
Inhibition of Ultraviolet B-Induced Expression of the Proinflammatory Cytokines TNF-α and VEGF in the Cornea by Fucoxanthin Treatment in a Rat Model

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
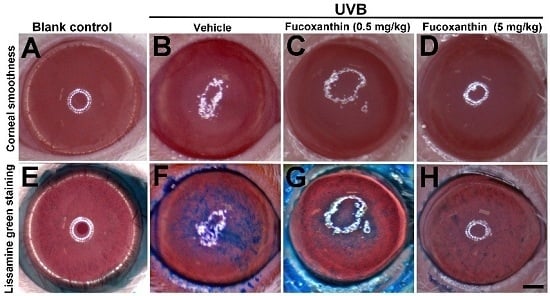
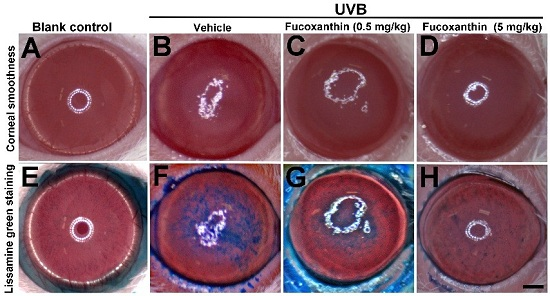
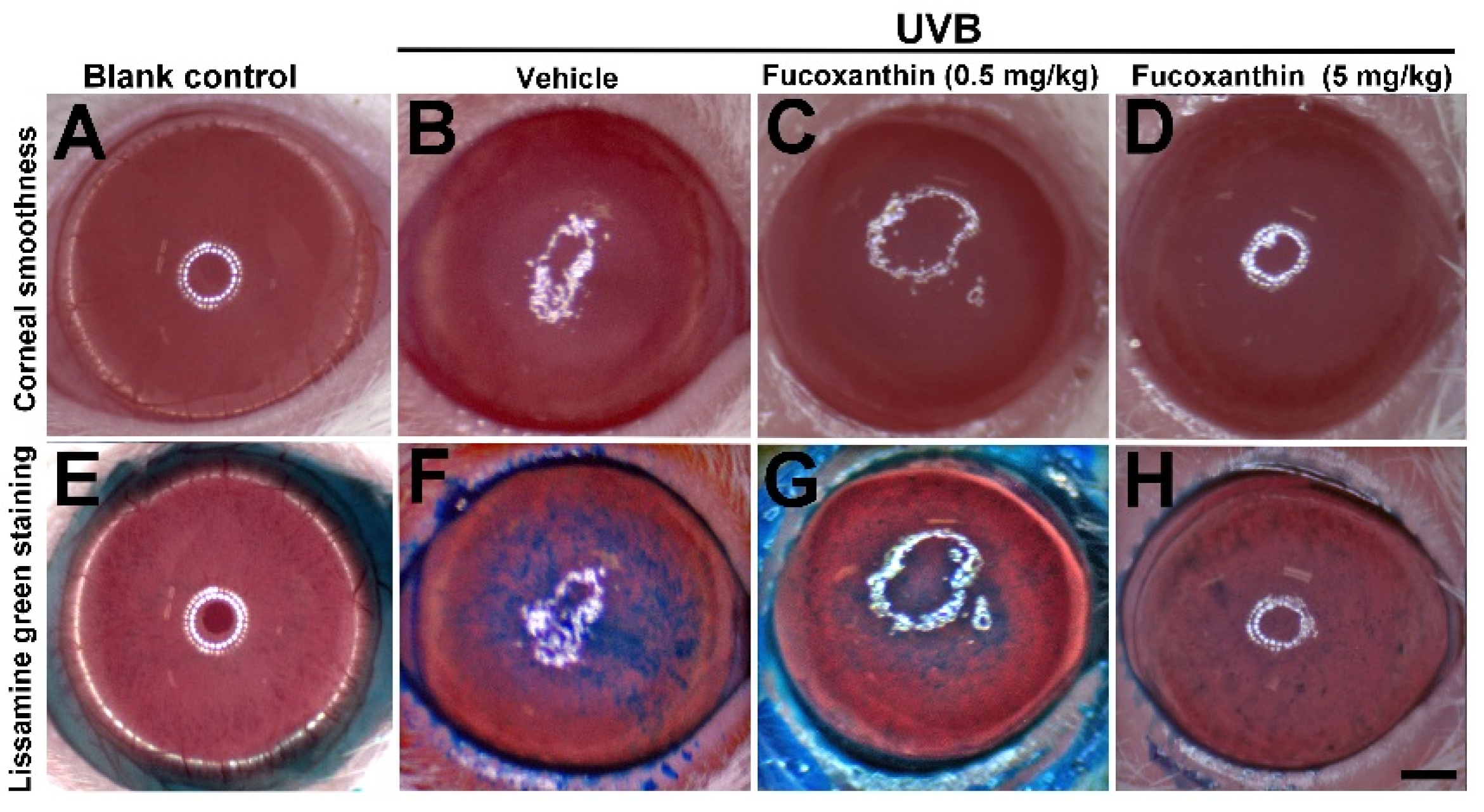
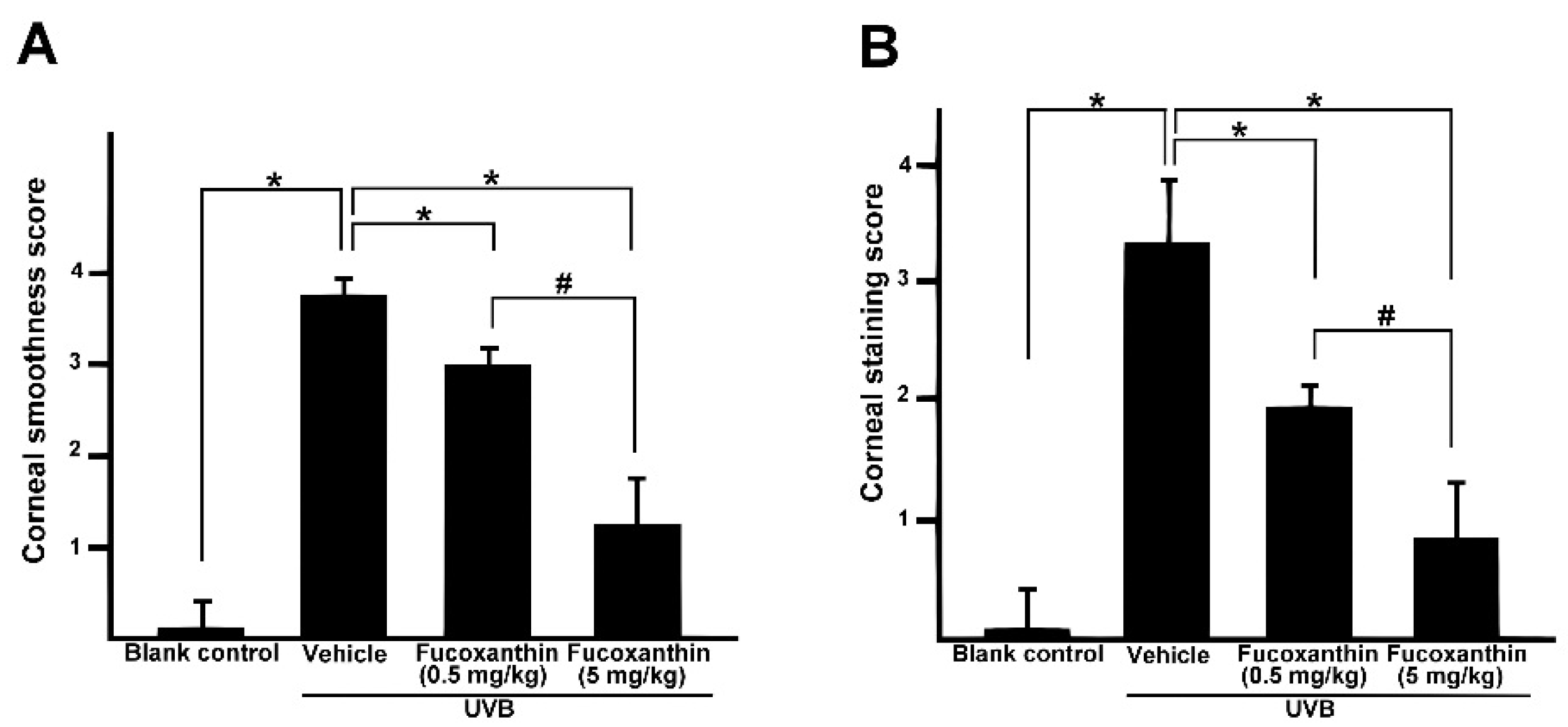
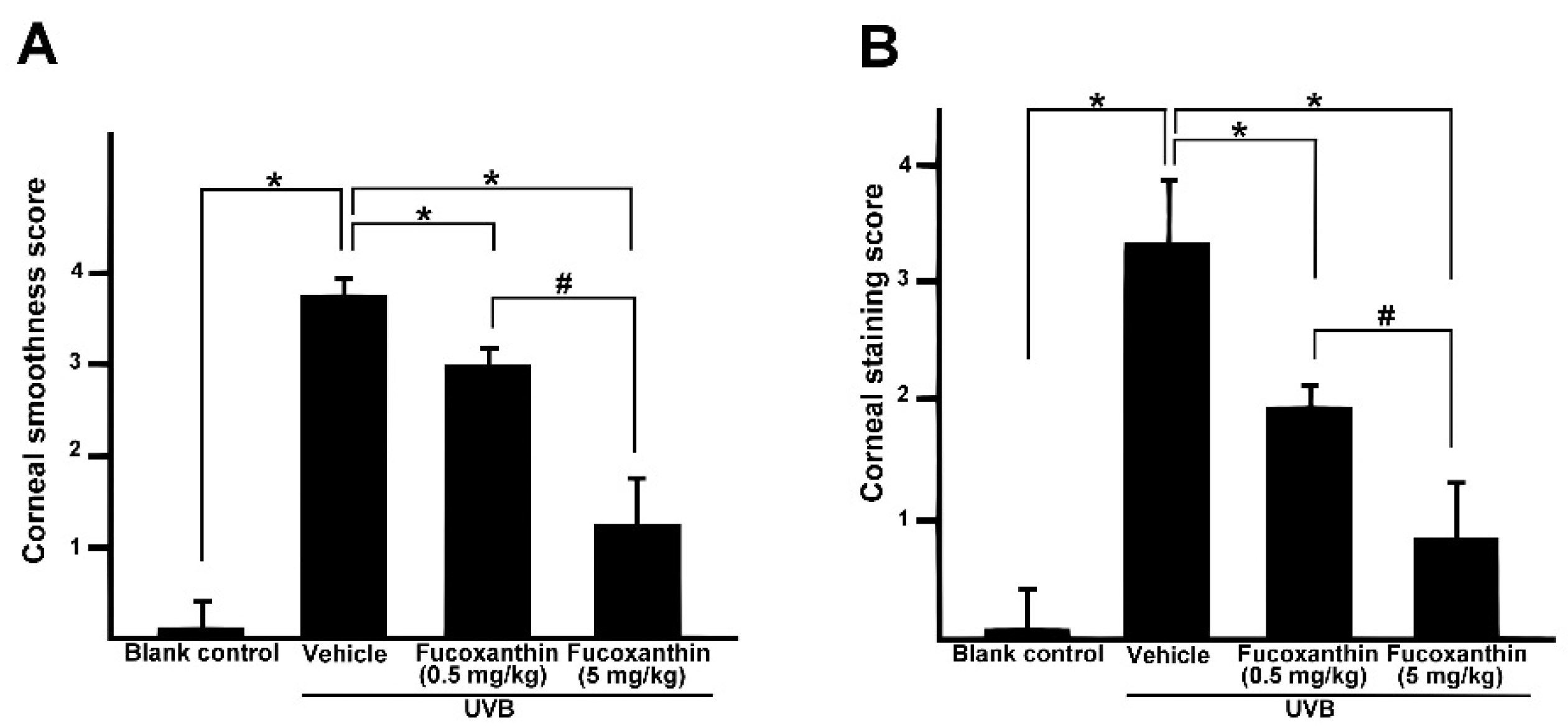
2.1. Protective Effects of Fucoxanthin on UVB-Induced Corneal Disorders

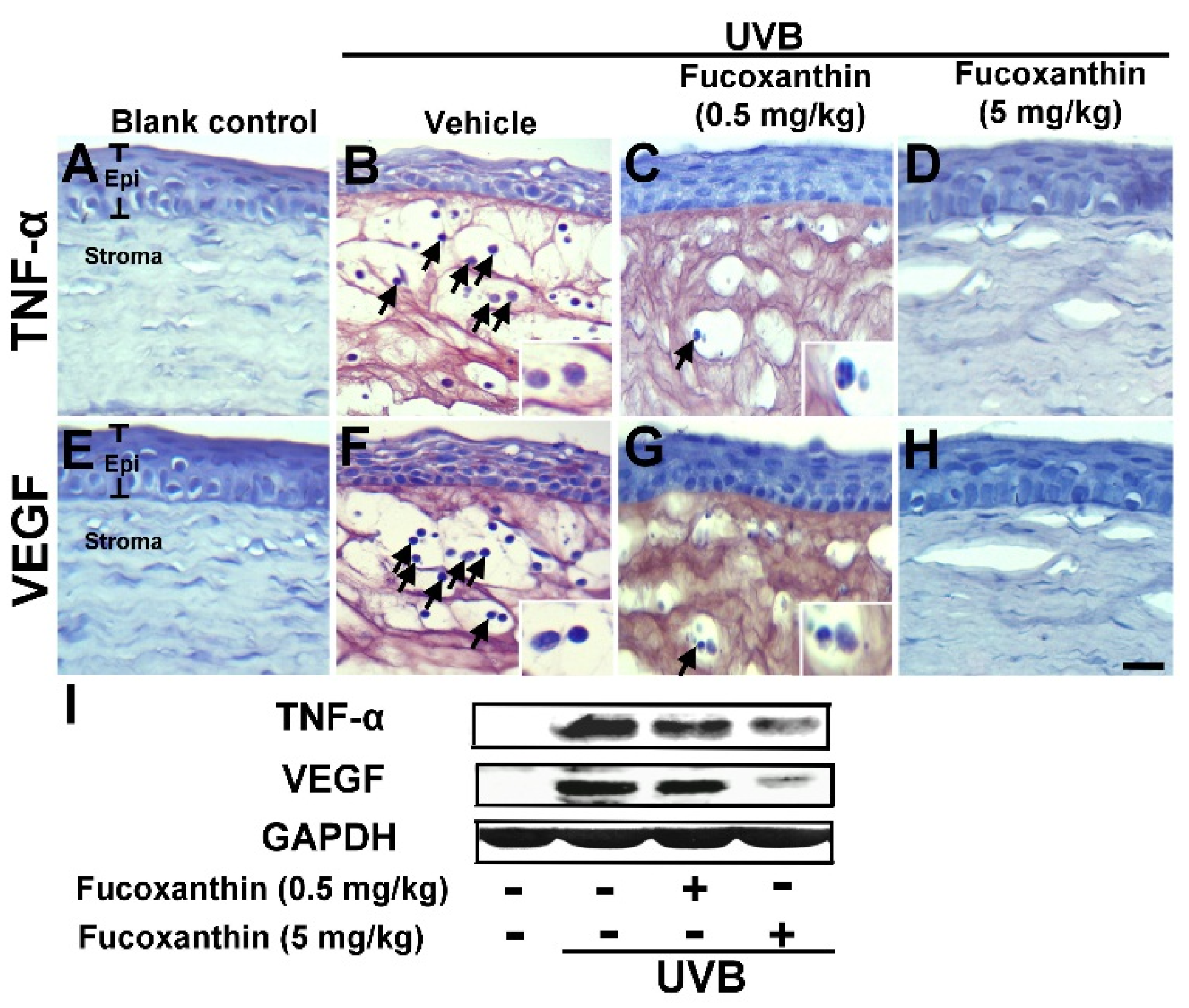
2.2. Inhibition of TNF-α and VEGF Activation Induced by UVB Irradiation with Fucoxanthin

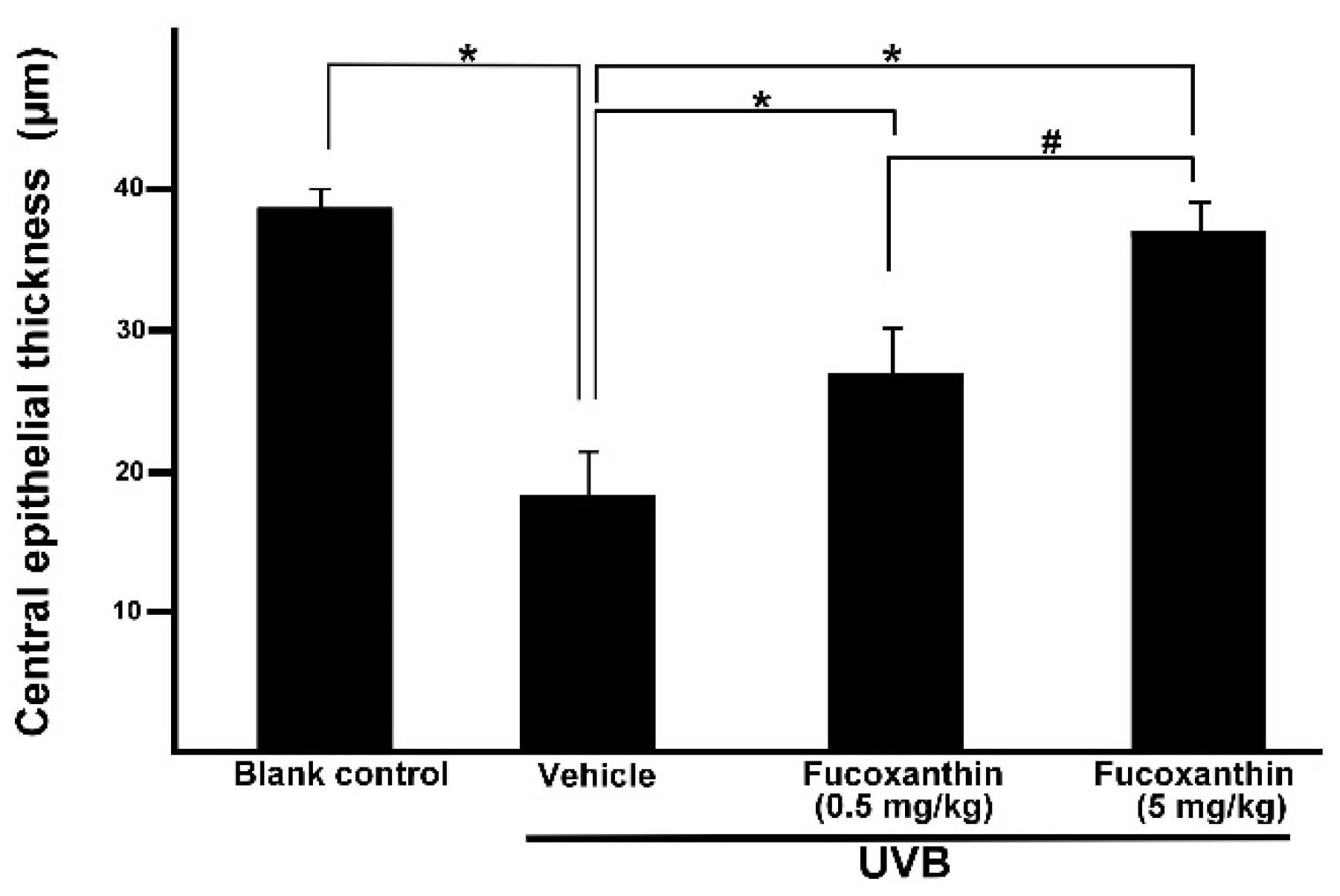
2.3. Effect of Fucoxanthin on the Thickness of Corneal Epithelial and Stroma Layers
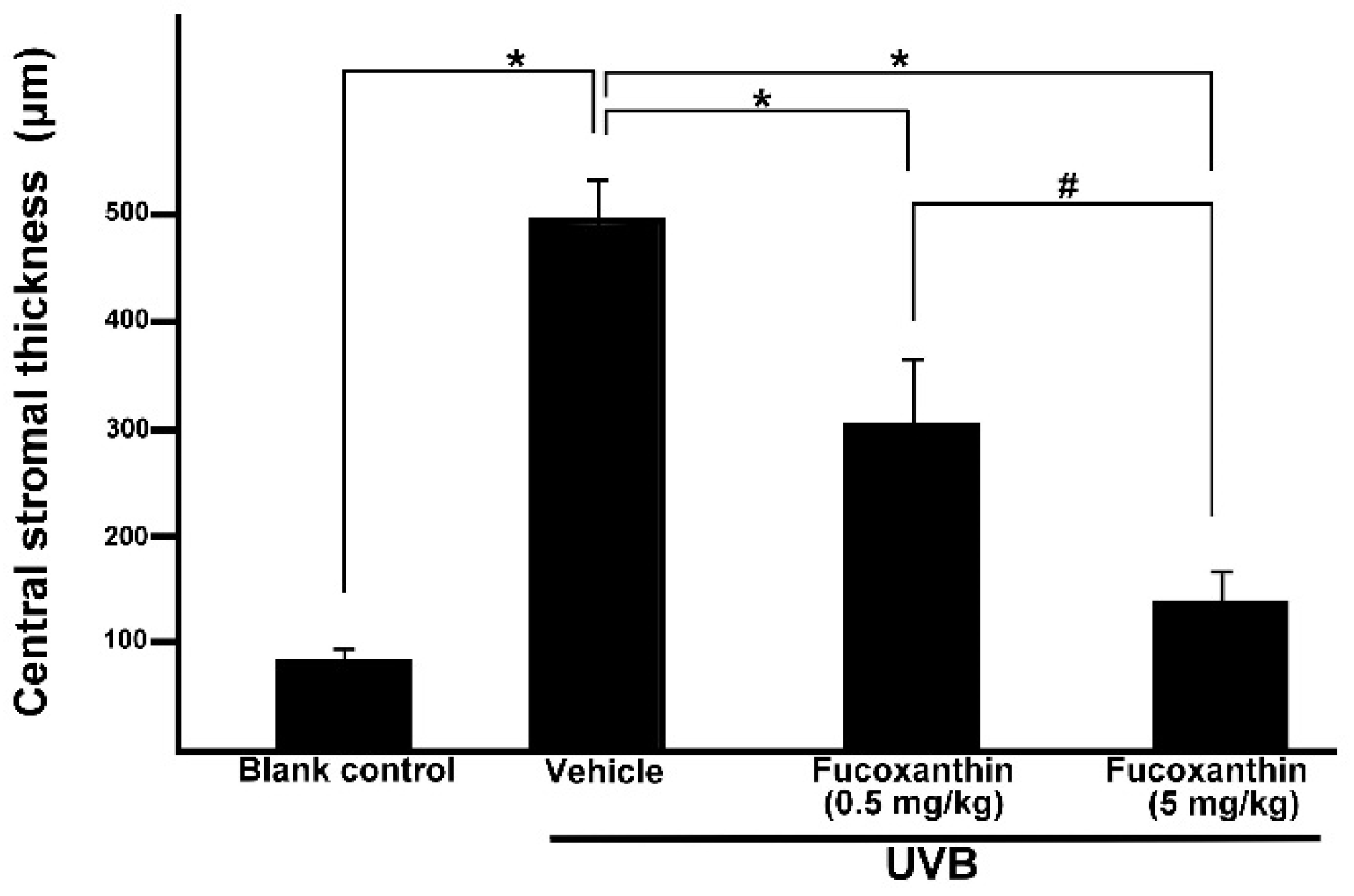
3. Discussions
4. Experimental Section
4.1. Experimental Animals
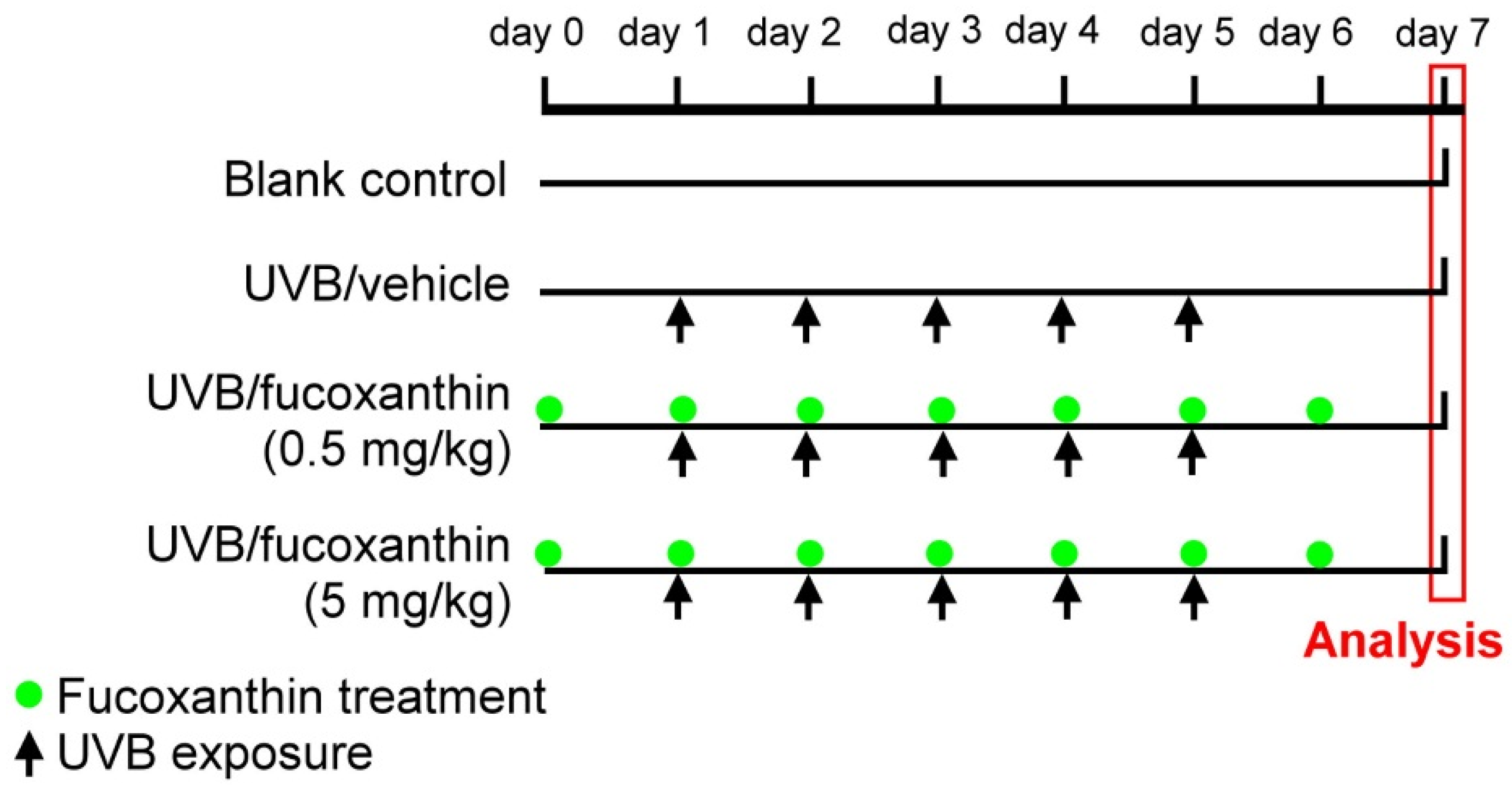
4.2. UVB-Induced Corneal Disorders and Fucoxanthin Treatment
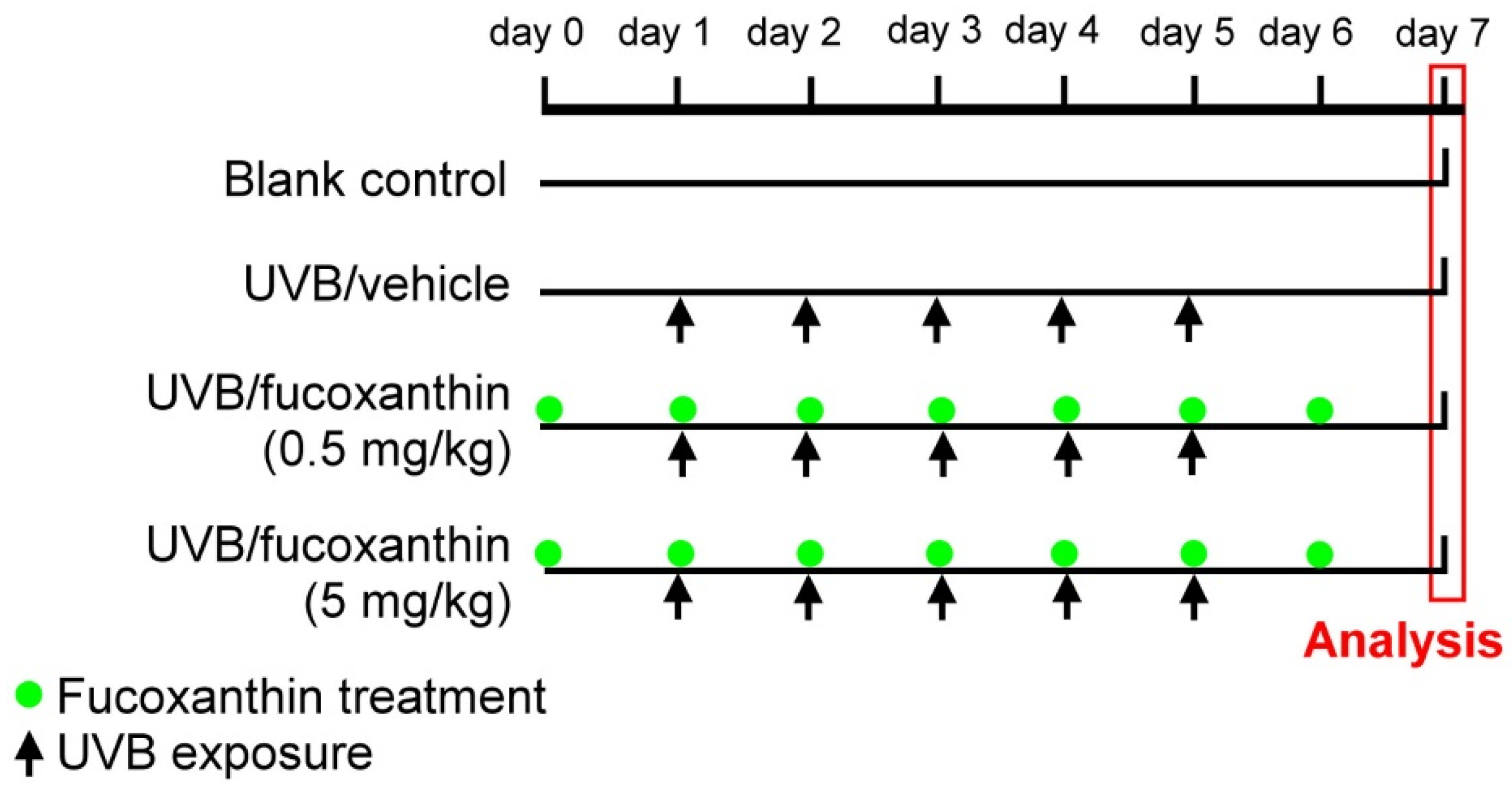
4.3. Determination of Corneal Disorders
4.4. Histopathological Analysis and Immunohistochemistry of Inflammatory Reaction in the Cornea
4.5. Western Blot Analysis
4.6. Corneal Thickness Measurements
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Diffey, B.L. Sources and measurement of ultraviolet radiation. Methods 2002, 28, 4–13. [Google Scholar] [CrossRef]
- Kolozsvári, L.; Nogradi, A.; Hopp, B.; Bor, Z. UV absorbance of the human cornea in the 240- to 400-nm range. Investig. Ophthalmol. Vis. Sci. 2002, 43, 2165–2168. [Google Scholar] [PubMed]
