Abstract
Five new dibenzoxazepinone derivatives, mycemycins A–E (1–5), were isolated from the ethanol extracts of mycelia of two different streptomycetes. 1 and 2 were isolated from an acidic red soil-derived strain, Streptomyces sp. FXJ1.235, and 3–5 from a gntR gene-disrupted deep-sea strain named Streptomyces olivaceus FXJ8.012Δ1741. The structures of mycemycins were elucidated by a combination of spectroscopic analyses, including 1D- and 2D-NMR techniques.
1. Introduction
Actinomycetes are a proven source of diverse natural products and account for more than 40% of the currently known microbial bioactive compounds, including antibiotics, enzyme inhibitors, immunoactive agents, etc. [1,2,3]. While the great majority of bioactive compound-producing actinomycetes are from terrestrial habitats, in recent years, marine actinomycetes have become an up-and-coming source of novel natural products [4,5]. At the same time, increasing genome sequencing data of actinomycetes exhibit far more putative secondary metabolic gene clusters than the “active” clusters and relevant natural compounds, which means that the vast majority of the secondary metabolic potential remains untapped [6], and reinforces the idea that reexamination of well-studied actinomycete strains is still meaningful [7]. To explore new products of these silent gene clusters, several approaches have been developed including culture medium optimization, strain improvement, manipulation of global or pathway-specific regulators, heterologous expression, ribosome engineering, co-cultivation, etc. [8].
Recently, we reported tetroazolemycins and spoxazomicins from a deep-sea derived actinomycete, Streptomyces olivaceus strain FXJ8.012, which was isolated from a water sample collected from the south Indian Ocean [9]. A genome survey of this strain indicated that it contains a number of silent gene clusters that may be responsible for the biosynthesis of novel compounds, including halometabolites. By disruption of a putative transcriptional regulator gene orf-1741, we successfully activated the production of a cluster of novel natural products (3–5, Figure 1). Interestingly, these compounds exhibited similar UV spectra with another two compounds (1–2, Figure 1) that we have isolated from wild type Streptomyces sp. strain FXJ1.235, a terrestrial actinomycete derived from an acidic red soil sample collected from Jiangxi Province, China. Furthermore, all the compounds except 1 exhibited halogenated traits according to MS analysis. So we decided to study them together. Results demonstrated that they do possess high structural similarity. Herein we describe the isolation, structural elucidation and bioactivities of these novel compounds.
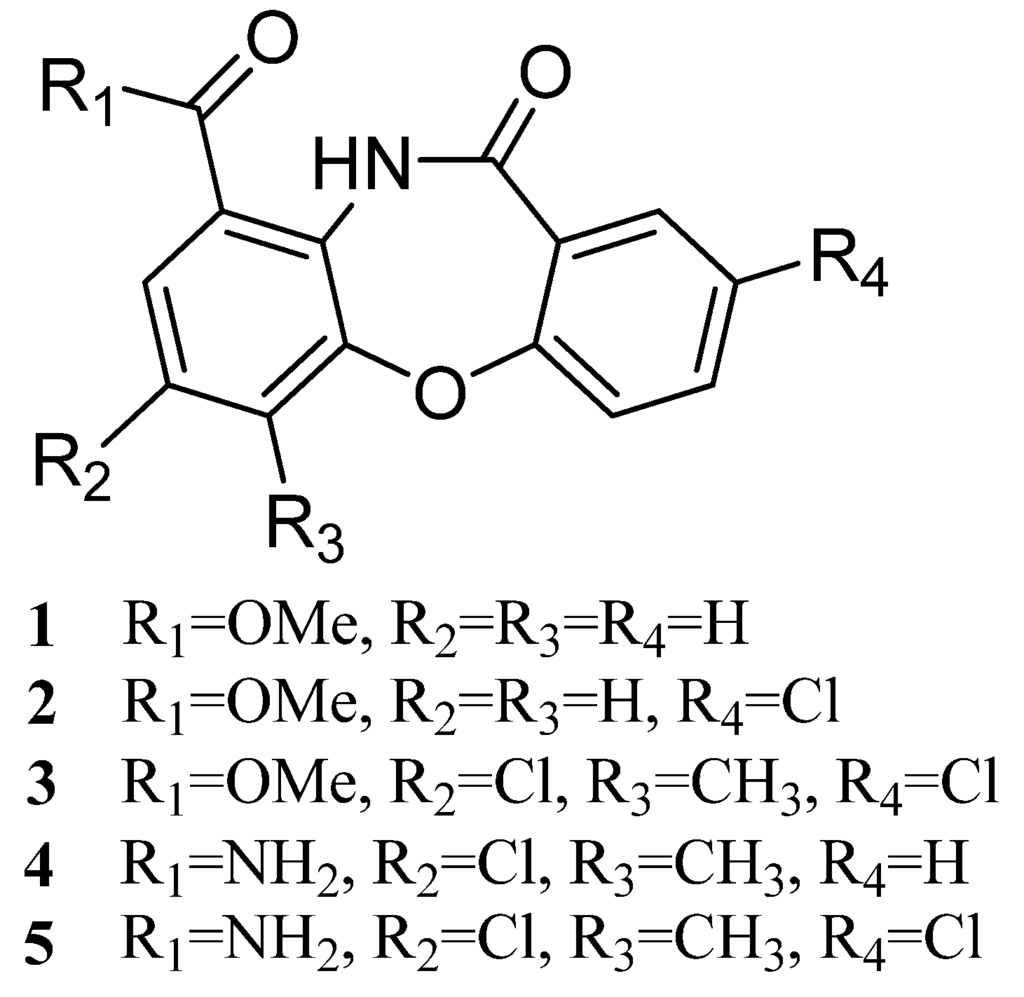
Figure 1.
