An Overview on the Marine Neurotoxin, Saxitoxin: Genetics, Molecular Targets, Methods of Detection and Ecological Functions

Abstract
:1. Marine Toxins Overview

{kind=link}
{kind=link}
| Backbone Structure | LD50 a, ARfD b | Organisms | Health Impacts c | Molecular Target | Pharmacology |
|---|---|---|---|---|---|
 | |||||
| Alkaloid | 3–10, 0.7 | Dinoflagellates: Pyrodinium bahamense, Alexandrium spp. Gymnodinium catenatum | PSP | Voltage-gated ion channels: Na (site 1); K; Ca | Pore blocker |
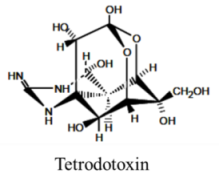 | |||||
| Alkaloid | 8, n/a | Fish and Bacteria | PFP | Na channel site 1 | Pore blocker |
 | |||||
| Polyketide | 170, n/a | Dinoflagellates | NSP | Na channel site 5 | enhanced activation and inactivation block |
 | |||||
| Polyketide | 380–460, 50 | Dinoflagellates | NSP | Unknown | Unknown; proposed interaction with cytoskeletal |
 | |||||
| Polyketide | 0.45, n/a | Dinoflagellates | CFP | Na channel site 5 | Shift in activation gating |
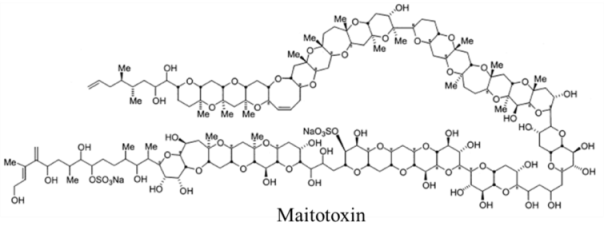 | |||||
| Polyketide | 0.15, n/a | Dinoflagellates | CFP | Cation channels | Channel modifier; allows non-selective ion passage |
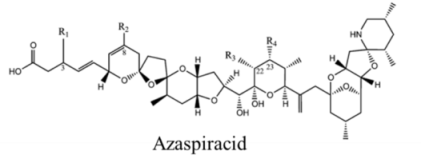 | |||||
| Polyketide | 200, 280 | Dinoflagellates | AZP | K channel | Pore blocker |
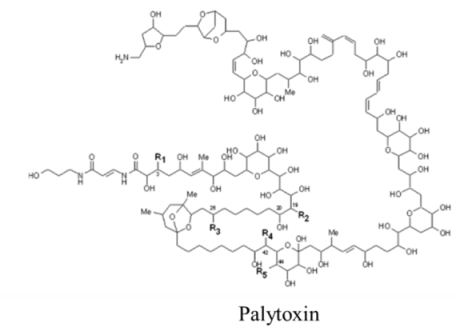 | |||||
| Polyketide | 0.45, n/a | Dinoflagellate | PP | Na:K ATPase | Channel modifier; allows non-selective ion passage |
| Cooliatoxin: structure uncharacterized | |||||
| Polyketide | 1000, n/a | Dinoflagellate | Symptoms similar to NSP | Unknown | Unknown |
| Ostreotoxin 3: structure uncharacterized | |||||
| Polyketide | 32100, n/a | Dinoflagellate | Symptoms similar to NSP | Unknown | Inactivation gating modifier |
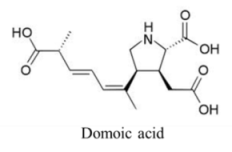 | |||||
| Amino acid | 3600, 100 | Diatom | ASP | Glutamate receptors | Depolarization via prolonged influx of Ca+ and Na+ |
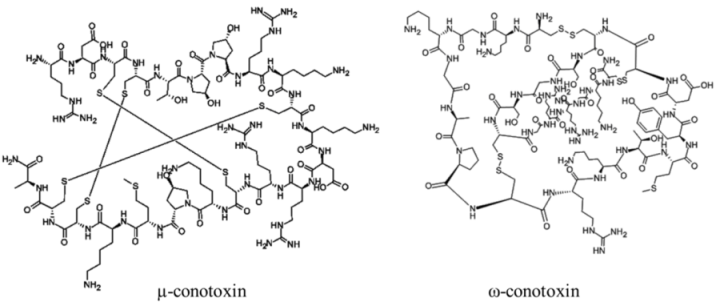 | |||||
| Polypeptide | 12000–30000, n/a | Cone snails | N/A | Na channel sites 1 (μ) and 6 (δ); Ca channel (ω); nicotinic acetylcholine receptors (α) | Pore blocker (μ); prolonged channel opening (δ) |
 | |||||
| Polypeptide | 170, n/a | Sea anemones | N/A | Na channel site 3; K channel | Slow/block channel inactivation |
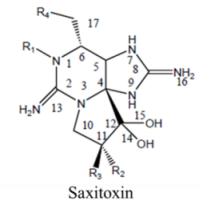
2. Saxitoxin and Derivatives: Structure and Chemistry
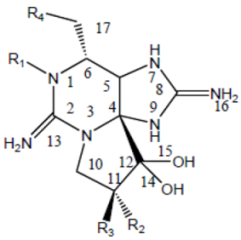 | |||||
|---|---|---|---|---|---|
| Division | Name a | R1 | R2 | R3 | R4 |
| STX | H | H | H | OCONH2 | |
| NeoSTX | OH | H | H | OCONH2 | |
| GTX1 | OH | OSO3− | H | OCONH2 | |
| Carbamate | GTX2 | H | OSO3− | H | OCONH2 |
| GTX3 | H | H | OSO3− | OCONH2 | |
| GTX4 | OH | H | OSO3− | OCONH2 | |
| GTX5 (B1) | H | H | H | OCONHSO3− | |
| GTX6 (B2) | OH | H | H | OCONHSO3− | |
| C1 | H | OSO3− | H | OCONHSO3− | |
| N-sulfocarbamoyl | C2 | H | H | OSO3− | OCONHSO3− |
| C3 | OH | OSO3− | H | OCONHSO3− | |
| C4 | OH | H | OSO3− | OCONHSO3− | |
| dcSTX | H | H | H | OH | |
| dcNeoSTX | OH | H | H | OH | |
| dcGTX1 | OH | OSO3− | H | OH | |
| Decarbamoyl | dcGTX2 | H | OSO3− | H | OH |
| dcGTX3 | H | H | OSO3− | OH | |
| dcGTX4 | OH | H | OSO3− | OH | |
| doSTX | H | H | H | H | |
| Deoxydecarbamoyl | doGTX2 | H | H | OSO3− | H |
| doGTX3 | H | OSO3− | H | H | |
3. Molecular Targets and Pharmacology
3.1. Ion Channel Structure
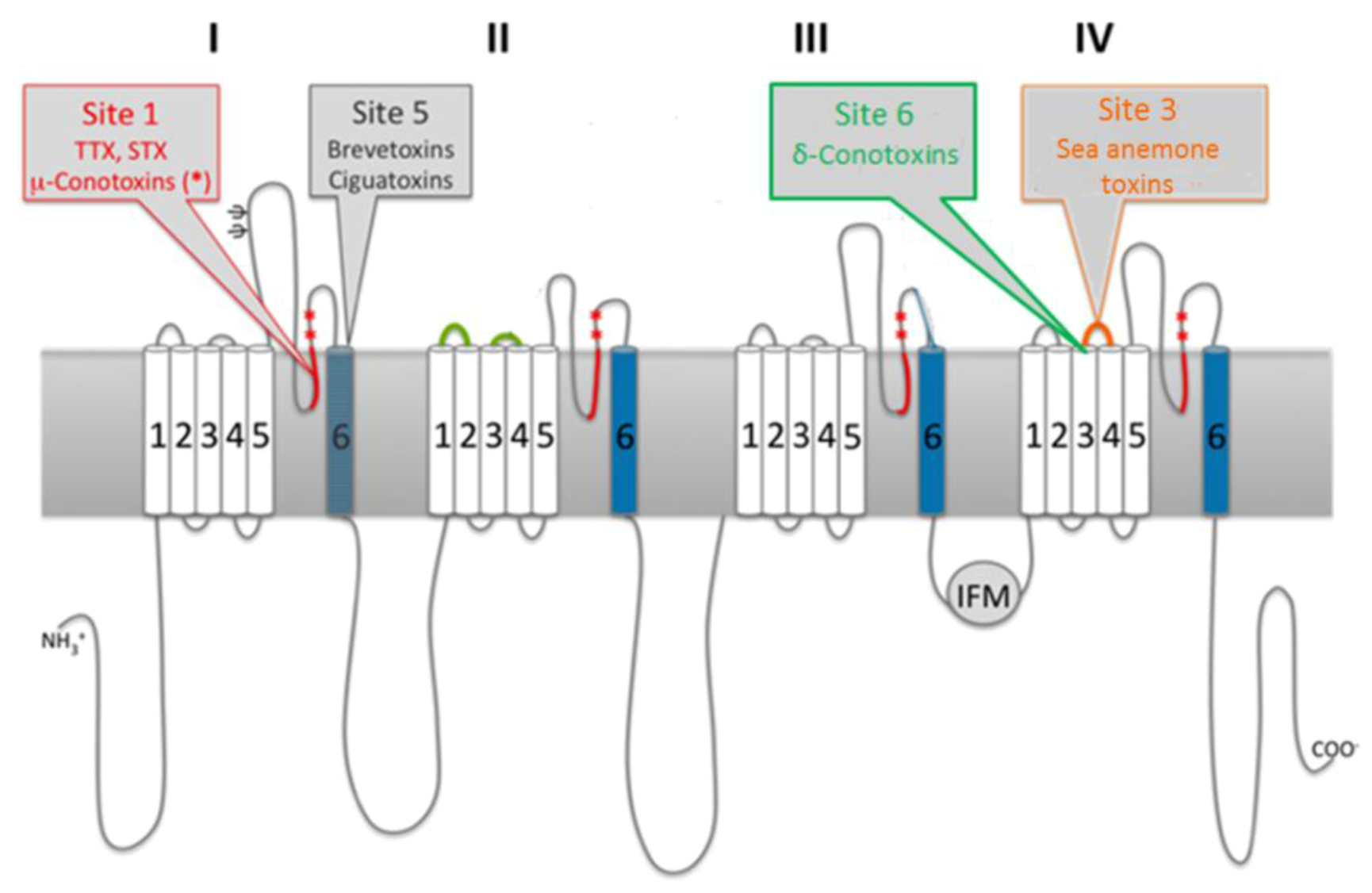
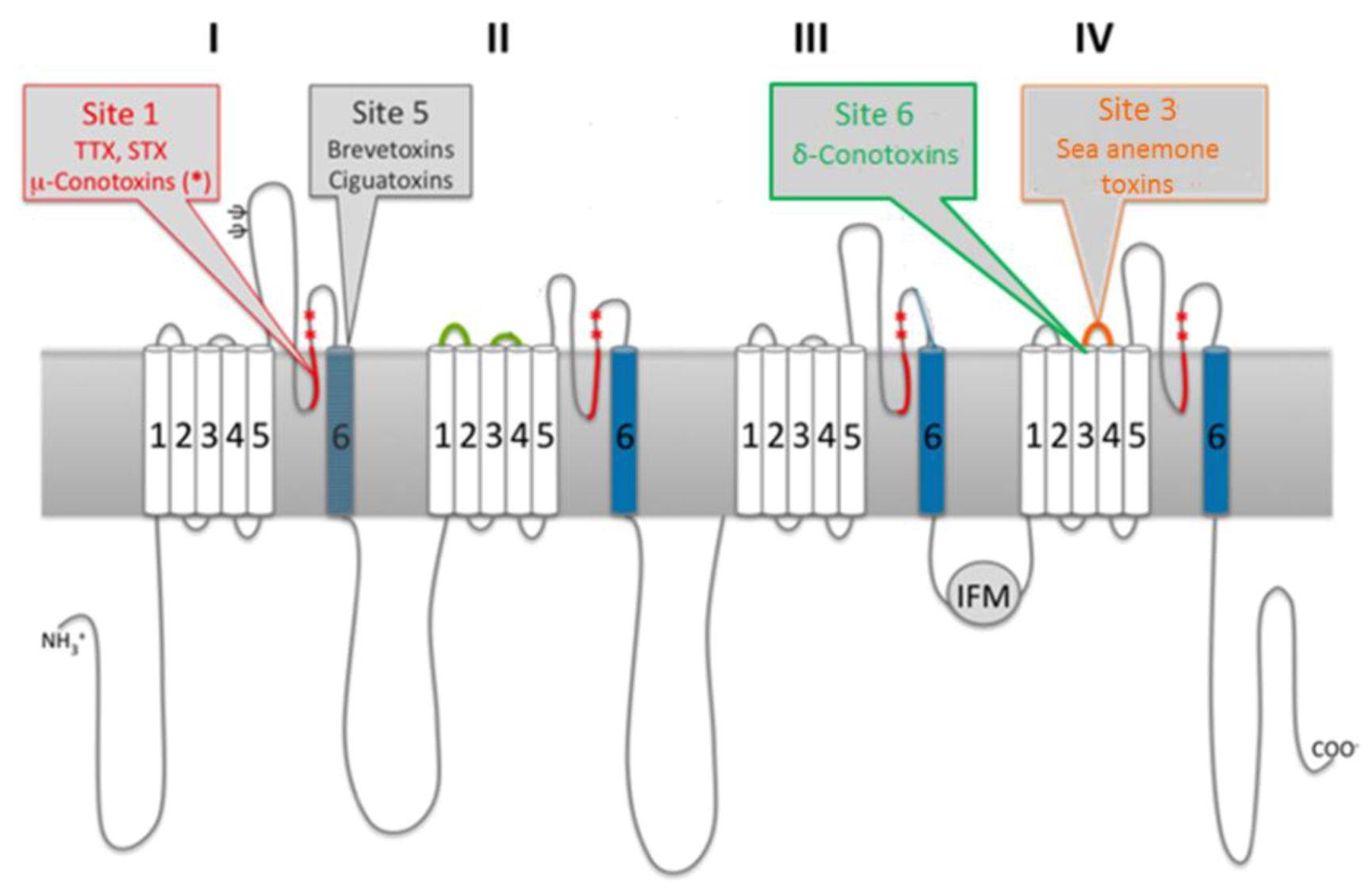
3.2. Marine Neurotoxins Binding Sites
3.3. Molecular Targets of Saxitoxin
3.3.1. Ion Channels
3.3.2. Saxiphilin
3.3.3. Copper Transporters
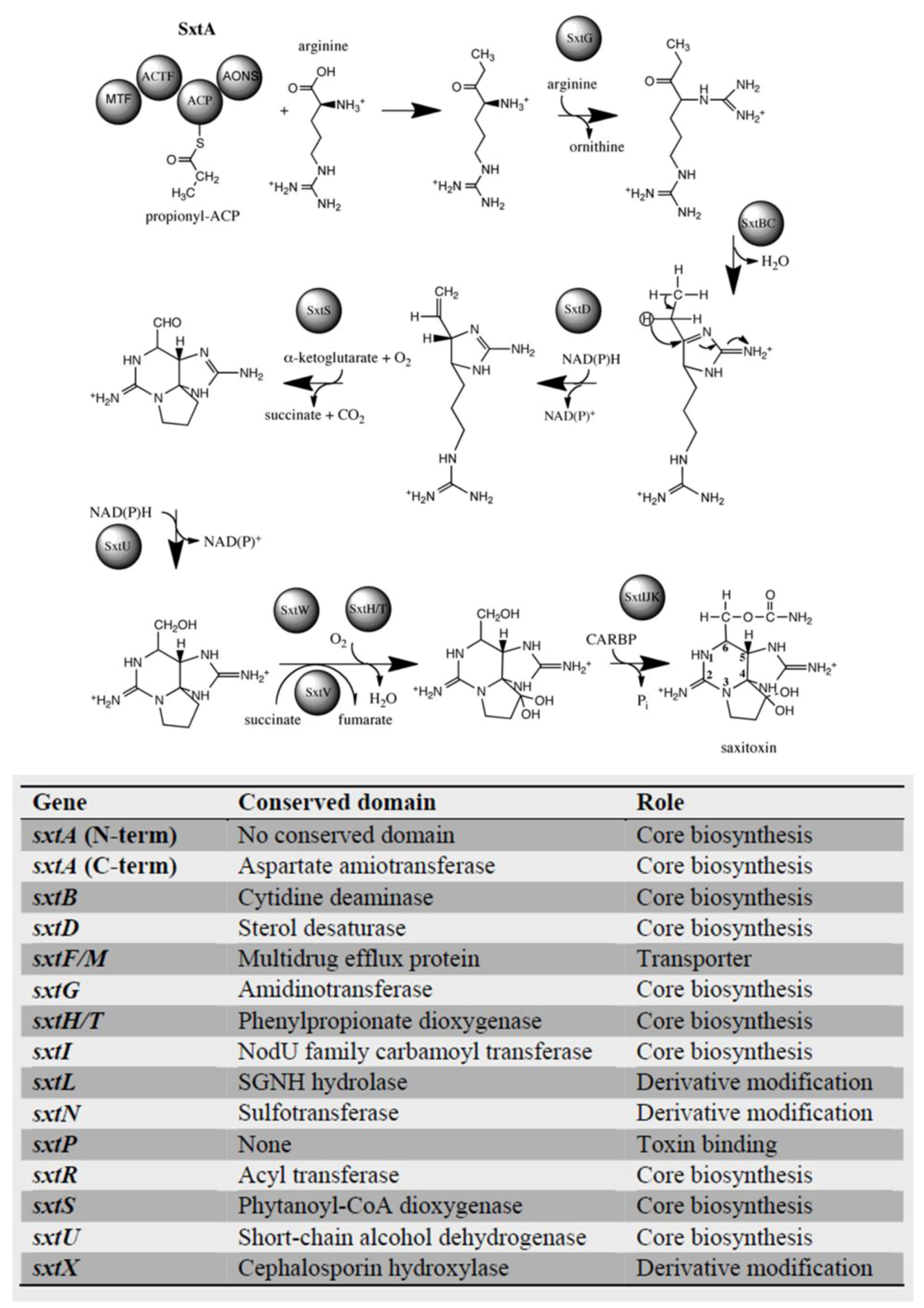
4. Genetics of Saxitoxin Production
4.1. Dinoflagellate Genome Overview
4.2. Dinoflagellate Saxitoxin Biosynthesis Genes