- Schein, O.D. Phototoxicity and the cornea. J Natl. Med. Assoc. 1992, 84, 579–583. [Google Scholar] [PubMed]
- Kennedy, M.; Kim, K.H.; Harten, B.; Brown, J.; Planck, S.; Meshul, C.; Edelhauser, H.; Rosenbaum, J.T.; Armstrong, C.A.; Ansel, J.C. Ultraviolet irradiation induces the production of multiple cytokines by human corneal cells. Investig. Ophthalmol. Vis. Sci. 1997, 38, 2483–2491. [Google Scholar]
- Alexander, G.; Carlsen, H.; Blomhoff, R. Corneal NF-kappaB activity is necessary for the retention of transparency in the cornea of UV-B-exposed transgenic reporter mice. Exp. Eye Res. 2006, 82, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Pauloin, T.; Dutot, M.; Joly, F.; Warnet, J.M.; Rat, P. High molecular weight hyaluronan decreases UVB-induced apoptosis and inflammation in human epithelial corneal cells. Mol. Vis. 2009, 15, 577–583. [Google Scholar] [PubMed]
- Kimura, K.; Teranishi, S.; Fukuda, K.; Kawamoto, K.; Nishida, T. Delayed disruption of barrier function in cultured human corneal epithelial cells induced by tumor necrosis factor-alpha in a manner dependent on NF-kappaB. Investig. Ophthalmol. Vis Sci. 2008, 49, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Golu, A.; Gheorghişor, I.; Bălăşoiu, A.T.; Baltă, F.; Osiac, E.; Mogoantă, L.; Bold, A. The effect of ultraviolet radiation on the cornea-experimental study. Rom. J Morphol. Embryol. 2013, 54, 1115–1120. [Google Scholar] [PubMed]