Structures of compounds 1–5.
Figure 1.
Structures of compounds 1–5.
2. Results and Discussion
2.1. Genetic Manipulation of S. olivaceus FXJ8.012 and Isolation of Compounds 1–5
In the genome of S. olivaceus FXJ8.012, orf-1741 is flanked by putative secondary metabolic genes and encodes a 126-amino-acid peptide which was proposed as a member of GntR-family transcriptional regulator. GntR-like regulators play an important role in the control of actinomycete development and antibiotic biosynthesis [10,11]. Some members of the GntR family have previously been reported as repressors of secondary metabolite biosynthesis in actinomycetes. For example, MtdA represses nucleoside antibiotic A201A [12], SCO3269 represses actinorhodin and undecylprodigiosin [13], and ptnR1 represses platensimycin [14]. Thus, in order to activate the expression of related silent secondary metabolic gene clusters, we disrupted orf-1741 by replacing it with a kanamycin-resistance gene (neo). The disruption resulted in a different HPLC-based metabolic profile of the mutant strain FXJ8.012Δ1741 cultivated in modified R2 medium [15]. Consequently, three new halogenated dibenzoxazepinone derivatives (3–5) were detected from the mutant strain, while no new products were detected from the wild type strain (Supplementary Information Figure S1). Purification of the ethanol extracts of a 20 L fermentation culture of FXJ8.012Δ1741 led to the isolation of mycemycins C–E (3–5).
In addition, purification of the ethanol extracts of a 10 L fermentation culture of S. sp. FXJ1.235 led to the isolation of mycemycins A and B (1–2), with much higher yields than 3 and 4 and better solubility than 5. So we chose to elucidate the structures of compounds 1 and 2 first.
2.2. Structural Identification of Mycemycins A–E
Mycemycins A (1) was obtained as a white solid. Its molecular formula was established as C15H11NO4 according to the [M + H]+ peak at m/z 270.0776 (calcd for C15H12NO4, 270.0766) in the high resolution-electrospray ionization-mass (HR-ESI-MS) spectrum combined with the 13C-nuclear magnetic resonance (NMR) data, corresponding to 11 degrees of unsaturation.
The 1H NMR spectrum (Table 1 and Figure S2) showed seven aromatic protons, with four main doublets at δ 7.15, 7.81, 8.04 and 8.08 and three main triplets at δ 7.03, 7.46 and 7.48, and one methoxyl group at δ 4.06. Besides conventional coupling correlations, the aromatic “w” type coupling could also be found from protons at δ 7.48 and 8.04 with small J = 1.5 Hz. The 13C NMR spectrum (Table 1 and Figure S2) showed 15 carbon signals, and, combined with the DEPT and HSQC spectra (Figure S2), revealed one methoxyl group (δ 52.4), seven phenyl methine groups (δ 115.0, 117.7, 119.6, 124.8, 127.2, 127.5 and 134.3), and seven quaternary carbons (δ 110.0, 121.3, 139.4, 149.8, 159.4, and two possible ester/amido carbon signals at δ 164.4 and 165.6).
Detailed analysis of 2D-NMR spectra (Figure 2 and Figure S2) revealed that 1 was comprised of two aromatic ring systems. The 1H-1H correlation spectroscopy (COSY) spectrum established the connections of H11(δ 8.08)-H10 (δ 7.46)-H9 (δ 8.04); the heteronuclear multiple bond correlation (HMBC) experiment revealed the key correlations from H11 and 15-Me to C14, H9 and H11 to C13, and from H9 and H10 to C12 and C8. According to the above data, the first aromatic ring from C8 to C15 was established. Furthermore, the chemical shifts of C15 (δ 52.4) and C8 (δ 149.8) indicated that both of the two carbons were connected to an oxygen atom. The COSY spectrum revealed the connection of H3 (δ 8.04)-H4 (δ 7.03)-H5 (δ 7.48)-H6 (δ 7.15); the HMBC correlations from H3 to C1 and from H4 and H6 to C2 suggested the connections of C1-C2-C3; the HMBC correlations from H3, H5 and H6 to C7 and the chemical shift of C7 (δ 159.4) revealed the connections of C2-C7-C6 where C7 was connected to an oxygen atom. Based on these data, the second aromatic ring from C1 to C7 was established. Moreover, according to the supposed molecular formula, the two ring systems should connect with each other though C7-O-C8 and C11-NH-C13.

Table 1.
1H- (500 MHz) and 13C-NMR (125 MHz) data of 1–4 in CDCl3 (δ in ppm, J in Hz).