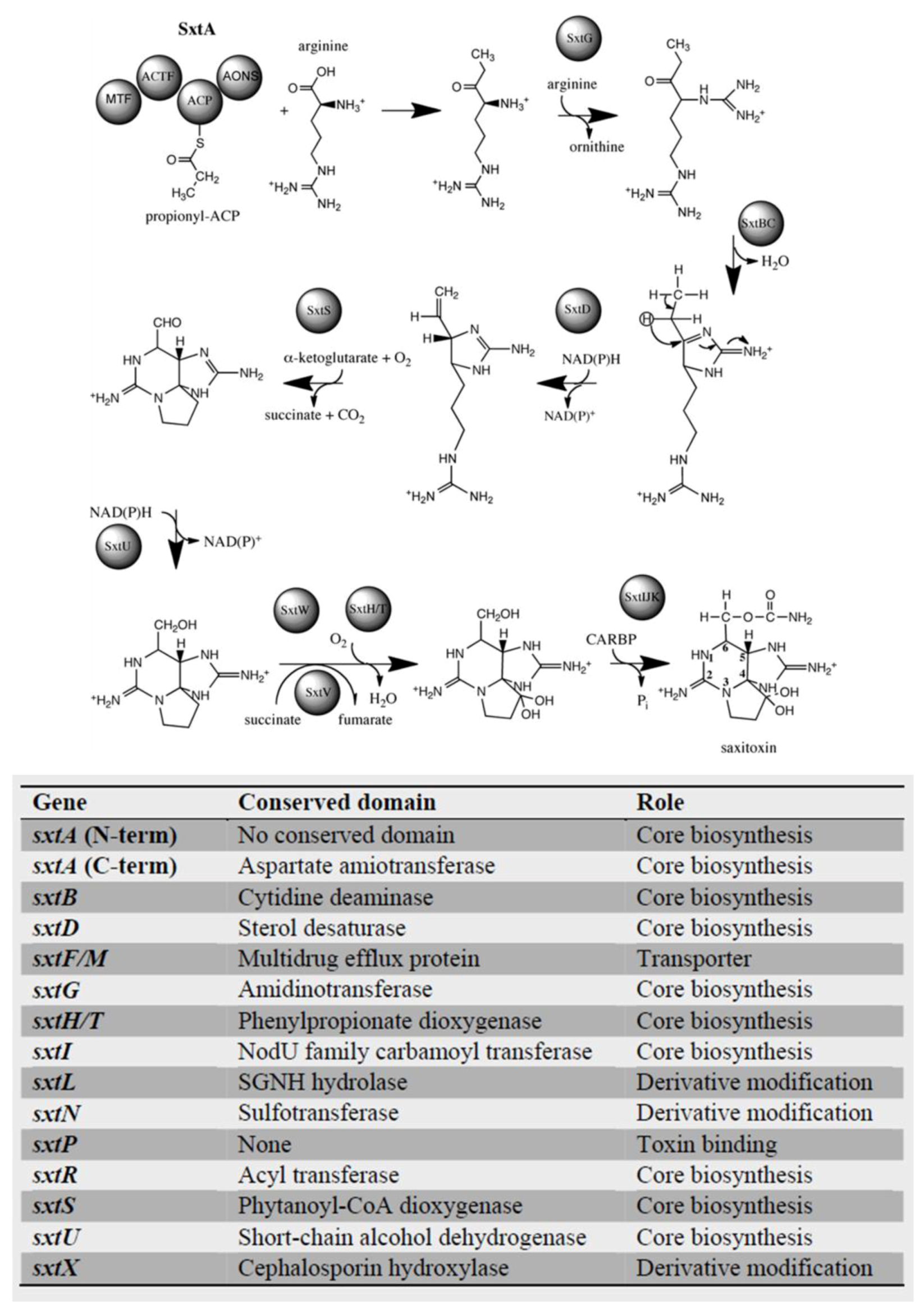
4.3. Saxitoxin-Related Enzymes
5. Effects of Saxitoxin on Human and Environmental Health
6. Methods of Detection
6.1. In Vivo Animal Bioassays
6.2. Analytical Techniques
6.3. In Vitro Functional Assays
6.4. In Vitro Cell Assays
6.5. Immunoassays
6.6. Molecular Tools
7. Ecological Functions
8. Conclusions
References
- Wang, D.-Z. Neurotoxins from marine dinoflagallates: A brief review. Mar. Drugs 2008, 6, 349–371. [Google Scholar] [CrossRef]
- Rein, K.S.; Borrone, J. Polyketides from dinoflagellates: Origins, pharmacology and biosynthesis. Comp. Biochem. Physiol. B 1999, 124, 117–131. [Google Scholar] [CrossRef]
- Anderson, P.D. Bioterrorism: Toxins as weapons. J. Pharm. Pract. 2012, 25, 121–129. [Google Scholar] [CrossRef]
- Harada, T.; Oshima, Y.; Yasumoto, T. Studies on paralytic shellfish poisoning in tropical waters: 4. Structures of 2 paralytic shellfish toxins, gonyautoxin-V and gonyautoxin-VI isolated from a tropical dinoflagellate, Pyrodinium bahamense var. compressa. Agric. Biol. Chem. 1982, 46, 1861–1864. [Google Scholar] [CrossRef]
- Oshima, Y.; Hasegawa, M.; Yasumoto, T.; Hallegraeff, G.; Blackburn, S. Dinoflagellate Gymnodinium catenatum as the source of paralytic shellfish toxins in Tasmanian shellfish. Toxicon 1987, 25, 1105–1111. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Steffensen, D.A.; Wetherbee, R. Three estuarine Australian dinoflagellates that can produce paralytic shellfish toxins. J. Plankton Res. 1988, 10, 533–541. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Sullivan, J.J.; Hall, S.; Lee, C. Dynamics and physiology of saxitoxin production by the dinoflagellates Alexandrium spp. Mar. Biol. 1990, 104, 511–524. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Evans, W.R.; Yin, Q.Q.; Bell, P.; Moczydlowski, E. Evidence for paralytic shellfish poisons in the freshwater cyanobacterium Lyngbya wollei (Farlow ex Gomont) comb. nov. Appl. Environ. Microbiol. 1997, 63, 3104–3110. [Google Scholar]
- Negri, A.P.; Jones, G.J. Bioaccumulation of paralytic shellfish poisoning (PSP) toxins from the cyanobacterium Anabaena circinalis by the freshwater mussel Alathyria condola. Toxicon 1995, 33, 667–678. [Google Scholar] [CrossRef]
- Lagos, N.; Onodera, H.; Zagatto, P.A.; Andrinolo, D.; Azevedo, S.M.; Oshima, Y. The first evidence of paralytic shellfish toxins in the freshwater cyanobacterium Cylindrospermopsis raciborskii, isolated from Brazil. Toxicon 1999, 37, 1359–1373. [Google Scholar] [CrossRef]
- Pomati, F.; Sacchi, S.; Rossetti, C.; Giovannardi, S.; Onodera, H.; Oshima, Y.; Neilan, B.A. The freshwater cyanobacterium Planktothrix sp. FP1: Molecular identification and detection of paralytic shellfish poisoning toxins. J. Phycol. 2000, 36, 553–562. [Google Scholar] [CrossRef]
- Ferreira, F.M.B.; Soler, J.M.F.; Fidalgo, M.L.; Fernandez-Vila, P. PSP toxins from Aphanizomenon flos-aquae (cyanobacteria) collected in the Crestuma-Lever reservoir (Douro river, northern Portugal). Toxicon 2001, 39, 757–761. [Google Scholar] [CrossRef]
- Shimizu, Y. Microalgal metabolites. Chem. Rev. 1993, 93, 1685–1698. [Google Scholar] [CrossRef]
- Pearson, L.; Mihali, T.; Moffitt, M.; Kellmann, R.; Neilan, B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar. Drugs 2010, 8, 1650–1680. [Google Scholar] [CrossRef]
- Araoz, R.; Molgo, J.; Tandeau de Marsac, N. Neurotoxic cyanobacterial toxins. Toxicon 2010, 56, 813–828. [Google Scholar] [CrossRef]
- Schantz, E.J.; Ghazarossian, V.E.; Schnoes, H.K.; Strong, F.M.; Springer, J.P.; Pezzanite, J.O.; Clardy, J. Structure of saxitoxin. J. Am. Chem. Soc. 1975, 97, 1238–1239. [Google Scholar] [CrossRef]