- Chang, J.H.; Gabison, E.E.; Kato, T.; Azar, D.T. Corneal neovascularization. Curr. Opin. Ophthalmol. 2001, 12, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Rubio, R.G.; Adamis, A.P. Ocular Angiogenesis: Vascular Endothelial Growth Factor and Other Factors. Dev. Ophthalmol. 2016, 55, 28–37. [Google Scholar] [PubMed]
- Bilgihan, A.; Bilgihan, K.; Yis, O.; Sezer, C.; Akyol, G.; Hasanreisoglu, B. Effects of topical vitamin E on corneal superoxide dismutase, glutathione peroxidase activities and polymorphonuclear leucocyte infiltration after photorefractive keratectomy. Acta Ophthalmol. Scand. 2003, 81, 177–180. [Google Scholar] [CrossRef]
- Nomura, T.; Kikuchi, M.; Kubodera, A.; Kawakami, Y. Proton-donative antioxidant activity of fucoxanthin with 1,1-diphenyl-2-picrylhydrazyl (DPPH). Biochem. Mol. Bio. Int. 1997, 42, 361–370. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Maoka, T. Allenic and cumulenic lipids. Prog. Lipid Res. 2007, 46, 328–375. [Google Scholar] [CrossRef] [PubMed]
- Fung, A.; Hamid, N.; Lu, J. Fucoxanthin content and antioxidant properties of Undaria pinnatifida. Food Chem. 2013, 136, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Terasaki, M.; Baba, Y.; Yasui, H.; Saga, N.; Hosokawa, M.; Miyashita, K. Evaluation of recoverable functional lipid components with special reference to fucoxanthin and fucosterol contents of several brown seaweeds of Japan. J. Phycol. 2009, 45, 974–980. [Google Scholar] [CrossRef]
