CS5931, a Novel Polypeptide in Ciona savignyi, Represses Angiogenesis via Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs)

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
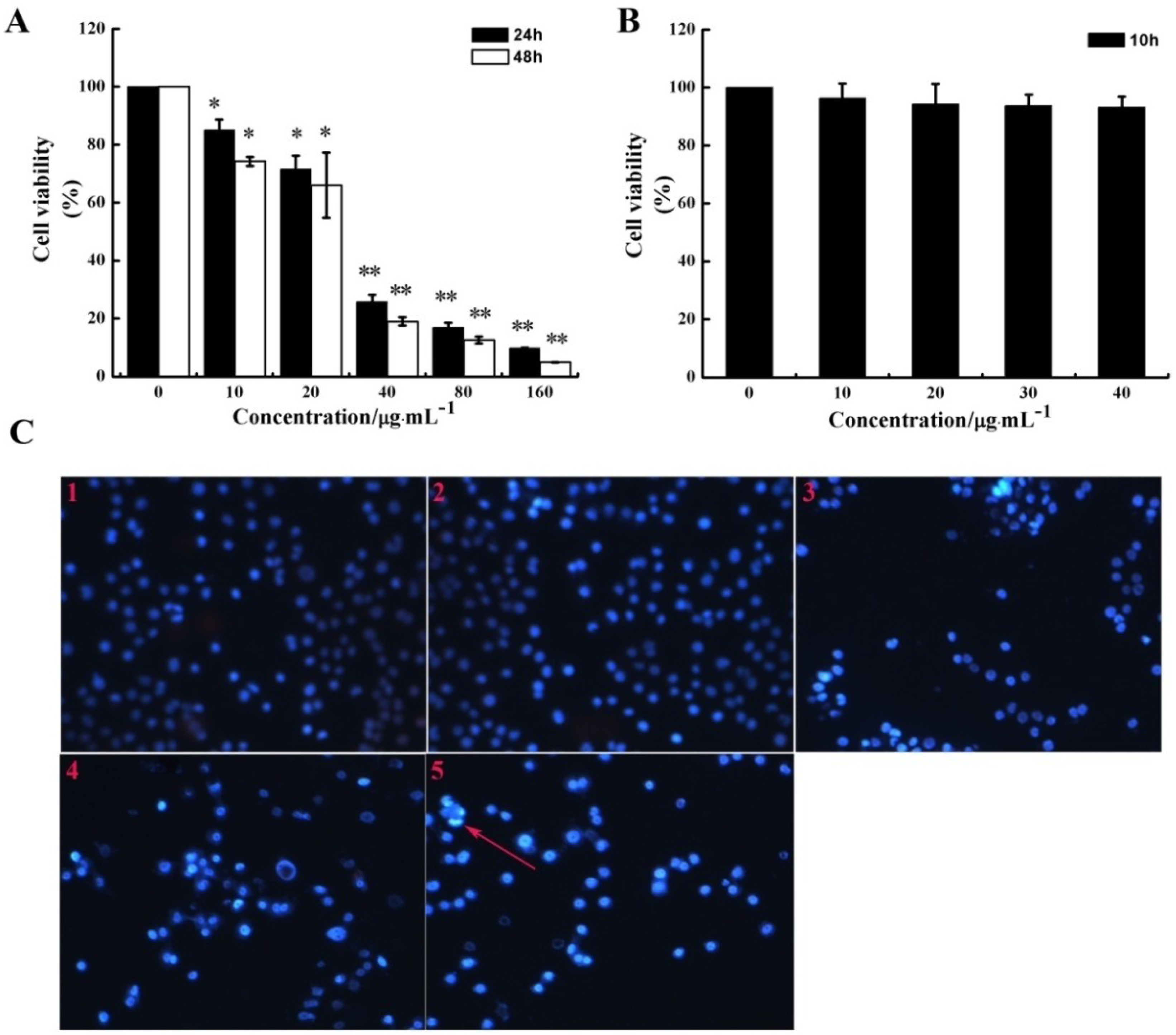
2.1. CS5931 Inhibits the Proliferation of HUVECs
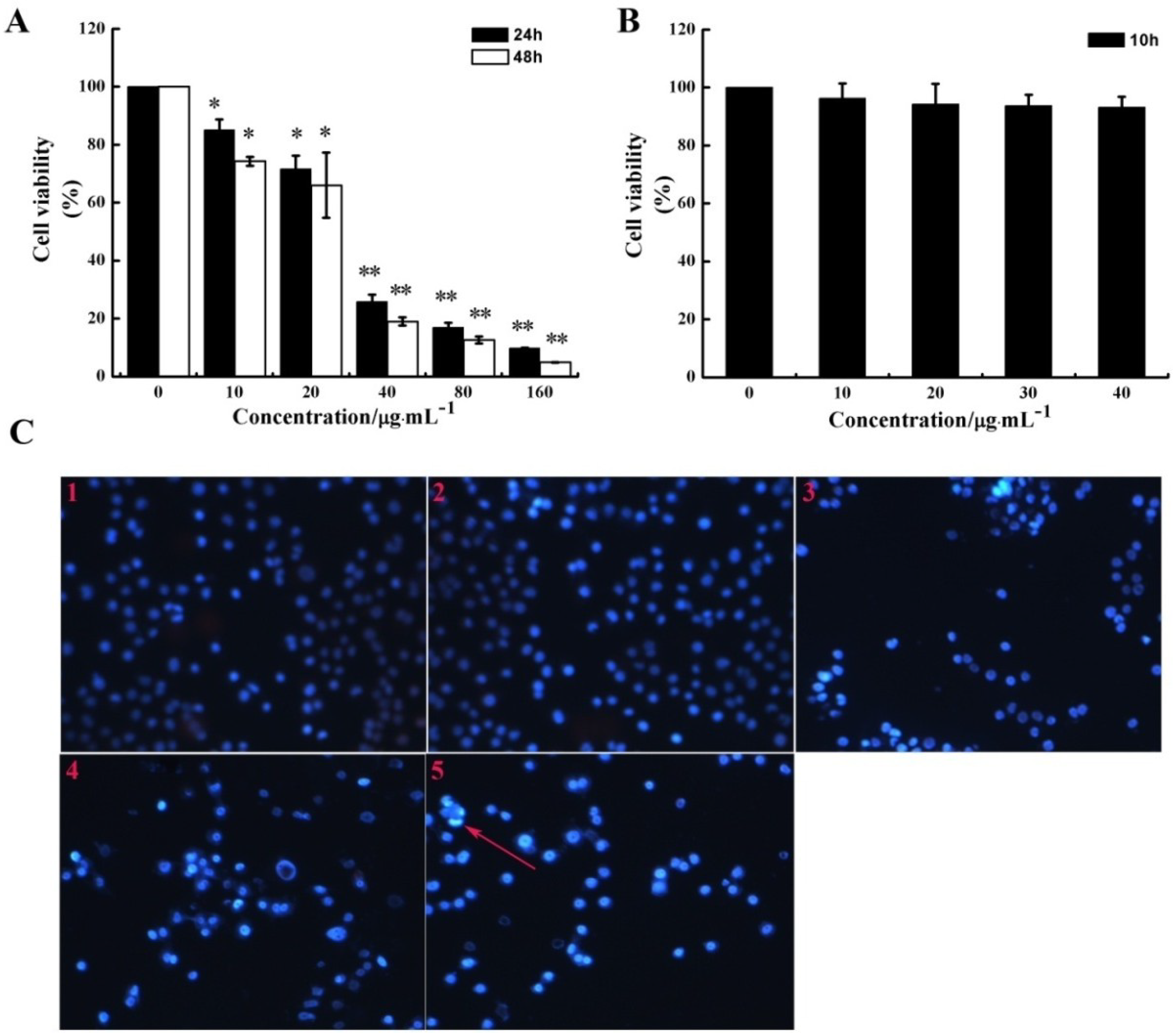
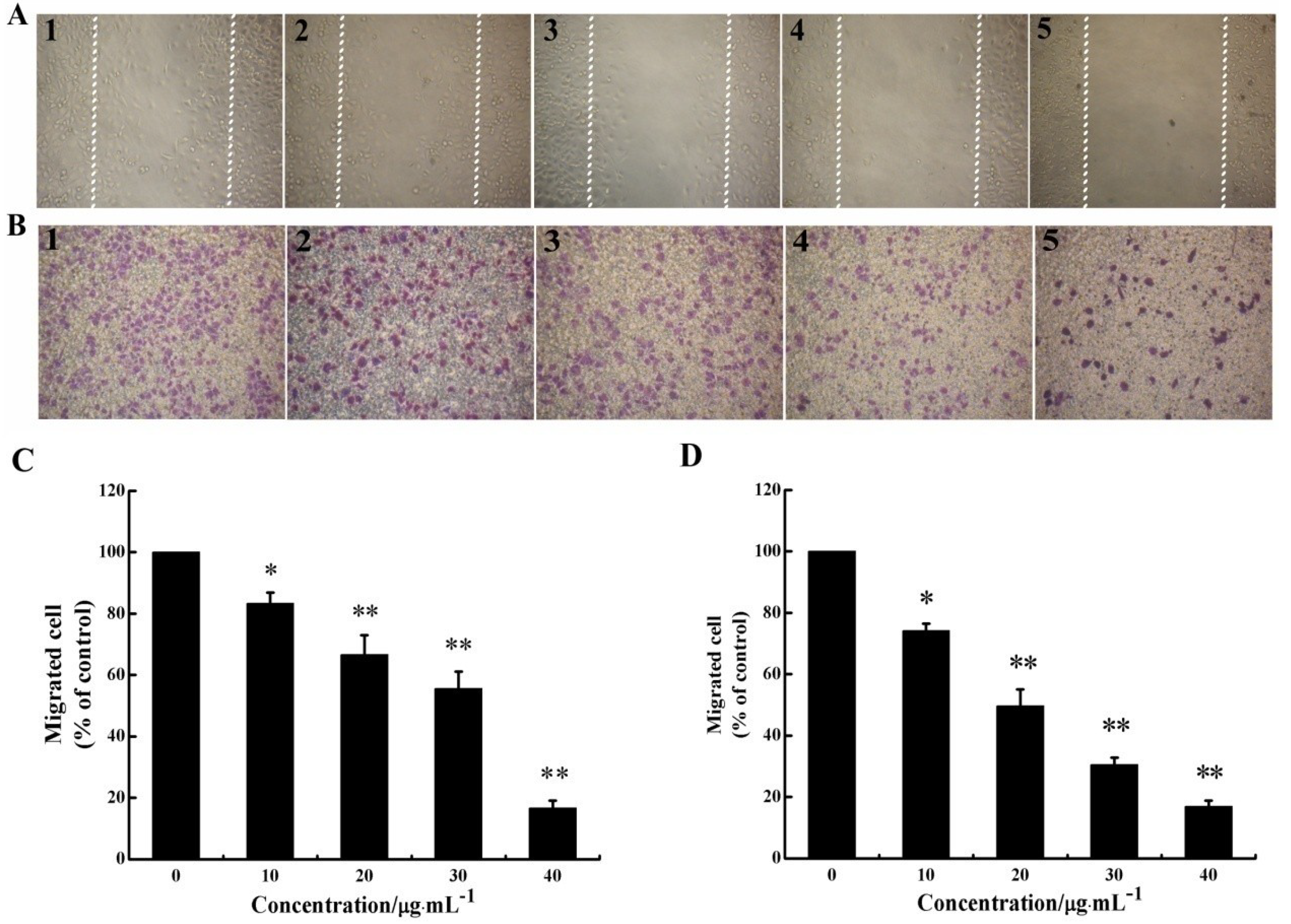
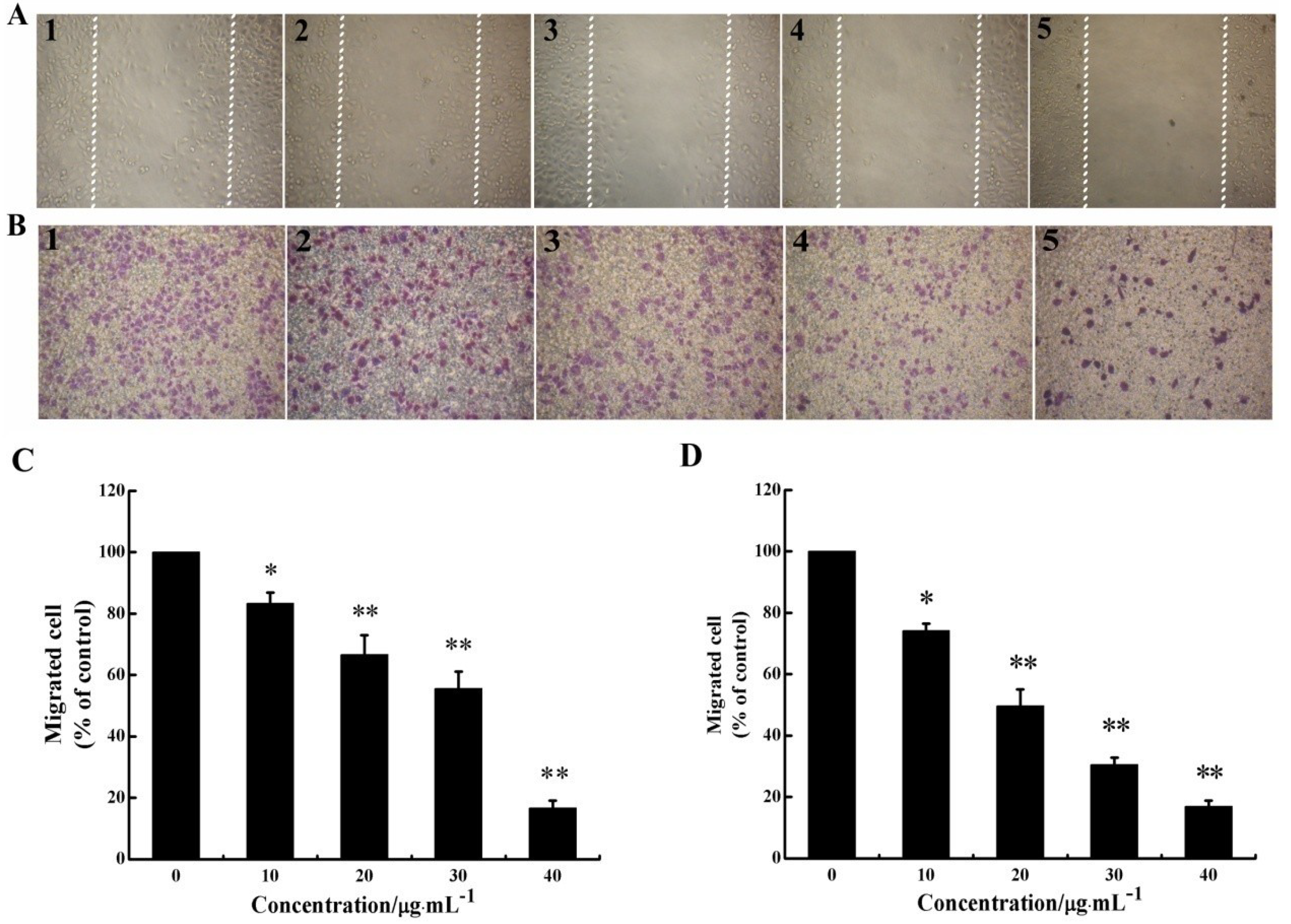
2.2. CS5931 Represses the Migration of HUVECs
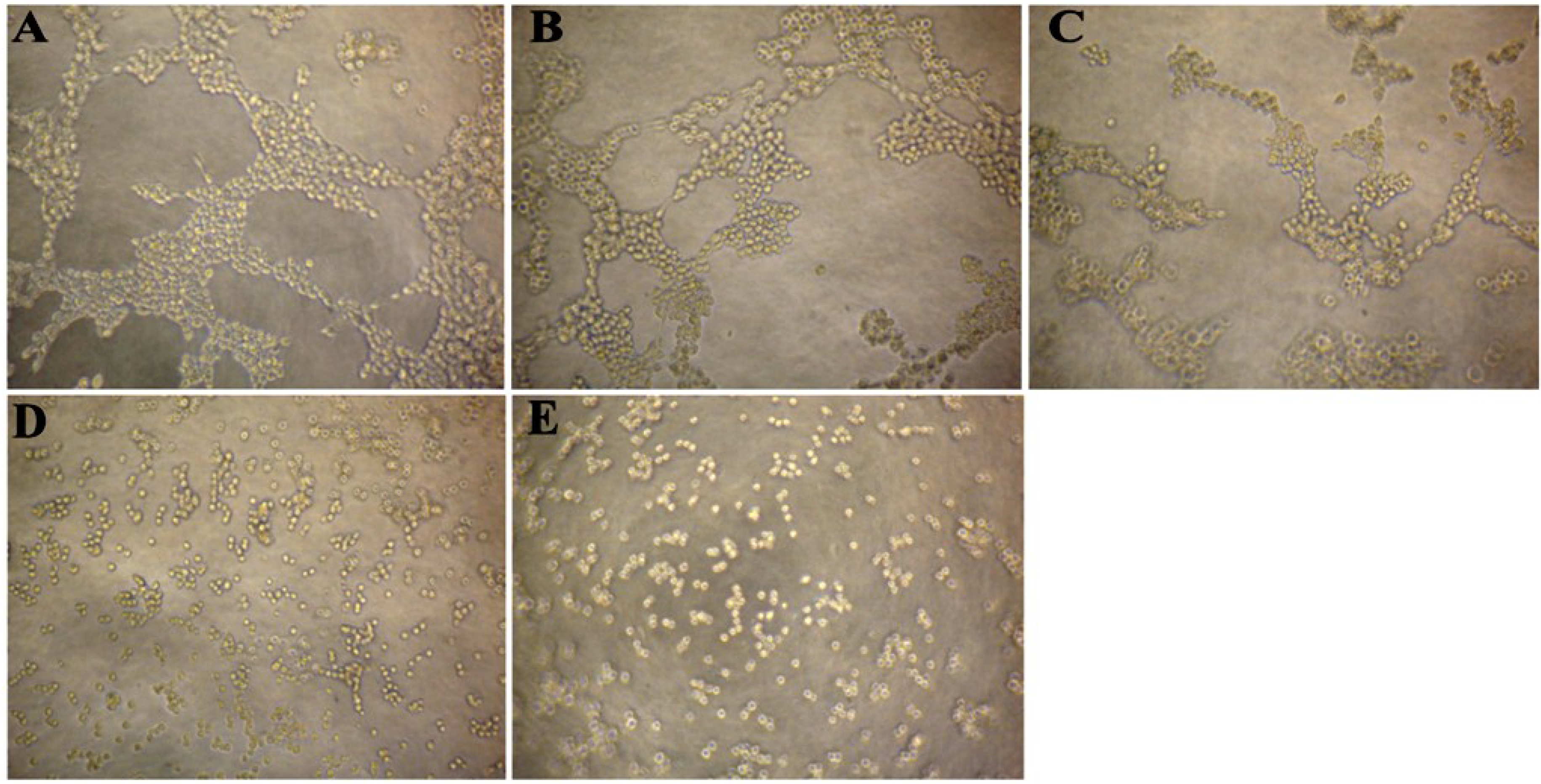
2.3. CS5931 Disrupts the Cord Formation of HUVECs
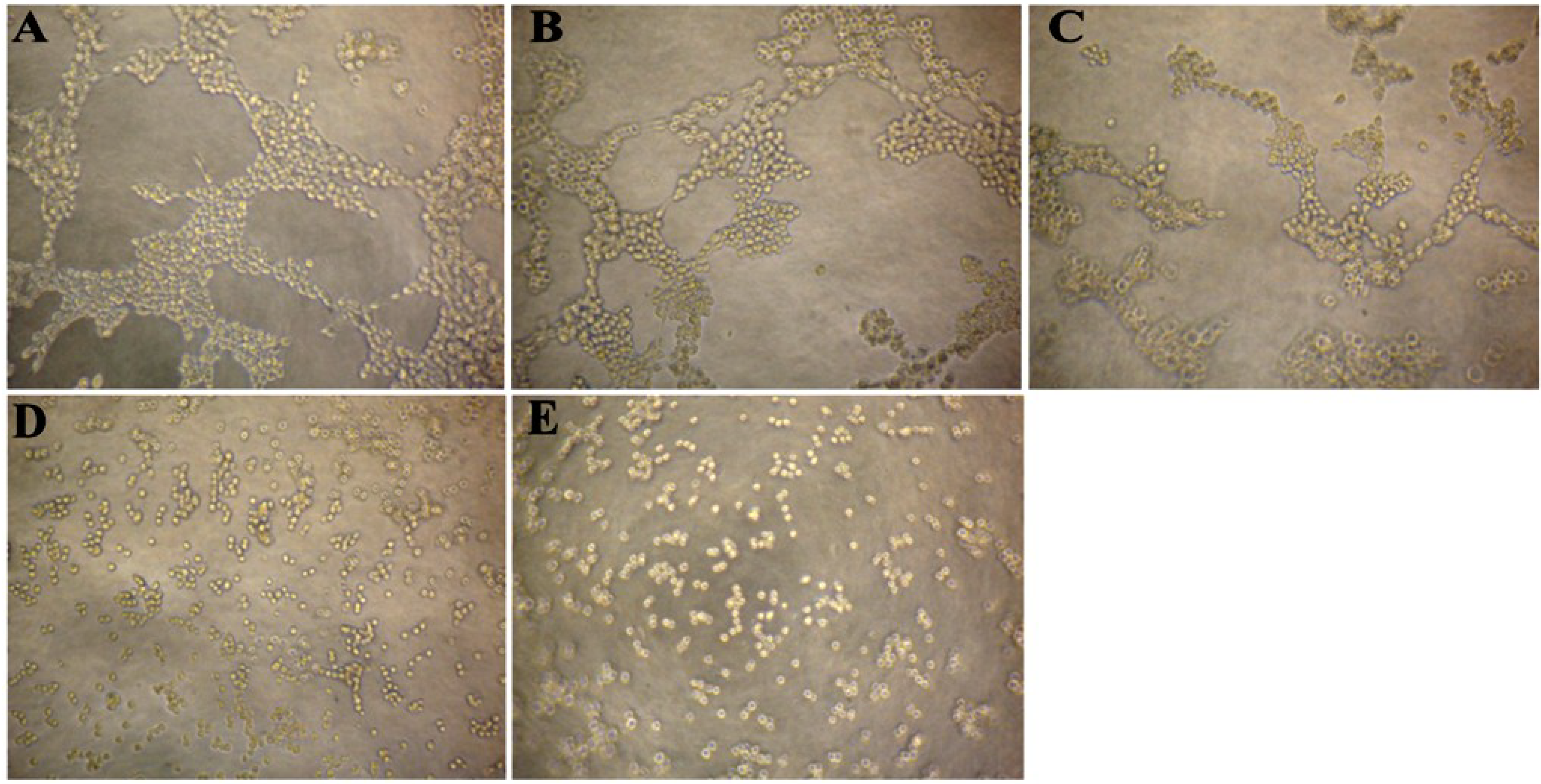
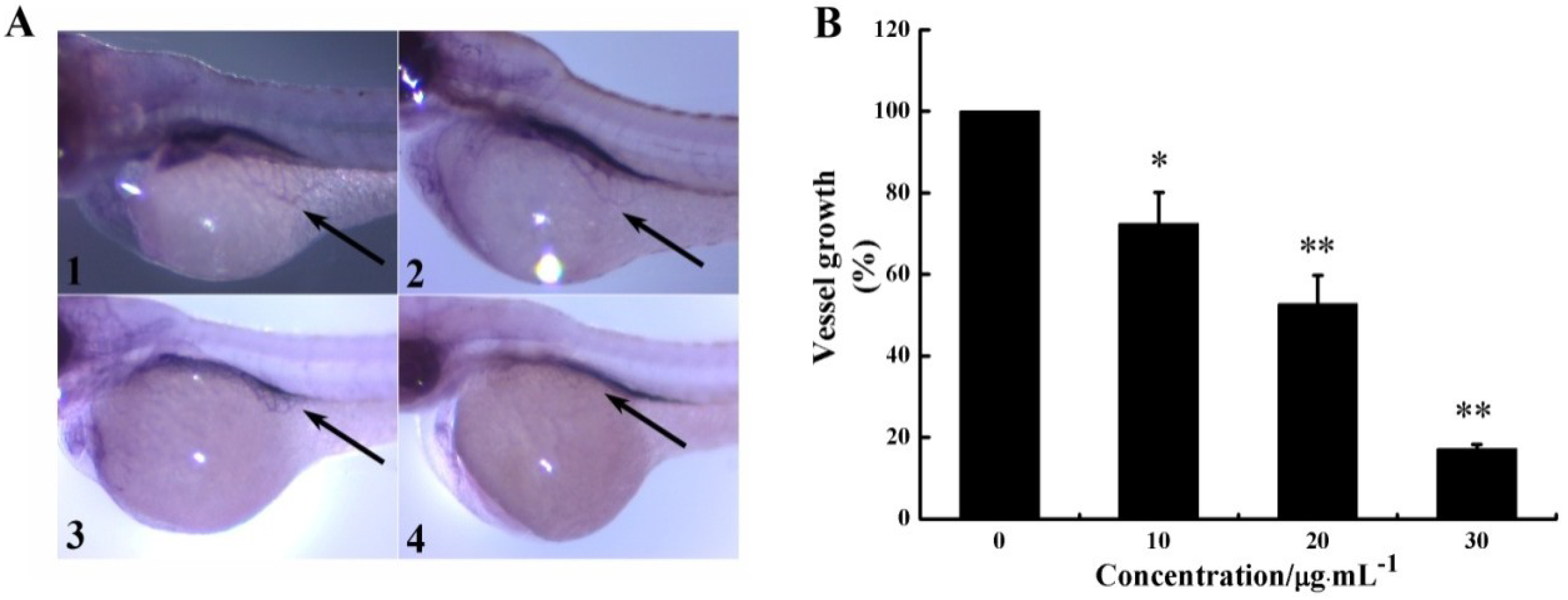
2.4. CS5931 Blocks Vessel Formation in Zebrafish Embryos in Vivo

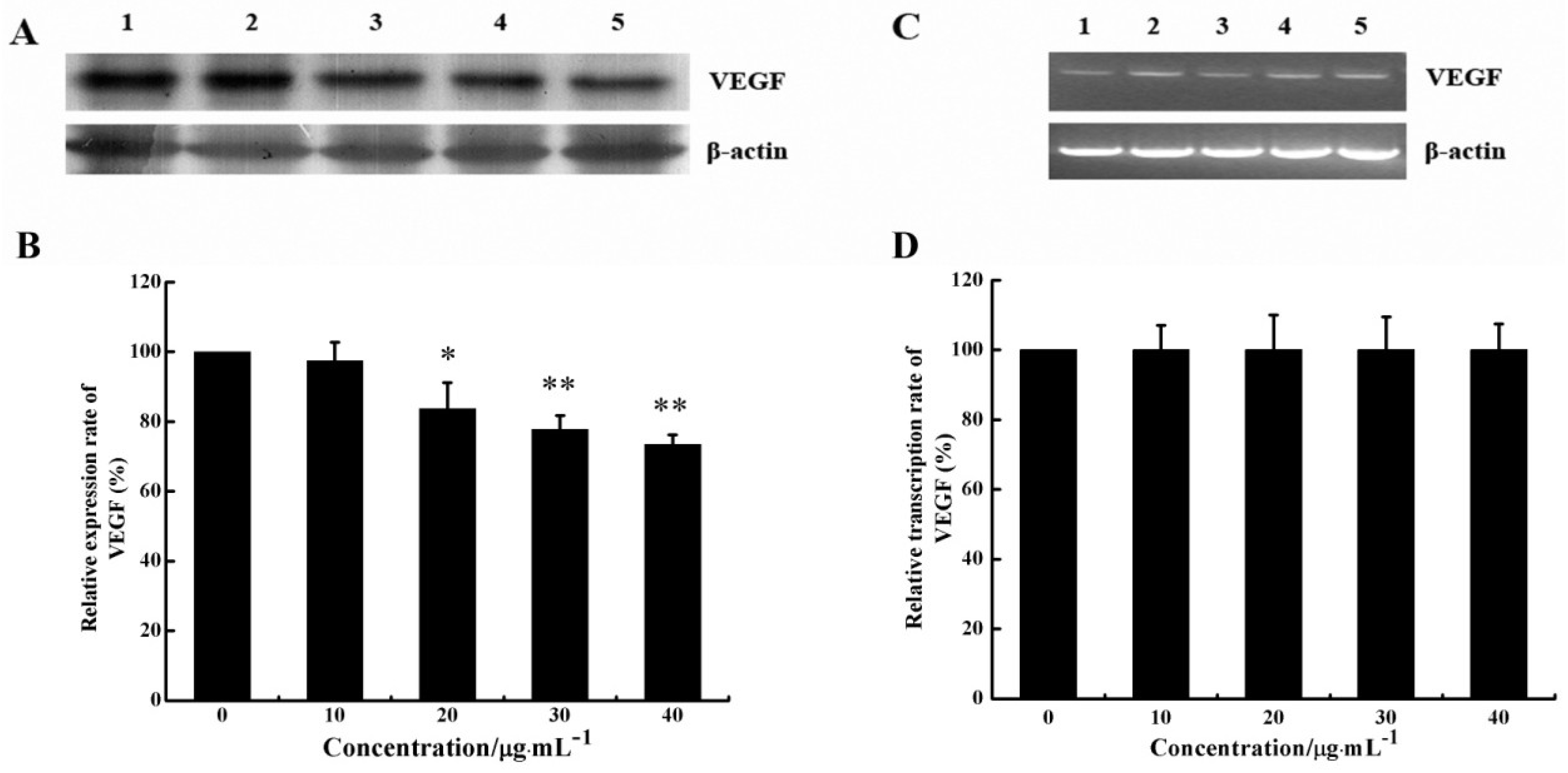
2.5. CS5931 Reduces VEGF Expression in HUVECs

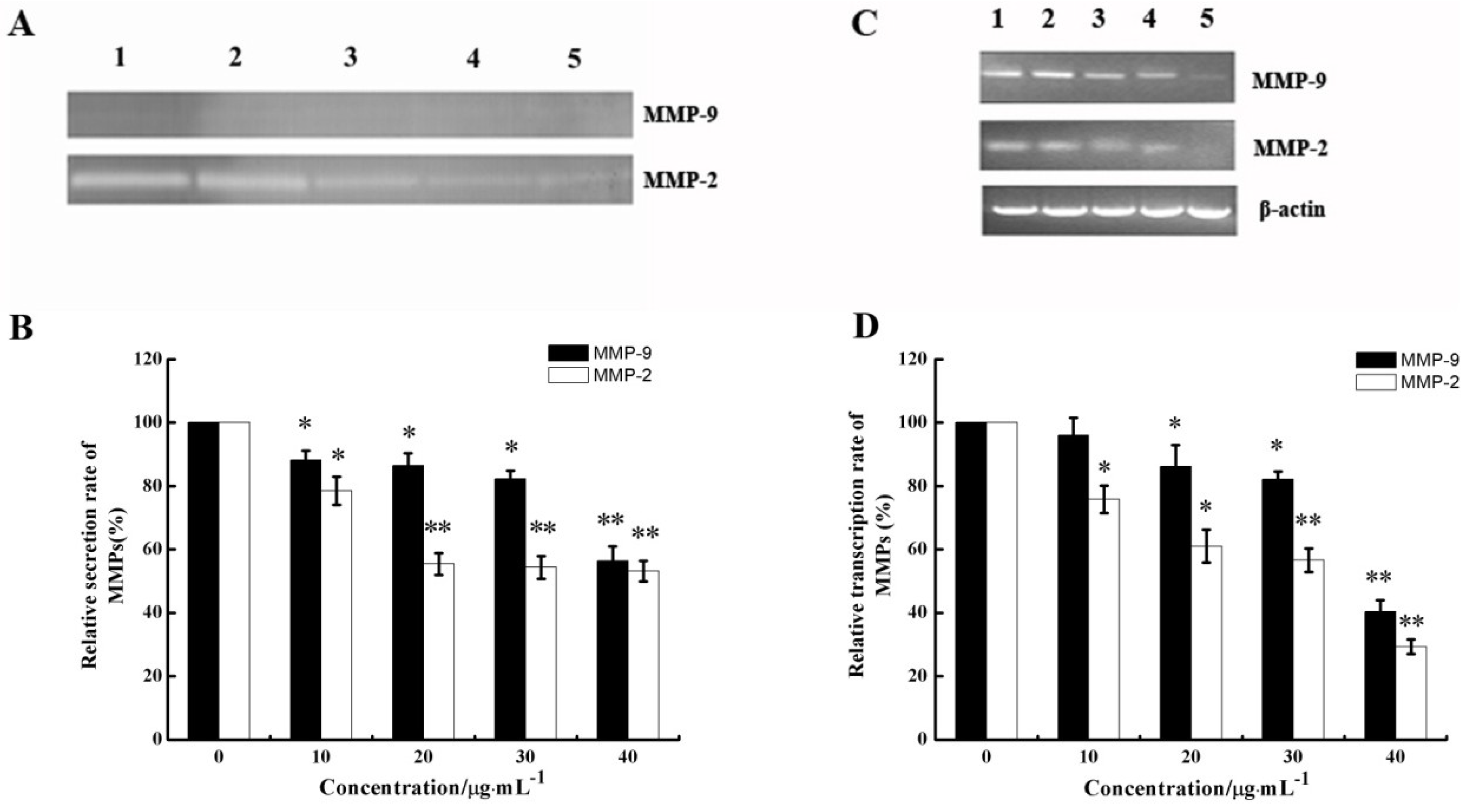
2.6. CS5931 Down-Regulates the Secretion and mRNA Expression of MMPs
3. Experimental Section
3.1. Materials and Reagents
3.2. Cells and Cell Culture
3.3. HUVEC Proliferation Assay
3.4. PI/Hoechst 33258 Staining Assay
3.5. Scratch-Wound Cell Migration Assay
3.6. Transwell Migration Assay
3.7. HUVEC Tube Formation Assay
3.8. Alkaline Phosphatase Staining for Visual Inspection in Zebrafish Embryos
3.9. Western Blotting Analysis
3.10. Production of Matrix Metalloproteinases (MMPs)
3.11. Semi-Quantitative Reverse Transcription and Polymerase Chain Reaction (RT-PCR)
