
Positional Distribution of Fatty Acids in Triacylglycerols and Phospholipids from Fillets of Atlantic Salmon (Salmo Salar) Fed Vegetable and Fish Oil Blends

 and
and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Lipid Content and Fatty Acid Compositions of Fillets

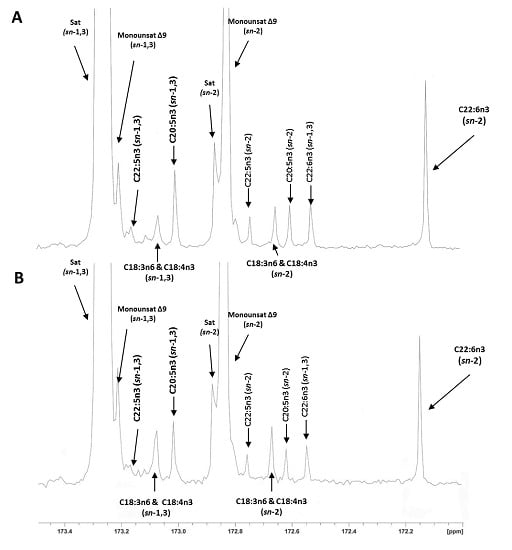
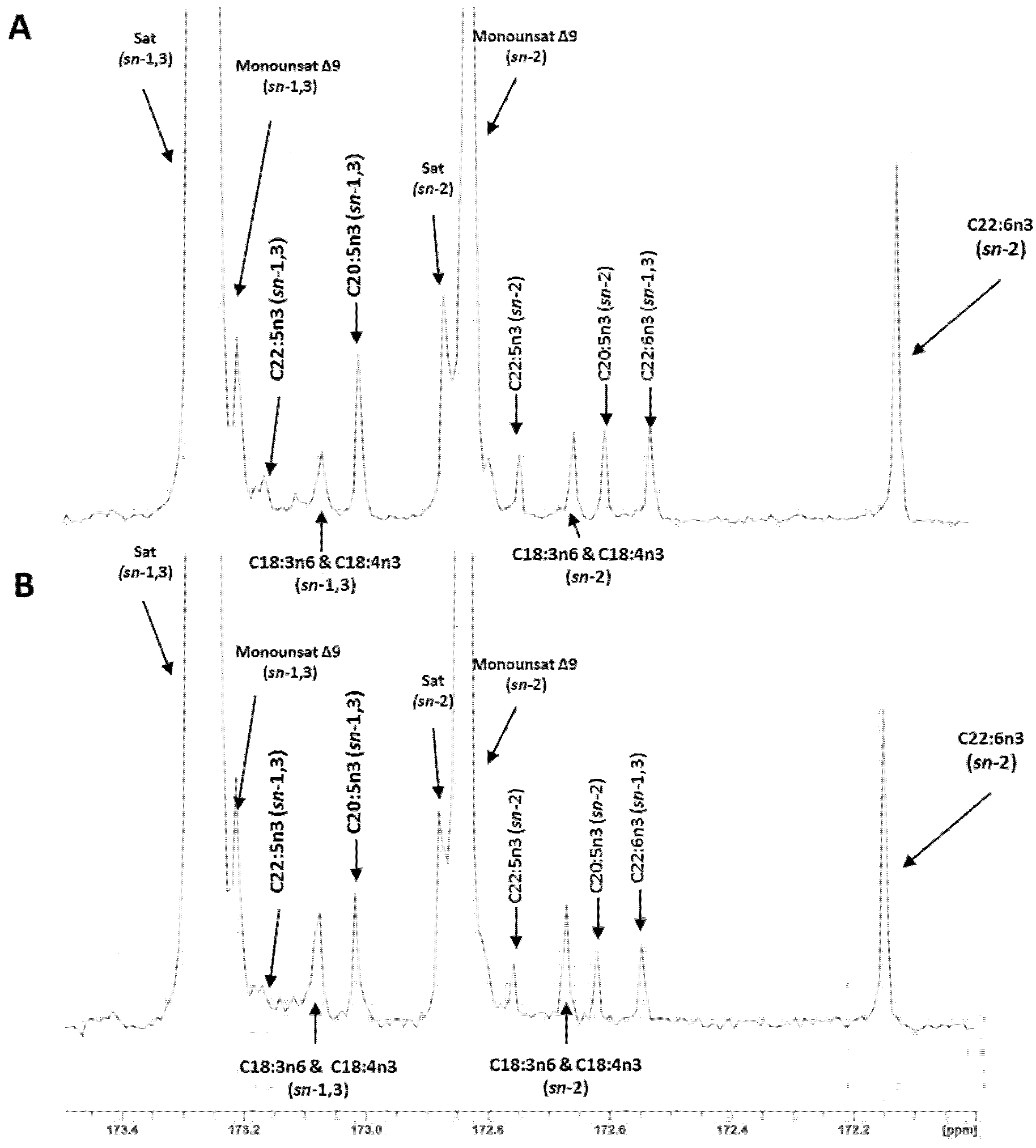
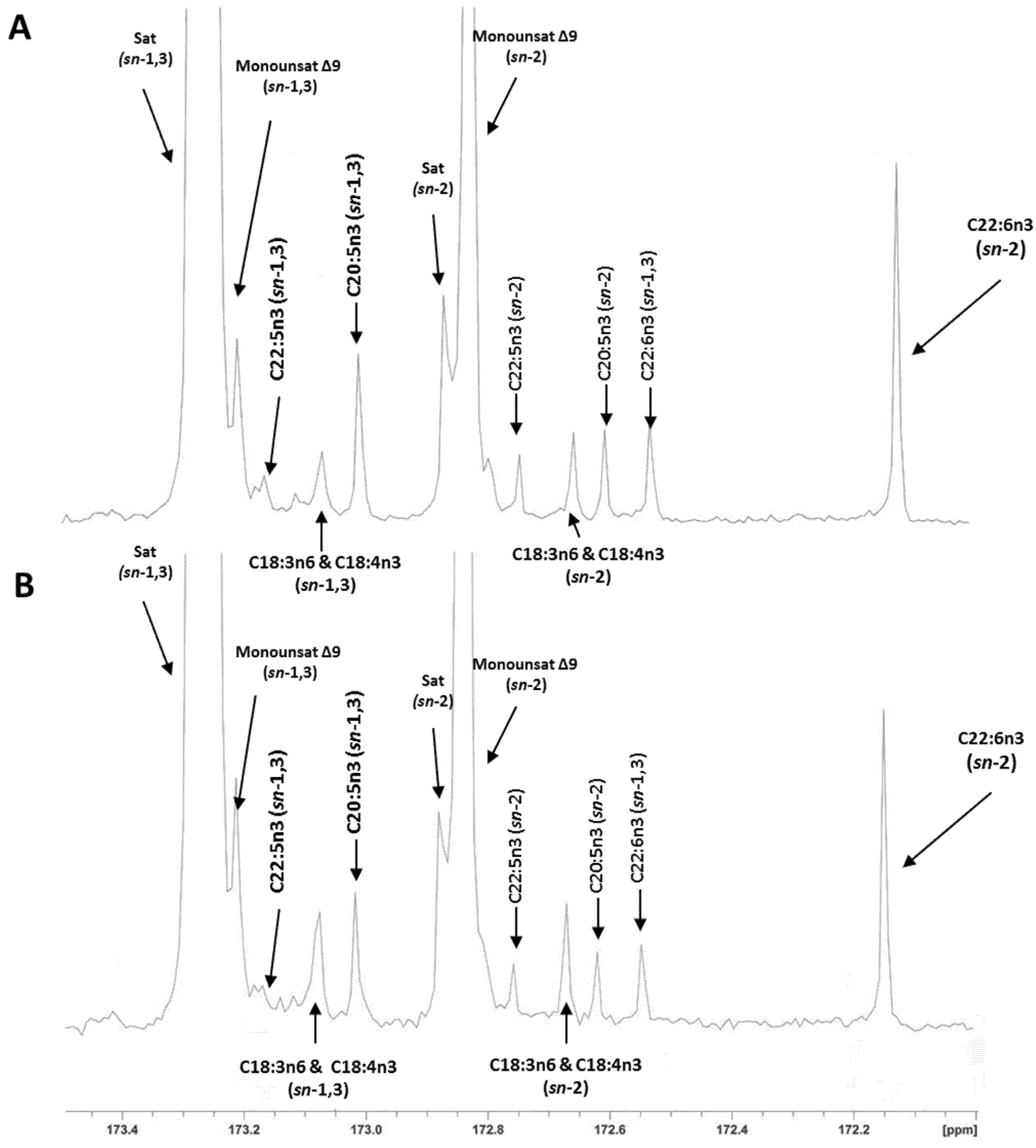
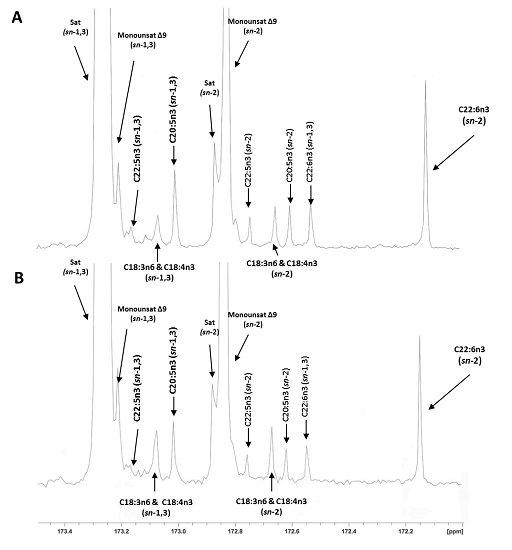{kind=link}
{kind=link}
| H-ED | L-ED | p-Value a | |
|---|---|---|---|
| C14:0 | 3.8 ± 0.1 | 1.9 ± 0.1 | 0.012 |
| C16:0 | 14.8 ± 0.6 | 15.2 ± 0.6 | 0.685 |
| C16:1n-7 | 3.3 ± 0.1 | 1.8 ± 0.1 | 0.021 |
| C18:0 | 3.4 ± 0.1 | 3.5 ± 0.1 | 0.581 |
| C18:1n-9 | 35.7 ± 0.5 | 44.4 ± 0.4 | 0.061 |
| C18:1n-7 | 2.2 ± 0.3 | 1.7 ± 0.1 | 0.056 |
| C18:2n-6 | 11.4 ± 0.4 | 13.3 ± 0.4 | 0.095 |
| C18:3n-3 | 3.4 ± 0.1 | 3.3 ± 0.1 | 0.732 |
| C18:4n-3 | 1.0 ± 0.1 | 0.8 ± 0.1 | 0.127 |
| C20:1n-9 | 6.9 ± 0.2 | 4.5 ± 0.2 | 0.021 |
| C20:2n-6 | 1.0 ± 0.0 | 1.0 ± 0.0 | 0.283 |
| C20:3n-6 | 0.4 ± 0.0 | 0.8 ± 0.0 | 0.013 |
| C20:4n-6 | 0.3 ± 0.0 | 0.3 ± 0.0 | 0.681 |
| C20:3n-3 | 0.3 ± 0.0 | 0.2 ± 0.0 | 0.105 |
| C20:4n-3 | 0.9 ± 0.0 | 0.5 ± 0.0 | 0.019 |
| C20:5n-3 | 3.1 ± 0.1 | 1.5 ± 0.1 | 0.018 |
| C22:1 | 7.6 ± 0.3 | 3.3 ± 0.3 | 0.010 |
| C22:5n-3 | 1.3 ± 0.1 | 0.6 ± 0.1 | 0.024 |
| C22:6n-3 | 8.1 ± 0.1 | 4.9 ± 0.1 | 0.005 |
| ∑SFA b | 22.5 ± 0.9 | 21.1 ± 0.9 | 0.398 |
| ∑MUFA c | 56.1 ± 2.3 | 56.2 ± 2.3 | 0.989 |
| ∑n-3 d | 18.3 ± 0.4 | 12.1 ± 0.4 | 0.009 |
| ∑n-6 e | 13.5 ± 0.5 | 16.1 ± 0.5 | 0.070 |
| n3/n6 | 1.4 ± 0.01 | 0.8 ± 0.01 | 0.001 |
| %TL (FW) | 11.0 ± 0.4 | 10.5 ± 0.4 | 0.469 |
2.2. Fillet Lipid Class Composition, TAG, and PC Fatty Acid Analysis
| H-ED | L-ED | p-Value b | |
|---|---|---|---|
| TAG | 92.3 ± 1.0 | 92.5 ± 1.0 | 0.864 |
| DG | 0.8 ± 0.1 | 0.7 ± 0.1 | 0.786 |
