
Occurrence of Lipophilic Marine Toxins in Shellfish from Galicia (NW of Spain) and Synergies among Them

 ,
,
Abstract
:1. Introduction
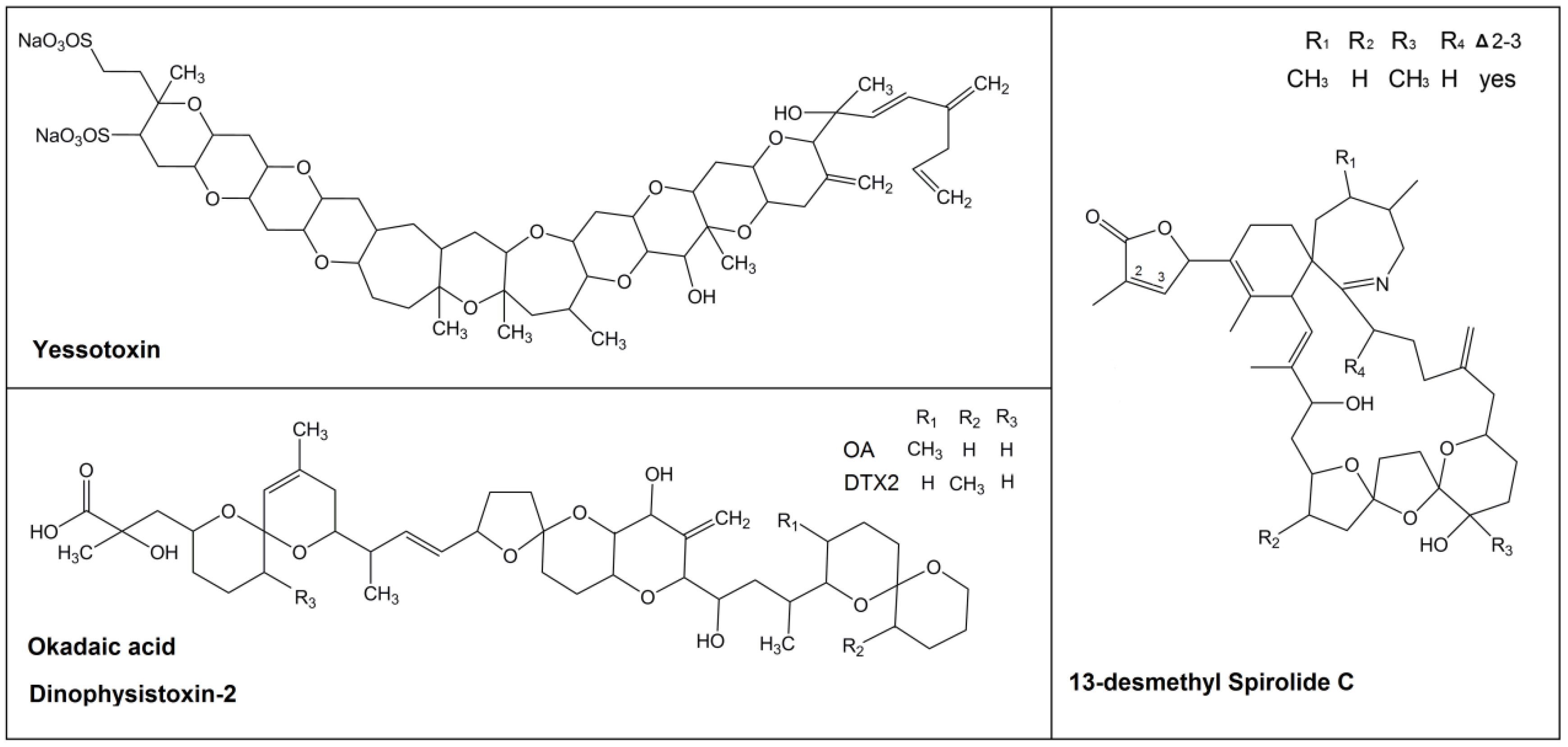
2. Results and Discussion
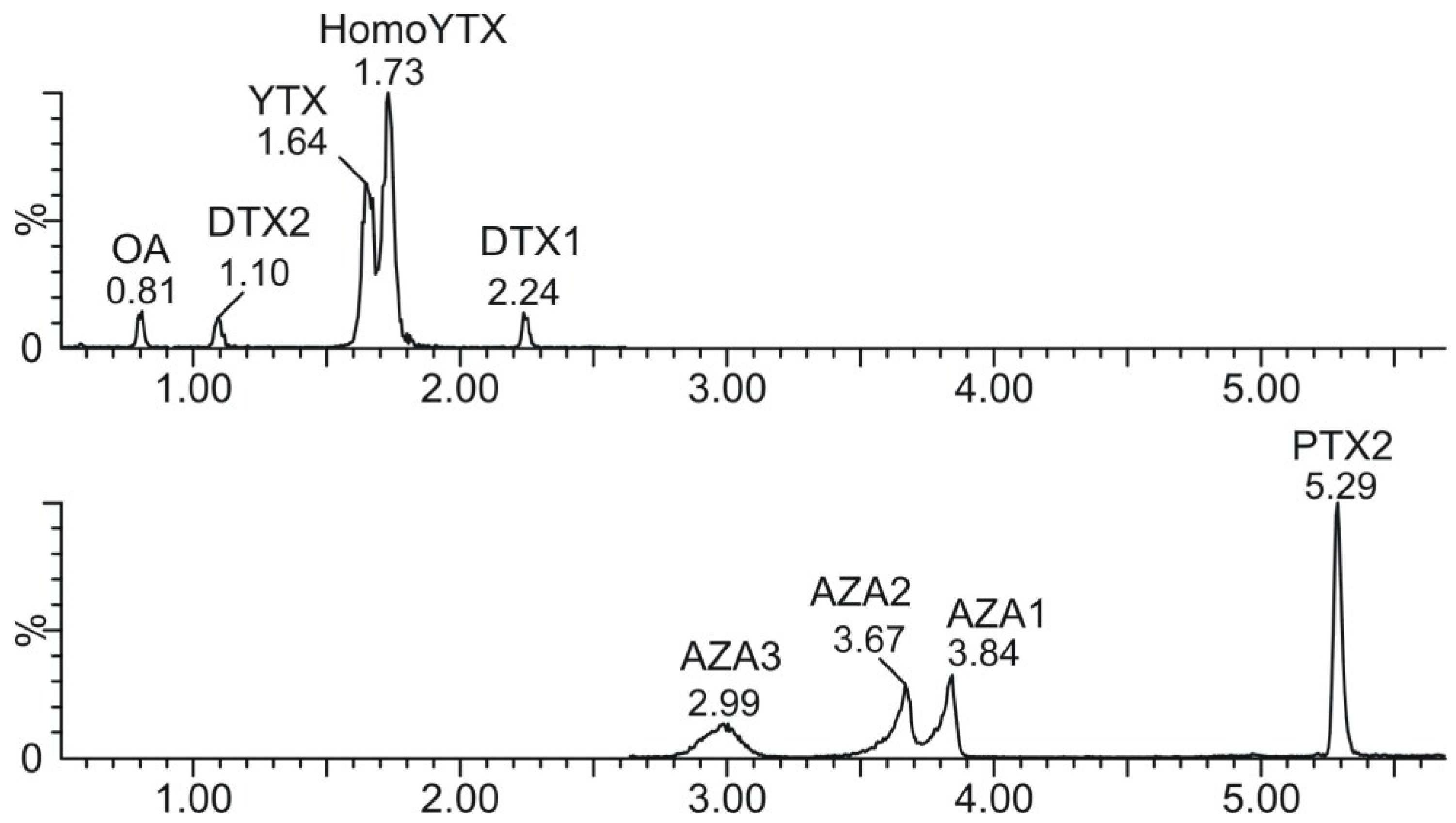
2.1. Lipophilic Toxins Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
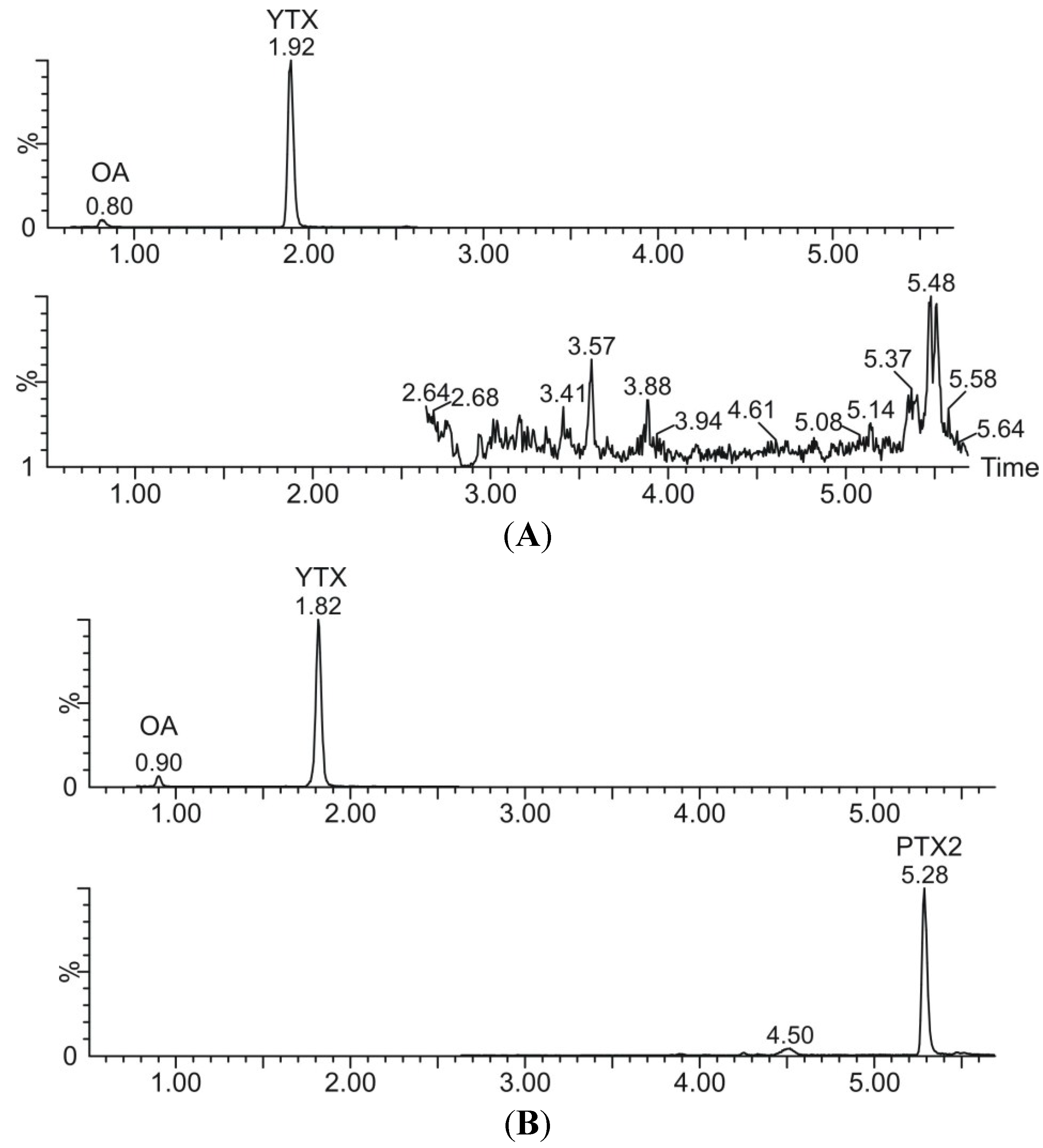{kind=link}
| Matrix | Free OA (μg eq OA/kg) | Total OA (μg eq OA/kg) | Free DTX2 (μg eq OA/kg) | Total DTX2 (μg eq OA/kg) | YTX (mg eq YTX/kg) | 45-OH-YTX (mg eq YTX/kg) | Total YTX (mg eq YTX/kg) | PTX-2 (μg eq AO/kg) |
|---|---|---|---|---|---|---|---|---|
| M. galloprovincialis | 64 | 108 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 85 | 128 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 98 | 129 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 36 | 79 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 46 | 129 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 52 | 85 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 73 | 118 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 128 | 142 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 47 | 135 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 46 | 128 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 56 | 147 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 86 | 162 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 55 | 92 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 26 | 78 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 41 | 180 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 