3.12. Statistical Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences | Base pairs | |
|---|---|---|---|
| P1 | β-actin primer forward | 5′-ACACTGTGCCCATCTAGGAGG -3′ | 21 |
| P2 | β-actin primer reverse | 5′-AGGGGCCGGACTCGTCATACT-3′ | 21 |
| P3 | VEGF primer forward | 5′-TTGCTGCTCTACCTCCAC-3′ | 18 |
| P4 | VEGF primer reverse | 5′-AATGCTTTCTCCGCTCTG-3′ | 18 |
| P5 | MMP-2 primer forward | 5′-GGCCCTGTCACTCCTGAGAT-3′ | 20 |
| P6 | MMP-2 primer reverse | 5′-GGCATCCAGGTTATCGGGGA-3′ | 20 |
| P7 | MMP-9 primer forward | 5′-CGGAGCACGGAGACGGGTAT-3′ | 20 |
| P8 | MMP-9 primer reverse | 5′-TGAAGGGGAAGACGCACAGC-3′ | 20 |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, Z.; Zheng, L.; Yang, S.; Niu, R.; Chu, E.; Lin, X. N-acetylchitooligosaccharide is a potent angiogenic inhibitor both in vivo and in vitro. Biochem. Biophys. Res. Commun. 2007, 357, 26–31. [Google Scholar] [CrossRef]
- Ellis, L.M.; Liu, W.; Ahmad, S.A.; Fan, F.; Jung, Y.D.; Shaheen, R.M.; Reinmuth, N. Overview of angiogenesis: biologic implications for antiangiogenic therapy. Semin. Oncol. 2001, 28, 94–104. [Google Scholar] [CrossRef]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef]
- Folkman, J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nature Med. 1995, 1, 27–30. [Google Scholar] [CrossRef]
- Taraboletti, G.; Poli, M.; Dossi, R.; Manenti, L.; Borsotti, P.; Faircloth, G.T.; Broggini, M.; D’Incalci, M.; Ribatti, D.; Giavazzi, R. Antiangiogenic activity of aplidine, a new agent of marine origin. Br. J. Cancer 2004, 90, 2418–2424. [Google Scholar]
