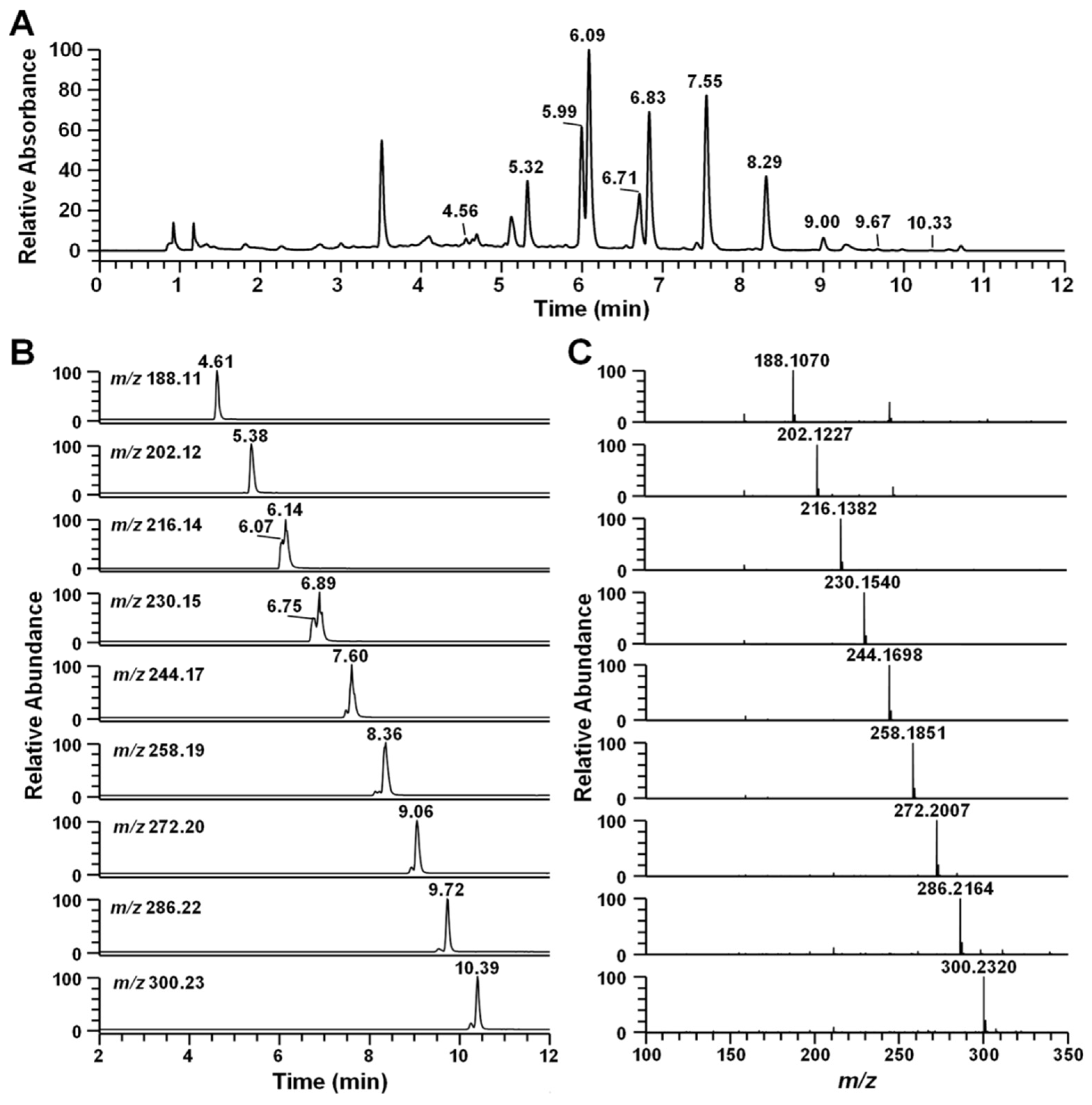
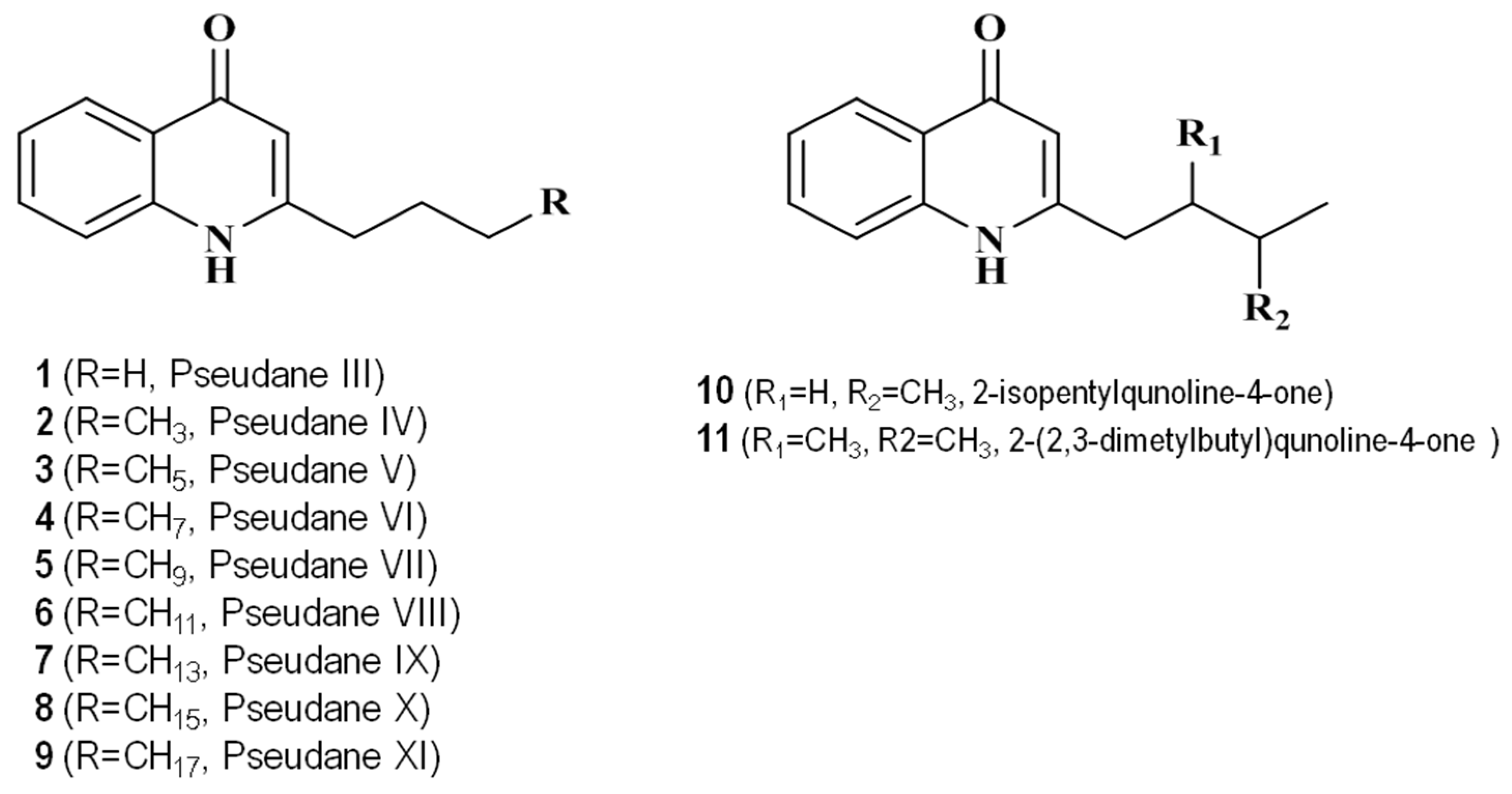
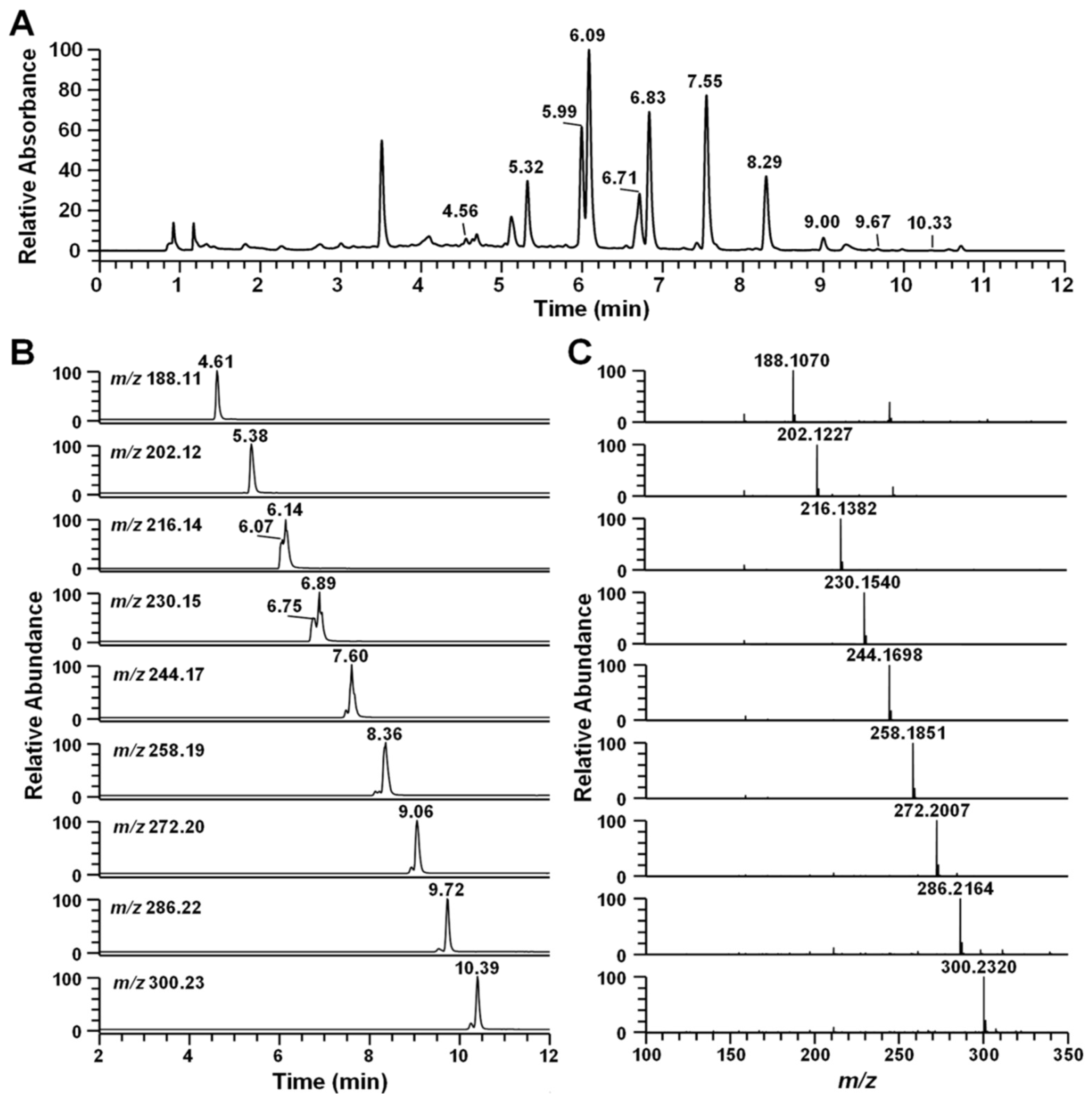
2.2. Identification of Secondary Metabolites Using High-Resolution Mass Spectrometry

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT (min) | Compound | [M + H]+ (m/z) | Formula | ∆ppm | MS/MS Fragment Ion | Ref. |
|---|---|---|---|---|---|---|
| 4.56 | Pseudane-III | 188.1070 | C12H14ON | −0.003 | 132, 146, 159, 170, 188 | |
| 5.32 | Pseudane-IV | 202.1227 | C13H16ON | 0.491 | 132, 146, 159, 172, 183, 202 | * |