| Position | 1 | 2 | 3 | 4 | ||||
|---|---|---|---|---|---|---|---|---|
| δH | δC | δH | δC | δH | δC | δH | δC | |
| 1 | ― | 164.4, C | ― | 163.1, C | ― | 163.3, C | ― | 163.9, C |
| 2 | ― | 110, C | ― | 111, C | ― | 110.8, C | ― | 109.8, C |
| 3 | 8.04 (dd, 1.5, 8.0) | 127.2, CH | 8.01 (d, 2.5) | 126.4, CH | 8.00 (d, 2.5) | 128.4, CH | 8.10 (d, 7.5) | 127.83, CH |
| 4 | 7.03 (t, 7.5) | 119.6, CH | ― | 124.5, C | ― | 124.5, C | 7.10 (t, 7.5) | 120.4, CH |
| 5 | 7.48 (dt, 1.5, 7.8) | 134.3, CH | 7.42 (dd, 2.5, 8.5–9.0) | 134.1, CH | 7.43 (dd, 2.5, 8.5–9.0) | 134.2, CH | 7.53 (t, 7.5) | 134.9, CH |
| 6 | 7.15 (d, 8.5) | 117.7, CH | 7.10 (d, 9.0) | 119.3, CH | 7.10 (d, 9.0) | 119.3, CH | 7.15 (d, 8.0) | 117.7, CH |
| 7 | ― | 159.4, C | ― | 157.9, C | ― | 157.9, C | ― | 158.4, C |
| 8 | ― | 149.8, C | ― | 149.8, C | ― | 149.3, C | ― | 148.7, C |
| 9 | 7.81 (d, 8.5) | 115, CH | 7.82 (d, 8.0) | 115.1, CH | ― | 125.4, C | ― | 124.3, C |
| 10 | 7.46 (t, 8.0) | 124.8, CH | 7.49 (t, 8.0) | 125.2, CH | ― | 131.4, C | ― | 131.9, C |
| 11 | 8.08 (d, 7.5) | 127.5, CH | 8.10 (d, 8.0) | 127.7, CH | 8.10 (s) | 126.3, CH | 8.23 (s) | 127.8, CH |
| 12 | ― | 121.3, C | ― | 121.6, C | ― | 119.1, C | ― | 120.8, C |
| 13 | ― | 139.4, C | ― | 139.2, C | ― | 137.3, C | ― | 136, C |
| 14 | ― | 165.6, C | ― | 165.4, C | ― | 164.5, C | ― | 164.5, C |
| 15 | 4.06 (s) | 52.4, CH3 | 4.06 (s) | 52.5, CH3 | 4.04 (s) | 52.6, CH3 | ― | ― |
| 9-Me | ― | ― | ― | ― | 2.69 (s) | 13.7, CH3 | 2.69 (s) | 13.6, CH3 |
| 13-NH | ― | ― | ― | ― | 11.65 (s, w) | ― | 10.27 (s) | ― |
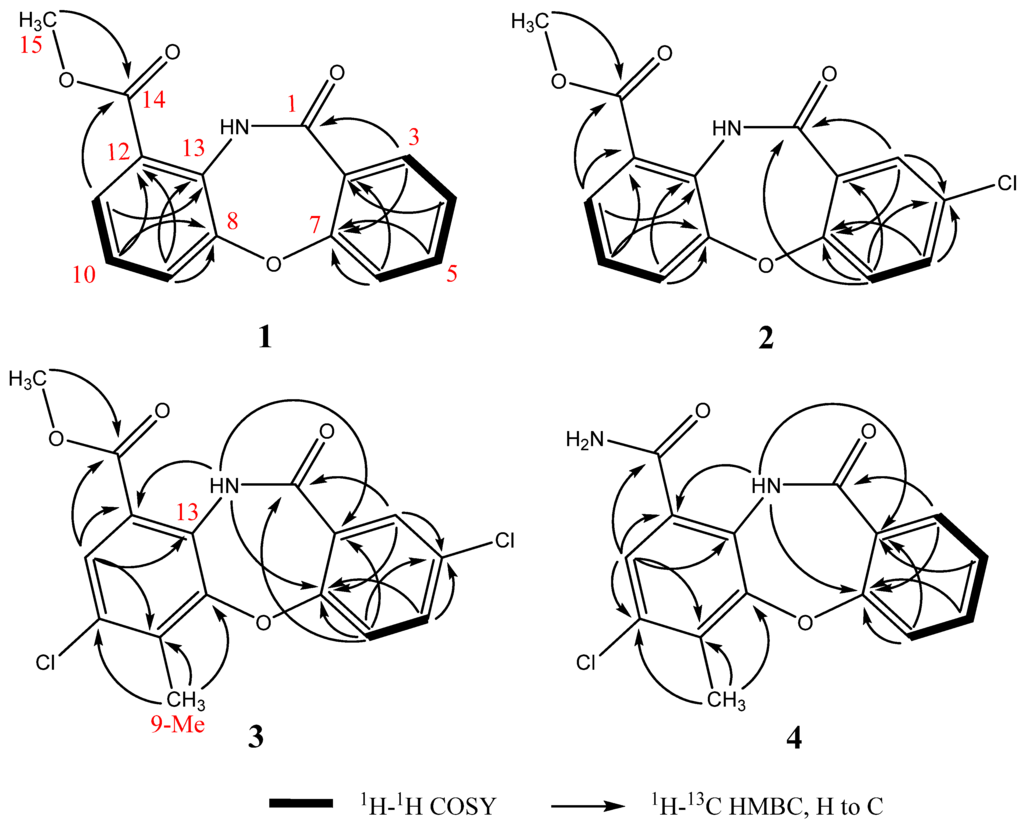
Figure 2.
Key 1H-1H COSY and HMBC correlations of 1–4.
Figure 2.
Key 1H-1H COSY and HMBC correlations of 1–4.