- Jubb, A.M.; Oates, A.J.; Holden, S.; Koeppen, H. Predicting benefit from anti-angiogenic agents in malignancy. Nature Rev. Cancer 2006, 6, 626–635. [Google Scholar] [CrossRef]
- Taraboletti, G.; Margosio, B. Antiangiogenic and antivascular therapy for cancer. Curr. Opin. Pharmacol. 2001, 1, 378–384. [Google Scholar] [CrossRef]
- Fidler, I.J.; Singh, R.K.; Yoneda, J.; Kumar, R.; Xu, L.; Dong, Z.; Bielenberg, D.R.; McCarty, M.; Ellis, L.M. Critical determinants of neoplastic angiogenesis. Cancer J. 2000, 6, S225–S236. [Google Scholar]
- Li, A.; Varney, M.L.; Valasek, J.; Godfrey, M.; Dave, B.J.; Singh, R.K. Autocrine role of interleukin-8 in induction of endothelial cell proliferation, survival, migration and MMP-2 production and angiogenesis. Angiogenesis 2005, 8, 63–71. [Google Scholar] [CrossRef]
- Folkman, J. What is the evidence that tumors are angiogenesis dependent? J. Natl. Cancer Inst. 1990, 82, 4–7. [Google Scholar] [CrossRef]
- Brower, V. Tumor angiogenesis-new drugs on the block. Nat. Biotechnol. 1999, 17, 963–968. [Google Scholar] [CrossRef]
- Loges, S.; Schmidt, T.; Carmeliet, P. Mechanisms of resistance to anti-angiogenic therapy and development of third-generation anti-angiogenic drug candidates. Genes Cancer 2010, 1, 12–25. [Google Scholar] [CrossRef]