- Asai, A.; Sugawara, T.; Ono, H.; Nagao, A. Biotransformation of fucoxanthinol into amarouciaxanthin A in mice and HepG2 cells: Formation and cytotoxicity of fucoxanthinmetabolites. Drug Metab. Dispos. 2004, 32, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, T.; Matsubara, K.; Akagi, R.; Mori, M.; Hirata, T. Antiangiogenic activity of brown algae fucoxanthin and its deacetylated product, fucoxanthinol. J. Agric. Food Chem. 2006, 54, 9805–9810. [Google Scholar] [CrossRef] [PubMed]
- Afolayan, A.F.; Bolton, J.J.; Lategan, C.A.; Smith, P.J.; Beukes, D.R. Fucoxanthin, tetraprenylated toluquinone and toluhydroquinone metabolites from Sargassum heterophyllum inhibit the in vitro growth of the malaria parasite Plasmodium falciparum. Z. Naturforsch. C 2008, 63, 848–852. [Google Scholar] [CrossRef]
- Woo, M.N.; Jeon, S.M.; Shin, Y.C.; Lee, M.K.; Kang, M.A.; Choi, M.S. Anti-obese property of fucoxanthin is partly mediated by altering lipid-regulating enzymes and uncoupling proteins of visceral adipose tissue in mice. Mol. Nutr. Food Res. 2009, 53, 1603–1611. [Google Scholar] [CrossRef] [PubMed]
- Beppu, F.; Hosokawa, M.; Yim, M.J.; Shinoda, T.; Miyashita, K. Down-regulation of hepatic stearoyl-CoA desaturase-1 expression by fucoxanthin via leptin signaling in diabetic/obese KK-A(y) mice. Lipids 2013, 48, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Rengarajan, T.; Rajendran, P.; Nandakumar, N.; Balasubramanian, M.P.; Nishigaki, I. Cancer preventive efficacy of marine carotenoid fucoxanthin: Cell cycle arrest and apoptosis. Nutrients 2013, 5, 4978–4989. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.P.; Hou, Y.H. First Evidence for the Anti-inflammatory Activity of Fucoxanthin in High-Fat-Diet-Induced Obesity in Mice and the Antioxidant Functions in PC12 Cells. Inflammation 2014, 37, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Morandi, A.C.; Molina, N.; Guerra, B.A.; Bolin, A.P.; Otton, R. Fucoxanthin in association with vitamin C acts as modulators of human neutrophil function. Eur. J. Nutr. 2014, 53, 779–792. [Google Scholar] [CrossRef]
- Pourzand, C.; Tyrrell, R.M. Apoptosis, the role of oxidative stress and the example of solar UV radiation. Photochem. Photobiol. 1999, 70, 380–390. [Google Scholar] [CrossRef]
- Urikura, I.; Sugawara, T.; Hirata, T. Protective effect of fucoxanthin against UVB-induced skin photoaging in hairless mice. Biosci. Biotechnol. Biochem. 2011, 75, 757–760. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, H.; Yoshii, Y.; Asai, T.; Furukawa, T.; Takaichi, S.; Fujibayashi, Y. Photo-excitation of carotenoids causes cytotoxicity via singlet oxygen production. Biochem. Biophys. Res. Commun. 2012, 417, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Petrescu, M.S.; Larry, C.L.; Bowden, R.A.; Williams, G.W.; Gagen, D.; Li, Z.; Smith, C.W.; Burns, A.R. Neutrophil interactions with keratocytes during corneal epithelial wound healing: A role for CD18 integrins. Investig. Ophthalmol. Vis. Sci. 2007, 48, 5023–5029. [Google Scholar] [CrossRef] [PubMed]
- Edelman, J.L.; Castro, M.R.; Wen, Y. Correlation of VEGF expression by leukocytes with the growth and regression of blood vessels in the rat cornea. Investig. Ophthalmol. Vis. Sci. 1999, 40, 1112–1123. [Google Scholar]
