
Identification of Antiviral Agents Targeting Hepatitis B Virus Promoter from Extracts of Indonesian Marine Organisms by a Novel Cell-Based Screening Assay




Abstract
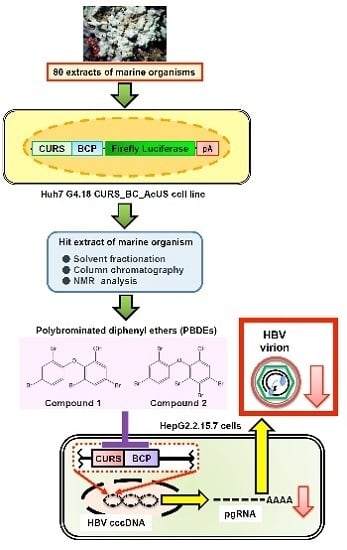
:
1. Introduction
2. Results and Discussion
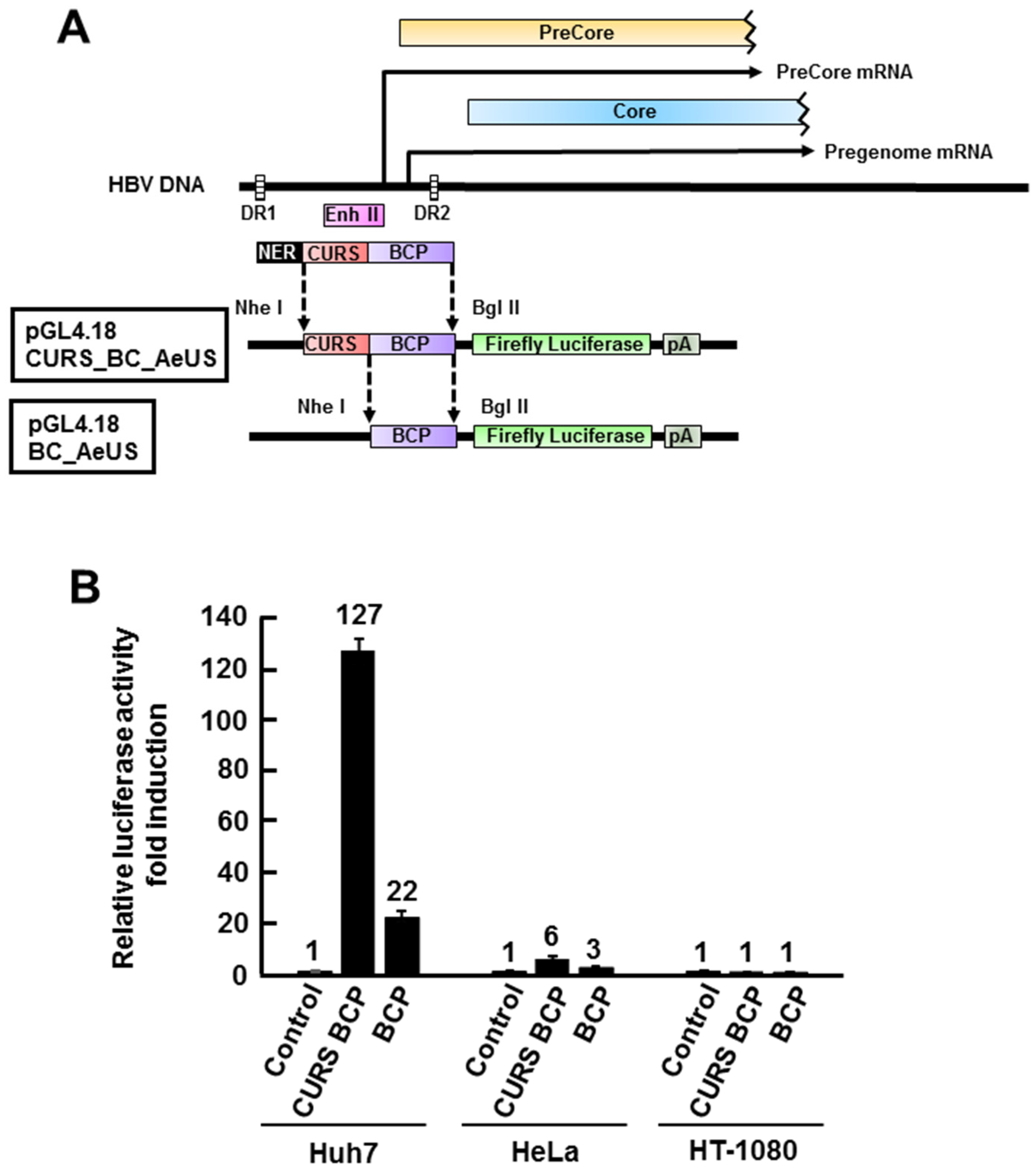
2.1. Establishment of HBV Core Promoter Reporter Cell Line
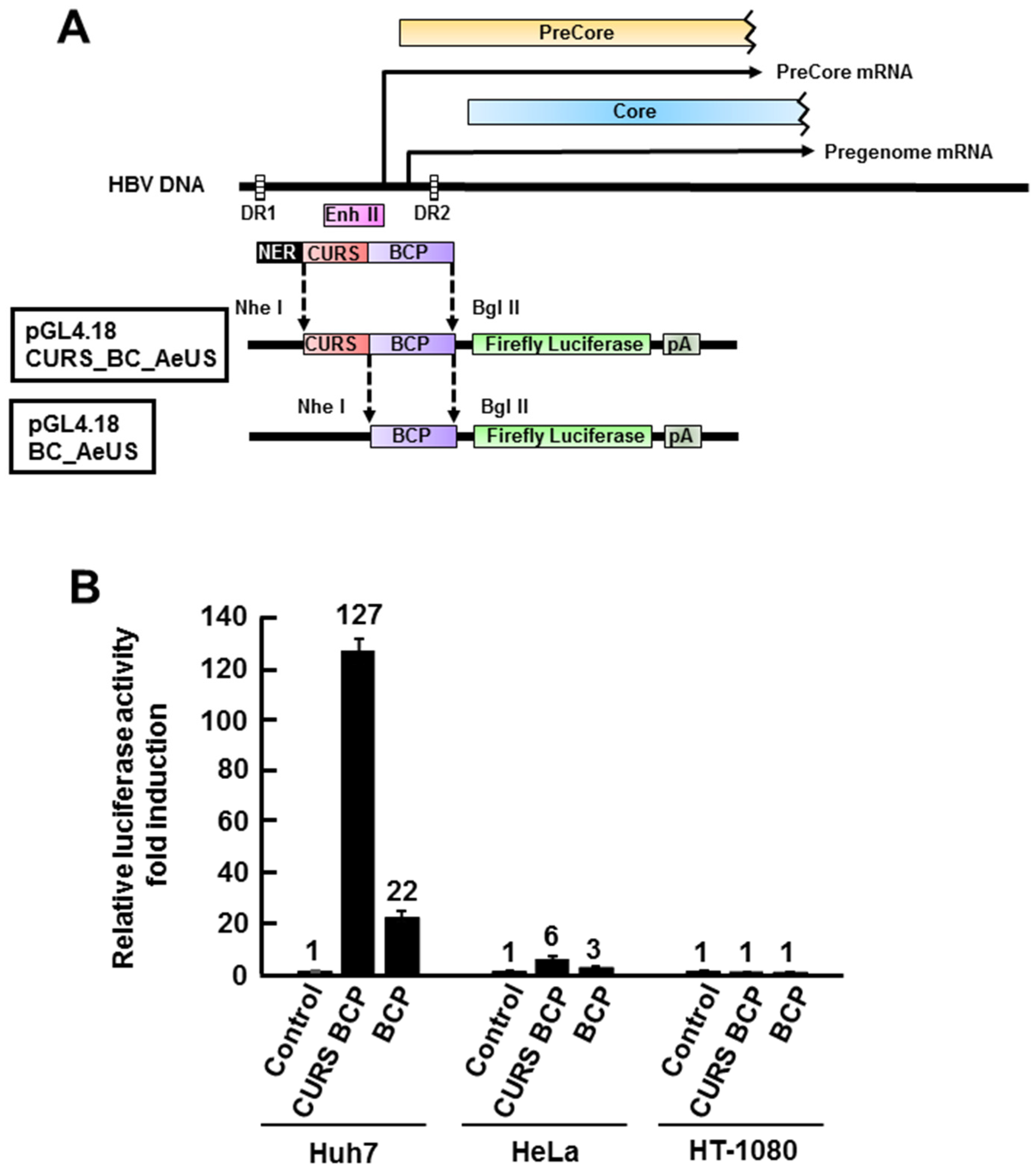
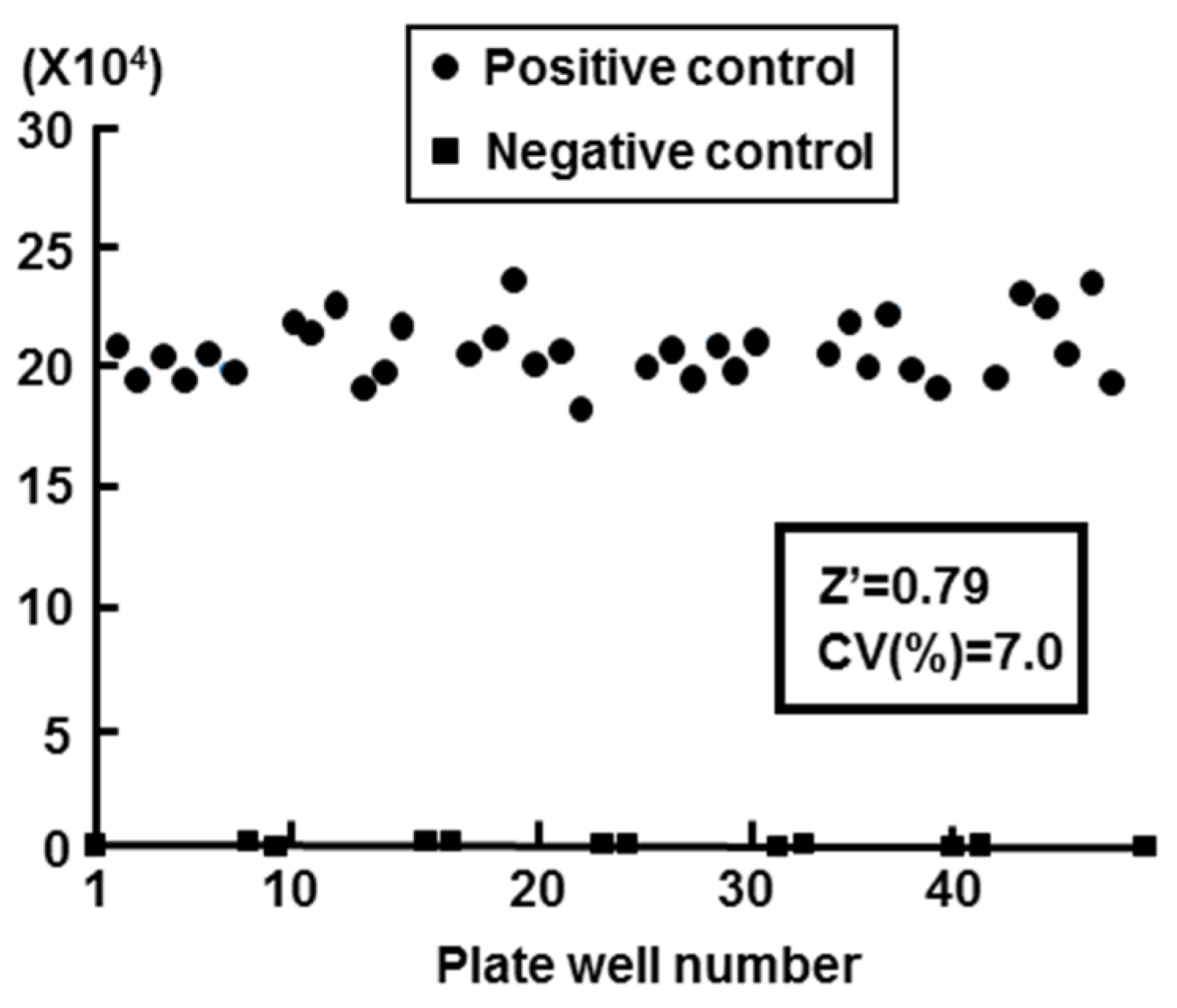
2.2. Validation of Cell-Based HBV Core Promoter Assay
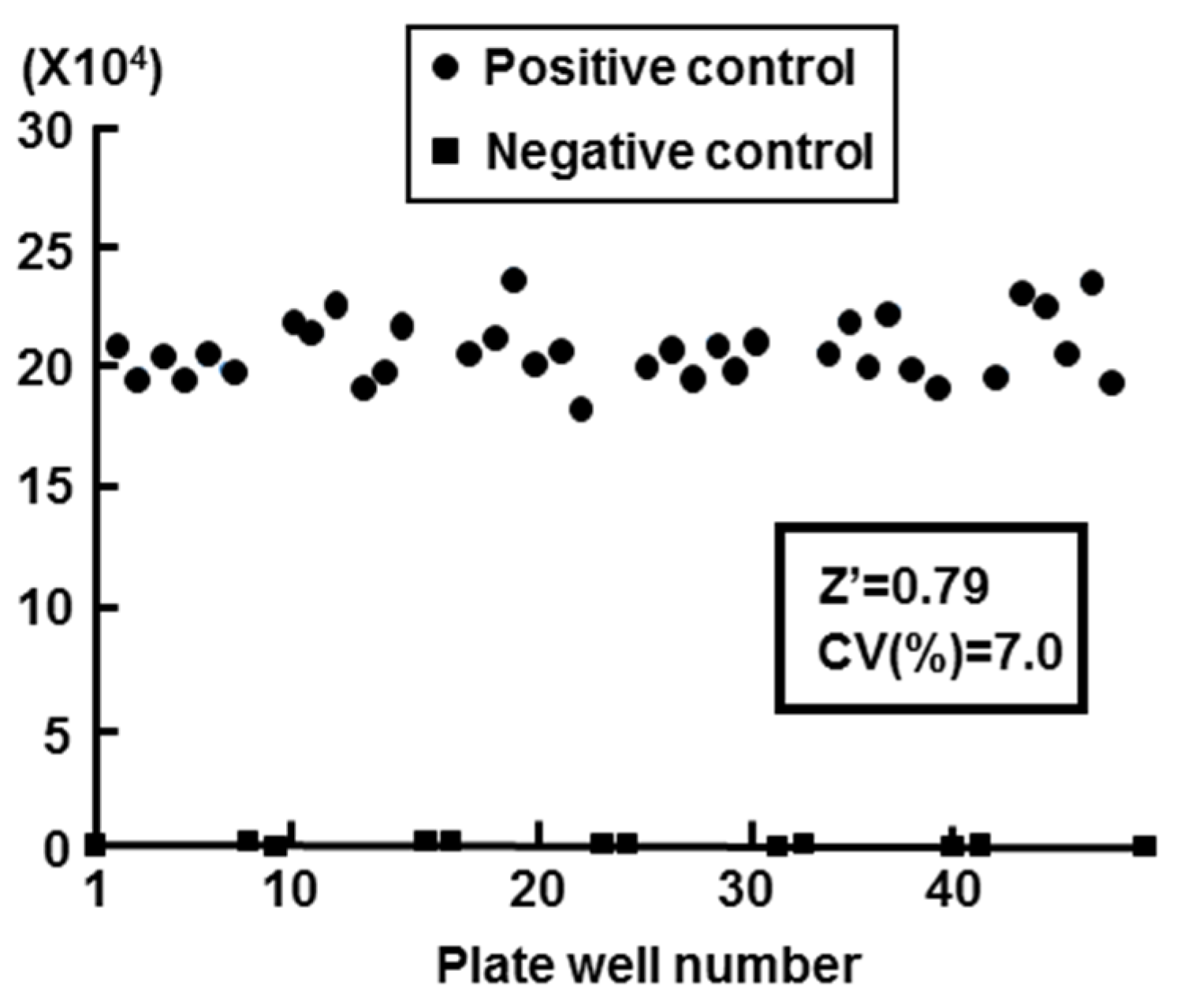
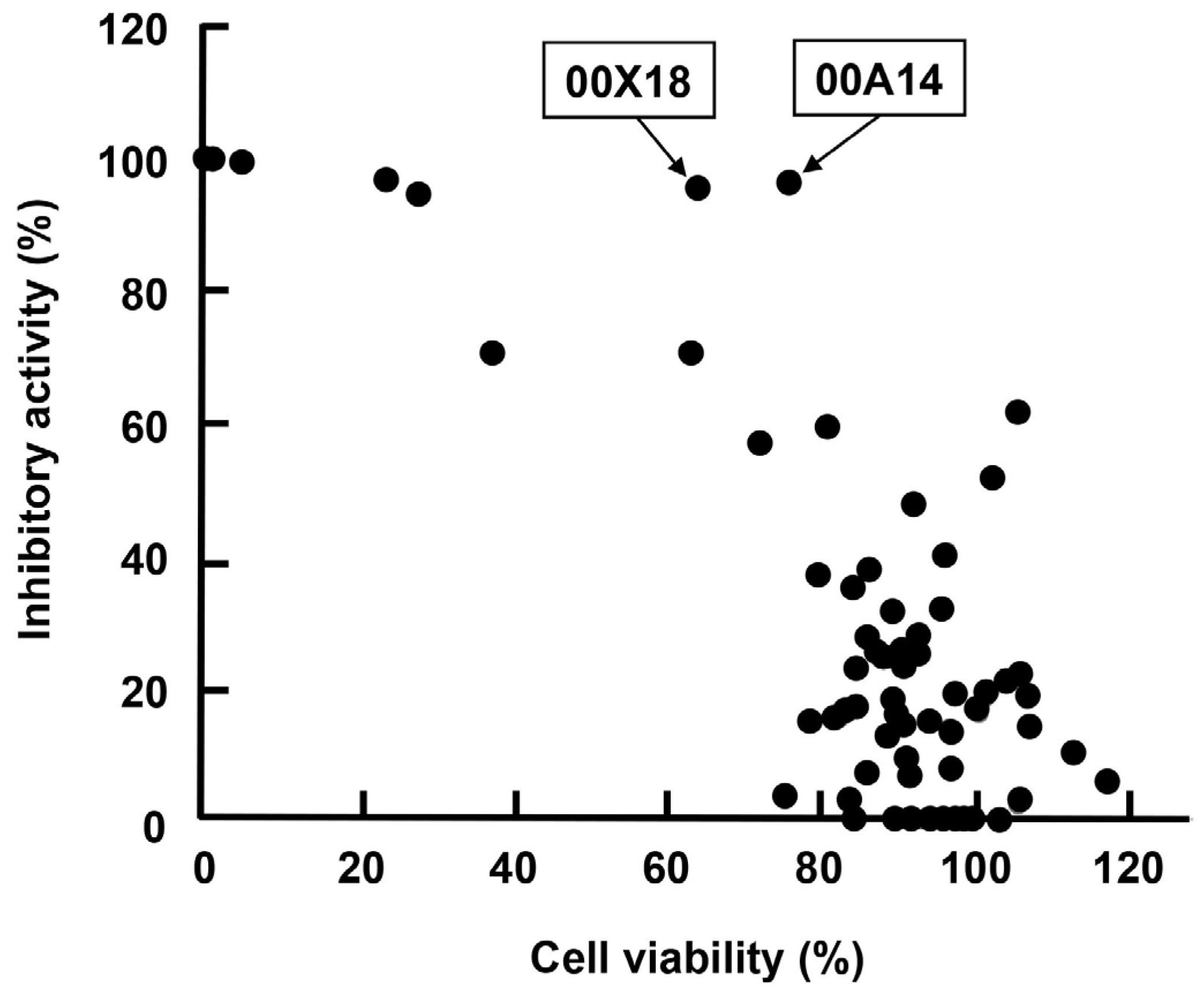
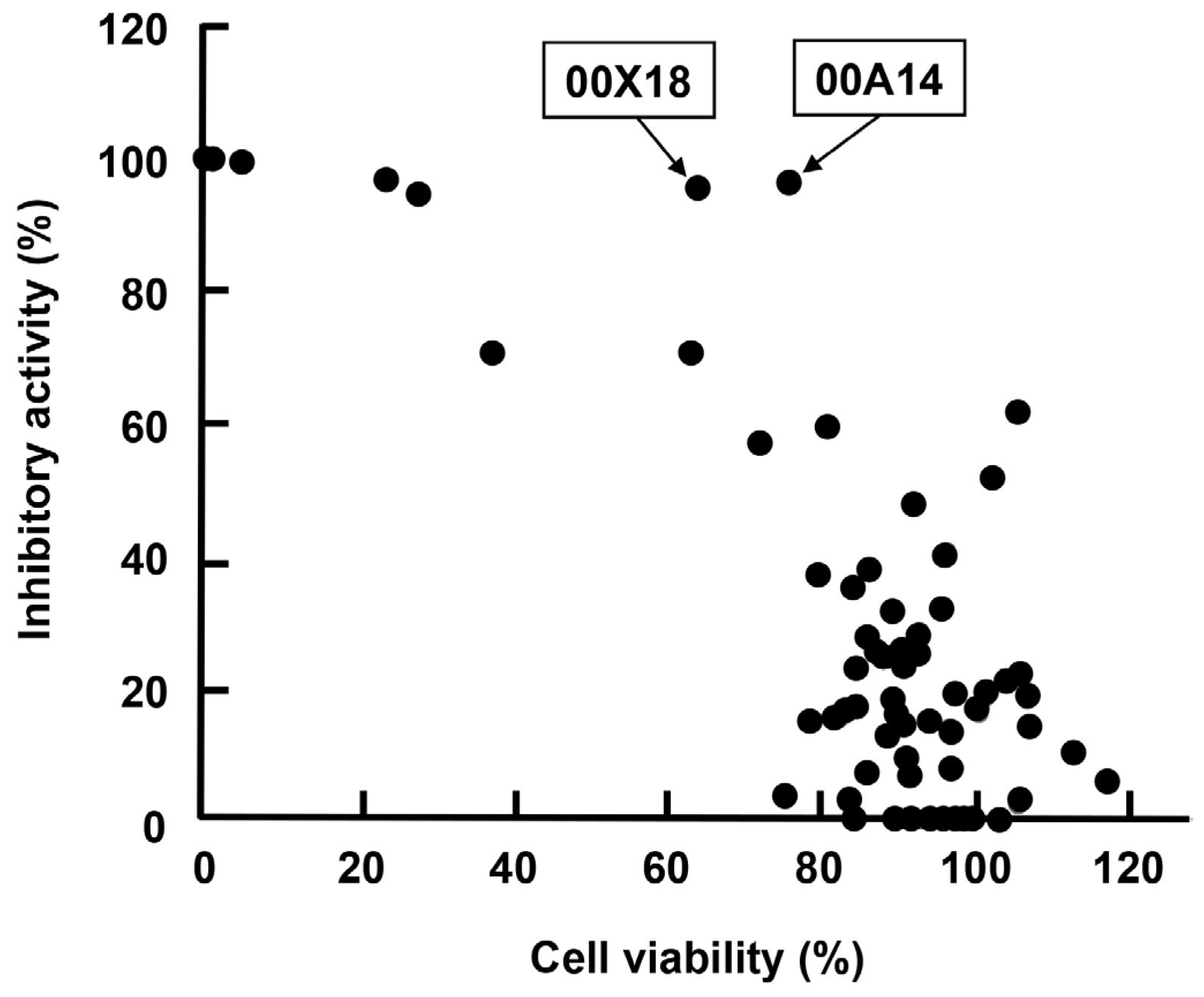
2.3. High-Throughput Screening for Extracts of Marine Organisms Inhibiting HBV Core Promoter Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Sample Code Name | Specimen | Phylum | Inhibitory Activity (%) | Cell Viability (% of Control) | Collection Site |
|---|---|---|---|---|---|---|
| 1 | 00A01 | Callyspongia sp. | Porifera | 0 | 101.5 | Simua Island |
| 2 | 00A05 | Xestospongia sp. | Porifera | 2.3 | 97.7 | Simua Island |
| 3 | 00A07 | Ircinia ramosa | Porifera | 0 | 177.4 | Simua Island |
| 4 | 00A08 | Liosina sp. | Porifera | 99.4 | 0 | Simua Island |
| 5 | 00A09 | Clathria sp. | Porifera | 98.6 | 0 | Simua Island |
| 6 | 00A10 | Unidentified | Chordata | 18.1 | 108.3 | Simua Island |
| 7 | 00A11 | Hippospongia sp. | Porifera | 40.3 | 97.7 | Simua Island |
| 8 | 00A12 | Petrosia sp. | Porifera | 6.9 | 92.8 | Simua Island |
| 9 | 00A13 | Callyspongia cf. aerizusa | Porifera | 5.4 | 119.1 | Simua Island |
| 10 | 00A14 | Dysidea granulosa | Porifera | 96.5 | 77.3 | Simua Island |
| 11 | 00B15 | Spheciospongia vagabunda | Porifera | 0 | 104.8 | Kajuongia Island |
| 12 | 00B16 | Callyspongia sp. | Porifera | 0 | 101.3 | Kajuongia Island |
| 13 | 00B17 | Unidentified | Porifera | 0 | 91.2 | Kajuongia Island |
| 14 | 00J85 | Unidentified | Porifera | 0 | 96.6 | Buton Island |
| 15 | 00J86 | Unidentified | Porifera | 0 | 97.6 | Buton Island |
| 16 | 00J87 | Unidentified | Porifera | 0 | 95.2 | Buton Island |
| 17 | 00J88 | Phyllospongia sp. | Porifera | 17.9 | 90.7 | Buton Island |
| 18 | 00J89 | Unidentified | Porifera | 0 | 101.3 | Buton Island |