- Deplanque, G.; Harris, A. Anti-angiogenic agents: Clinical trial design and therapies in development. Eur. Cancer 2000, 36, 1713–1724. [Google Scholar] [CrossRef]
- Suarez-Jimenez, G.-M.; Burgos-Hernandez, A.; Ezquerra-Brauer, J.-M. Bioactive peptides and depsipeptides with anticancer potential: Sources from marine animals. Mar. Drugs 2012, 10, 963–986. [Google Scholar] [CrossRef]
- Jha, R.K.; Zi-rong, X. Biomedical compounds from marine organisms. Mar. Drugs 2004, 2, 123–146. [Google Scholar]
- Zheng, L.; Lin, X.; Wu, N.; Liu, M.; Zheng, Y.; Sheng, J.; Ji, X.; Sun, M. Targeting cellular apoptotic pathway with peptides from marine organisms. Biochim. Biophys. Acta 2013, 1836, 42–48. [Google Scholar]
- Wrasidlo, W.; Mielgo, A.; Torres, V.A.; Barbero, S.; Stoletov, K.; Suyama, T.L.; Klemke, R.L.; Gerwick, W.H.; Carson, D.A.; Stupack, D.G. The marine lipopeptide somocystinamide A triggers apoptosis via caspase 8. Proc. Natl. Acad. Sci. USA 2008, 105, 2313–2318. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, C.; Liu, H.; Wang, F.; Zheng, L.; Zhao, J.; Chu, E.; Lin, X. A Novel Polypeptide Extracted From Ciona savignyi Induces Apoptosis Through a Mitochondrial-Mediated Pathway in Human Colorectal Carcinoma Cells. Clin. Colorectal Cancer 2012, 11, 207–214. [Google Scholar] [CrossRef]
- Zhao, J.; Wei, J.; Liu, M.; Xiao, L.; Wu, N.; Liu, G.; Huang, H.; Zhang, Y.; Zheng, L.; Lin, X. Cloning, characterization and expression of a cDNA encoding a granulin-like polypeptide in Ciona savignyi. Biochimie 2013, 95, 1611–1619. [Google Scholar] [CrossRef]