- Imanishi, J.; Kamiyama, K.; Iguchi, I.; Kita, M.; Sotozono, C.; Kinoshita, S. Growth factors: Importance in wound healing and maintenance of transparency of the cornea. Prog. Retin. Eye Res. 2000, 19, 113–129. [Google Scholar] [CrossRef]
- Lai, C.M.; Spilsbury, K.; Brankov, M.; Zaknich, T.; Rakoczy, P.E. Inhibition of corneal neovascularization by recombinant adenovirus mediated antisense. VEGF RNA. Exp. Eye Res. 2002, 75, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Klenkler, B.; Sheardown, H. Growth factors in the anterior segment: Role in tissue maintenance, wound healing and ocular pathology. Exp. Eye Res. 2004, 79, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, S.; Xu, S.; Yu, X.; Ma, D.; Hu, X.; Cao, X. In vivo induction of apoptosis by fucoxanthin, a marine carotenoid, associated with down-regulating STAT3/EGFR signaling in sarcoma 180 (S180) xenografts-bearing mice. Mar. Drugs 2012, 10, 2055–2068. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.N.; Heo, S.J.; Yoon, W.J.; Kang, S.M.; Ahn, G.; Yi, T.H.; Jeon, Y.J. Fucoxanthin inhibits the inflammatory response by suppressing the activation of NF-κB and MAPKs in lipopolysaccharide-induced RAW 264.7 macrophages. Eur. J. Pharmacol. 2010, 649, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Shiratori, K.; Ohgami, K.; Ilieva, I.; Jin, X.H.; Koyama, Y.; Miyashita, K.; Yoshida, K.; Kase, S.; Ohno, S. Effects of fucoxanthin on lipopolysaccharide-induced inflammation in vitro and in vivo. Exp. Eye Res. 2005, 81, 422–428. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.Y.; Lin, D.P.; Wu, C.Y.; Teng, M.C.; Sun, C.Y.; Tsai, Y.T.; Su, K.C.; Wang, S.R.; Chang, H.H. Dietary zerumbone prevents mouse cornea from UVB-induced photokeratitis through inhibition of NF-κB, iNOS, and TNF-α expression and reduction of MDA accumulation. Mol. Vis. 2011, 17, 854–863. [Google Scholar] [PubMed]
- Lennikov, A.; Kitaichi, N.; Fukase, R.; Murata, M.; Noda, K.; Ando, R.; Ohguchi, T.; Kawakita, T.; Ohno, S.; Ishida, S. Amelioration of ultraviolet-induced photokeratitis in mice treated with astaxanthin eye drops. Mol. Vis. 2012, 18, 455–464. [Google Scholar] [PubMed]
- Tsai, C.F.; Lu, F.J.; Hsu, Y.W. Protective effects of Dunaliella salina—A carotenoids-rich alga—Against ultraviolet B-induced corneal oxidative damage in mice. Mol. Vis. 2012, 18, 1540–1547. [Google Scholar] [PubMed]
- Tanito, M.; Takanashi, T.; Kaidzu, S.; Yoshida, Y.; Ohira, A. Cytoprotective effects of rebamipide and carteolol hydrochloride against ultraviolet B-induced corneal damage in mice. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2980–2985. [Google Scholar] [CrossRef] [PubMed]
- Kim, J. The use of vital dyes in corneal disease. Curr. Opin. Ophthalmol. 2000, 11, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Li, S.; Nikulina, K.; Porco, T.; Gallup, M.; McNamara, N. Immune profile of squamous metaplasia development in autoimmune regulator-deficient dry eye. Mol. Vis. 2009, 15, 563–576. [Google Scholar] [PubMed]