76 | 201 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 261 | 361 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 46 | 103 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 87 | 193 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 76 | 162 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 169 | 376 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 69 | 107 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 42 | 83 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | <LOQ | 44 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 96 | 177 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 41 | 59 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 42 | 67 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 29 | 64 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 52 | 78 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 43 | 104 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 157 | 259 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 73 | 135 | <LOQ | <LOQ | 0.49 | <LOQ | 0.49 | <LOQ |
| M. galloprovincialis | 80 | 183 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | <LOQ | 64 | <LOQ | <LOQ | 0.25 | <LOQ | 0.25 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.59 | 0.11 | 0.7 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.3 | 0.08 | 0.38 | <LOQ |
| M. galloprovincialis | <LOQ | 43 | <LOQ | <LOQ | 0.19 | <LOQ | 0.19 | <LOQ |
| M. galloprovincialis | <LOQ | 56 | <LOQ | <LOQ | 0.21 | <LOQ | 0.21 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.11 | <LOQ | 0.11 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.12 | <LOQ | 0.12 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.27 | 0.08 | 0.35 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.09 | <LOQ | 0.09 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.15 | <LOQ | 0.15 | <LOQ |
| M. galloprovincialis | <LOQ | 42 | <LOQ | <LOQ | 0.24 | <LOQ | 0.24 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.08 | <LOQ | 0.08 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.14 | <LOQ | 0.14 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.16 | <LOQ | 0.16 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.1 | <LOQ | 0.1 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.09 | <LOQ | 0.09 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.09 | <LOQ | 0.09 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.15 | <LOQ | 0.15 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.2 | <LOQ | 0.2 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.18 | <LOQ | 0.18 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.13 | <LOQ | 0.13 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.23 | <LOQ | 0.23 | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | 0.17 | <LOQ | 0.17 | <LOQ |
| M. galloprovincialis | 63 | 105 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| M. galloprovincialis | 697 | 890 | 87 | 92 | <LOQ | <LOQ | 0.45 | <LOQ |
| M. galloprovincialis | 84 | 129 | <LOQ | <LOQ | 0.40 | 0.07 | <LOQ | 46 |