| 19 | 00J90 | Unidentified | Porifera | 0 | 96.3 | Buton Island |
| 20 | 00J91 | Parazoanthus sp. | Cnidaria | 24.2 | 90.1 | Buton Island |
| 21 | 00K92 | Unidentified | Porifera | 14.8 | 95.9 | Buton Island |
| 22 | 00K94 | Ianthella basta | Porifera | 94.2 | 28.8 | Tobea Island |
| 23 | 00K95 | Unidentified | Chordata | 9.2 | 92.4 | Tobea Island |
| 24 | 00K97 | Higginsia mixta | Porifera | 16.1 | 84.5 | Tobea Island |
| 25 | 00L00 | Thrinacophora cervicornis | Porifera | 0 | 85.7 | Magintin Island |
| 26 | 00L02 | Unidentified | Chordata | 3.1 | 76.6 | Magintin Island |
| 27 | 00M03 | Gelliodes fibulata | Porifera | 13.7 | 98.2 | Masaloka Island |
| 28 | 00M04 | Clavularia viridis | Cnidaria | 99.6 | 1.4 | Masaloka Island |
| 29 | 00M05 | Coelocarteria sp. | Porifera | 15.1 | 91.1 | Masaloka Island |
| 30 | 00M06 | Mycale sp. | Porifera | 37.6 | 87.6 | Masaloka Island |
| 31 | 00M07 | Unidentified | Porifera | 28.2 | 94.2 | Masaloka Island |
| 32 | 00M08 | Unidentified | Porifera | 96.3 | 24.3 | Masaloka Island |
| 33 | 00N09 | Unidentified | Porifera | 15.3 | 83.7 | Buton strait |
| 34 | 00N10 | Myrmekioderma granulatum | Porifera | 0 | 92.9 | Buton strait |
| 35 | 00N11 | Callyspongia samarensis | Porifera | 0 | 95.5 | Buton strait |
| 36 | 00N12 | Biemna sp. | Porifera | 6.8 | 87.5 | Buton strait |
| 37 | 00N13 | Biemna triraphis | Porifera | 24.1 | 92.6 | Buton strait |
| 38 | 00N14 | Xestospongia exigua | Porifera | 99.4 | 1.6 | Buton strait |
| 39 | 00P16 | Unidentified | Cnidaria | 0 | 101.0 | Muna Island |
| 40 | 00Q17 | Unidentified | Porifera | 16.1 | 101.9 | Buton strait |
| 41 | 00Q18 | Unidentified | Porifera | 2.5 | 84.8 | Buton strait |
| 42 | 00Q19 | Axinyssa sp. | Porifera | 51.5 | 104.0 | Buton strait |
| 43 | 00Q20 | Mycale sp. | Porifera | 22.7 | 86.0 | Buton strait |
| 44 | 00R22 | Clavularia inflate | Cnidaria | 22.1 | 107.0 | Buton Island |
| 45 | 00R23 | Paralemnalia sp. | Cnidaria | 19.2 | 99.0 | Buton Island |
| 46 | 00R24 | Junceella fragilis | Cnidaria | 18.6 | 102.9 | Buton Island |
| 47 | 00R25 | Nephthea sp. | Cnidaria | 0 | 94.6 | Buton Island |