| FFA | 0.9 ± 0.2 | 1.0 ± 0.2 | 0.653 |
| PC | 3.7 ± 0.3 | 3.7 ± 0.3 | 0.927 |
| PE | 1.7 ± 0.1 | 1.6 ± 0.1 | 0.943 |
| PI | 0.3 ± 0.0 | 0.3 ± 0.0 | 0.642 |
| PS | 0.3 ± 0.0 | 0.2 ± 0.0 | 0.682 |
| TAG | PC | |||||
|---|---|---|---|---|---|---|
| H-ED | L-ED | p-Value a | H-ED | L-ED | p-Value a | |
| C14:0 | 3.7 ± 0.0 | 1.9 ± 0.0 | 0.0003 | 2.2 ± 0.1 | 1.0 ± 0.1 | 0.003 |
| C16:0 | 13.1 ± 0.3 | 14.0 ± 0.2 | 0.148 | 20.8 ± 0.2 | 22.5 ± 0.2 | 0.022 |
| C16:1n-7 | 3.2 ± 0.1 | 2.0 ± 0.0 | 0.003 | 1.4 ± 0.1 | 0.7 ± 0.1 | 0.018 |
| C18:0 | 2.7 ± 0.1 | 3.2 ± 0.1 | 0.053 | 0.9 ± 0.1 | 1.0 ± 0.1 | 0.581 |
| C18:1n-9 | 32.5 ± 0.6 | 42.1 ± 0.5 | 0.005 | 11.7 ± 0.1 | 15.4 ± 0.1 | 0.010 |
| C18:2n-6 | 10.9 ± 0.2 | 12.9 ± 0.2 | 0.015 | 3.8 ± 0.2 | 7.2 ± 0.2 | 0.008 |
| C18:3n-3 | 3.2 ± 0.0 | 3.3 ± 0.0 | 0.024 | 2.0 ± 0.1 | 2.7 ± 0.1 | 0.025 |
| C20:1n-9 | 6.4 ± 0.1 | 4.3 ± 0.1 | 0.001 | 0.8 ± 0.0 | 0.5 ± 0.0 | 0.031 |
| C20:2n-6 | 0.8 ± 0.0 | 0.9 ± 0.0 | 0.365 | 0.5 ± 0.0 | 0.6 ± 0.0 | 0.095 |
| C20:3n-6 | 0.3 ± 0.0 | 0.6 ± 0.0 | 0.016 | 0.6 ± 0.1 | 2.3 ± 0.1 | 0.004 |
| C20:4n-6 | 0.2 ± 0.0 | 0.2 ± 0.0 | 0.249 | 0.7 ± 0.0 | 1.3 ± 0.0 | 0.005 |
| C20:4n-3 | 0.8 ± 0.0 | 0.5 ± 0.0 | 0.008 | 0.9 ± 0.0 | 0.9 ± 0.0 | 0.935 |
| C20:5n-3 | 2.5 ± 0.1 | 1.2 ± 0.1 | 0.008 | 10.8 ± 0.4 | 8.1 ± 0.3 | 0.031 |
| C22:1 | 7.1 ± 0.1 | 3.3 ± 0.1 | 0.0009 | 0.3 ± 0.0 | 0.3 ±0.0 | 0.770 |
| C22:5n-3 | 1.0 ± 0.0 | 0.5 ± 0.0 | 0.018 | 2.4 ± 0.2 | 2.1 ± 0.2 | 0.295 |
| C22:6n-3 | 5.1 ± 0.2 | 2.9 ± 0.2 | 0.010 | 34.5 ± 01.0 | 28.0 ± 0.9 | 0.040 |
| ∑SFA b | 19.9 ± 0.3 | 19.6 ± 0.2 | 0.474 | 24.0 ± 0.2 | 24.7 ± 0.2 | 0.168 |
| ∑MUFA c | 52.1 ± 0.4 | 54.6 ± 0.4 | 0.037 | 15.8 ± 0.3 | 18.3 ± 0.3 | 0.034 |
| ∑n-3 d | 14.4 ± 0.2 | 9.6 ± 0.2 | 0.001 | 51.9 ± 0.5 | 44.4 ± 0.5 | 0.010 |
| ∑n-6 e | 12.2 ± 0.1 | 14.8 ± 0.2 | 0.002 | 5.9 ± 0.31 | 11.7 ± 0.28 | 0.005 |