| Venerupis spp | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Chlamys varia | <LOQ | 391 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Chlamys varia | <LOQ | 104 | <LOQ | 83 | <LOQ | <LOQ | <LOQ | <LOQ |
| Chlamys varia | <LOQ | 100 | <LOQ | 185 | <LOQ | <LOQ | <LOQ | <LOQ |
| Chlamys opercularis | <LOQ | 101 | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
| Pecten maximus | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ | <LOQ |
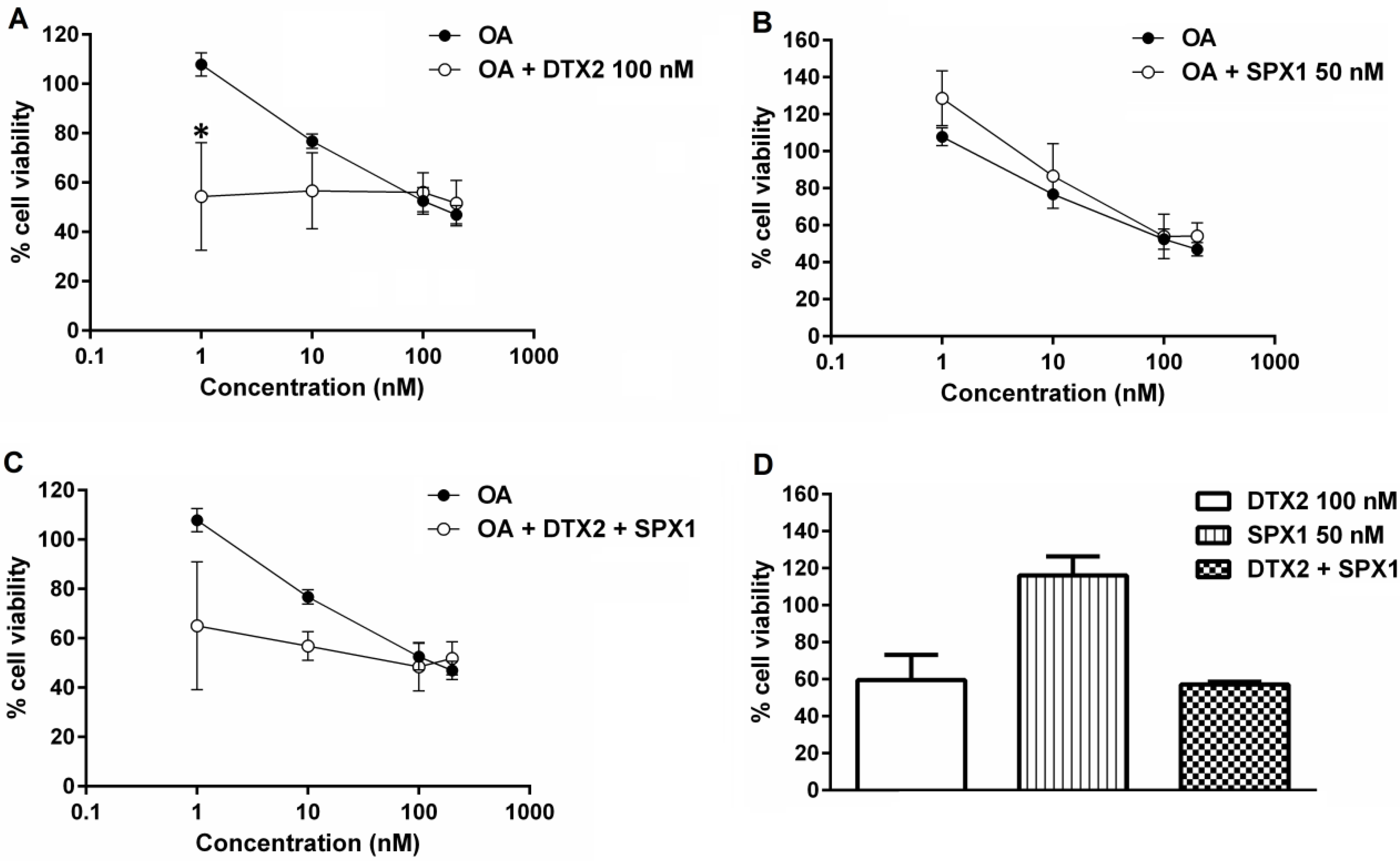
2.2. Effects of Toxins on Cell Viability

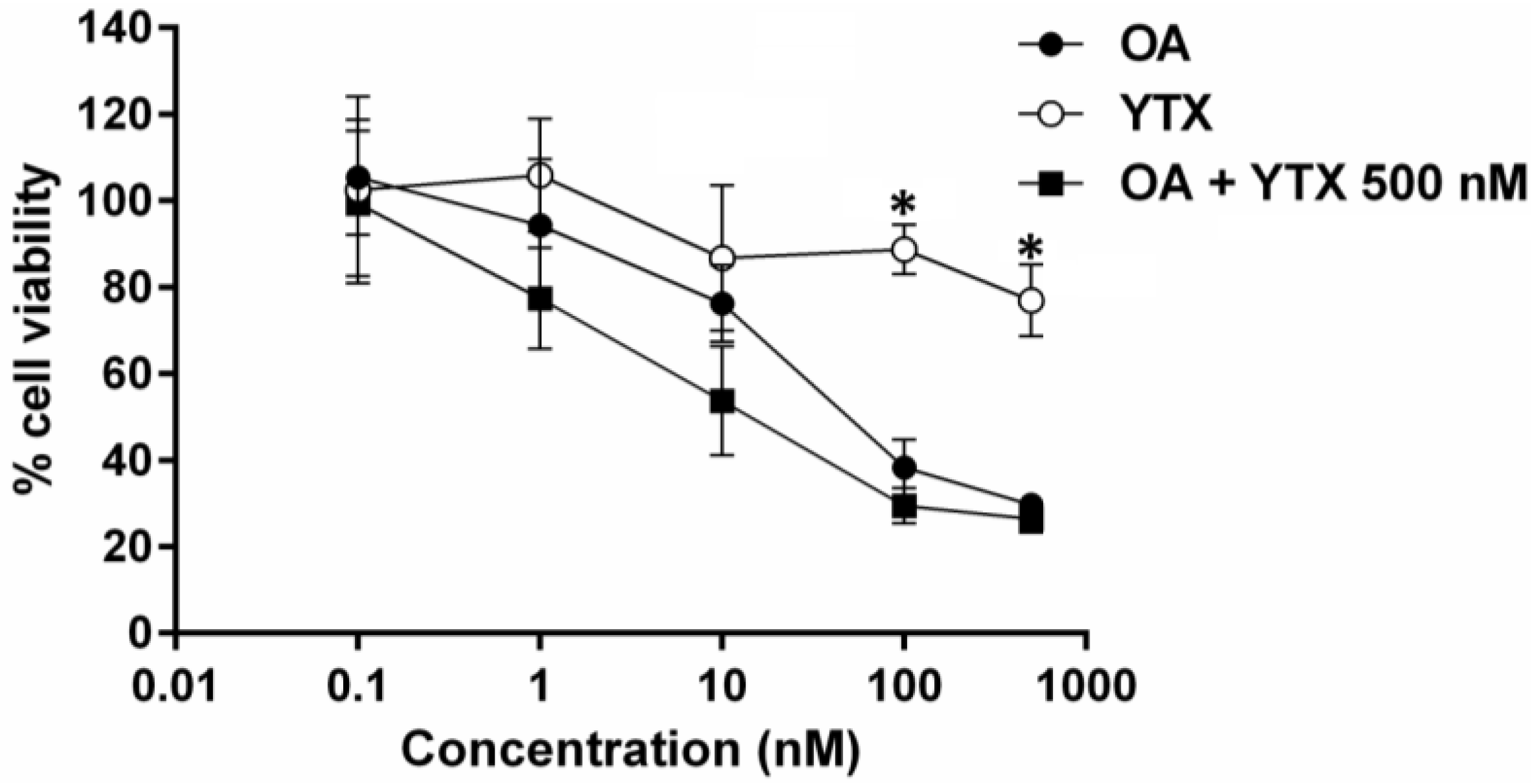
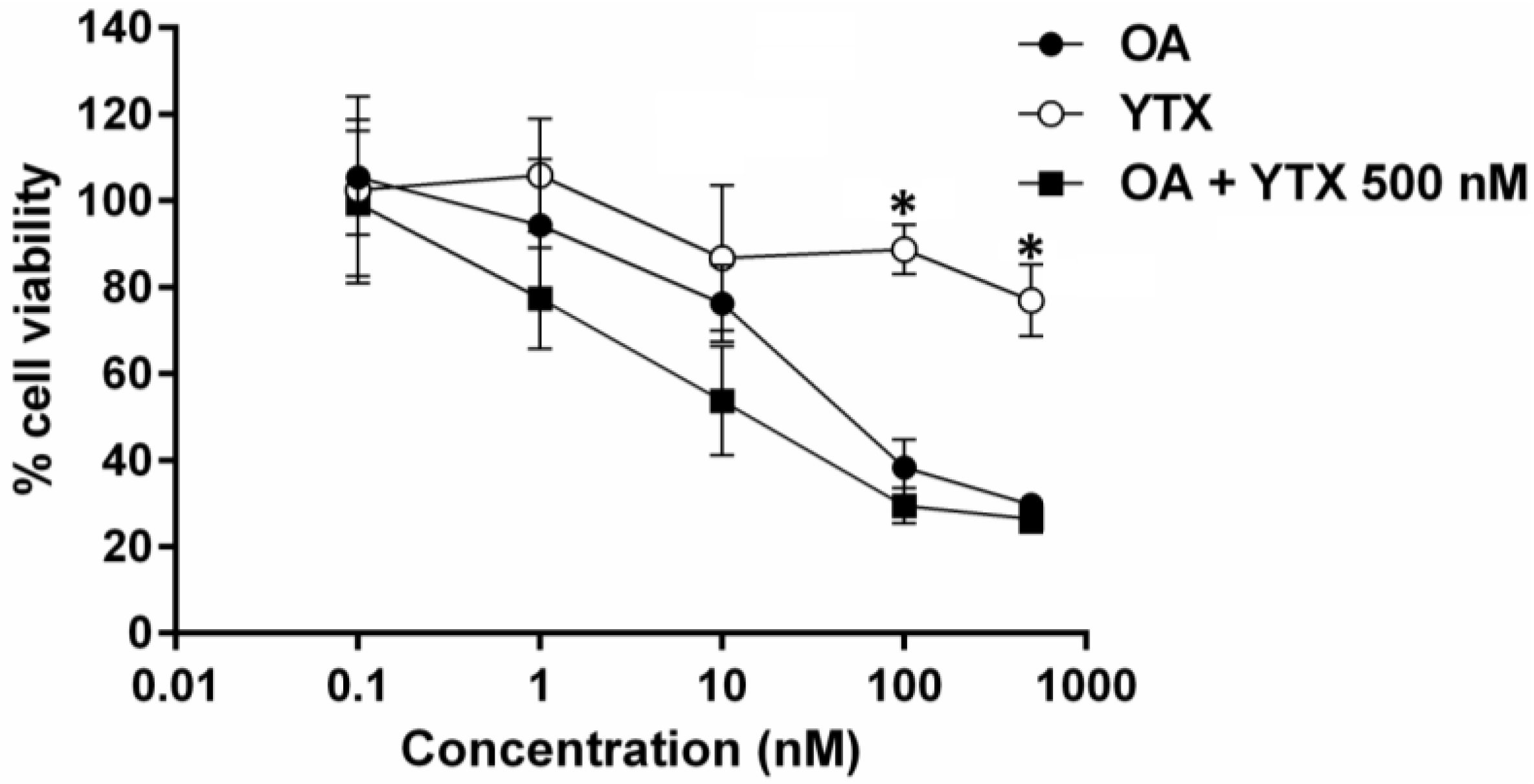
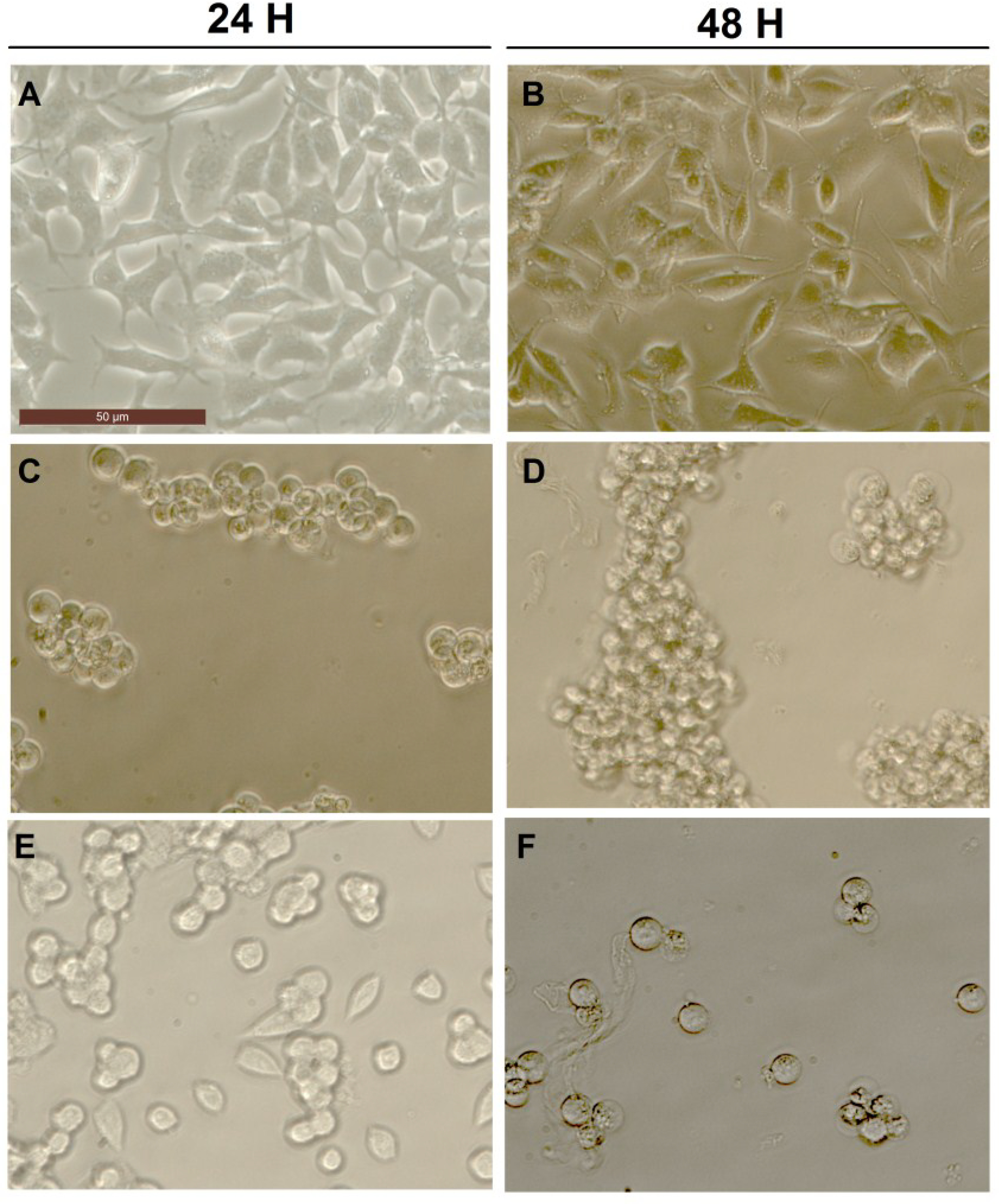
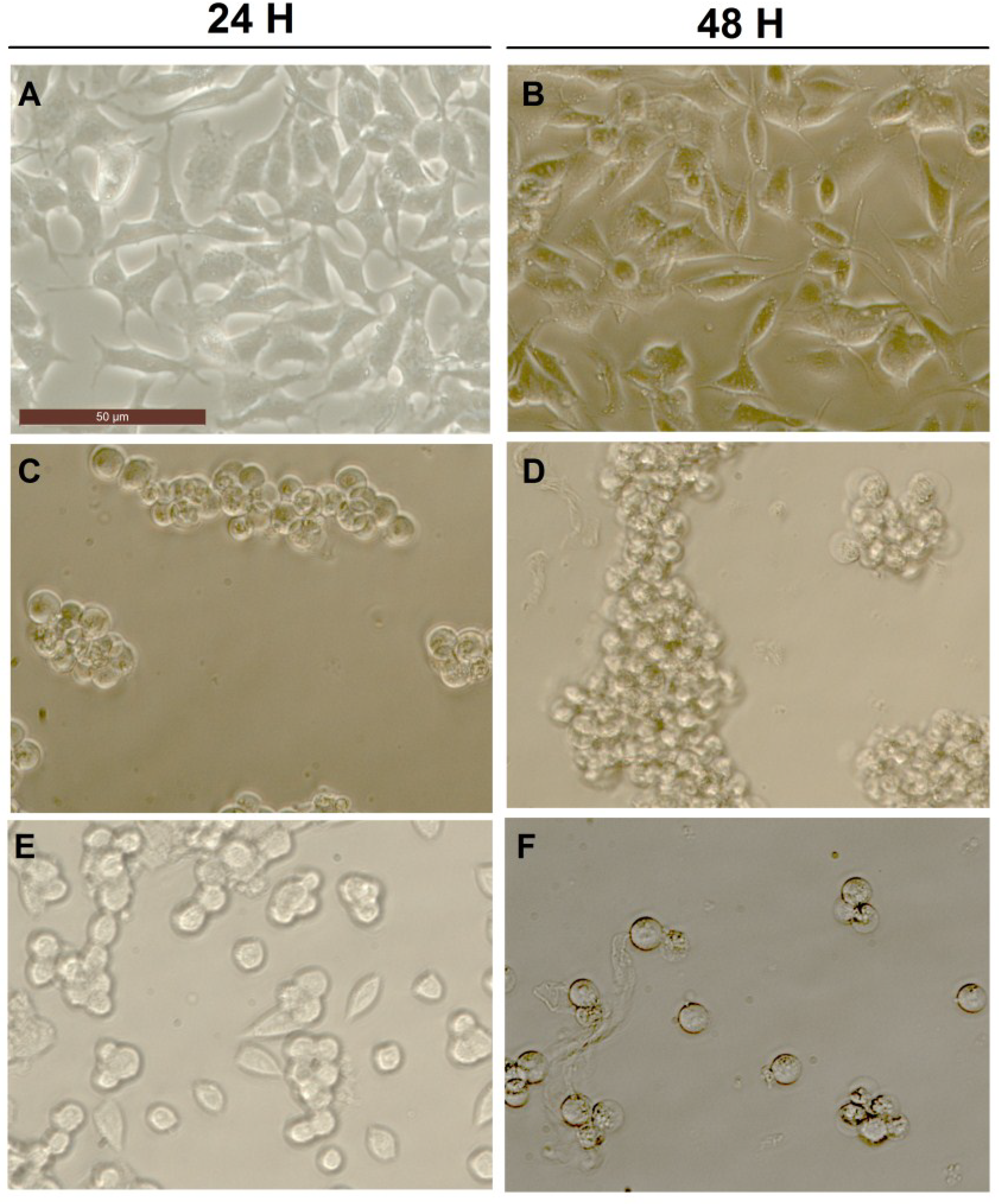
2.3. Effect of Okadaic Acid and Yessotoxin on Cell Morphology
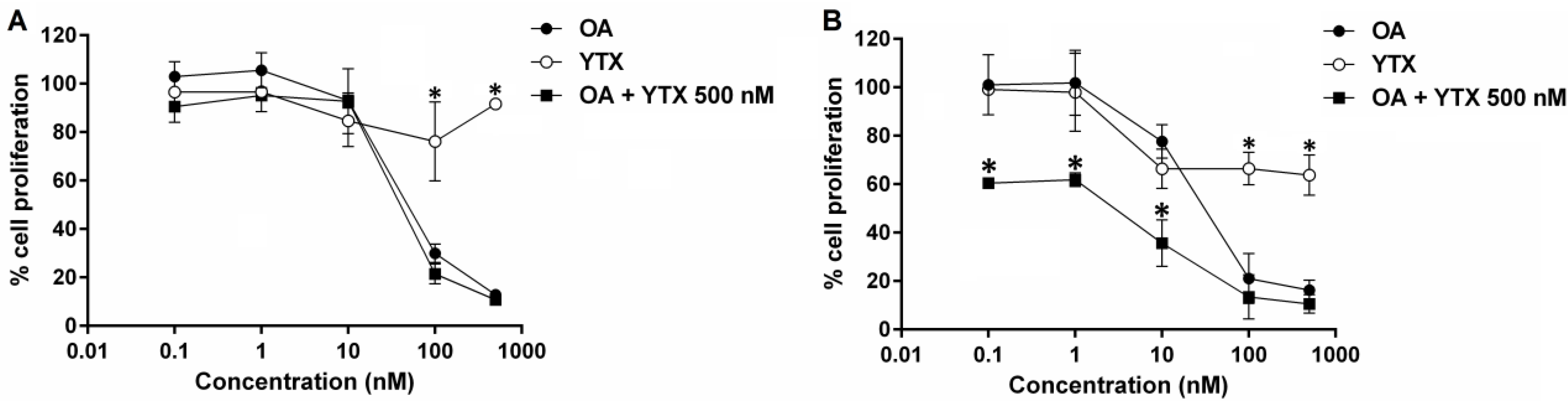
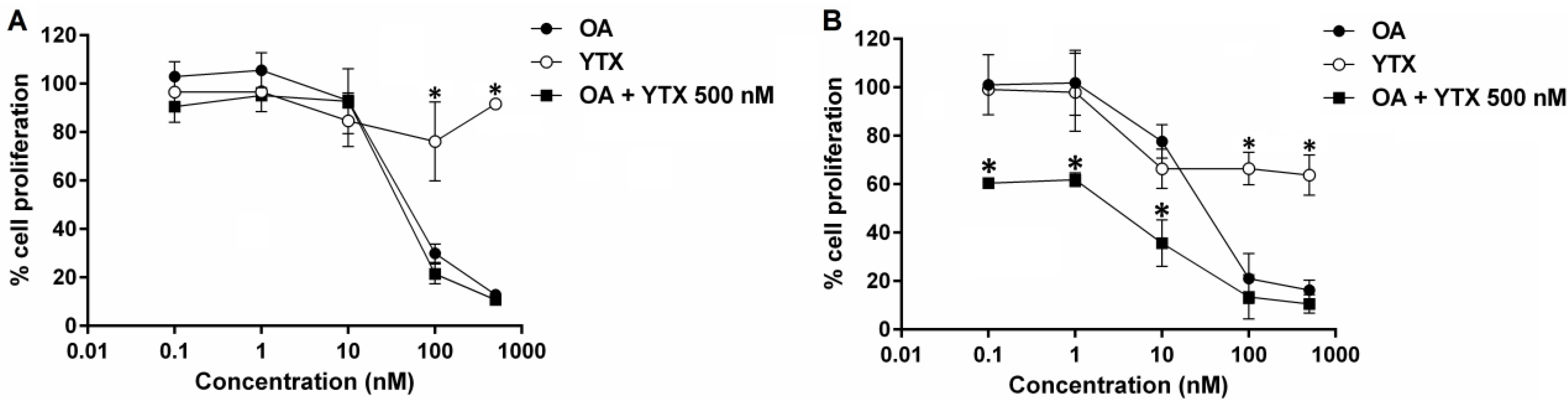
2.4. Effects of Okadaic Acid and Yessotoxin on Cell Proliferation
3. Experimental Section
3.1. Reagents and Toxin Standards
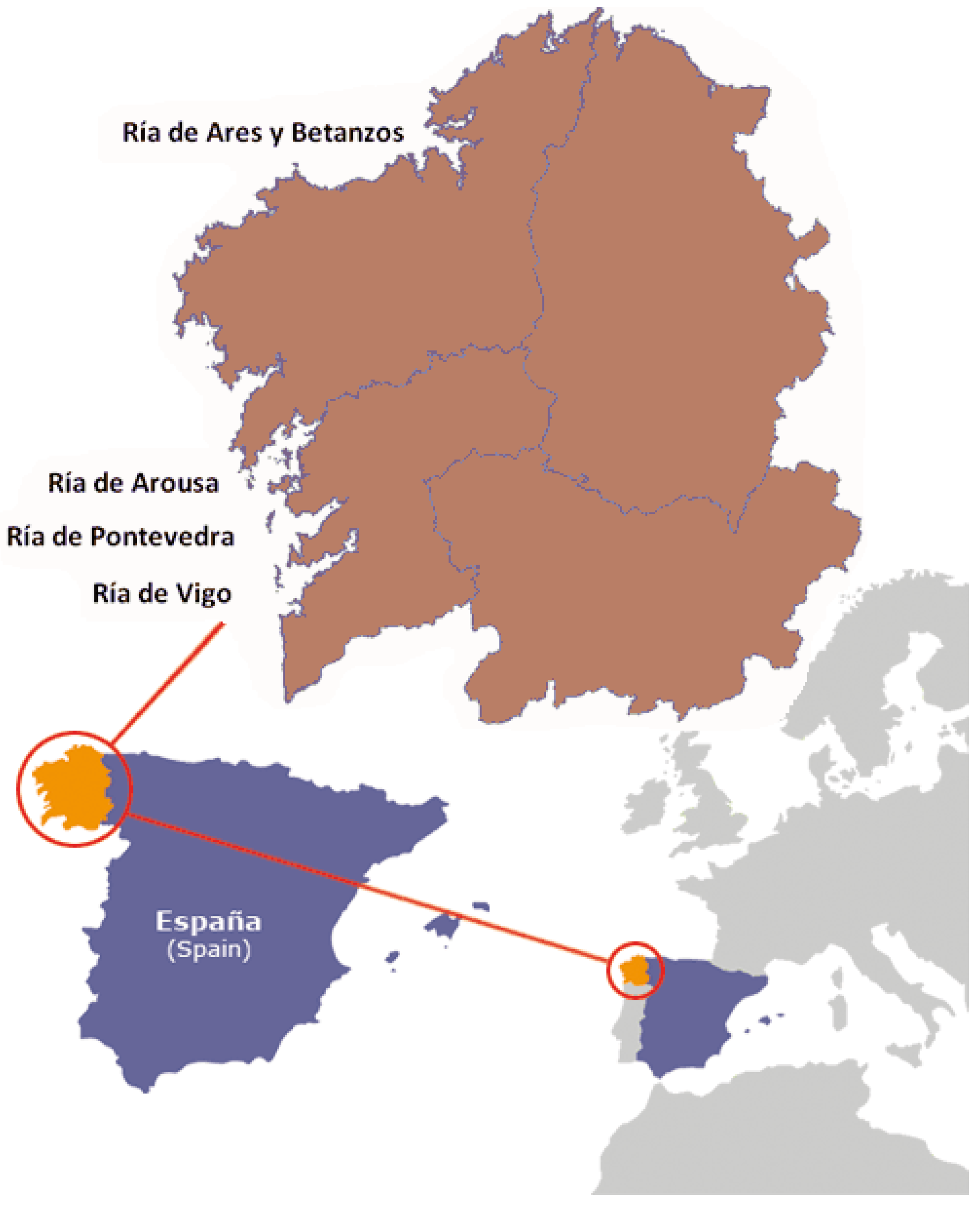
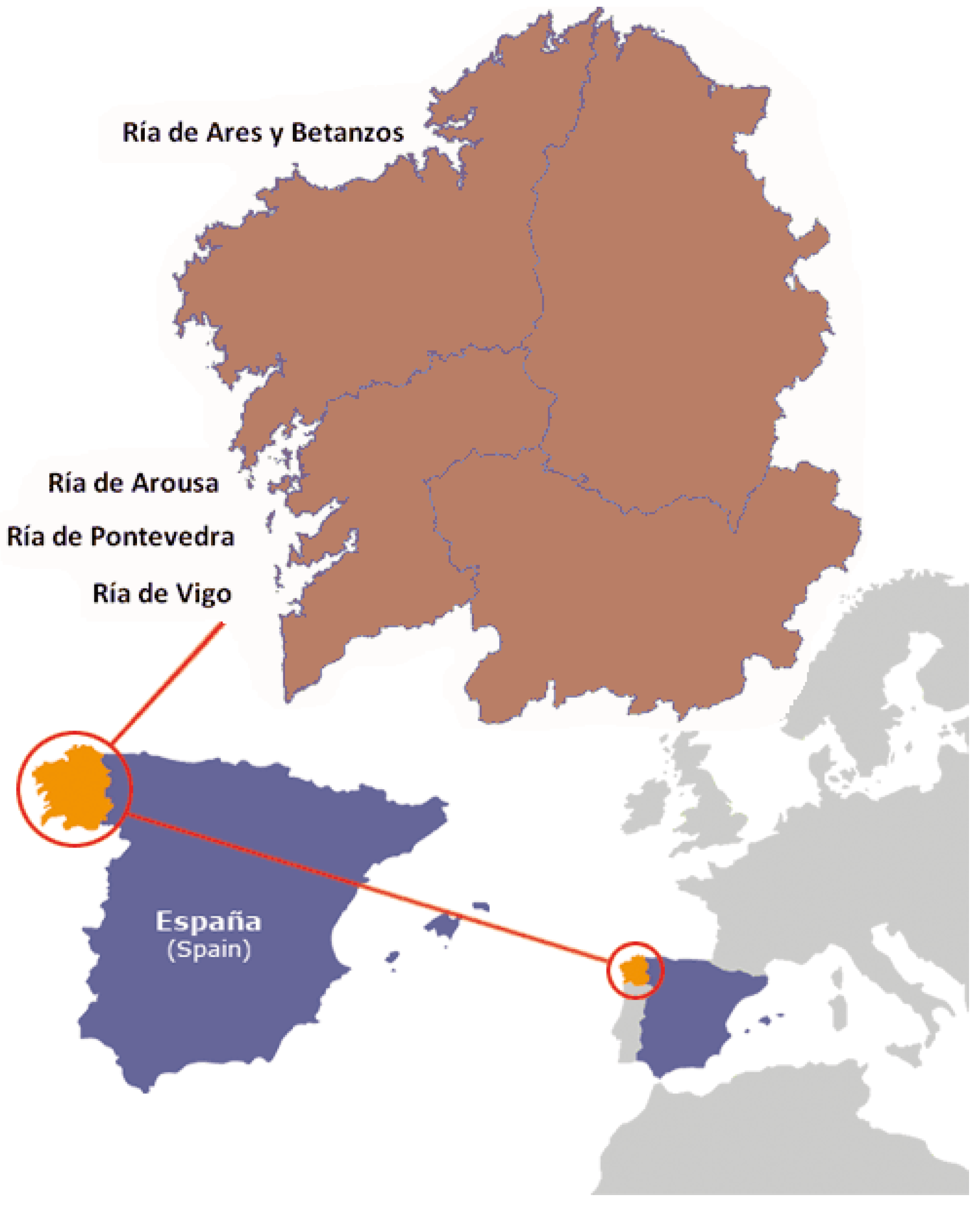
3.2. Sampling
3.3. Sample Extraction
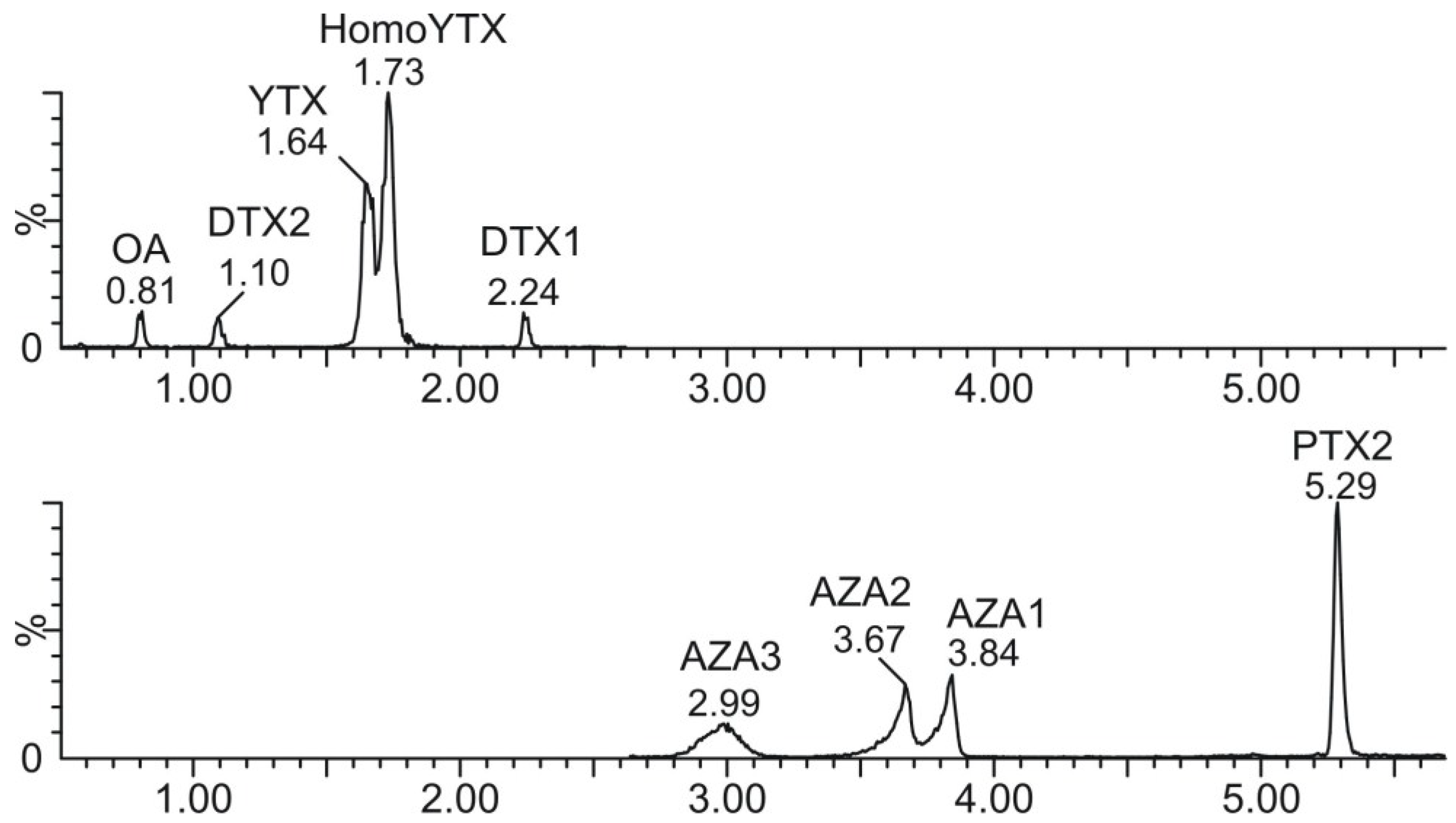
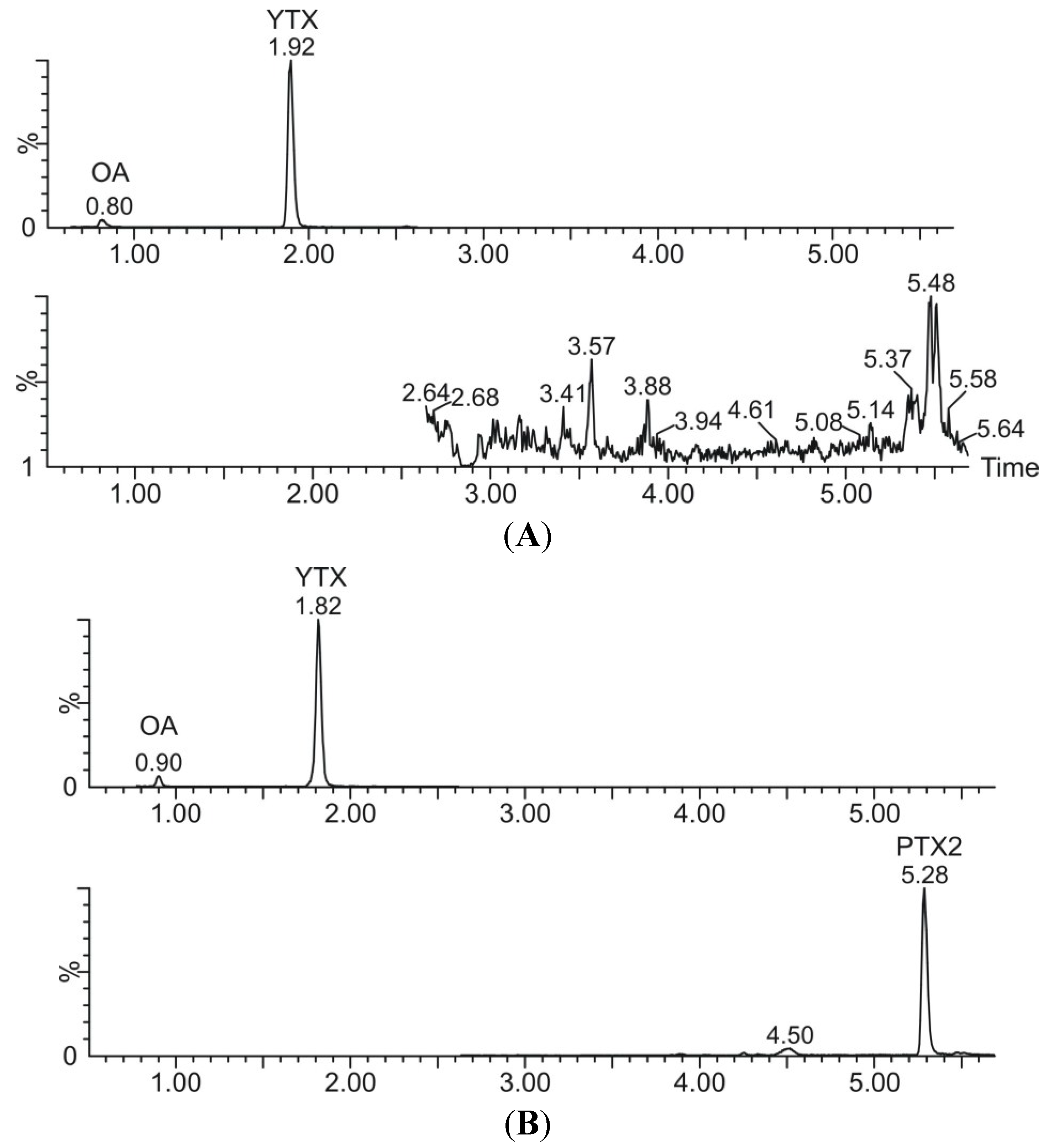
3.4. Liquid Chromatography–Mass Spectrometry (LC–MS/MS)
| Column | Waters Acquity UPLC BEH C18 1.7 µm 2.1 × 100 mm | |
| Flow | 0.40 mL/min | |
| Injection volume | 2 μL | |
| Column temperature | 45 °C | |
| Maximum pressure | 15,000 psi | |
| Time (min) | % Mobile phase A | % Mobile phase B |
| Ammonia 6.7 mM in water | Ammonia 6.7 mM in acetonitrile | |
| initial | 70 | 30 |
| 0.50 | 70 | 30 |
| 6.00 | 10 | 90 |
| 6.50 | 0 | 100 |
| 9.50 | 0 | 100 |
| 9.60 | 70 | 30 |