| 48 | 00S26 | Svenzea sp. | Porifera | 25.5 | 88.5 | Tobea Island |
| 49 | 00S27 | Unidentified | Cnidaria | 32.0 | 97.3 | Tobea Island |
| 50 | 00S28 | Coelogorgia sp. | Cnidaria | 0 | 97.7 | Tobea Island |
| 51 | 00T29 | Theonella sp. | Porifera | 35.3 | 85.4 | Tobea Island |
| 52 | 00T30 | Unidentified | Porifera | 47.6 | 93.6 | Tobea Island |
| 53 | 00T31 | Higginsia cf. mixta | Porifera | 14.7 | 142.0 | Tobea Island |
| 54 | 00T32 | Paratelesto sp. | Cnidaria | 0 | 94.7 | Tobea Island |
| 55 | 00U33 | Pycnoclabella sp. | Chordata | 25.8 | 91.9 | Muna Island |
| 56 | 00U34 | Lissoclinum patella | Chordata | 27.8 | 87.2 | Muna Island |
| 57 | 00X01 | Polycarpa contecta | Chordata | 25.9 | 93.6 | Simua Island |
| 58 | 00X02 | Dysidea sp. | Porifera | 56.8 | 73.4 | Tobea Island |
| 59 | 00X04 | Nephthea sp. | Cnidaria | 0 | 99.2 | Beromasidi Island |
| 60 | 00X05 | Haliclona fascigera | Porifera | 7.8 | 98.7 | Torobulu |
| 61 | 00X06 | Axinyssa sp. | Porifera | 98.9 | 4.9 | Torobulu |
| 62 | 00X07 | Unidentified | Porifera | 16.7 | 86.0 | Torobulu |
| 63 | 00X08 | Unidentified | Cnidaria | 12.9 | 87.0 | Torobulu |
| 64 | 00X10 | Niphates olemda | Porifera | 70.2 | 38.5 | Buton Island |
| 65 | 00X11 | Unidentified | Porifera | 99.7 | 1.1 | Tobea Island |
| 66 | 00X12 | Unidentified | Porifera | 70.6 | 63.9 | Tobea Island |
| 67 | 00X13 | Unidentified | Porifera | 13.9 | 108.7 | Magintin Island |
| 68 | 00X14 | Xestospongia sp. | Porifera | 21.1 | 105.6 | Magintin Island |
| 69 | 00X15 | Dysidea/Euryspongia | Porifera | 10.2 | 114.4 | Magintin Island |
| 70 | 00X16 | Unidentified | Chordata | 0 | 130.3 | Buton strait |
| 71 | 00X17 | Dysidea cf. arenaria | Porifera | 14.7 | 80.0 | Buton strait |
| 72 | 00X18 | Dysidea sp. | Porifera | 95.0 | 65.3 | Buton strait |
| 73 | 00X19 | Unidentified | Porifera | 23.0 | 92.5 | Buton strait |
| 74 | 00X21 | Gelliodes/Niphates | Porifera | 36.7 | 80.9 | Buton strait |
| 75 | 00X22 | Amphimedon/Haliclona | Porifera | 31.2 | 90.9 | Buton strait |
| 76 | 00X23 | Dysidea cf. arenaria | Porifera | 0 | 92.8 | Buton strait |
| 77 | 00X24 | Unidentified | Porifera | 0 | 99.2 | Buton strait |