| n3/n6 | 1.2 ± 0.0 | 0.65 ± 0.0 | 0.001 | 8.7 ± 0.25 | 3.8 ± 0.23 | 0.004 |
2.3. Regiospecific Analysis of Salmon Fillet Triacylclycerol and Phosphatidylcholine Classes
| EPA | DHA | |
|---|---|---|
| Diet | ||
| H-ED | 50.1 ± 1.3 | 84.9 ± 0.7 |
| L-ED | 49.3 ± 1.3 | 83.1 ± 0.7 |
| Method | ||
| Lipase | 50.1 ± 1.1 | 81.7 ± 0.7 |
| NMR | 49.2 ± 1.0 | 86.3 ± 0.6 |
| Diet × Method | ||
| H-ED Lipase | 50.7 ± 1.6 | 83.5 ± 1.0 |
| H-ED NMR | 49.4 ± 1.4 | 86.3 ± 0.8 |
| L-ED Lipase | 49.4 ± 1.4 | 79.9 ± 1.0 |
| L-ED NMR | 49.1 ± 1.4 | 86.4 ± 0.8 |
| Probability a | ||
| Diet | 0.7286 | 0.1377 |
| Method | 0.4323 | 0.0020 |
| Diet × Method | 0.6219 | 0.0758 |
| H-ED | L-ED | p-Value a | |
|---|---|---|---|
| C14:0 | 21.3 ± 2.6 | 19.5 ± 2.6 | 0.680 |
| C16:0 | 16.4 ± 1.9 | 13.3 ± 1.9 | 0.378 |
| C18:1n-9 | 52.6 ± 1.4 | 57.4 ± 1.4 | 0.150 |
| C18:2n-6 | 45.5 ± 2.3 | 58.1 ± 2.3 | 0.063 |
| C18:3n-3 | 25.7 ± 1.0 | 31.2 ± 1.0 | 0.063 |
| C20:3n-6 | 53.5 ± 1.3 | 66.3 ± 1.3 | 0.022 |
| C20:4n-6 | 89.1 ± 0.8 | 93.2 ± 0.8 | 0.080 |
| C20:5n-3 | 77.4 ± 0.9 | 83.5 ± 0.9 | 0.040 |
| C22:5n-3 | 80.3 ± 1.3 | 85.7 ± 1.3 | 0.099 |
| C22:6n-3 | 89.7 ± 0.9 | 91.6 ± 0.9 | 0.312 |
3. Experimental Section
3.1. Animals and Diets
| Ingredients (as Fed Basis g/kg) | H-ED | L-ED |
|---|---|---|
| Wheat a | 35.0 | 35.0 |
| Wheat gluten b | 149.6 | 149.9 |
| Sunflower meal c | 88.4 | 84.8 |
| Fava beans, dehulled d | 60.0 | 60.0 |
| Soy Protein Concentrate e | 310.0 | 310.0 |
| Fishmeal NA f | 100.0 | 100.0 |
| Palm oil g | 2.1 | 46.7 |
| Rapeseed oil h | 91.9 | 146.5 |
| Fish oil NA i | 123.1 | 26.1 |
| Astaxanthin 10% j | 0.4 | 0.4 |
| Vitamin and mineral mix k | 32.1 | 32.1 |
| Water | 7.1 | 7.4 |
| Analyzed composition | ||
| Moisture, % | 6.4 | 6.6 |
| Total fat, % | 25.1 | 25.5 |