- Rogers, R.S.; Rapoport, H. The pKas of saxitoxin. J. Am. Chem. Soc. 1980, 102, 7335–7339. [Google Scholar] [CrossRef]
- Shimizu, Y.; Hsu, C.P.; Genenah, A. Structure of saxitoxin in solutions and stereochemistry of dihydrosaxitoxins. J. Am. Chem. Soc. 1981, 103, 605–609. [Google Scholar] [CrossRef]
- Schantz, E.J.; Lynch, J.M.; Vayvada, G.; Matsumot, K.; Rapoport, H. Purification and characterization of poison produced by Gonyaulax catenella in axenic culture. Biochemistry 1966, 5, 1191–1195. [Google Scholar] [CrossRef]
- Strichartz, G. Structural determinants of the affinity of saxitoxin for neuronal sodium channels—Electrophysiological studies on frog peripheral nerve. J. Gen. Physiol. 1984, 84, 281–305. [Google Scholar] [CrossRef]
- Genenah, A.A.; Shimizu, Y. Specific toxicity of paralytic shellfish poisons. J. Agric. Food Chem. 1981, 29, 1289–1291. [Google Scholar] [CrossRef]
- Hall, S.; Strichartz, G.; Moczydlowski, E.; Ravindran, A.; Reichardt, P.B. The Saxitoxins: Sources, Chemistry, and Pharmacology. In Marine Toxins: Origin, Structure, and Molecular Pharmacology; Hall, S., Strichartz, G., Eds.; American Chemical Society Symposium Series 418; American Chemical Society: Washington, DC, USA, 1990; pp. 29–65. [Google Scholar]
- Llewellyn, L.E. Predicitive toxinology: An initial foray using calculated molecular descriptors to decribe toxicity using saxitoxin as a model. Toxicon 2007, 50, 901–913. [Google Scholar] [CrossRef]
- Cestele, S.; Catterall, W.A. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Strong, M.; Chandy, K.G.; Gutman, G.A. Molecular evolution of voltage-sensitive ion channel genes: On the origins of electrical excitability. Mol. Biol. Evol. 1993, 10, 221–242. [Google Scholar] [Green Version]
- Charalambous, K.; Wallace, B.A. NaChBac: The long lost sodium channel ancestor. Biochemistry 2011, 50, 6742–6752. [Google Scholar] [CrossRef]
- Stevens, M.; Peigneur, S.; Tytgat, J. Neurotoxins and their binding areas on voltage-gated sodium channels. Front. Pharmacol. 2011, 2, 1–13. [Google Scholar]
- Catterall, W.A.; Cestele, S.; Yarov-Yarovoy, V.; Yu, F.H.; Konoki, K.; Scheuer, T. Voltage-gated ion channels and gating modifier toxins. Toxicon 2007, 49, 124–141. [Google Scholar] [CrossRef]
- Catterall, W.A. The Voltage Sensitive Sodium Channel: A Receptor for Multiple Neurotoxins. In Toxic Dinoflagellates; Anderson, D.M., White, A.W., Baden, D.G., Eds.; Elsevier Science Publishing Co., Inc.: New York, NY, USA, 1985; pp. 329–342. [Google Scholar]
- Noda, M.; Suzuki, H.; Numa, S.; Stuhmer, W. A single point mutation confers tetrodotoxin and saxitoxin insensitivity on the sodium channel II. FEBS Lett. 1989, 259, 213–216. [Google Scholar] [CrossRef]
- Terlau, H.; Heinemann, S.H.; Stuhmer, W.; Pusch, M.; Conti, F.; Imoto, K.; Numa, S. Mapping the site of block by tetrodotoxin and saxitoxin of sodium channel II. FEBS Lett. 1991, 293, 93–96. [Google Scholar] [CrossRef]
- Hartshorne, R.P.; Catterall, W.A. The sodium channel from rat brain—Purification and subunit composition. J. Biol. Chem. 1984, 259, 1667–1675. [Google Scholar]
- Kao, C.Y.; Walker, S.E. Active groups of saxitoxin and tetrodoxin as deduced from actions of saxitoxin analogs on frog muscle and squid axon. J. Physiol. 1982, 323, 619–637. [Google Scholar]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200–222. [Google Scholar] [CrossRef]
- Baden, D.G.; Trainer, V.L. The Mode and Action of Toxins and Seafood Poisoning. In Algal Toxins in Seafood and Drinking Water; Falconer, I., Ed.; Academic Press: San Diego, CA, USA, 1993. [Google Scholar]
- Wang, J.X.; Salata, J.J.; Bennett, P.B. Saxitoxin is a gating modifier of hERG K+ channels. J. Gen. Physiol. 2003, 121, 583–598. [Google Scholar] [CrossRef]