| 78 | 00X26 | Anthelia sp. | Cnidaria | 61.5 | 107.4 | Buton strait |
| 79 | 00X27 | Unidentified | Chordata | 14.8 | 85.2 | Tobea Island |
| 80 | 00X28 | Clathria sp. | Porifera | 58.9 | 82.0 | Tobea Island |
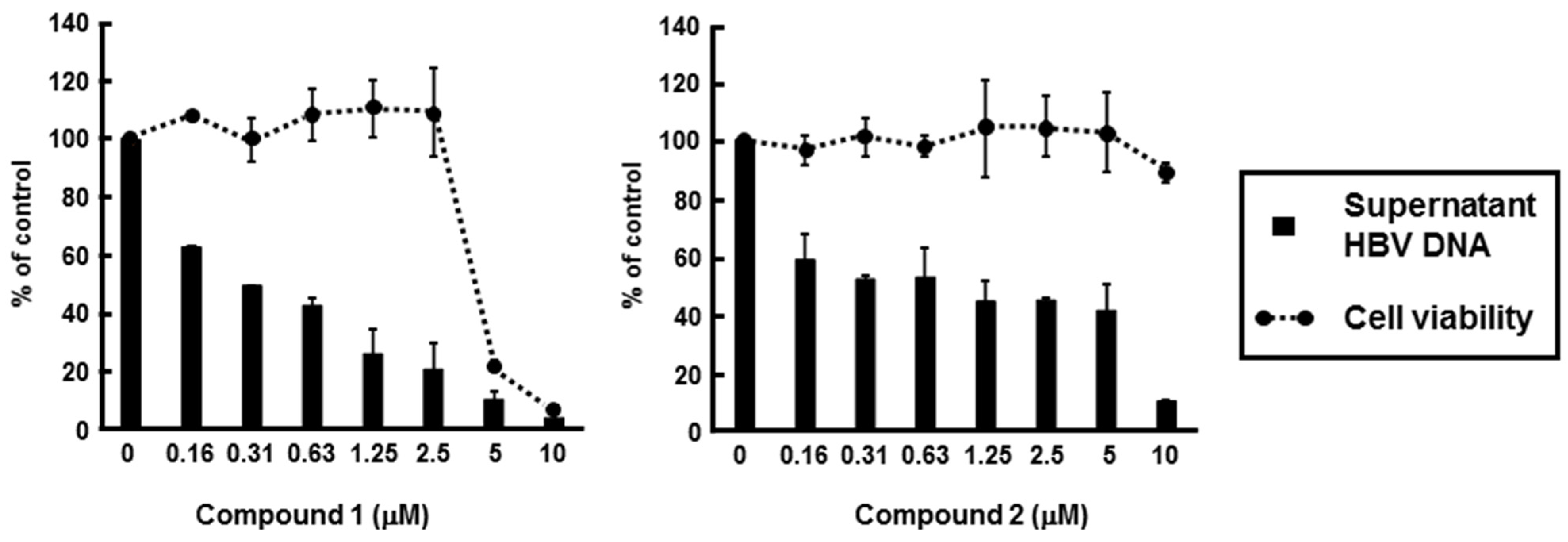
2.4. Identification of PBDEs as the Inhibitory Compounds of HBV Production via HBV Core Promoter Activity

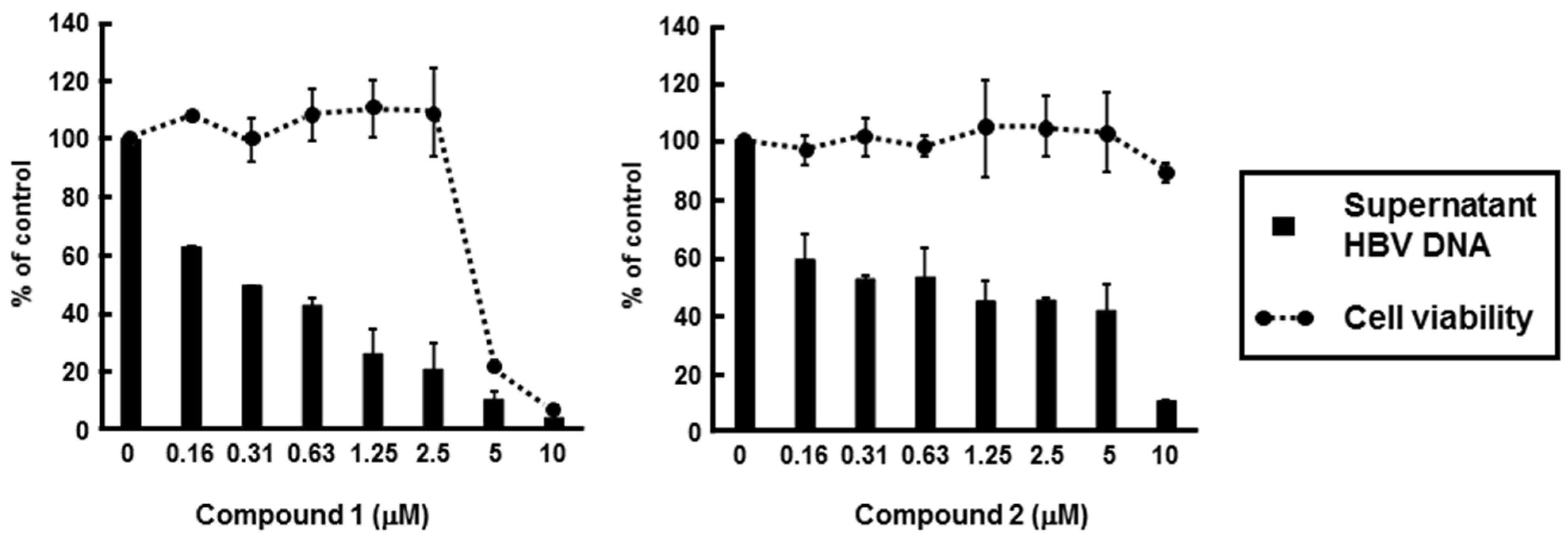
| Compound | EC50 a (µM) | CC50 b (µM) | Selectivity c Index |
|---|---|---|---|
| Compound 1 | 0.23 ± 0.07 | 4.19 ± 0.12 | 18.2 |
| Compound 2 | 0.80 ± 0.34 | 10.26 ± 3.69 | 12.8 |
| Entecavir | 0.021 ± 0.003 | >100 | >4761 |
3. Experimental Section
3.1. Cell Culture
3.2. Plasmid Construction and Transient or Stable Expression
3.3. Validation of Screening Method
3.4. Cell-Based HBV Promoter Assay
3.5. Determination of Cytotoxicity
3.6. Preparation of Extracts from Marine Organisms
3.7. Separation of PBDEs
3.8. Determination of Anti HBV Activity and Cytotoxicity in HepG2.2.15.7 Cells
3.9. Reagents
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ott, J.J.; Stevens, G.A.; Groeger, J.; Wiersma, S.T. Global epidemiology of hepatitis B virus infection: New estimates of age-specific HBsAg seroprevalence and endemicity. Vaccine 2012, 30, 2212–2219. [Google Scholar] [CrossRef] [PubMed]
- Liaw, Y.F.; Chu, C.M. Hepatitis B virus infection. Lancet 2009, 373, 582–592. [Google Scholar] [CrossRef]
- Trepo, C.; Chan, H.L.; Lok, A. Hepatitis B virus infection. Lancet 2014, 384, 2053–2063. [Google Scholar] [CrossRef]
- Locarnini, S.; Littlejohn, M.; Aziz, M.N.; Yuen, L. Possible origins and evolution of the hepatitis B virus (HBV). Semin. Cancer Biol. 2013, 23, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.; Nassal, M. Hepatitis B virus replication. World J. Gastroenterol. 2007, 13, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Kramvis, A.; Kew, M.C. The core promoter of hepatitis B virus. J. Viral Hepat. 1999, 6, 415–427. [Google Scholar] [CrossRef] [PubMed]
- Moolla, N.; Kew, M.; Arbuthnot, P. Regulatory elements of hepatitis B virus transcription. J. Viral Hepat. 2002, 9, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Nassal, M. Hepatitis B virus replication: Novel roles for virus-host interactions. Intervirology 1999, 42, 100–116. [Google Scholar] [CrossRef] [PubMed]