| Crude Protein, % | 45.2 | 45.2 |
| Ash, % | 4.5 | 4.6 |
| Fatty acids (g/100 g total fatty acids) | ||
| C14:0 | 3.8 | 1.7 |
| C16:0 | 10.6 | 13.0 |
| C18:1n-9 | 27.3 | 38.8 |
| C18:2n-6 | 11.8 | 16.3 |
| C18:3n-3 | 4.2 | 5.2 |
| C20:5n-3 | 4.8 | 2.1 |
| C22:6n-3 | 5.5 | 2.5 |
| ∑SFA l | 17.7 | 17.8 |
| ∑MUFA m | 47.2 | 49.8 |
| ∑ (n-3) | 17.2 | 10.8 |
| ∑ (n-6) | 12.8 | 16.7 |
| n-3/n-6 | 1.34 | 0.64 |
3.2. Sampling
3.3. Feed Analysis
3.4. Tissue Total Lipid Extraction and Analysis
3.5. Lipid Classes Analysis
3.6. Positional Analysis of TAG
3.6.1. 13C NMR Spectroscopy
3.6.2. Enzymatic Analysis
3.7. Positional Analysis of PC
3.8. Fatty Acid Methyl Esters Analysis
3.9. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Miller, M.R.; Nichols, P.D.; Barnes, J.; Davies, N.W.; Peacock, E.J.; Carter, C.G. Regiospecificity profiles of storage and membrane lipids from the gill and muscle tissue of Atlantic salmon (Salmo salar L.) grown at elevated temperature. Lipids 2006, 41, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.E.E. Triacylglycerol structure and interesterification of palmitic and stearic acid-rich fats: An overview and implications for cardiovascular disease. Nutr. Res. Rev. 2009, 22, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Hunter, J.E. Studies on effects of dietary fatty acids as related to their position on triglycerides. Lipids 2001, 36, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Thies, F.; Garry, J.M.C.; Yaqoob, P.; Rerkasem, K.; Williams, J.; Shearman, C.P.; Gallagher, P.J.; Calder, P.C.; Grimble, R.F. Association of n-3 polyunsaturated fatty acids with stability of atherosclerotic plaques: A randomised controlled trial. Lancet 2003, 361, 477–485. [Google Scholar] [CrossRef]