| 10.00 | 70 | 30 |
| Compound | Parent Ion (Da) | Fragment Ion (Da) | ESI | Cone Voltage (V) | Collision Voltage (eV) | Dwell (s) |
|---|---|---|---|---|---|---|
| AZA3 * | 828.50 | 362.20 | + | 47 | 55 | 0.007 |
| AZA3 | 828.50 | 640.40 | + | 47 | 55 | 0.007 |
| AZA1 | 842.50 | 362.00 | + | 50 | 50 | 0.038 |
| AZA1 * | 842.50 | 654.50 | + | 50 | 55 | 0.036 |
| AZA2 * | 856.50 | 654.50 | + | 27 | 45 | 0.038 |
| AZA2 | 856.50 | 672.50 | + | 27 | 45 | 0.038 |
| PTX2 * | 876.50 | 823.50 | + | 36 | 25 | 0.034 |
| PTX2 | 876.50 | 841.50 | + | 36 | 20 | 0.034 |
| PTX1 | 892.50 | 213.50 | + | 36 | 20 | 0.034 |
| PTX1 * | 892.50 | 821.50 | + | 36 | 25 | 0.034 |
| PTX1 | 892.50 | 839.50 | + | 36 | 25 | 0.034 |
| YTX | 570.40 | 396.30 | − | 45 | 35 | 0.007 |
| YTX * | 570.40 | 467.40 | − | 45 | 30 | 0.007 |
| h-YTX | 577.50 | 403.40 | − | 48 | 30 | 0.007 |
| h-YTX * | 577.50 | 474.40 | − | 48 | 30 | 0.007 |
| 45-OH-YTX | 578.40 | 396.40 | − | 75 | 35 | 0.007 |
| 45-OH-YTX * | 578.40 | 467.40 | − | 75 | 30 | 0.007 |
| 45-OH-homo-YTX | 585.50 | 403.40 | − | 75 | 30 | 0.007 |
| 45-OH-homo-YTX * | 585.50 | 474.40 | − | 75 | 30 | 0.007 |
| AO-DTX2 | 803.50 | 113.00 | − | 86 | 56 | 0.007 |
| AO-DTX2 * | 803.50 | 255.20 | − | 86 | 46 | 0.007 |
| DTX1 * | 817.50 | 255.20 | − | 86 | 48 | 0.007 |
| DTX1 | 817.50 | 113.1 | − | 86 | 43 | 0.007 |
3.5. Cell Culture
3.6. Cell Viability Assay
3.7. Light Microscopy
3.8. Cell Proliferation Assay
3.9. Data Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAO. Fisheries and Aquaculture Department. Statistical Collections. Available online: http://www.fao.org/fishery/statistics/en (accessed on 27 December 2014).