- Murray, S.A.; Mihali, T.K.; Neilan, B.A. Extraordinary conservation, gene loss, and positive selection in the evolution of an ancient neurotoxin. Mol. Biol. Evol. 2011, 28, 1173–1182. [Google Scholar] [CrossRef]
- Su, Z.; Sheets, M.; Ishida, H.; Li, F.H.; Barry, W.H. Saxitoxin blocks L-type ICa. J. Pharmacol. Exp. Ther. 2004, 308, 324–329. [Google Scholar]
- Zakon, H.H. Adaptive evolution of voltage-gated sodium channels: The first 800 million years. Proc. Natl. Acad. Sci. USA 2012, 109, 10619–10625. [Google Scholar] [CrossRef]
- Mahar, J.; Lukacs, G.L.; Li, Y.; Hall, S.; Moczydlowski, E. Pharmacologiocal and biochemical properties of saxiphilin, a soluble saxitoxin-binding protein from the bullfrog (Rana catesbeiana). Toxicon 1991, 29, 53–71. [Google Scholar] [CrossRef]
- Morabito, M.A.; Moczydlowski, E. Molecular cloning of bullfrog saxiphilin—A unique relative of the transferrin family that binds saxitoxin. Proc. Natl. Acad. Sci. USA 1994, 91, 2478–2482. [Google Scholar] [CrossRef]
- Gaffney, J.P.; Valentine, A.M. Beyond bilobal: Transferrin homologs having unusual domain architectures. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 212–217. [Google Scholar] [CrossRef]
- Morabito, M.A.; Llewellyn, L.E.; Moczydlowski, E.G. Expression of saxiphilin in insect cells and localization of the saxitoxin-binding site to the C-terminal domain homologous to the C-lobe of transferrins. Biochemistry 1995, 34, 13027–13033. [Google Scholar] [CrossRef]
- Llewellyn, L.E.; Moczydlowski, E.G. Characterization of saxitoxin binding to saxiphilin, a relative of the transferrin family that displays pH-dependent ligand-binding. Biochemistry 1994, 33, 12312–12322. [Google Scholar] [CrossRef]
- Cusick, K.D.; Minkin, S.C.; Dodani, S.C.; Chang, C.J.; Wilhelm, S.W.; Sayler, G.S. Inhibition of copper uptake in yeast reveals the copper transporter Ctr1p as a potential molecular target of saxitoxin. Environ. Sci. Technol. 2012, 46, 2959–2966. [Google Scholar] [CrossRef]
- Hill, K.; Hassett, R.; Kosman, D.; Merchant, S. Regulated copper uptake in Chlamydomonas reinhardtii in response to copper availabilty. Plant. Physiol. 1996, 112, 697–704. [Google Scholar]
- Dancis, A.; Haile, D.; Yuan, D.S.; Klausner, R.D. The Saccharomyces cerevisiae copper transport protein (Ctr1p). Biochemical characterization, regulation by copper, and physiologic role in copper uptake. J. Biol. Chem. 1994, 269, 25660–25667. [Google Scholar]
- Dancis, A.; Yuan, D.S.; Haile, D.; Askwith, C.; Eide, D.; Moehle, C.; Kaplan, J.; Klausner, R.D. Molecular characterization of a copper transport protein in S.cerevisiae: An unexpected role for copper in iron transport. Cell 1994, 76, 393–402. [Google Scholar] [CrossRef]
- Page, M.D.; Kropat, J.; Hamel, P.P.; Merchant, S.S. Two Chlamydomonas CTR copper transporters with a novel Cys-Met motif are localized to the plasma membrane and function in copper assimilation. Plant Cell 2009, 21, 928–943. [Google Scholar] [CrossRef]
- Aller, S.G.; Unger, V.M. Projection structure of the human copper transporter CTR1 at 6-Ã resolution reveals a compact trimer with a novel channel-like architecture. Proc. Natl. Acad. Sci. USA 2006, 103, 3627–3632. [Google Scholar] [CrossRef]
- Henderson, R.; Ritchie, J.M.; Strichartz, G.R. Evidence that tetrodotoxin and saxitoxin act at a metal cation binding site in sodium channels of nerve membrane. Proc. Natl. Acad. Sci. USA 1974, 71, 3936–3940. [Google Scholar] [CrossRef]
- De Feo, C.J.; Aller, S.G.; Unger, V.M. A structural perspective on copper uptake in eukaryotes. Biometals 2007, 20, 705–716. [Google Scholar] [CrossRef]
- Shimizu, Y. Microalgal metabolites: A new perspective. Annu. Rev. Microbiol. 1996, 50, 431–465. [Google Scholar] [CrossRef]