- Seeger, C.; Mason, W.S. Hepatitis B virus biology. Microbiol. Mol. Biol. Rev. 2000, 64, 51–68. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Banks, K.E.; Anderson, A.L.; McLachlan, A. Hepatitis B virus transcription and replication. Drug News Perspect. 2001, 14, 325–334. [Google Scholar] [PubMed]
- Lok, A.S. Personalized treatment of hepatitis B. Clin. Mol. Hepatol. 2015, 21, 1–6. [Google Scholar] [CrossRef] [PubMed]
- You, C.R.; Lee, S.W.; Jang, J.W.; Yoon, S.K. Update on hepatitis B virus infection. World J. Gastroenterol. 2014, 20, 13293–13305. [Google Scholar] [CrossRef] [PubMed]
- Zoulim, F.; Durantel, D. Antiviral therapies and prospects for a cure of chronic hepatitis B. Cold Spring Harb. Perspect. Med. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Gish, R.; Jia, J.D.; Locarnini, S.; Zoulim, F. Selection of chronic hepatitis B therapy with high barrier to resistance. Lancet Infect. Dis. 2012, 12, 341–353. [Google Scholar] [CrossRef]
- Grimm, D.; Thimme, R.; Blum, H.E. HBV life cycle and novel drug targets. Hepatol. Int. 2011, 5, 644–653. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Newman, D.J. Natural products: A continuing source of novel drug leads. Biochim. Biophys. Acta 2013, 1830, 3670–3695. [Google Scholar] [CrossRef] [PubMed]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Donia, M.; Hamann, M.T. Marine natural products and their potential applications as anti-infective agents. Lancet Infect. Dis. 2003, 3, 338–348. [Google Scholar] [CrossRef]
- Sagar, S.; Kaur, M.; Minneman, K.P. Antiviral lead compounds from marine sponges. Mar. Drugs 2010, 8, 2619–2638. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Chen, M.; Yen, T.S.; Ou, J.H. Hepatocyte-specific expression of the hepatitis B virus core promoter depends on both positive and negative regulation. Mol. Cell. Biol. 1993, 13, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Honigwachs, J.; Faktor, O.; Dikstein, R.; Shaul, Y.; Laub, O. Liver-specific expression of hepatitis B virus is determined by the combined action of the core gene promoter and the enhancer. J. Virol. 1989, 63, 919–924. [Google Scholar] [PubMed]