- Abeywardena, M.Y.; Patten, G.S. Role of ω3 long-chain polyunsaturated fatty acids in reducing cardio-metabolic risk factors. Endocr. Metab. Immun. Disord. Drug Targ. 2011, 11, 232–246. [Google Scholar] [CrossRef]
- Siriwardhana, N.; Kalupahana, N.S.; Moustaid-Moussa, N. Health benefits of n-3 polyunsaturated fatty acids: Eicosapentaenoic acid and docosahexaenoic acid. Adv. Food Nutr. Res. 2012, 65, 211–222. [Google Scholar] [PubMed]
- Yoshinaga, K.; Sasaki, K.; Watanabe, H.; Nagao, K.; Inoue, N.; Shirouchi, B.; Yanagita, T.; Nagai, T.; Mizobe, H.; Kojima, K.; et al. Differential effects of triacylglycerol positional isomers containing n-3 series highly unsaturated fatty acids on lipid metabolism in C57BL/6J mice. J. Nutr. Biochem. 2015, 26, 431–432. [Google Scholar] [CrossRef]
- Torstensen, B.E.; Bell, J.G.; Rosenlund, G.; Henderson, R.J.; Graff, I.E.; Tocher, D.R.; Lie, O.; Sargent, J.R. Tailoring of Atlantic salmon (Salmo salar L.) flesh lipid composition and sensory quality by replacing fish oil with a vegetable oil blend. J. Agric. Food Chem. 2005, 53, 10166–10178. [Google Scholar] [CrossRef] [PubMed]
- Menoyo, D.; Lopez-Bote, C.J.; Diez, A.; Obach, A.; Bautista, J.M. Impact of n-3 fatty acid chain length and n-3/n-6 ratio in Atlantic salmon (Salmo salar) diets. Aquaculture 2007, 267, 248–259. [Google Scholar] [CrossRef]
- Sørensen, M.; Berge, G.M.; Thomassen, M.S.; Ruyter, B.; Hatlen, B.; Ytrestøyl, T.; Synnøve, T.; Ǻsgard, T. Today’s and Tomorrow's Feed Ingredients in Norwegian Aquaculture; Nofima AS: Trømso, Norway, 2011; pp. 1–32. [Google Scholar]
- Turchini, G.M.; Torstensen, B.E.; Ng, W.K. Fish oil replacement in finfish nutrition. Rev. Aquacult. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Segura, J.; Ruiz-López, N.; Menoyo, D.; Cambero, M.I.; López-Bote, C.J. Comparison of analytical techniques for the determination of the positional distribution of fatty acids in triacylglycerols. Relationship with pig fat melting point and hardness. Grasas Aceites 2015, 66, 2. [Google Scholar] [CrossRef]