- Blanco, J. Evaluación del impacto de los métodos y niveles utilizados para el control de toxinas en el mejillón. Available online: http://www.magrama.gob.es/app/jacumar/planes_nacionales/documentos/97_if_culmites_anexo_galicia.pdf (accessed on 27 December 2014).
- Ferreira, M.; Lago, J.; Vieites, J.M.; Cabado, A.G. World production of bivalve mollusks and socioeconomic facts related to the impact of marine biotoxins. In Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection, 3rd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 291–311. [Google Scholar]
- Gestal-Otero, J.J. Epidemiology of marine toxins. In Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection, 3rd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 123–195. [Google Scholar]
- Álvarez-Salgado, X.A.; Labarta, U.; Fernández-Reiriz, M.J.; Figueiras, F.G.; Rosón, G.; Piedracoba, S.; Filgueira, R.; Cabanas, J.M. Renewal time and the impact of harmful algal blooms on the extensive mussel raft culture of the Iberian coastal upwelling system (SW Europe). Harmful Algae 2008, 7, 849–855. [Google Scholar] [CrossRef]
- Dittami, S.; Pazos, Y.; Laspra, M.; Medlin, L. Microarray testing for the presence of toxic algae monitoring programme in Galicia (NW Spain). Environ. Sci. Pollut. Res. 2013, 20, 6778–6793. [Google Scholar] [CrossRef]
- Galicia, X.D. Anuario de pesca 2009. Available online: http://www.pescadegalicia.com/Publicaciones/AnuarioPesca2009/index.htm (accessed on 19 February 2014).