| 5.99 | 2-isopentylquinolin-4-one | 216.1382 | C14H18ON | −0.188 | 132, 146, 159, 172, 186, 200, 216 | Novel |
| 6.09 | Pseudane-V | 216.1382 | C14H18ON | −0.188 | 132, 146, 159, 172, 186, 197, 216 | [15] |
| 6.71 | 2-(2,3-dimethylbutyl) quinolin-4-one | 230.1539 | C15H20ON | 0.083 | 132, 146, 159, 172, 186, 200, 230 | Novel |
| 6.83 | Pseudane-VI | 230.1539 | C15H20ON | −0.090 | 132, 146, 159, 172, 186, 200, 230 | * |
| 7.55 | Pseudane-VII | 244.1695 | C16H22ON | −0.372 | 132, 146, 159, 172, 186, 200, 244 | [16] |
| 8.29 | Pseudane-VIII | 258.1851 | C17H24ON | −0.701 | 132, 146, 159, 172, 186, 200, 258 | * |
| 9.00 | Pseudane-IX | 272.2007 | C18H26ON | −0.885 | 132, 146, 159, 172, 186, 200, 272 | [16] |
| 9.67 | Pseudane-X | 286.2164 | C19H28ON | −0.131 | 146, 159, 172, 186, 200, 214, 286 | [17] |
| 10.33 | Pseudane-XI | 300.2322 | C20H30ON | −0.001 | 146, 159, 172, 186, 200, 214, 300 | [17] |
2.3. NMR Analysis of the New Compounds
| No. C/H | δC ppm | δH (ppm), Integration, Multiplicity, J (Hz) |
|---|---|---|
| 2-isopentylquinolin-4-one | ||
| 1 (CH3) | 22.8 | 0.99 (3H, m, J = 8.3, 7.0, 1.3 Hz) |
| 2 (CH3) | 22.8 | 0.99 (3H, m, J = 8.3, 7.0, 1.3 Hz) |
| 5 (CH) | 29.2 | 1.67 (1H, m, J = 7.3 Hz) |