- Hanahan, D.; Bergers, G.; Bergsland, E. Less is more, regularly: metronomic dosing of cytotoxic drugs can target tumor angiogenesis in mice. J. Clin. Investig. 2000, 105, 1045–1047. [Google Scholar] [CrossRef]
- Kerbel, R.; Folkman, J. Clinical translation of angiogenesis inhibitors. Nature Rev. Cancer 2002, 2, 727–739. [Google Scholar] [CrossRef]
- Schirner, M. Antiangiogenic chemotherapeutic agents. Cancer Metastasis Rev. 2000, 19, 67–73. [Google Scholar] [CrossRef]
- Miller, K.D.; Sweeney, C.J.; Sledge, G.W. Redefining the target: chemotherapeutics as antiangiogenics. J. Clin. Oncol. 2001, 19, 1195–1206. [Google Scholar]
- Nicoletti, M.; Lucchini, V.; Massazza, G.; Abbott, B.; D’Incalci, M.; Giavazzi, R. Antitumor activity of taxol (NSC-125973) in human ovarian carcinomas growing in the peritoneal cavity of nude mice. Ann. Oncol. 1993, 4, 151–155. [Google Scholar]
- Yamamoto, T.; Sudo, K.; Fujita, T. Significant inhibition of endothelial cell growth in tumor vasculature by an angiogenesis inhibitor, TNP-470 (AGM-1470). Anticancer Res. 1994, 14, 1–3. [Google Scholar]
- Belotti, D.; Vergani, V.; Drudis, T.; Borsotti, P.; Pitelli, M.R.; Viale, G.; Giavazzi, R.; Taraboletti, G. The microtubule-affecting drug paclitaxel has antiangiogenic activity. Clin. Cancer Res. 1996, 2, 1843–1849. [Google Scholar]