- Kellmann, R.; Mihali, T.K.; Jeon, Y.J.; Pickford, R.; Pomati, F.; Neilan, B.A. Biosynthetic intermediate analysis and functional homology reveal a saxitoxin gene cluster in cyanobacteria. App. Environ. Microbiol. 2008, 74, 4044–4053. [Google Scholar] [CrossRef]
- Kellmann, R.; Neilan, B.A. Biochemical characterization of paralytic shellfish toxin biosynthesis in vitro. J. Phycol. 2007, 43, 497–508. [Google Scholar] [CrossRef]
- Mihali, T.K.; Kellmann, R.; Neilan, B.A. Characterisation of the paralytic shellfish toxin biosynthesis gene clusters in Anabaena circinalis AWQC131C and Aphanizomenon sp. NH-5. BMC Biochem. 2009, 10, 8. [Google Scholar] [CrossRef]
- Mihali, T.K.; Carmichael, W.W.; Neilan, B.A. A putative gene cluster from a Lyngbya wollei bloom that encodes paralytic shellfish toxin biosynthesis. PLoS One 2011, 6, e14657. [Google Scholar]
- Lin, S.J. Genomic understanding of dinoflagellates. Res. Microbiol. 2011, 162, 551–569. [Google Scholar] [CrossRef]
- Moustafa, A.; Evans, A.N.; Kulis, D.M.; Hackett, J.D.; Erdner, D.L.; Anderson, D.M.; Bhattacharya, D. Transcriptome profiling of a toxic dinoflagellate reveals a gene-rich protist and a potential impact on gene expression due to bacterial presence. PLoS One 2010, 5, e9688. [Google Scholar] [CrossRef]
- Hou, Y.; Lin, S. Distinct gene number-genome size relationships for eukaryotes and non-eukaryotes: Gene content estimation for dinoflagellate genomes. PLoSOne 2009, 4, e6978. [Google Scholar]
- Erdner, D.L.; Anderson, D.M. Global transcriptional profiling of the toxic dinoflagellate Alexandrium fundyense using massively parallel signature sequencing. BMC Genomics 2006, 7, 88. [Google Scholar] [CrossRef]
- Stuken, A.; Orr, R.J.S.; Kellmann, R.; Murray, S.A.; Neilan, B.A.; Jakobsen, K.S. Discovery of nuclear-encoded genes for the neurotoxin saxitoxin in dinoflagellates. PLoS One 2011, 6, e20096. [Google Scholar]
- Hackett, J.D.; Wisecarver, J.H.; Brosnahan, M.L.; Kulis, D.M.; Anderson, D.M.; Bhattacharya, D.; Plumley, F.G.; Erdner, D.L. Evolution of saxitoxin synthesis in cyanobacteria and dinoflagellates. Mol. Biol. Evol. 2013, 30, 70–78. [Google Scholar] [CrossRef]
- Bachvaroff, T.R.; Place, A.R. From stop to start: Tandem gene arrangement, copy number and trans-splicing sites in the dinoflagellate Amphidinium carterae. PLoS One 2008, 3, e2929. [Google Scholar] [CrossRef]
- Le, Q.H.; Markovic, P.; Hastings, J.W.; Jovine, R.V.M.; Morse, D. Structure and organization of the peridinin chlorophyll a binding protein gene in Gonyaulax polyedra. Mol. Gen. Genet. 1997, 255, 595–604. [Google Scholar] [CrossRef]
- Moustafa, A.; Loram, J.E.; Hackett, J.D.; Anderson, D.M.; Plumley, F.G.; Bhattacharya, D. Origin of saxitoxin biosynthetic genes in cyanobacteria. PLoS One 2009, 4, e5758. [Google Scholar]
- Salcedo, T.; Upadhyay, R.J.; Nagasaki, K.; Bhattacharya, D. Dozens of toxin-related genes are expressed in a nontoxic strain of the dinoflagellate Heterocapsa circularisquama. Mol. Biol. Evol. 2012, 29, 1503–1506. [Google Scholar] [CrossRef]
- Yoshida, T.; Sako, Y.; Uchida, A.; Kakutani, T.; Arakawa, O.; Noguchi, T.; Ishida, Y. Purification and characterization of sulfotransferase specific to O-22 of 11-hydroxy saxitoxin from the toxic dinoflagellate Gymnodinium catenatum (Dinophyceae). Fish. Sci. 2002, 68, 634–642. [Google Scholar] [CrossRef]
- Sako, Y.; Yoshida, T.; Uchida, A.; Arakawa, O.; Noguchi, T.; Ishida, Y. Purification and characterization of a sulfotransferase specific to N-21 of saxitoxin and gonyautoxin 2 + 3 from the toxic dinoflagellate Gymnodinium catenatum (Dinophyceae). J. Phycol. 2001, 37, 1044–1051. [Google Scholar] [CrossRef]