| 3 (CH2) | 33.2 | 2.72 (2H, m, J = 8.5 Hz) |
| 4 (CH2) | 39.3 | 1.63 (2H, m) |
| 6 (CH) | 108.8 | 6.22 (1H, s) |
| 7 (CH) | 119.2 | 7.58 (1H, ddd, J = 8.46 Hz) |
| 8 (CH) | 125.2 | 7.38 (1H, d, J = 8.16, 7.03, 1.13 Hz) |
| 11 (C) | 125.6 | |
| 9 (CH) | 126.11 | 8.20 (1H,dd, J = 8.53, 1.26 Hz) |
| 10 (CH) | 133.5 | 7.68 (1H, t, J = 7.39 Hz) |
| 12 (C) | 141.7 | |
| 13 (C) | 157.5 | |
| 14 (C) | 180.8 | |
| 2-(2,3-dimethylbutyl)quinolin-4-one | ||
| 1 (CH3) | 15.2 | 0.86 (3H, d, J = 6.78 Hz) |
| 2 (CH3) | 18.2 | 0.95 (3H, d, J = 6.78 Hz) |
| 3 (CH3) | 20.6 | 0.97 (3H, d, J = 6.78 Hz) |
| 5 (CH) | 33.4 | 1.69 (1H, d, J = 13.58, 6.82, 4.39 Hz) |
| 4 (CH2) | 39.9 | 2.83 (1H, dd, J = 13.8, 5.52 Hz) |
| 6 (CH) | 40.5 | 1.89 (1H, m) |
| 7 (CH) | 109.8 | 6.21 (1H, s) |
| 8 (CH) | 119.2 | 7.39 (1H, t, J = 7.54 Hz) |
| 9 (CH) | 125.2 | 7.59 (1H, d, J = 8.03 Hz) |
| 12 (C) | 125.6 | |
| 10 (CH) | 126.1 | 8.21 (1H, d, J = 8.2 Hz) |
| 11 (CH) | 133.5 | 7.68 (1H, m) |
| 13 (C) | 141.7 | |
| 14 (C) | 156.8 | |
| 15 (C) | 180.5 | |
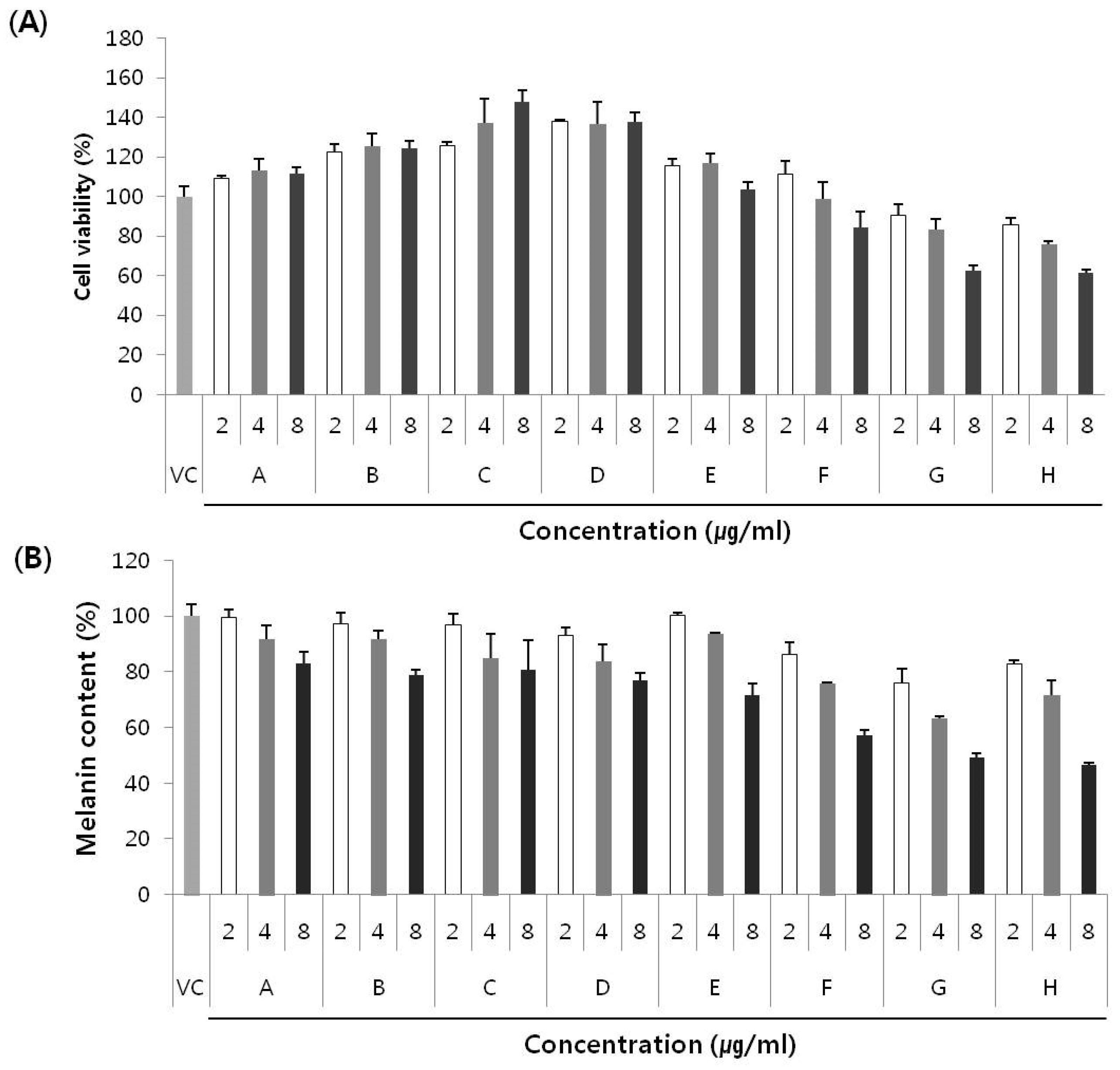
2.4. Anti-Melanogenic Effect of Secondary Metabolites and New Compounds