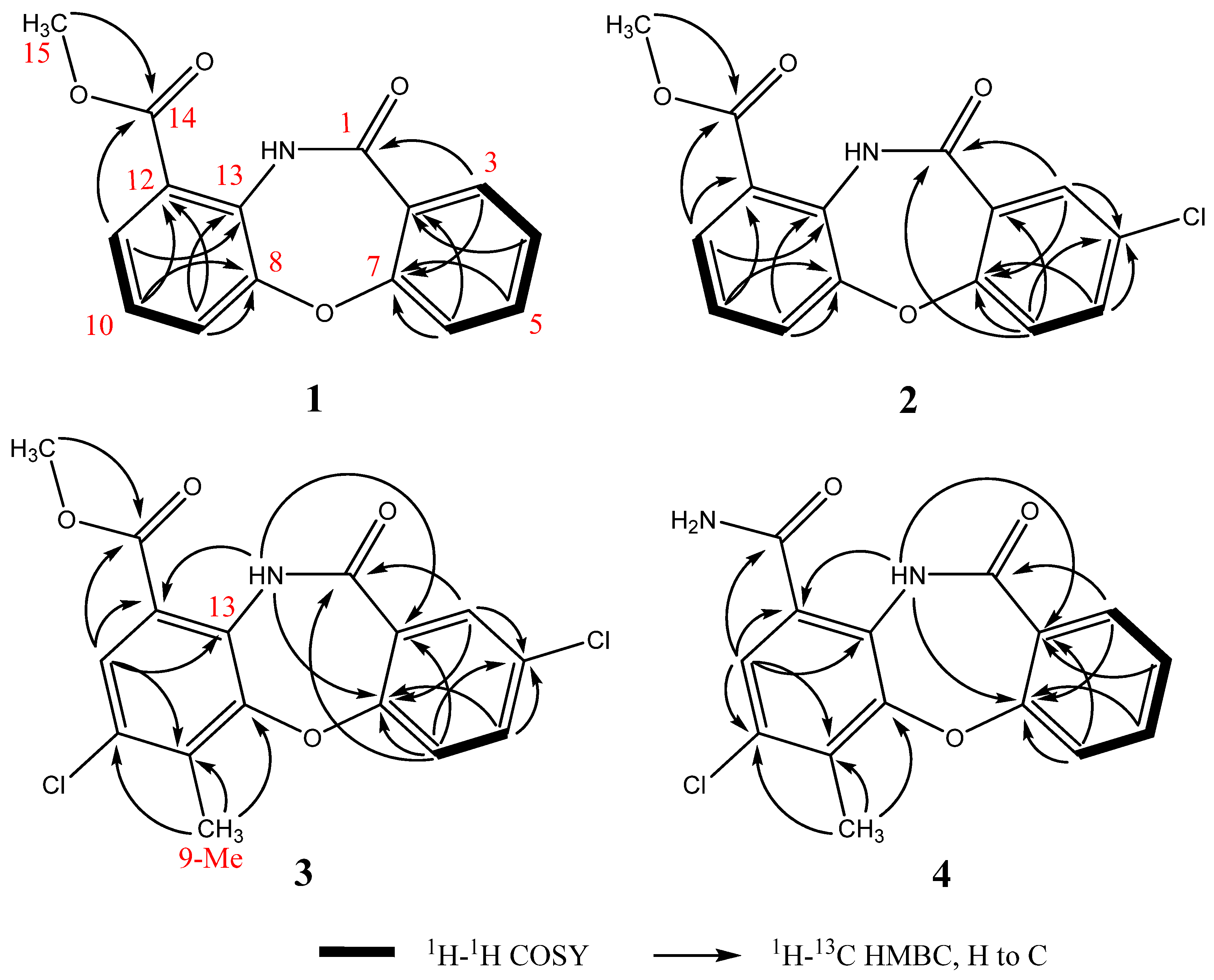
Mycemycins B (2) was obtained as a white solid. Its molecular formula was established as C15H10ClNO4 according to the [M + H]+ peak at m/z 304.0378 (calcd for C15H11ClNO4, 304.0377) in the HR-ESI-MS spectrum combined with its 13C-NMR data, corresponding to 11 degrees of unsaturation. Analyses of its 1H and 13C NMR data (Table 1, Figure S3) revealed similar structural features to those of 1. The most obvious differences between 1 and 2 lay in the positions from C3 to C5. The splitting pattern of H3 changed from “dd” in 1 (coupled with H4 and H5) to “d” in 2 (coupled only with H5). Equally, that of H5 also changed from “dt” in 1 (coupled with H3, H4 and H6) to “dd” in 2 (coupled with H3 and H6). All the changes were due to the lack of a hydrogen atom at the C4 position, which was supported by the DEPT and HSQC correlations (Figure 2 and Figure S3), and by the chemical shift of C4 that changed from δ 119.6 in 1 to δ 124.5 in 2. The exhibited HMBC signals from H6 (strong signal), H3 (moderate signal) and H5 (weak signal) to the quaternary carbon-C4 further confirmed this deduction (Figure 2 and Figure S3). According to the suggested molecular formula, a Cl- atom should be assigned to C4.
Mycemycins C (3) was obtained as a white solid. Its molecular formula was established as C16H11Cl2NO4 according to the [M + H]+ peak at m/z 352.0137 (calcd for C16H12Cl2NO4, 352.0143) in the HR-ESI-MS spectrum combined with its 13C-NMR data, corresponding to 11 degrees of unsaturation. The 1H and 13C NMR data (Table 1, Figure S4) revealed its similar structural features to those of 2, especially that the coupling and chemical shifts of H3-H6, C1-C8 and C13-C15 were in good accordance, which were confirmed by the COSY and HMBC correlations (Figure 2, Figure S4). Major differences lay in the positions from C9 to C12. The NMR data of CH (δH 8.10/δC 126.3, confirmed by HSQC, Figure S4), which showed accordance with C11 of 1 and 2, suggested that this carbon was at position 11, too. However, the strong “s” peak of H11 in the 1H-NMR spectrum indicated the absence of hydrogen atoms at both C10 and C12 compared with 1 and 2. According to the HMBC spectrum, the strong correlation signals from 9-Me to C8, C9 and C10 confirmed the connection of C9 and 9-Me; signals from H11 to C12 (weak signal, Figure S4), C13 and C14, and from NH to C12, combined with the chemical shifts of C12 (δ 119.1) and C13 (δ 137.3), confirmed the connections between C11 and C12, C12 and C14, and between NH and C13. The chemical shift of NH (δ 11.65), which revealed the hydrogen bond between 13-NH and 14-carbonyl oxygen atom, also supported the connection between C12 and C14. A further HMBC signal found from H11 to C9, combined with the correlations described above and the chemical shifts from C8 to C13, revealed the overall perspective of the aromatic system. Finally, according to the suggested molecular formula, the second Cl- atom should be assigned to the idle C10 position.