- Yee, J.K. A liver-specific enhancer in the core promoter region of human hepatitis B virus. Science 1989, 246, 658–661. [Google Scholar] [CrossRef] [PubMed]
- Yuh, C.H.; Ting, L.P. The genome of hepatitis B virus contains a second enhancer: Cooperation of two elements within this enhancer is required for its function. J. Virol. 1990, 64, 4281–4287. [Google Scholar] [PubMed]
- Fatokun, A.A.; Liu, J.O.; Dawson, V.L.; Dawson, T.M. Identification through high-throughput screening of 4′-methoxyflavone and 3′,4′-dimethoxyflavone as novel neuroprotective inhibitors of parthanatos. Br. J. Pharmacol. 2013, 169, 1263–1278. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Chung, T.D.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, V.; El Gamal, A.A.; Yamanaka, K.; Poth, D.; Kersten, R.D.; Schorn, M.; Allen, E.E.; Moore, B.S. Biosynthesis of polybrominated aromatic organic compounds by marine bacteria. Nat. Chem. Biol. 2014, 10, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Teuten, E.L.; Xu, L.; Reddy, C.M. Two abundant bioaccumulated halogenated compounds are natural products. Science 2005, 307, 917–920. [Google Scholar] [CrossRef] [PubMed]
- Handayani, D.; Edrada, R.A.; Proksch, P.; Wray, V.; Witte, L.; van Soest, R.W.; Kunzmann, A.; Soedarsono. Four new bioactive polybrominated diphenyl ethers of the sponge Dysidea herbacea from west sumatra, indonesia. J. Nat. Prod. 1997, 60, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Hanif, N.; Tanaka, J.; Setiawan, A.; Trianto, A.; de Voogd, N.J.; Murni, A.; Tanaka, C.; Higa, T. Polybrominated diphenyl ethers from the indonesian sponge Lamellodysidea herbacea. J. Nat. Prod. 2007, 70, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Sionov, E.; Roth, D.; Sandovsky-Losica, H.; Kashman, Y.; Rudi, A.; Chill, L.; Berdicevsky, I.; Segal, E. Antifungal effect and possible mode of activity of a compound from the marine sponge Dysidea herbacea. J. Infect. 