- Moroño, A.; Arévalo, F.; Fernández, M.L.; Maneiro, J.; Pazos, Y.; Salgado, C.; Blanco, J. Accumulation and transformation of DSP toxins in mussels Mytilus galloprovincialis during a toxic episode caused by Dinophysis acuminata. Aquat. Toxicol. 2003, 26, 269–280. [Google Scholar] [CrossRef]
- Regueiro, J.; Rossignoli, A.E.; Álvarez, G.; Blanco, J. Automated on-line solid-phase extraction coupled to liquid chromatography tandem mass spectrometry for determination of lipophilic marine toxins in shellfish. Food Chem. 2011, 129, 533–540. [Google Scholar] [CrossRef]
- Blanco, J.; Mariño, C.; Marín, H.; Acosta, C.P. Anatomical distribution of diarrhetic shellfish poisoning (DSP) toxins in the mussel Mytilus galloprovincialis. Toxicon 2007, 50, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Villar-González, A.; Rodríguez-Velasco, M.L.; Ben-Gigirey, B.; Botana, L.M. Lipophilic toxin profile in Galicia (Spain): 2005 toxic episode. Toxicon 2007, 49, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, G.; Paz, B.; Franco, J.M.; Suzuki, T.; Reguera, B. First detection of pectenotoxin-11 and confirmation of OA-D8 diol-ester in Dinophysis acuta from European waters by LC-MS/MS. Toxicon 2008, 52, 889–896. [Google Scholar] [CrossRef] [PubMed]
- Arévalo, F.; Pazos, Y.; Correa, J.; Salgado, C.; Moroño, A.; Paz, B.; Franco, J.M. First report of yessotoxins in mussels of Galician Rias during a bloom of Lingulodinium polyedra Stein (Dodge). In Fifth International Conference on Molluscan Shellfish Safety; Henshilwood, K., Deegan, B., McMahon, T., Cusak, C., Keaveney, S., Silke, J.O., Cinneide, M., Lyons, D., Hess, P., Eds.; Marine Institute: Galway, Ireland, 2006; pp. 184–189. [Google Scholar]
- Magdalena, A.B.; Lehane, M.; Krys, S.; Fernández, M.L.; Furey, A.; James, K.J. The first identification of azaspiracids in shellfish from France and Spain. Toxicon 2003, 42, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Villar González, A.; Rodríguez-Velasco, M.L.; Ben-Gigirey, B.; Botana, L.M. First evidence of spirolides in Spanish shellfish. Toxicon 2006, 48, 1068–1074. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, H. Joint FAO/WHO/IOC activities to provide scientific advice on marine biotoxins (research report). Mar. Pollut. Bull. 2006, 52, 1735–1745. [Google Scholar] [CrossRef] [PubMed]
- FAO. Food and nutrition paper 80: Marine biotoxins. Available online: http://www.fao.org/docrep/007/y5486e/y5486e00.HTM (accessed on 27 December 2014).
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.K.; Clardy, J. Diarrethic shellfish toxins. Tetrahedron Lett. 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion of the panel on contaminants in the food chain. Marine biotoxins in shellfish-okadaic acid and analogues. EFSA J. 2008, 589, 1–62. [Google Scholar]
- Tachibana, K.; Scheuer, P.J.; Tsukitani, Y.; Kikuchi, H.; van Engen, D.; Clardy, J.; Gopichand, Y.; Schmitz, F.J. Okadaic acid, a cytotoxic polyether from two marine sponges of the genus Halichondria. J. Am. Chem. Soc. 1981, 103, 2469–2471. [Google Scholar] [CrossRef]
- Dickey, R.W.; Bobzin, S.C.; Faulkner, D.J.; Bencsath, F.A.; Andrzejewski, D. Identification of okadaic acid from a caribbean dinoflagellate, Prorocentrum concavum. Toxicon 1990, 28, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Bravo, I.; Fernández, M.L.; Ramilo, I.; Martínez, A. Toxin composition of the toxic dinoflagellate Prorocentrum lima isolated from different locations along the Galician coast (NW Spain). Toxicon 2001, 39, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.L.; Reguera, B.; González-Gil, S.; Míguez, A. Pectenotoxin-2 in single-cell isolates of Dinophysis caudata and Dinophysis acuta from the Galician Rías (NW Spain). Toxicon 2006, 48, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Escalera, L.; Reguera, B.; Moita, T.; Pazos, Y.; Cerejo, M.; Cabanas, J.M.; Ruiz-Villarreal, M. Bloom dynamics of Dinophysis acuta in an upwelling system: In situ growth versus transport. Harmful Algae 2010, 9, 312–322. [Google Scholar] [CrossRef]
- Lee, J.S.; Igarashi, T.; Fraga, S.; Dahl, E.; Hovgaard, P.; Yasumoto, T. Determination of diarrhetic shellfish toxins in various dinoflagellate species. J. Appl. Phycol. 1989, 1, 147–152. [Google Scholar] [CrossRef]
- Quilliam, M.A.; Gilgan, M.W.; Pleasance, S.; de Freitas, A.S.W.; Douglas, D.; Fritz, L. Confirmation of an incident of diarrhetic shellfish poisoning in Eastern Canada. In Toxic Phytoplankton Blooms in the Sea; Smayda, T., Shimuzu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 547–552. [Google Scholar]
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in Japan and chemical properties of the toxin. In Toxic Dinoflagellates Blooms; Taylor, D., Seliger, H., Eds.; Elsevier: Amsterdam, The Netherlands, 1979; pp. 495–502. [Google Scholar]
- Hu, T.; Doyle, J.; Jackson, D.; Marr, J.; Nixon, E.; Pleasance, S.; Quilliam, M.A.; Walter, J.A.; Wright, J.L.C. Isolation of a new diarrhetic shellfish poison from Irish mussels. J. Chem. Soc. Chem. Commun. 1992, 39–41. [Google Scholar] [CrossRef]
- Blanco, J.; Fernández, M.; Marino, J.; Reguera, B.; Míguez, A.; Maneiro, J. From Dinophysis spp. toxicity to DSP outbrakes: A preliminary model of toxin accumulation in mussels. In Harmful Marine Algal Blooms; Lassus, P., Arzul, G., Erard, E., Gentien, P., Marcaillou, C., Eds.; Lavoisier Science Publishers: Paris, France, 1995; pp. 777–782. [Google Scholar]
- Vale, P.; Sampayo, M.A. Dinophysistoxin-2: A rare diarrhoeic toxin associated with Dinophysis acuta. Toxicon 2000, 38, 1599–1606. [Google Scholar] [CrossRef] [PubMed]
- Vale, P.; de M. Sampayo, M.A. Esters of okadaic acid and dinophysistoxin-2 in Portuguese bivalves related to human poisonings. Toxicon 1999, 37, 1109–1121. [Google Scholar] [CrossRef] [PubMed]
- Vale, P.; de M. Sampayo, M.A. Esterification of DSP toxins by portuguese bivalves from the northwest coast determined by LC–MS—A widespread phenomenon. Toxicon 2002, 40, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Smienk, H.; Domínguez, E.; Rodríguez-Velasco, M.L.; Clarke, D.; Kapp, K.; Katikou, P.; Cabado, A.G.; Otero, A.; Vieites, J.M.; Razquin, P.; et al. Quantitative determination of the okadaic acid toxins group by a colorimetric phosphatase inhibition assay: Interlaboratory study. J. AOAC Int. 2013, 96, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Kumagai, M.; Lee, J.S.; Yasumoto, T. Isolation and structure of yessotoxin, a novel polyether compound implicated in diarrhetic shellfish poisoning. Tetrahedron Lett. 1987, 28, 5869–5872. [Google Scholar] [CrossRef]
- Satake, M.; MacKenzie, L.; Yasumoto, T. Identification of Protoceratium reticulatum as the biogenetic origin of yessotoxin. Nat. Toxins 1997, 5, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Draisci, R.; Ferretti, E.; Palleschi, L.; Marchiafava, C.; Poletti, R.; Milandri, A.; Ceredi, A.; Pompei, M. High levels of yessotoxin in mussels and presence of yessotoxin and homoyessotoxin in dinoflagellates of the Adriatic Sea. Toxicon 1999, 37, 1187–1193. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.L.; McNabb, P.; De Salas, M.; Briggs, L.; Beuzenberg, V.; Gladstone, M. Yessotoxin production by Gonyaulax spinifera. Harmful Algae 2006, 5, 148–155. [Google Scholar] [CrossRef]
- Paz, B.; Riobó, P.; Luisa Fernández, M.; Fraga, S.; Franco, J.M. Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture. Toxicon 2004, 44, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Paz, B.; Blanco, J.; Franco, J.M. Yessotoxins production during the culture of Protoceratium reticulatum strains isolated from Galician Rias Baixas (NW Spain). Harmful Algae 2013, 21–22, 13–19. [Google Scholar]
- Domínguez, H.J.; Paz, B.; Daranas, A.H.; Norte, M.; Franco, J.M.; Fernández, J.J. Dinoflagellate polyether within the yessotoxin, pectenotoxin and okadaic acid toxin groups: Characterization, analysis and human health implications. Toxicon 2010, 56, 191–217. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Regulation (EC) No 853/2004 of the European Parliament and of the Council of April 29, 2004 laying down specific hygiene rules for food of animal origin. Off. J. Eur. 2004, L139, 55–205. [Google Scholar]
- European Commission. Comission regulation (EU) No 786/2013 of 16 August 2013 amending Annex III to regulation (EC) No 854/2204 of the European Parliament and of the Council as regards the permitted limits of yessotoxins in live bivalve molluscs. Off. J. Eur. Communities. 2013, L220, 14. [Google Scholar]
- Ogino, H.; Kumagai, M.; Yasumoto, T. Toxicologic evaluation of yessotoxin. Nat. Toxins 1997, 5, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Biré, R.; Krys, S.; Frémy, J.M.; Dragacci, S.; Stirling, D.; Kharrat, R. First evidence on occurrence of gymnodimine in clams from Tunisia. J. Nat. Toxins 2002, 11, 269–275. [Google Scholar] [PubMed]
- Molgó, J.; Girard, E.; Benoit, E. Cyclic imines: An insight into this emerging group of bioactive marine toxins. In Phycotoxins: Chemistry and Biochemistry; Botana, L.M., Ed.; Blackwell Publishing: Ames, IA, USA, 2007; pp. 319–335. [Google Scholar]