- Landsberg, J.H.; Hall, S.; Johannessen, J.N.; White, K.D.; Conrad, S.M.; Abbott, J.P.; Flewelling, L.J.; Richardson, R.W.; Dickey, R.W.; Jester, E.L.E.; et al. Saxitoxin puffer fish poisoning in the United States, with the first report of Pyrodinium bahamense as the putative toxin source. Environ. Health Perspect. 2006, 114, 1502–1507. [Google Scholar] [CrossRef]
- Etheridge, S.M. Paralytic shellfish poisoning: Seafood safety and human health perspectives. Toxicon 2010, 56, 108–122. [Google Scholar]
- Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-traditional vectors for paralytic shellfish poisoning. Mar. Drugs 2008, 6, 308–348. [Google Scholar] [CrossRef]
- Van Dolah, F.M. Marine algal toxins: Origins, health effects, and their increased occurrence. Environ. Health Perspect. 2000, 108, 133–141. [Google Scholar] [CrossRef]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar]
- Abbott, J.P.; Flewelling, L.J.; Landsberg, J.H. Saxitoxin monitoring in three species of Florida puffer fish. Harmful Algae 2009, 8, 343–348. [Google Scholar] [CrossRef]
- Tamplin, M.L. A Bacterial Source of Tetrodotoxins and Saxitoxins. In Marine Toxins: Origin, Structure, and Molecular Pharmacology; Hall, S.L., Strichartz, G., Eds.; American Chemical Society: Washington, DC, USA, 1990; pp. 78–86. [Google Scholar]
- Schantz, E.J.; McFarren, E.F.; Schafer, M.L.; Lewis, K.H. Purified shellfsih poison for bioassay standardization. J. Assoc. Off. Anal. Chem. 1958, 41, 160–168. [Google Scholar]
- Aune, T.; Ramstad, H.; Heidenreich, B.; Landsverk, T.; Waaler, T.; Egaas, E.; Julshamn, K. Zinc accumulation in oysters giving mouse deaths in paralytic shellfish poisoning bioassay. J. Shellfish Res. 1998, 17, 1243–1246. [Google Scholar]
- Turner, A.D.; Dhanji-Rapkova, M.; Algoet, M.; Suarez-Isla, B.A.; Cordova, M.; Caceres, C.; Murphy, C.J.; Casey, M.; Lees, D.N. Investigations into matrix components affecting the performance of the official bioassay reference method for quantitation of paralytic shellfish poisoning toxins in oysters. Toxicon 2012, 59, 215–230. [Google Scholar]
- Aune, T.; Aasen, J.A.B.; Miles, C.O.; Larsen, S. Effect of mouse strain and gender on LD50 of yessotoxin. Toxicon 2008, 52, 535–540. [Google Scholar]
- Food and Agricultural Organization. Marine Biotoxins FAO Food and Nutrition Paper 80; Food and Agricultural Organization of the United Nations: Rome, Italy, 2004.
- Humpage, A.R.; Magalhaes, V.F.; Froscio, S.M. Comparison of analytical tools and biological assays for detection of paralytic shellfish poisoning toxins. Anal. Bioanal. Chem. 2010, 397, 1655–1671. [Google Scholar] [CrossRef]
- Louzao, M.C.; Vieytes, M.R.; Baptista de Sousa, J.M.V.; Leira, F.; Botana, L.M. A fluorometric method based on changes in membrane potential for screening paralytic shellfish toxins in mussels. Anal. Biochem. 2001, 289, 246–250. [Google Scholar] [CrossRef]
- Cheng, J.P.; Pi, S.S.; Ye, S.F.; Gao, H.M.; Yao, L.; Jiang, Z.Y.; Song, Y.L.; Xi, L. A new simple screening method for the detection of paralytic shellfish poisoning toxins. Chin. J. Ocean. Limnol. 2012, 30, 786–790. [Google Scholar] [CrossRef]
- Sullivan, J.J. High-Performance Liquid Chromatographic Method Applied to Paralytic Shellfish Poisoning Research. In Marine Toxins: Origin, Structure, and Molecular Pharmacology; Hall, S., Strichartz, G., Eds.; American Chemical Society: Washington, DC, USA, 1990; pp. 66–77. [Google Scholar]
- Turner, A.D.; Hatfield, R.C. Refinement of AOAC official method (SM) 2005.06 liquid chromatography-fluorescence detection method to improve performance characteristics for the determination of paralytic shellfish toxins in king and queen scallops. J. AOAC Int. 2012, 95, 129–142. [Google Scholar] [CrossRef]