Mycemycins D (4) was obtained as a white solid. Its molecular formula was established as C15H11ClN2O3 according to the [M + H]+ peak at m/z 303.0538 (calcd for C15H12ClN2O3, 303.0536) in the HR-ESI-MS spectrum combined with its 13C-NMR data, corresponding to 11 degrees of unsaturation. Its structural fragment from C1 to C7 was identical with that of 1, well supported by the similarities in their 1H and 13C NMR data and 2D-NMR correlations (Table 1, Figure 2 and Figure S5). The structural fragment from C8 to C14 was similar to that of 3, sustained by the NMR data, especially by the similar HMBC correlations from positions 8 to 14, albeit that the chemical shift of H11 in 4 (δ 8.23) was slightly different from that in 1–3 (δ 8.08–8.10). Comparisons of the 1H and 13C NMR spectra between 4 and 1–3 revealed the absence of 15-OMe (δH 4.04–4.06/δC 52.4–52.6 in 1–3) in 4 and that, considering the suggested molecular formula, C14 should be connected to an -NH2 group.
Mycemycins E (5) was obtained as a white solid. Its molecular formula was established as C15H10Cl2N2O3 according to the [M + H]+ peak at m/z 337.0149 (calcd for C15H11Cl2N2O3, 337.0146) in the HR-ESI-MS spectrum. This purified compound did not dissolve in any common solvents such as methanol, acetone, acetonitrile, benzene, chloroform, cyclohexane, dichloromethane, dimethylformamide, dimethylsulfoxide, ethyl acetate, formamide, n-hexane, pyridine, tetrahydrofuran and toluene, thus, the NMR analysis could not be applied, and no desirable crystals could be obtained for X-ray analysis either. Therefore, its chemical structure (Figure 1) was suggested mainly according to the highly similar MS/MS pattern with that of 3 (Figure S6) and the fact that co-production of 3–5 is a biogenetic consideration.
2.3. Bioactivities of Compounds 1–4
Mycemycins A–D (1–4) were evaluated for their antimicrobial activity. These compounds turned out to be inactive against microbes Staphyloccocus aureus, Escherichia coli, Bacillus subtilis, Mycobacterium gilvum, Micrococcus luteus, Pseudomonas aeruginosa, Fusarium oxysporum and Trichoderma viride. The bioactivity of compounds 1–4 were further predicted using Reverse Docking Method [16], and the results indicated potential anti-HIV-1 reverse transcriptase (RT) activity of the compounds. However, because mycemycins had very poor solubility in the HIV-1 RT enzyme reaction buffer and precipitated out before the enzyme was added, their HIV-1 RT inhibitory activity could not be properly measured.
2.4. Discussion
Mycemycins are structurally classified as dibenzoxazepinones with two aromatic ring systems. Dibenzoxazepinones have been reported to have various bioactivities and be potential antioxidants [17], calcium channel antagonists [18,19], HIV-1 RT inhibitors [20], etc. Most of these compounds are artificially designed and synthesized, with only two natural compounds derived from leaves of herbaceous plant Carex distachya [17]. This is the first report of natural dibenzoxazepinones discovered from the microbial kingdom. Interestingly, mycemycins A and B were derived from a terrestrial streptomycete recovered from red soil in China and mycemycins C–E from a marine streptomycete isolated from deep seawater in south Indian Ocean, and these two strains belong to different Streptomyces species with a 96.5% 16S rRNA gene sequence similarity. These observations indicate a wide distribution of mycemycins. Moreover, all the marine-derived mycemycins are mono/bis-chlorinated, underpinning that the ocean is a promising source for the discovery of halogenated natural products [21].
GntR-like regulators are often located adjacent to the genes that they control, but there are many exceptions as well [11]. Although the regulatory gene orf-1741 is flanked by putative secondary metabolic genes, it is impossible in this study to predict the corresponding metabolites that the regulator may involve based on the genome survey sequences. As the halogenase genes (orf-1802 and orf-1805) in FXJ8.012 genome are located relatively far from orf-1741 (around 50 kb), our results suggest that the GntR encoded by orf-1741 is likely to be an orphan regulator [11] or an upstream repressor of mycemycin biosynthesis in S. olivaceus FXJ8.012. Further efforts need to be made to verify this inference. More experiments are also required to enhance the solubility of mycemycins, especially compound 5, to better evaluate their bioactivities.
3. Experimental Section
3.1. General
UV spectra were recorded on a Beckman Coulter DU800 UV/Vis Spectrophotometer, 200 to 400 nm. IR spectra were recorded on a Thermo Nicoiet 8700 spectrometer. HR-ESI-MS spectra were obtained on a Waters Xevo G2 QTOF mass spectrometer with an ACQUITY UPLC BEH C18 column (2.1 mm × 50 mm, 1.7 µm). NMR spectra were measured on a Bruker AV 500 NMR spectrometer. The concentration was performed on an EYELA N-1100S-W rotary evaporator. The analytical and semi-preparative HPLC were both performed on a Shimadzu SPD-M20A HPLC system with a Waters Xbridge ODS column (4.5 × 150 mm, 5 μm, for analysis) and a Sigma-aldrich Ascentis RP-Amide column (4.6 × 150 mm, 5 μm, for semi-preparation). Column chromatography was carried out on silica gel (100–200 mesh) (Qingdao Haiyang Chemical Group Corp., Qingdao, China) and Sephadex LH-20 (Pharmacia).