- Takada, N.; Umemura, N.; Suenaga, K.; Chou, T.; Nagatsu, A.; Haino, T.; Yamada, K.; Uemura, D. Pinnatoxins B and C, the most toxic components in the pinnatoxin series from the Okinawan bivalve Pinna muricata. Tetrahedron Lett. 2001, 42, 3491–3494. [Google Scholar] [CrossRef]
- Takada, N.; Umemura, N.; Suenaga, K.; Uemura, D. Structural determination of pteriatoxins A, B and C, extremely potent toxins from the bivalve Pteria penguin. Tetrahedron Lett. 2001, 42, 3495–3497. [Google Scholar] [CrossRef]
- Torigoe, K.; Murata, M.; Yasumoto, T. Prorocentrolide, a toxic nitrogenous macrocycle from a marine dinoflagellate, Prorocentrum lima. J. Am. Chem. Soc. 1988, 110, 7876–7877. [Google Scholar] [CrossRef]
- Uemura, D.; Chou, T.; Haino, T.; Nagatsu, A.; Fukuzawa, S.; Zheng, S.-Z.; Chen, H.-S. Pinnatoxin A: A toxic amphoteric macrocycle from the Okinawan bivalve Pinna muricata. J. Am. Chem. Soc. 1995, 117, 1155–1156. [Google Scholar] [CrossRef]
- Hu, T.; Curtis, J.M.; Oshima, Y.; Quilliam, M.A.; Walter, J.A.; Watson-Wright, W.M.; Wright, J.L.C. Spirolides B and D, two novel macrocycles isolated from the digestive glands of shellfish. J. Chem. Soc. Chem. Commun. 1995, 20, 2159–2161. [Google Scholar] [CrossRef]
- Cembella, A.D.; Lewis, N.I.; Quilliam, M.A. Spirolide composition of micro-extracted pooled cells isolated from natural plankton assemblages and from cultures of the dinoflagellate Alexandrium ostenfeldii. Nat. Toxins 1999, 7, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Cembella, A.D.; Lewis, N.I.; Quilliam, M.A. The marine dinoflagellate Alexandrium ostenfeldii (Dinophyceae) as the causative organism of spirolide shellfish toxins. Phycologia 2000, 39, 67–74. [Google Scholar] [CrossRef]
- Touzet, N.; Franco, J.M.; Raine, R. Morphogenetic diversity and biotoxin composition of Alexandrium (Dinophyceae) in Irish coastal waters. Appl. Environ. Microbiol. 2008, 7, 782–797. [Google Scholar]
- EFSA. Scientific opinion on marine biotoxins in shellfish—Cyclic imines (spirolides, gymnodimines, pinnatoxins and pteriatoxins). EFSA J. 2010, 8, 1–39. [Google Scholar]
- Moroño, A.; Pazos, Y.; Doval, M.D.; Maneiro, J. Floraciones algales nocivas y condiciones oceanográficas en las rías gallegas durante 2001 y 2002. In VIII Reunión Ibérica sobre fitoplancton tóxico y biotoxinas, Proceedings of the VIII Reunión Ibérica sobre fitoplancton tóxico y biotoxinas, La Laguna, Spain, 3–5 November 2003; Norte, M., Fernández, J.J., Eds.; Campus: La Laguna, Spain, 2004; pp. 195–210. [Google Scholar]
- Pazos, Y.; Moroño, A. Microplancton tóxico y nocivo en las rías gallegas en los años 2003 a 2006. In Avances y tendencias en fitoplancton tóxico y biotoxinas, Proceedings of the IX Reunión Ibérica sobre Fitoplancton Tóxico y Biotoxinas, Cartagena, Spain, 7–10 May 2007; Gilabert, J., Ed.; Universidad Politécnica de Cartagena: Cartagena, Spain, 2008; pp. 13–28. [Google Scholar]
- Fernández, M.L.; Míguez, A.; Moroño, A.; Arévalo, F.; Pazos, Y.; Salgado, C.; Correa, J.; Blanco, J.; González-Gil, S.; Reguera, B. First report of pectenotoxin-2 in phytoplankton net-hauls and mussels from the Galician Rias Bajas during proliferations of Dinophysisacuta and Dinophysis caudata. In Proceedings of Forth International Conference on Molluscan Shellfish Safety, Santiago de Compostela, Spain, 4–8 June 2002; Villalba, A., Reguera, B., Romalde, J.R., Beiras, R., Eds.; Xunta de Galicia and IOC of UNESCO: Santiago de Compostela, Spain, 2003; pp. 75–83. [Google Scholar]
- Lamas, C.; Valverde, I.; Lago, J.; Vieites, J.M.; Cabado, A.G. Comparison between cytotoxic effects induced by PTX-2 in vitro in two human cell lines. Egypt. J. Nat. Toxins 2006, 3, 1–16. [Google Scholar]
- Leira, F.; Álvarez, C.; Vieites, J.M.; Vieytes, M.R.; Botana, L.M. Characterization of distinct apoptotic changes induced by okadaic acid and yessotoxin in the BE(2)-M17 neuroblastoma cell line. Toxicol. In Vitro 2002, 16, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Santaclara, F.; Lago, J.; Vieites, J.M.; Cabado, A.G. Effect of okadaic acid on integrins and structural proteins in BE(2)-M17 cells. Arch. Toxicol. 2005, 79, 582–586. [Google Scholar] [CrossRef] [PubMed]
- Valverde, I.; Lago, J.; Reboreda, A.; Vieites, J.M.; Cabado, A.G. Characteristics of palytoxin-induced cytotoxicity in neuroblastoma cells. Toxicol. In Vitro 2008, 22, 1432–1439. [Google Scholar] [CrossRef] [PubMed]
- Valverde, I.; Lago, J.; Vieites, J.M.; Cabado, A.G. Neuroblastoma cells as a cell model to study BADGE(bisphenol A diglycidyl-ether) and BFDGE (bisphenol F diglycidyl-ether) toxicity. In Focus on Neuroblastoma Research; Fernandes, J.A., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2007; pp. 119–147. [Google Scholar]
- Louzao, M.C.; Lago, J.; Ferreira, M.; Alfonso, A. Advances in knowledge of phycotoxins, new information about their toxicology and consequences on European legislation. In New Trends in Marine and Freshwater Toxins; Cabado, A.G., Vieites, J.M., Eds.; Nova Science: New York, NY, USA, 2013; pp. 149–202. [Google Scholar]
- Smienk, H.G.; Calvo, D.; Razquin, P.; Domínguez, E.; Mata, L. Single laboratory validation of ready-to-use phosphatase inhibition assay for detection of okadaic acid toxins. Toxins 2012, 4, 339–352. [Google Scholar] [CrossRef] [PubMed]
- Bourne, Y.; Radic, Z.; Aráoz, R.; Talley, T.T.; Benoit, E.; Servent, D.; Taylor, P.; Molgó, J.; Marchot, P. Structural determinants in phycotoxins and AChBP conferring high affinity binding and nicotinic AChR antagonism. Proc. Natl. Acad. Sci. USA 2010, 107, 6076–6081. [Google Scholar] [CrossRef] [PubMed]
- Munday, R.; Quilliam, M.A.; LeBlanc, P.; Lewis, N.; Gallant, P.; Sperker, S.A.; Ewart, H.S.; MacKinnon, S.L. Investigations into the toxicology of spirolides, a group of marine phycotoxins. Toxins 2012, 4, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cabado, A.G.; Leira, F.; Vieytes, M.; Vieites, J.M.; Botana, L.M. Cytoskeletal disruption is the key factor that triggers apoptosis in okadaic acid-treated neuroblastoma cells. Arch. Toxicol. 2004, 78, 74–85. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific opinion of the panel on contaminants in the food chain. Marine biotoxins in shellfish-yessotoxin group. EFSA J. 2008, 907, 1–62. [Google Scholar]
- Alfonso, A.; Alfonso, C. Yessotoxins, pharmacology and mechanism of action: Biological detection. In Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection; Botana, L.M., Ed.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2008; pp. 315–327. [Google Scholar]
- INTECMAR. Instituto Tecnolóxico para o Control do Medio Mariño de Galicia. Technological Institute for the Control of the Marine Environmet in Galicia. Available online: http://www.intecmar.org/ (accessed on 27 December 2014).
- (EU-RL-MB). EU-Harmonised Standard Operation Procedure for Determination of Lipophilic Marine Biotoxins in Molluscs by LC-MS/MS. Available online: http://aesan.msssi.gob.es/en/CRLMB/web/procedimientos_crlmb/crlmb_standard_operating_procedures.shtml (accessed on 12 August 2013).
- EFSA. Scientific opinion of the panel on contaminants in the food chain on a request from the European Commission on marine biotoxins in shellfish-Summary on regulated marine biotoxins. EFSA J. 2009, 1306, 1–23. [Google Scholar]
- European Commission. 2002/657/EC: Commission decision 2002/657/EC implementing Council directive 96/23/EC concerning the performance of analytical methods and the interpretation of results. Off. J. Eur. Communities 2002, L221, 8–36. [Google Scholar]
- Horwitz, W.; Albert, R. The Horwitz ratio (Horrat): A useful index of method performance with respect to precision. J. AOAC Int. 2006, 89, 1095–1109. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez, L.P.; González, V.; Martínez, A.; Paz, B.; Lago, J.; Cordeiro, V.; Blanco, L.; Vieites, J.M.; Cabado, A.G. Occurrence of Lipophilic Marine Toxins in Shellfish from Galicia (NW of Spain) and Synergies among Them. Mar. Drugs 2015, 13, 1666-1687. https://doi.org/10.3390/md13041666
Rodríguez LP, González V, Martínez A, Paz B, Lago J, Cordeiro V, Blanco L, Vieites JM, Cabado AG. Occurrence of Lipophilic Marine Toxins in Shellfish from Galicia (NW of Spain) and Synergies among Them. Marine Drugs. 2015; 13(4):1666-1687. https://doi.org/10.3390/md13041666
Chicago/Turabian StyleRodríguez, Laura P., Virginia González, Aníbal Martínez, Beatriz Paz, Jorge Lago, Victoria Cordeiro, Lucía Blanco, Juan Manuel Vieites, and Ana G. Cabado. 2015. "Occurrence of Lipophilic Marine Toxins in Shellfish from Galicia (NW of Spain) and Synergies among Them" Marine Drugs 13, no. 4: 1666-1687. https://doi.org/10.3390/md13041666
APA StyleRodríguez, L. P., González, V., Martínez, A., Paz, B., Lago, J., Cordeiro, V., Blanco, L., Vieites, J. M., & Cabado, A. G. (2015). Occurrence of Lipophilic Marine Toxins in Shellfish from Galicia (NW of Spain) and Synergies among Them. Marine Drugs, 13(4), 1666-1687. https://doi.org/10.3390/md13041666