- Lively, G.D.; Jiang, B.; Hedberg-Buenz, A.; Chang, B.; Petersen, G.E.; Wang, K.; Kuehn, M.H.; Anderson, M.G. Genetic dependence of central corneal thickness among inbred strains of mice. Investig. Ophthalmol. Vis. Sci. 2010, 51, 160–171. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-J.; Lee, C.-J.; Lin, T.-B.; Liu, H.-J.; Huang, S.-Y.; Chen, J.-Z.; Tseng, K.-W. Inhibition of Ultraviolet B-Induced Expression of the Proinflammatory Cytokines TNF-α and VEGF in the Cornea by Fucoxanthin Treatment in a Rat Model. Mar. Drugs 2016, 14, 13. https://doi.org/10.3390/md14010013
Chen S-J, Lee C-J, Lin T-B, Liu H-J, Huang S-Y, Chen J-Z, Tseng K-W. Inhibition of Ultraviolet B-Induced Expression of the Proinflammatory Cytokines TNF-α and VEGF in the Cornea by Fucoxanthin Treatment in a Rat Model. Marine Drugs. 2016; 14(1):13. https://doi.org/10.3390/md14010013
Chicago/Turabian StyleChen, Shiu-Jau, Ching-Ju Lee, Tzer-Bin Lin, Hsiang-Jui Liu, Shuan-Yu Huang, Jia-Zeng Chen, and Kuang-Wen Tseng. 2016. "Inhibition of Ultraviolet B-Induced Expression of the Proinflammatory Cytokines TNF-α and VEGF in the Cornea by Fucoxanthin Treatment in a Rat Model" Marine Drugs 14, no. 1: 13. https://doi.org/10.3390/md14010013
APA StyleChen, S.-J., Lee, C.-J., Lin, T.-B., Liu, H.-J., Huang, S.-Y., Chen, J.-Z., & Tseng, K.-W. (2016). Inhibition of Ultraviolet B-Induced Expression of the Proinflammatory Cytokines TNF-α and VEGF in the Cornea by Fucoxanthin Treatment in a Rat Model. Marine Drugs, 14(1), 13. https://doi.org/10.3390/md14010013