- Dell’Aversano, C.; Eaglesham, G.K.; Quilliam, M.A. Analysis of cyanobacterial toxins by hydrophilic interaction liquid chromatography-mass spectrometry. J. Chromatogr. A 2004, 1028, 155–164. [Google Scholar] [CrossRef]
- Dell’Aversano, C.; Hess, P.; Quilliam, M.A. Hydrophilic interaction liquid chromatography-mass spectrometry for the analysis of paralytic shellfish poisoning (PSP) toxins. J. Chromatogr. A 2005, 1081, 190–201. [Google Scholar] [CrossRef]
- Halme, M.; Rapinoja, M.L.; Karjalainen, M.; Vanninen, P. Verification and quantification of saxitoxin from algal samples using fast and validated hydrophilic interaction liquid chromatography-tandem mass spectrometry method. J. Chromatogr. B 2012, 880, 50–57. [Google Scholar] [CrossRef]
- Davio, S.R.; Fontelo, P.A. A competitive displacement assay to detect saxitoxin and tetrodotoxin. Anal. Biochem. 1984, 141, 199–204. [Google Scholar] [CrossRef]
- Vieytes, M.R.; Cabado, A.G.; Alfonso, A.; Louzao, M.C.; Botana, A.M.; Botana, L.M. Solid-phase radioreceptor assay for paralytic shellfish toxins. Anal. Biochem. 1993, 211, 87–93. [Google Scholar] [CrossRef]
- Doucette, G.J.; Logan, M.M.; Ramsdell, J.S.; van Dolah, F.M. Development and preliminary validation of a microtiter plate-based receptor binding assay for paralytic shellfish poisoning toxins. Toxicon 1997, 35, 625–636. [Google Scholar] [CrossRef]
- Usup, G.; Leaw, C.-P.; Cheah, M.-Y.; Ahmad, A.; Ng, B.-K. Analysis of paralytic shellfish poisoning congeners by a sodium channel receptor binding assay. Toxicon 2004, 44, 37–43. [Google Scholar] [CrossRef]
- Van Dolan, F.M.; Fire, S.E.; Leighfield, T.A.; Mikulski, C.M.; Doucette, G.J. Determination of paralytic shellfish toxins in shellfish by Receptor Binding Assay: Collaborative study. J. AOAC Int. 2012, 95, 795–812. [Google Scholar] [CrossRef]
- Kogure, K.; Tamplin, M.L.; Simidu, U.; Colwell, R.R. A tissue culture assay for tetrodotoxin, saxitoxin, and related toxins. Toxicon 1988, 26, 191–197. [Google Scholar] [CrossRef]
- Jellett, J.F.; Marks, L.J.; Stewart, J.E.; Dorey, M.I.; Watson-Wright, W.; Lawrence, J.F. Paralytic shellfish poison (saxitoxin family) bioassays: Automated endpoint determination and stadardization of the in vitro tissue culture bioassay, and comparison with the standard mouse bioassay. Toxicon 1992, 30, 1143–1156. [Google Scholar] [CrossRef]
- Gallacher, S.; Birkbeck, T.H. A tissue culture assay for direct detection of sodium channel blocking toxins in bacterial culture supernatents. FEMS Microbiol. Lett. 1992, 92, 101–108. [Google Scholar] [CrossRef]
- Manger, M.L.; Lega, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Tetrazolium-based bioassay for neurotoxins active on voltage-sensitive sodium channels: Semiautomated assay for saxitoxins, brevetoxins, and ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef]
- Jellett, J.F.; Doucette, L.I.; Belland, E.R. The MIST (TM) shipable cell bioassay kits for PSP: An alternative to the mouse bioassay. J. Shellfish Res. 1998, 17, 1653–1655. [Google Scholar]
- Nicholson, R.A.; Li, G.H.; Buenaventura, E.; Graham, D. A rapid and sensitive assay for paralytic shellfish poison (PSP) toxins using mouse brain synaptoneurosomes. Toxicon 2002, 40, 831–838. [Google Scholar] [CrossRef]
- Louzao, M.C.; Vieytes, M.R.; Cabado, A.G.; de Sousa, J.; Botana, L.M. A fluorimetric microplate assay for detection and quantitation of toxins causing paralytic shellfish poisoning. Chem. Res. Toxicol. 2003, 16, 433–438. [Google Scholar] [CrossRef]
- Manger, R.; Woodle, D.; Berger, A.; Hungerford, J. Flow cytometric detection of saxitoxins using fluorescent voltage-sensitive dyes. Anal. Biochem. 2007, 366, 149–155. [Google Scholar] [CrossRef]