- Standal, I.B.; Axelson, D.E.; Aursand, M. Differentiation of fish oils according to species by 13C-NMR regiospecific analyses of triacyglycerols. J. Am. Oil Chem. Soc. 2009, 86, 401–407. [Google Scholar] [CrossRef]
- Tengku-Rozaina, T.M.; Birch, E.J. Positional distribution of fatty acids on hoki and tuna oil triglycerides by pancreatic lipase and 13C NMR analysis. Eur. J. Lipid Sci. Technol. 2014, 116, 272–281. [Google Scholar] [CrossRef]
- Pratoomyot, J.; Bendiksen, E.A.; Bell, J.G.; Tocher, D.R. Comparison of effects of vegetable oils blended with southern hemisphere fish oil and decontaminated northern hemisphere fish oil on growth performance, composition and gene expression in Atlantic salmon (Salmo salar L.). Aquaculture 2008, 280, 170–178. [Google Scholar] [CrossRef]
- Bell, J.G.; Pratoomyot, J.; Strachan, F.; Henderson, R.J.; Fontanillas, R.; Hebard, A.; Guy, D.R.; Hunter, D.; Tocher, D.R. Growth, flesh adiposity and fatty acid composition of Atlantic salmon (Salmo salar) families with contrasting flesh adiposity: Effects of replacement of dietary fish oil with vegetable oils. Aquaculture 2010, 306, 225–232. [Google Scholar] [CrossRef]
- Pratoomyot, J.; Bendiksen, E.A.; Bell, J.G.; Tocher, D.R. Effects of increasing replacement of dietary fishmeal with plant protein sources on growth performance and body lipid composition of Atlantic salmon (Salmo salar L.). Aquaculture 2010, 305, 124–132. [Google Scholar] [CrossRef]
- Bell, J.G.; McEvoy, J.; Tocher, D.R.; McGhee, F.; Campbell, P.J.; Sargent, J.R. Replacement of fish oil with rapeseed oil in diets of Atlantic salmon (Salmo salar) affects tissue lipid compositions and hepatocyte fatty acid metabolism. J. Nutr. 2001, 131, 1535–1543. [Google Scholar] [PubMed]