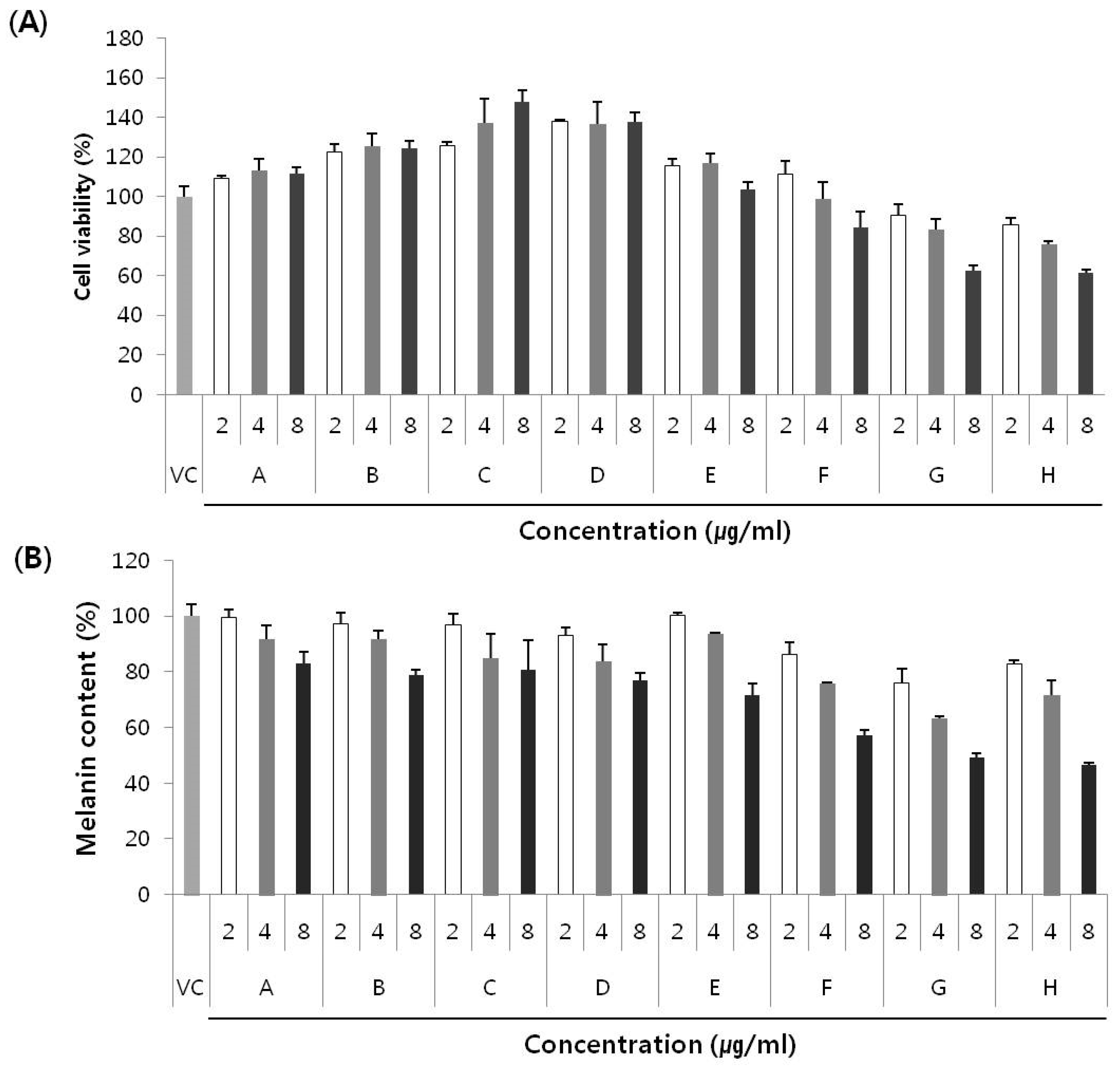
3. Experimental Section
3.1. Isolation of Pseudane-Producing Bacterium
3.2. Identification of Pseudoalteromonas sp.
3.3. Secondary Metabolite-Profiling Using LC-MS
3.4. Extraction and Purification of Secondary Metabolites
3.5. Nuclear Magnetic Resonance (NMR) Analysis
3.6. Cell Cultures
3.7. Cell Viability Assay
3.8. Measurement of Melanin Content
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Paul, V.J. (Ed.) Ecological Roles of Marine Natural Products (Explorations in Chemical Ecology); Cornstock Press: Ithaca, NY, USA, 1992; p. 245.