3.2. Strains, Plasmids and Media
Bacterial strains, plasmids and primers used in this study are listed in Tables S1,2 in the supplemental material.
S. sp. FXJ1.235 was isolated from an acidic red soil sample collected from Boyang lake, Jiangxi Province, China. The isolation medium was M6 agar and the growth medium was M11 agar [22]. The strain was identified by morphology and 16S rRNA gene sequence analysis using regular procedures [23]. Its 16S rRNA gene sequence (GenBank accession number KJ152035) showed the highest similarity (99%) to Streptomyces galbus DSM 40089T (GenBank accession number NR_026178). Detailed information of S. olivaceus FXJ8.012 (16S rRNA gene sequence HQ622478) was reported previously [9]. Escherichia coli Top 10 was used as a host strain for general cloning, and E. coli ET12567/pUZ8002 was used for conjugation between E. coli and streptomycetes.
GYM agar medium [9] was used for sporulation of strains FXJ1.235 and FXJ8.012Δ1741; GYM liquid medium was used for seed culture of strains FXJ1.235 and FXJ8.012Δ1741; GYM liquid medium with 1 g/L NaCl was used as the fermentation medium for FXJ1.235; modified R2 medium [15] was used as the fermentation medium for FXJ8.012Δ1741. LB and PDA agars were used for antibacterial and antifungal tests, respectively, as previously described [9]. E. coli strains carrying plasmids for nucleic acid manipulation were cultured in liquid or agar LB medium, added with ampicillin (100 μg/mL), nalidixic acid (25 μg/mL), apramycin (50 μg/mL), chloramphenicol (25 μg/mL) or kanamycin (100 μg/mL) as appropriate.
3.3. Nucleic Acid Manipulations and Mutant Construction
The DNA sequence of orf-1741 has been deposited in the GenBank database with an accession number KT357292.
Nucleic acid manipulations for E. coli and streptomycetes were performed according to standard approaches [24,25]. Inactivation of orf-1741 was performed by gene replacement via double-crossover homologous recombination. The kanamycin resistance gene neo was obtained by amplification from a recombinant plasmid pUC119::neo with primer pairs neo-EcoRI-Forward and neo-EcoRI-Reverse (Table S2) [26]. Upstream and downstream regions of approximately 2 kb flanking orf-1741 were amplified by PCR with primer pairs 1741-L-Forward/1741-L-Reverse and 1741-R-Forward/1741-R-Reverse (Table S2). These PCR products were digested with HindIII/EcoRI or XbaI/EcoRI, then separated by agarose gel electrophoresis and purified from the gel. The resulting fragments were ligated with the EcoRI-digested neo gene and XbaI- and HindIII-digested plasmid pKC1139 to generate pKC1139::1741::neo, which was subsequently introduced into S. olivaceus FXJ8.012 by conjugation via ET12567/pUZ8002. Spores of exconjugants were harvested and spread on MS agar [25] containing kanamycin. After incubation at 38 °C for a week, apramycin-sensitive and kanamycin-resistant colonies were identified, and further verified as orf-1741 disruption mutants (Δ1741) by PCR with primer pairs Δ1741-Foward and Δ1741-Reverse (Figure S7) and sequencing of the resulting amplimers.
3.4. Fermentation, Extraction and Isolation
3.4.1. S. sp. FXJ1.235
After growth on GYM agar in petri dishes at 28 °C for 7 days, a suitable amount of spores of S. sp. FXJ1.235 were transferred into a 500-mL shake flask containing 100 mL of GYM liquid medium. The resulting culture was incubated on a rotary shaker at 170–180 rpm and 28 °C for 5 days to afford a seed culture, which was used to inoculate 100 shake flasks (500 mL) each containing 100 mL of GYM liquid medium supplemented with NaCl. The inoculated flasks were incubated under similar conditions for 7 days.
After fermentation, the culture broth (10 L) was centrifuged at 5000 rpm for 15 min to separate mycelia and supernatant. The resulting mycelium cake was extracted three times with an equal volume of ethanol for 8 h. The ethanol extracts were combined and concentrated in vacuo, and then subjected to column chromatography on silica gel (100–200 mesh) using acetone as the elution solvent to obtain a mixture of crude products (1.1–1.2 g). The mixture was passed repeatedly through a Sephadex LH-20 column eluting with MeOH/CHCl3 (4:1) to remove fatty acids, nucleosides and other impurities. Final purification was carried out by RP-HPLC using a Sigma-aldrich Ascentis RP-Amide column eluting with acetonitrile/H2O (7:3) to yield 1 (7.1 mg) and 2 (5.8 mg).
3.4.2. S. olivaceus FXJ8.012
The inoculation and fermentation process for FXJ8.012Δ1741 was almost the same as that for FXJ1.235. Differences lay in the large scale fermentation where 200 shake flasks (500 mL) each containing 100 mL of modified R2 medium [15] were used for FXJ8.012Δ1741.
The separation and purification procedures for the metabolites of FXJ8.012Δ1741 were almost identical with those for 1 and 2. About 5.1 g crude products were obtained after silica gel column chromatography; and in the final RP-HPLC procedure, the mixture of compounds was eluted with acetonitrile/H2O (76:24) to yield 3 (1.3 mg), 4 (0.8 mg) and 5 (about 5–6 mg).
Mycemycin A (1): white solid; UV (MeOH) λmax 244, 262, 272(sh), 292(sh), 305, 332, 345(sh) nm; IR (KBr) νmax 2954, 2917, 1716, 1634, 1599, 1549, 1488, 1426, 1305, 1259, 1244, 1192, 1160, 1140, 1061, 1013, 939, 876, 787, 753, 709, 537, 493, 480 cm−1; 1H- and 13C-NMR data, Table 1; HR-ESI-MS m/z 270.0776 [M + H]+ (calcd for C15H12NO4, 270.0766).