- Bell, J.G.; Henderson, R.J.; Tocher, D.R.; McGhee, F.; Dick, J.R.; Porter, A.; Smullen, R.P.; Sargent, J.R. Substituting fish oil with crude palm oil in the diet of Atlantic salmon (Salmo salar) affects muscle fatty acid composition and hepatic fatty acid metabolism. J. Nutr. 2002, 132, 222–230. [Google Scholar] [PubMed]
- Menoyo, D.; Lopez-Bote, C.J.; Bautista, J.M.; Obach, A. Growth, digestibility and fatty acid utilization in large Atlantic salmon (Salmo salar) fed varying levels of n-3 and saturated fatty acids. Aquaculture 2003, 225, 295–293. [Google Scholar] [CrossRef]
- Sargent, J.R.; Tocher, D.R.; Bell, J.G. The lipids. In Fish Nutrition, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: San Diego, CA, USA, 2002; pp. 181–257. [Google Scholar]
- Bell, J.G.; Dick, J.R.; Mc Vicar, A.H.; Sargent, J.R.; Thompson, K.D. Dietary sunflower, linseed and fish oils affect phospholipid fatty acid composition, development of cardiac lesions, phospholipase activity and eicosanoid production in Atlantic salmon. Prost. Leuko. Essen. Fatty Acids 1993, 49, 665–673. [Google Scholar] [CrossRef]
- Alvheim, A.R.; Torstensen, B.E.; Lin, Y.H.; Lillefosse, H.H.; Lock, E.J.; Madsen, L.; Hibbeln, J.R.; Malde, M.K. Dietary linoleic acid elevates endogenous 2-arachidonoylglycerol and anandamide in Atlantic salmon (Salmo salar L.) and mice, and induces weight gain and inflammation in mice. Br. J. Nutr. 2013, 109, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Tocher, D.R.; Bendiksen, E.A.; Campbell, P.J.; Bell, J.G. The role of phospholipids in nutrition and metabolism of teleost fish. Aquaculture 2008, 280, 21–34. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Fish and Shrimp; The National Academies Press: Washington, DC, USA, 2011.