- Gauthier, G.; Gauthier, M.; Christen, R. Phylogenetic analysis of the genera Alteromonas, Shewanella, and Moritella using genes coding for small-subunit rRNA sequences and division of the genus Alteromonas into two genera, Alteromonas (emended) and Pseudoalteromonas gen. nov., and proposal of twelve new species combinations. Int. J. Syst. Bacteriol. 1995, 45, 755–761. [Google Scholar] [PubMed]
- Holmström, C.; Kjelleberg, S. Marine Pseudoalteromonas species are associated with higher organisms and produce biologically active extracellular agents. FEMS Microbiol. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Heeb, S.; Fletcher, M.P.; Chhabra, S.R.; Diggle, S.P.; Williams, P.; Camara, M. Quinolones: From antibiotics to autoinducers. FEMS Microbiol. Rev. 2011, 35, 247–274. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, D. The quinine connection. J. Antimicrob. Chemother. 1992, 30, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Pesci, E.C.; Milbank, J.B.; Pearson, J.P.; McKnight, S.; Kende, A.S.; Greenberg, E.P.; Iglewski, B.H. Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 1999, 96, 11229–11234. [Google Scholar] [CrossRef] [PubMed]
- Cardozo, V.F.; Oliveira, A.G.; Nishio, E.K.; Perugini, M.R.; Andrade, C.G.; Silveira, W.D.; Durán, N.; Andrade, G.; Kobayashi, R.K.; Nakazato, G. Antibacterial activity of extracellular compounds produced by a Pseudomonas strain against methicillin-resistant Staphylococcus aureus (MRSA) strains. Ann. Clin. Microbiol. Antimicrob. 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Perez-De, L.C.V.; Carrillo-Mora, P.; Santamaria, A. Qunolinic acid, an endogenous molecule combining excitotoxicity, oxidative stress and other toxic mechanisms. Int. J. Tryptophan Res. 2012, 5, 1–8. [Google Scholar]
- Rochfort, S. Metabolomics reviewed: A new “Omics” platform technology for systems biology and implications for natural products research. J. Nat. Prod. 2005, 68, 1813–1820. [Google Scholar] [CrossRef] [PubMed]
- Want, J.E.; Cravatt, B.F.; Siuzdak, G. The expanding role of mass spectrometry in metabolite profiling and characterization. ChemBioChem 2005, 6, 1941–1951. [Google Scholar] [CrossRef] [PubMed]
- Bino, R.J.; Hall, R.D.; Fiehn, O.; Kopka, J.; Saito, K.; Draper, J.; Nikolau, B.J.; Mendes, P.; Roessner-Tunali, U.; Beale, M.H.; et al. Potential of metabolomics as a functional genomics tool. Trends Plant Sci. 2004, 9, 418–425. [Google Scholar] [CrossRef] [PubMed]
- Krug, D.; Zurek, G.; Revermann, O.; Vos, M.; Velicer, G.J.; Müller, R. Discovering the hidden secondary metabolome of Myxococcus xanthus: A study of intraspecific diversity. Appl. Environ. Microbiol. 2008, 74, 3058–3068. [Google Scholar] [CrossRef] [PubMed]
- Villas-Bôas, S.G.; Mas, S.; Akesson, M.; Smedsgaard, J.; Nielsen, J. Mass spectrometry in metabolome analysis. Mass Spectrom. Rev. 2005, 24, 613–646. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kim, Y.S.; Park, S.; Kim, J.; Kang, S.J.; Lee, M.H.; Ryu, S.; Choi, J.M.; Oh, T.K.; Yoon, J.H. Exceptional production of both prodigiosin and cycloprodigiosin as major metabolic constituents by a novel marine bacterium, Zooshikella rubidus S1-1. Appl. Environ. Microbiol. 2011, 77, 4967–4973. [Google Scholar] [CrossRef] [PubMed]