- Prestayko, A.; D’Aoust, J.; Issell, B.; Crooke, S. Cisplatin (cis-diamminedichloroplatinum II). Cancer Treat. Rev. 1979, 6, 17–39. [Google Scholar] [CrossRef]
- Broggini, M.; Marchini, S.; Galliera, E.; Borsotti, P.; Taraboletti, G.; Erba, E.; Sironi, M.; Jimeno, J.; Faircloth, G.; Giavazzi, R. Aplidine, a new anticancer agent of marine origin, inhibits vascular endothelial growth factor (VEGF) secretion and blocks VEGF-VEGFR-1 (flt-1) autocrine loop in human leukemia cells MOLT-4. Leukemia 2003, 17, 52–59. [Google Scholar] [CrossRef]
- Zheng, L.; Ling, P.; Wang, Z.; Niu, R.; Hu, C.; Zhang, T.; Lin, X. A novel polypeptide from shark cartilage with potent anti-angiogenic activity. Cancer Biol. Ther. 2007, 6, 775–780. [Google Scholar] [CrossRef]
- Taraboletti, G.; Micheletti, G.; Rieppi, M.; Poli, M.; Turatto, M.; Rossi, C.; Borsotti, P.; Roccabianca, P.; Scanziani, E.; Nicoletti, M.I. Antiangiogenic and antitumor activity of IDN 5390, a new taxane derivative. Clin. Cancer Res. 2002, 8, 1182–1188. [Google Scholar]
- Mottet, D.; Bellahcène, A.; Pirotte, S.; Waltregny, D.; Deroanne, C.; Lamour, V.; Lidereau, R.; Castronovo, V. Histone deacetylase 7 silencing alters endothelial cell migration, a key step in angiogenesis. Circ. Res. 2007, 101, 1237–1246. [Google Scholar] [CrossRef]