- Torstensen, B.E.; Espe, M.; Sanden, M.; Stubhaug, I.; Waagbo, R.; Hemre, G.I.; Fontanillas, R.; Nordgarden, U.; Hevroy, E.M.; Olsvik, P.; et al. Novel production of Atlantic salmon (Salmo salar) protein based on combined replacement of fish meal and fish oil with plant meal and vegetable oil blends. Aquaculture 2008, 285, 193–200. [Google Scholar] [CrossRef]
- Grahlnielsen, O.; Barnung, T. Variations in the fatty-acid profile of marine animals caused by environmental and developmental-changes. Mar. Environ. Res. 1985, 17, 218–221. [Google Scholar] [CrossRef]
- Segura, J.; Lopez-Bote, C.J. A laboratory efficient method for intramuscular fat analysis. Food Chem. 2014, 145, 821–825. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.J.; Tocher, D.R. Thin-layer chromatography. In Lipid Analysis; Hamilton, R.J., Hamilton, S., Eds.; Oxford University Press: Oxford, UK, 1992; pp. 65–111. [Google Scholar]
- Sayanova, O.; Ruiz-Lopez, N.; Haslam, R.P.; Napier, J.A. The role of D6-desaturase acyl-carrier specificity in the efficient synthesis of long-chain polyunsaturated fatty acids in transgenic plants. Plant Biotechnol. J. 2012, 10, 195–206. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lopez, N.; Haslam, R.P.; Venegas-Caleron, M.; Li, T.B.; Bauer, J.; Napier, J.A.; Sayanova, O. Enhancing the accumulation of omega-3 long chain polyunsaturated fatty acids in transgenic arabidopsis thaliana via iterative metabolic engineering and genetic crossing. Transgenic Res. 2012, 21, 1233–1243. [Google Scholar] [CrossRef] [PubMed]
- Suarez, E.R.; Mugford, P.F.; Rolle, A.J.; Burton, I.W.; Walter, J.A.; Kralovec, J.A. 13C-NMR regioisomeric analysis of EPA and DHA in fish oil derived triacylglycerol concentrates. J. Am. Oil Chem. Soc. 2010, 87, 1425–1433. [Google Scholar] [CrossRef]
- Luddy, F.E.; Magidman, P.; Herb, S.F.; Riemenschneider, R.W.; Barford, R.A. Pancreatic lipase hydrolysis of triglycerides by semimicro technique. J. Am. Oil Chem. Soc. 1964, 41, 693–696. [Google Scholar] [CrossRef]
- Ruiz-Lopez, N.; Haslam, R.P.; Usher, S.; Napier, J.A.; Sayanova, O. An alternative pathway for the effective production of the omega-3 long-chain polyunsaturates EPA and ETA in transgenic oilseeds. Plant Biotechnol. J. 2015. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Force, E.; Ruiz-Lopez, N.; Garces, R. The determination of the asymmetrical stereochemical distribution of fatty acids in triacylglycerols. Anal. Biochem. 2004, 334, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Lopez, N.; Haslam, R.P.; Venegas-Caleron, M.; Larson, T.R.; Graham, I.A.; Napier, J.A.; Sayanova, O. The synthesis and accumulation of stearidonic acid in transgenic plants: A novel source of “heart-healthy” omega-3 fatty acids. Plant Biotechnol. J. 2009, 7, 704–716. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Lopez, N.; Stubhaug, I.; Ipharraguerre, I.; Rimbach, G.; Menoyo, D. Positional Distribution of Fatty Acids in Triacylglycerols and Phospholipids from Fillets of Atlantic Salmon (Salmo Salar) Fed Vegetable and Fish Oil Blends. Mar. Drugs 2015, 13, 4255-4269. https://doi.org/10.3390/md13074255
Ruiz-Lopez N, Stubhaug I, Ipharraguerre I, Rimbach G, Menoyo D. Positional Distribution of Fatty Acids in Triacylglycerols and Phospholipids from Fillets of Atlantic Salmon (Salmo Salar) Fed Vegetable and Fish Oil Blends. Marine Drugs. 2015; 13(7):4255-4269. https://doi.org/10.3390/md13074255
Chicago/Turabian StyleRuiz-Lopez, Noemi, Ingunn Stubhaug, Ignacio Ipharraguerre, Gerald Rimbach, and David Menoyo. 2015. "Positional Distribution of Fatty Acids in Triacylglycerols and Phospholipids from Fillets of Atlantic Salmon (Salmo Salar) Fed Vegetable and Fish Oil Blends" Marine Drugs 13, no. 7: 4255-4269. https://doi.org/10.3390/md13074255
APA StyleRuiz-Lopez, N., Stubhaug, I., Ipharraguerre, I., Rimbach, G., & Menoyo, D. (2015). Positional Distribution of Fatty Acids in Triacylglycerols and Phospholipids from Fillets of Atlantic Salmon (Salmo Salar) Fed Vegetable and Fish Oil Blends. Marine Drugs, 13(7), 4255-4269. https://doi.org/10.3390/md13074255