- Long, R.A.; Qureshi, A.; Faulkner, D.J.; Azam, F. 2-n-Pentyl-4-Quinolinol produced by a marine Alteromonas sp. and its potential ecological and biogeochemical roles. Appl. Environ. Microbiol. 2003, 69, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Ritter, C.; Luckner, M. Biosynthesis of 2-n-alkyl-4-hydroxyquinoline derivates (pseudane) in Pseudomonas aeruginosa. Eur. J. Biochem. 1971, 18, 391–400. (In German) [Google Scholar] [CrossRef] [PubMed]
- Lépine, F.; Milot, S.; Déziel, E.; He, J.; Rahme, L.G. Electrospray/mass spectrometric identification and analysis of 4-hydoxy-2-alkylquinolines (HAQs) produced by Pseudomonas aeruginosa. J. Am. Soc. Mass Spectrom. 2004, 15, 862–869. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.; Cha, B.J.; Lee, Y.S.; Kim, G.S.; Noh, H.J.; Kim, S.Y.; Kang, H.C.; Kim, J.H.; Baek, N.I. The potential of minor ginsenosides isolated from the leaves of Panax ginseng as inhibitors of melanogenesis. Int. J. Mol. Sci. 2015, 16, 1677–1690. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.H.; Jo, Y.O.; Cho, C.W.; Son, D.; Park, S.; Rho, J.; Choi, S.Y. Inhibitory effects of cinnamic acid on melanin biosynthesis in skin. Biol. Pharm. Bull. 2008, 31, 946–948. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Miura, K.I. Preparation of transforming deoxyribonucleic acid by phenol treatment. Biochim. Biophys. Acta 1963, 72, 619–629. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Tamura, K.; Jakobsen, I.B.; Nei, M. MEGA2: Molecular evolutionary genetics analysis software. Bioinformatics 2001, 17, 1244–1245. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, W.J.; Kim, Y.O.; Kim, J.H.; Nam, B.-H.; Kim, D.-G.; An, C.M.; Lee, J.S.; Kim, P.S.; Lee, H.M.; Oh, J.-S.; et al. Liquid Chromatography-Mass Spectrometry-Based Rapid Secondary-Metabolite Profiling of Marine Pseudoalteromonas sp. M2. Mar. Drugs 2016, 14, 24. https://doi.org/10.3390/md14010024
Kim WJ, Kim YO, Kim JH, Nam B-H, Kim D-G, An CM, Lee JS, Kim PS, Lee HM, Oh J-S, et al. Liquid Chromatography-Mass Spectrometry-Based Rapid Secondary-Metabolite Profiling of Marine Pseudoalteromonas sp. M2. Marine Drugs. 2016; 14(1):24. https://doi.org/10.3390/md14010024
Chicago/Turabian StyleKim, Woo Jung, Young Ok Kim, Jin Hee Kim, Bo-Hye Nam, Dong-Gyun Kim, Cheul Min An, Jun Sik Lee, Pan Soo Kim, Hye Min Lee, Joa-Sup Oh, and et al. 2016. "Liquid Chromatography-Mass Spectrometry-Based Rapid Secondary-Metabolite Profiling of Marine Pseudoalteromonas sp. M2" Marine Drugs 14, no. 1: 24. https://doi.org/10.3390/md14010024
APA StyleKim, W. J., Kim, Y. O., Kim, J. H., Nam, B.-H., Kim, D.-G., An, C. M., Lee, J. S., Kim, P. S., Lee, H. M., Oh, J.-S., & Lee, J. S. (2016). Liquid Chromatography-Mass Spectrometry-Based Rapid Secondary-Metabolite Profiling of Marine Pseudoalteromonas sp. M2. Marine Drugs, 14(1), 24. https://doi.org/10.3390/md14010024