Mycemycin B (2): white solid; UV (MeOH) λmax 244, 265, 275(sh), 295, 307, 340, 351(sh) nm; IR (KBr) νmax 2954, 2921, 1716, 1631, 1584, 1541, 1479, 1425, 1281, 1242, 1194, 1134, 1060, 1022, 946, 879, 822, 788, 759, 724, 547, 487 cm−1; 1H- and 13C-NMR data, Table 1; HR-ESI-MS m/z 304.0378 [M + H]+ (calcd for C15H11ClNO4, 304.0377).
Mycemycin C (3): white solid; UV (MeOH) λmax 251, 259(sh), 269(sh), 279(sh), 295(sh), 307(sh), 333, 345(sh) nm; 1H- and 13C-NMR data, Table 1; HR-ESI-MS m/z 352.0137 [M + H]+ (calcd for C16H12Cl2NO4, 352.0143).
Mycemycin D (4): white solid; UV (MeOH) λmax 244(sh), 250, 260(sh), 273(sh), 285(sh), 297(sh), 312, 344, 356(sh) nm; 1H- and 13C-NMR data, Table 1; HR-ESI-MS m/z 303.0538 [M + H]+ (calcd for C15H12ClN2O3, 303.0536).
Mycemycin E (5): white solid; UV (MeOH) λmax 244(sh), 251, 260(sh), 270(sh), 283(sh), 300(sh), 311, 340, 352(sh) nm; HR-ESI-MS m/z 337.0148 [M + H]+ (calcd for C15H11Cl2N2O3, 337.0146).
3.5. Bioactivity Assay
Primary antimicrobial assays were conducted using the agar-diffusion method. 50 μg of compound were transferred to a sterile paper disc of 5 mm diameter, which was put onto the surface of LB or PDA agar containing pathogenic bacteria or fungi, followed by incubation at 37 °C overnight for bacteria or 25 °C for 1–2 days for fungi. Bioactivity was determined by observing the inhibition zones.
The assay of HIV-1 RT inhibitory activity was carried out in triplicate using the Reverse Transcriptase Assay, colorimetric kit (Roche 11468120910), following the standard procedure provided by the manufacture. The initial concentration for compounds 1–4 was set to 1000 μg/mL, and that for the positive control EFV to 1000 ng/mL.
4. Conclusions
In this study, we applied genetic manipulation to a marine-derived streptomycete, leading to the discovery of three new halogenated compounds, mycemycins C–E (3–5), which were structurally similar to another two new compounds, mycemycins A and B (1 and 2) isolated from a soil-derived streptomycete. These new compounds were elucidated as dibenzoxazepinone derivatives, and are the first dibenzoxazepinones discovered from microbial source. These findings suggest that marine-derived actinomycetes have great potential to produce novel natural products, notably halogenated ones, as their terrestrial counterparts, and new approaches such as manipulation of regulators are effective at activating the biosynthesis of these cryptic marine compounds.
Supplementary Files
Supplementary File 1Acknowledgments
This research was supported by the program of China Ocean Mineral Resources R & D Association (grant No. DYXM-115-02-1-02), the National Basic Research Program of China (grant No. 2011CB808800), and by the National Natural Science Foundation of China (grant No. 31170010 and 31300010). We thank Huarong Tan (Institute of Microbiology, Chinese Academy of Sciences, Beijing, China) for providing E. coli ET12567/pUZ8002, pKC1139 and pUC119, and Yongtang Zheng and Liumeng Yang (Kunming Institute of Zoology, Chinese Academy of Sciences, Yunnan, China) for anti-HIV-1 RT assay.
Author Contributions
Ning Liu and Ying Huang conceived and designed the experiments; Ning Liu and Fangying Song performed the experiments; Ning Liu and Fei Shang analyzed the data; Ning Liu, Fangying Song and Ying Huang wrote the paper.
Conflicts of Interest
The authors declare no conflict of interest.