2005, 50, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Schmitz, F.J.; Govindan, M.; Abbas, S.A.; Hanson, K.M.; Horton, P.A.; Crews, P.; Laney, M.; Schatzman, R.C. Enzyme inhibitors: New and known polybrominated phenols and diphenyl ethers from four indo-pacific Dysidea sponges. J. Nat. Prod. 1995, 58, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Salam, K.A.; Furuta, A.; Noda, N.; Tsuneda, S.; Sekiguchi, Y.; Yamashita, A.; Moriishi, K.; Nakakoshi, M.; Tani, H.; Roy, S.R.; et al. Pbde: Structure-activity studies for the inhibition of hepatitis C virus NS3 helicase. Molecules 2014, 19, 4006–4020. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, H.; Sumilat, D.A.; Kanno, S.; Ukai, K.; Rotinsulu, H.; Wewengkang, D.S.; Ishikawa, M.; Mangindaan, R.E.; Namikoshi, M. A polybromodiphenyl ether from an indonesian marine sponge Lamellodysidea herbacea and its chemical derivatives inhibit protein tyrosine phosphatase 1B, an important target for diabetes treatment. J. Nat. Med. 2013, 67, 730–735. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.M.; Johnson, R.K.; Hecht, S.M. Polybrominated diphenyl ethers from a sponge of the Dysidea genus that inhibit Tie2 kinase. Bioorg. Med. Chem. 2005, 13, 657–659. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Skildum, A.; Stromquist, E.; Rose-Hellekant, T.; Chang, L.C. Bioactive polybrominated diphenyl ethers from the marine sponge Dysidea sp. J. Nat. Prod. 2008, 71, 262–264. [Google Scholar] [CrossRef] [PubMed]
- Cohen, D.; Adamovich, Y.; Reuven, N.; Shaul, Y. Hepatitis B virus activates deoxynucleotide synthesis in nondividing hepatocytes by targeting the R2 gene. Hepatology 2010, 51, 1538–1546. [Google Scholar] [CrossRef] [PubMed]
- Ogura, N.; Watashi, K.; Noguchi, T.; Wakita, T. Formation of covalently closed circular DNA in Hep38.7-Tet cells, a tetracycline inducible hepatitis B virus expression cell line. Biochem. Biophys. Res. Commun. 2014, 452, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Sells, M.A.; Chen, M.L.; Acs, G. Production of hepatitis B virus particles in Hep G2 cells transfected with cloned hepatitis B virus DNA. Proc. Natl. Acad. Sci. USA 1987, 84, 1005–1009. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, M.; Tanaka, Y.; Kato, T.; Orito, E.; Ito, K.; Acharya, S.K.; Gish, R.G.; Kramvis, A.; Shimada, T.; Izumi, N.; et al. Influence of hepatitis B virus genotypes on the intra- and extracellular expression of viral DNA and antigens. Hepatology 2006, 44, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Hussain, M.; Wong, S.; Fung, S.K.; Yim, H.J.; Lok, A.S. A genotype-independent real-time PCR assay for quantification of hepatitis B virus DNA. J. Clin. Microbiol. 2007, 45, 553–558. [Google Scholar] [CrossRef] [PubMed][Green Version]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamashita, A.; Fujimoto, Y.; Tamaki, M.; Setiawan, A.; Tanaka, T.; Okuyama-Dobashi, K.; Kasai, H.; Watashi, K.; Wakita, T.; Toyama, M.; et al. Identification of Antiviral Agents Targeting Hepatitis B Virus Promoter from Extracts of Indonesian Marine Organisms by a Novel Cell-Based Screening Assay. Mar. Drugs 2015, 13, 6759-6773. https://doi.org/10.3390/md13116759
Yamashita A, Fujimoto Y, Tamaki M, Setiawan A, Tanaka T, Okuyama-Dobashi K, Kasai H, Watashi K, Wakita T, Toyama M, et al. Identification of Antiviral Agents Targeting Hepatitis B Virus Promoter from Extracts of Indonesian Marine Organisms by a Novel Cell-Based Screening Assay. Marine Drugs. 2015; 13(11):6759-6773. https://doi.org/10.3390/md13116759
Chicago/Turabian StyleYamashita, Atsuya, Yuusuke Fujimoto, Mayumi Tamaki, Andi Setiawan, Tomohisa Tanaka, Kaori Okuyama-Dobashi, Hirotake Kasai, Koichi Watashi, Takaji Wakita, Masaaki Toyama, and et al. 2015. "Identification of Antiviral Agents Targeting Hepatitis B Virus Promoter from Extracts of Indonesian Marine Organisms by a Novel Cell-Based Screening Assay" Marine Drugs 13, no. 11: 6759-6773. https://doi.org/10.3390/md13116759
APA StyleYamashita, A., Fujimoto, Y., Tamaki, M., Setiawan, A., Tanaka, T., Okuyama-Dobashi, K., Kasai, H., Watashi, K., Wakita, T., Toyama, M., Baba, M., De Voogd, N. J., Maekawa, S., Enomoto, N., Tanaka, J., & Moriishi, K. (2015). Identification of Antiviral Agents Targeting Hepatitis B Virus Promoter from Extracts of Indonesian Marine Organisms by a Novel Cell-Based Screening Assay. Marine Drugs, 13(11), 6759-6773. https://doi.org/10.3390/md13116759