- van Mil, A.; Grundmann, S.; Goumans, M.-J.; Lei, Z.; Oerlemans, M.I.; Jaksani, S.; Doevendans, P.A.; Sluijter, J.P. MicroRNA-214 inhibits angiogenesis by targeting Quaking and reducing angiogenic growth factor release. Cardiovasc. Res. 2012, 93, 655–665. [Google Scholar] [CrossRef]
- Dimmeler, S.; Dernbach, E.; Zeiher, A.M. Phosphorylation of the endothelial nitric oxide synthase at Ser-1177 is required for VEGF-induced endothelial cell migration. FEBS Lett. 2000, 477, 258–262. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book: A Guide for the Laboratory Use of Zebrafish (Brachydanio rerio); Institution of Neuro Science: Eugene, OR, USA, 1993; pp. 231–236. [Google Scholar]
- He, Z.-H.; He, M.-F.; Ma, S.-C.; But, P.P.-H. Anti-angiogenic effects of rhubarb and its anthraquinone derivatives. J. Ethnopharmacol. 2009, 121, 313–317. [Google Scholar] [CrossRef]
- Serbedzija, G.N.; Flynn, E.; Willett, C.E. Zebrafish angiogenesis: A new model for drug screening. Angiogenesis 1999, 3, 353–359. [Google Scholar] [CrossRef]
- Wei, J.; Liu, M.; Liu, H.; Wang, H.; Wang, F.; Zhang, Y.; Han, L.; Lin, X. Oleanolic acid arrests cell cycle and induces apoptosis via ROS-mediated mitochondrial depolarization and lysosomal membrane permeabilization in human pancreatic cancer cells. J. Appl. Toxicol. 2013, 33, 756–765. [Google Scholar] [CrossRef]