References and Notes
- Baltz, R.H. Antimicrobials from actinomycetes: Back to the future. Microbe 2007, 2, 125–131. [Google Scholar]
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Bérdy, J. Thoughts and facts about antibiotics: Where we are now and where we are heading. J. Antibiot. 2012, 65, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Gontang, E.A.; Gaudencio, S.P.; Fenical, W.; Jensen, P.R. Sequence-based analysis of secondary-metabolite biosynthesis in marine actinobacteria. Appl. Environ. Microbiol. 2010, 76, 2487–2499. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; Abdel-Mageed, W.M.; Ebel, R.; Bull, A.T.; Goodfellow, M.; Fiedler, H.P.; Jaspars, M. Dermacozines H–J isolated from a deep-sea strain of Dermacoccus abyssi from Mariana Trench sediments. J. Nat. Prod. 2014, 77, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Ochi, K.; Hosaka, T. New strategies for drug discovery: Activation of silent or weakly expressed microbial gene clusters. Appl. Microbiol. Biotechnol. 2013, 97, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Rebets, Y.; Brotz, E.; Tokovenko, B.; Luzhetskyy, A. Actinomycetes biosynthetic potential: How to bridge in silico and in vivo? J. Ind. Microbiol. Biotechnol. 2014, 41, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chater, K.F.; Chandra, G.; Niu, G.; Tan, H. Molecular regulation of antibiotic biosynthesis in Streptomyces. Microbiol. Mol. Biol. Rev. 2013, 77, 112–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Shang, F.; Xi, L.; Huang, Y. Tetroazolemycins A and B, two new oxazole-thiazole siderophores from deep-sea Streptomyces olivaceus FXJ8.012. Mar. Drugs 2013, 11, 1524–1533. [Google Scholar] [CrossRef]
- Hillerich, B.; Westpheling, J. A new GntR family transcriptional regulator in Streptomyces coelicolor is required for morphogenesis and antibiotic production and controls transcription of an ABC transporter in response to carbon source. J. Bacteriol. 2006, 188, 7477–7487. [Google Scholar] [CrossRef] [PubMed]
- Hoskisson, P.A.; Rigali, S. Chapter 1: Variation in form and function the helix-turn-helix regulators of the GntR superfamily. Adv. Appl. Microbiol. 2009, 69, 1–22. [Google Scholar] [PubMed]
- Zhu, Q.; Li, J.; Ma, J.; Luo, M.; Wang, B.; Huang, H.; Tian, X.; Li, W.; Zhang, S.; Zhang, C.; et al. Discovery and engineered overproduction of antimicrobial nucleoside antibiotic A201A from the deep-sea marine actinomycete Marinactinospora thermotolerans SCSIO 00652. Antimicrob. Agents. Chemother. 2012, 56, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Horbal, L.; Fedorenko, V.; Bechthold, A.; Luzhetskyy, A. A transposon-based strategy to identify the regulatory gene network responsible for landomycin E biosynthesis. FEMS. Microbiol. Lett. 2013, 342, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Smanski, M.J.; Peterson, R.M.; Marchillo, K.; Andes, D.; Rajski, S.R.; Shen, B. Engineering of Streptomyces platensis MA7339 for overproduction of platencin and congeners. Org. Lett. 2010, 12, 1744–1747. [Google Scholar] [CrossRef] [PubMed]
- Modified R2 medium: sucrose (103 g), MgCl2·6H2O (10.12 g), difco casaminoacids (0.1 g), TES buffer (5.73 g), trace element solution (2 mL, containing ZnCl2 40 mg/L, FeCl3·6H2O 200 mg/L, CuCl2·2H2O 10 mg/L, MnCl2·4H2O 10 mg/L, Na2B4O7·10H2O 10 mg/L), distilled water 1000 mL, and added with KH2PO4 (0.5%) 10 mL, CaCl2·2H2O (5 M) 4 mL, NaOH (1 N) 5 mL, glucose (50%) 20 mL and l-proline (20%) 15 ml before use.
- Liu, X.; Ouyang, S.; Yu, B.; Liu, Y.; Huang, K.; Gong, J.; Zheng, S.; Li, Z.; Li, H.; Jiang, H. PharmMapper server: A web server for potential drug target identification using pharmacophore mapping approach. Nucleic Acids Res. 2010, 38, W609–W614. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, A.; D’Abrosca, B.; Pacifico, S.; Cefarelli, G.; Uzzo, P.; Monaco, P. Natural dibenzoxazepinones from leaves of Carex distachya: Structural elucidation and radical scavenging activity. Bioorg. Med. Chem. Lett. 2007, 17, 636–639. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Farmer, P.S.; Quillam, M.A.; Howlett, S.E. Dibenzothiazepinones as potential calcium channel antagonists. I. Drug. Des. Discov. 1993, 10, 331–342. [Google Scholar] [PubMed]
- Li, R.; Farmer, P.S.; Wang, J.; Boyd, R.J.; Cameron, T.S.; Quilliam, M.A.; Walter, J.A.; Howlett, S.E. Molecular geometries of dibenzothiazepinone and dibenzoxazepinone calcium antagonists. Drug. Des. Discov. 1995, 12, 337–358. [Google Scholar] [PubMed]
- Hargrave, K.D.; Schmidt, G.; Engel, W.; Schromm, K.; Schromm, W.E.K.; Hargrave, K.; Schmidit, G.; Nehai, S. Prevention and treatment of HIV infection-by administering di:benz-(B,F)-(1–4)-oxazepine (and thiazepin)-11(hydroxy)-one derivs. and salts. EP419861-B; EP419861-A2; EP419861-A; AU9061916-A; CA2024040-A; JP3163021-A; HU57589-T; ZA9006834-A; EP419861-A3; AU639255-B; EP419861-B1; DE69023311-E; US5571806-A; NZ235082-A; HU211077-B; JP2862980-B2; KR165108-B1; CA2024040-C; EP419861-A2; EP419861-A. 03 Apr 1991 199114 English AU9061916-A 07 Mar 1991 199117 JP3163021-A 15 Jul 1991 199134 ZA9006834-A 27 May 1992 C07K 199228.
- Gribble, G.W. Biological activity of recently discovered halogenated marine natural products. Mar. Drugs. 2015, 13, 4044–4136. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, N.; Li, X.; Ding, Y.; Shang, F.; Gao, Y.; Ruan, J.; Huang, Y. Red soils harbor diverse culturable actinomycetes that are promising sources of novel secondary metabolites. Appl. Environ. Microbiol. 2015, 81, 3086–3103. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, H.; Liu, M.; Gu, Q.; Zheng, W.; Huang, Y. Streptomyces alni sp. nov., a daidzein-producing endophyte isolated from a root of Alnus nepalensis D. Don. Int. J. Syst. Evol. Microbiol. 2009, 59, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; The John Innes Foundation: Norwich, UK, 2000. [Google Scholar]
- Pan, Y.; Liu, G.; Yang, H.; Tian, Y.; Tan, H. The pleiotropic regulator AdpA-L directly controls the pathway-specific activator of nikkomycin biosynthesis in Streptomyces ansochromogenes. Mol. Microbiol. 2009, 72, 710–723. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).