- Wang, H.; Wei, J.; Wu, N.; Liu, M.; Wang, C.; Zhang, Y.; Wang, F.; Liu, H.; Lin, X. Mere15, a novel polypeptide from Meretrix meretrix, inhibits adhesion, migration and invasion of human lung cancer A549 cells via down-regulating MMPs. Pharm. Biol. 2013, 51, 145–151. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Liu, G.; Liu, M.; Wei, J.; Huang, H.; Zhang, Y.; Zhao, J.; Xiao, L.; Wu, N.; Zheng, L.; Lin, X. CS5931, a Novel Polypeptide in Ciona savignyi, Represses Angiogenesis via Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs). Mar. Drugs 2014, 12, 1530-1544. https://doi.org/10.3390/md12031530
Liu G, Liu M, Wei J, Huang H, Zhang Y, Zhao J, Xiao L, Wu N, Zheng L, Lin X. CS5931, a Novel Polypeptide in Ciona savignyi, Represses Angiogenesis via Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs). Marine Drugs. 2014; 12(3):1530-1544. https://doi.org/10.3390/md12031530
Chicago/Turabian StyleLiu, Ge, Ming Liu, Jianteng Wei, Haijuan Huang, Yuyan Zhang, Jin Zhao, Lin Xiao, Ning Wu, Lanhong Zheng, and Xiukun Lin. 2014. "CS5931, a Novel Polypeptide in Ciona savignyi, Represses Angiogenesis via Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs)" Marine Drugs 12, no. 3: 1530-1544. https://doi.org/10.3390/md12031530
APA StyleLiu, G., Liu, M., Wei, J., Huang, H., Zhang, Y., Zhao, J., Xiao, L., Wu, N., Zheng, L., & Lin, X. (2014). CS5931, a Novel Polypeptide in Ciona savignyi, Represses Angiogenesis via Inhibiting Vascular Endothelial Growth Factor (VEGF) and Matrix Metalloproteinases (MMPs). Marine Drugs, 12(3), 1530-1544. https://doi.org/10.3390/md12031530