- Usleber, E.; Schneider, E.; Terplan, G. Direct enzyme immunoassay in microtitration plate and test strip format for the detection of saxitoxin in shellfish. Lett. Appl. Microbiol. 1991, 13, 275–277. [Google Scholar] [CrossRef]
- Chu, F.S.; Huang, X.; Hall, S. Production and characterization of antibodies against neosaxitoxin. J. AOAC Int. 1992, 75, 341–345. [Google Scholar]
- Burk, C.; Usleber, E.; Dietrich, R.; Martlbauer, E. Production and characterization of antibodies against neosaxitoxin utilizing a novel immunogen synthesis procedure. Food Agric. Immunol. 1995, 7, 315–322. [Google Scholar] [CrossRef]
- Micheli, L.; di Stefano, S.; Moscone, D.; Palleschi, G.; Marini, S.; Coletta, M.; Draisci, R.; Quadri, F.D. Production of antibodies and development of highly sensitive formats of enzyme immunoassay for saxitoxin analysis. Anal. Bioanal. Chem. 2002, 373, 678–684. [Google Scholar] [CrossRef]
- Huang, X.; Hsu, K.H.; Chu, F.S. Direct competitive enzyme-linked immunosorbent assay for saxitoxin and neosaxitoxin. J. Agric. Food Chem. 1996, 44, 1029–1035. [Google Scholar] [CrossRef]
- Kawatsu, K.; Hamano, Y.; Sugiyama, A.; Hashizume, K.; Noguchi, T. Development and application of an enzyme immunoassay based on a monoclonal antibody against gonyautoxin components of paralytic shellfish poisoning toxins. J. Food Prot. 2002, 65, 1304–1308. [Google Scholar]
- Dubois, M.; Demoulin, L.; Charlier, C.; Singh, G.; Godefroy, S.B.; Campbell, K.; Elliott, C.T.; Delahaut, P. Development of ELISAs for detecting domoic acid, okadaic acid, and saxitoxin and their applicability for the detection of marine toxins in samples collected in Belgium. Food Addit. Contam. Part A 2010, 27, 859–868. [Google Scholar] [CrossRef]
- Garneau, M.E.; Schnetzer, A.; Countway, P.D.; Jones, A.C.; Seubert, E.L.; Caron, D.A. Examination of the seasonal dynamics of the toxic dinoflagellate Alexandrium catenella at Redondo Beach, California, by quantitative PCR. Appl. Environ. Microbiol. 2011, 77, 7669–7680. [Google Scholar]
- Murray, S.A.; Wiese, M.; Stuken, A.; Brett, S.; Kellmann, R.; Hallegraeff, G.; Neilan, B.A. SxtA-based quantitative molecular assay to identify saxitoxin-producing harmful algal blooms in marine waters. Appl. Environ. Microbiol. 2011, 77, 7050–7057. [Google Scholar] [CrossRef]
- Al-Tebrineh, J.; Pearson, L.A.; Yasar, S.A.; Neilan, B.A. A multiplex qPCR targeting hepato- and neurotoxigenic cyanobacteria of global significance. Harmful Algae 2012, 15, 19–25. [Google Scholar] [CrossRef]
- Nagai, S.; Itakura, S. Specific detection of the toxic dinoflagellates Alexandrium tamarense and Alexandrium catenella from single vegetative cells by a loop-mediated isothermal amplification method. Mar. Genomics 2012, 7, 43–49. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonezawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucl. Acids Res. 2000, 28, 63. [Google Scholar] [CrossRef]
- Mori, Y.; Nagamine, K.; Tomita, N.; Notomi, T. Detection of loop-mediated isothermal amplification reaction by turbidity derived from magnesium pyrophosphate formation. Biochem. Biophys. Res. Commun. 2001, 289, 150–154. [Google Scholar] [CrossRef]
- Enosawa, M.; Kageyama, S.; Sawai, K.; Watanabe, K.; Notomi, T.; Onoe, S.; Mori, Y.; Yokomizo, Y. Use of loop-mediated isothermal amplification of the IS900 sequence for rapid detection of cultured Mycobacterium avium subsp. paratuberculosis. J. Clin. Microbiol. 2003, 41, 4359–4365. [Google Scholar] [CrossRef]
- Tang, X.H.; Yu, R.C.; Zhou, M.J.; Yu, Z.G. Application of rRNA probes and fluorescence in situ hybridization for rapid detection of the toxic dinoflagellate Alexandrium minutum. Chin. J. Ocean. Limnol. 2012, 30, 256–263. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, K.; De Salas, M. DNA probes, targeting large sub-unit rRNA, for the rapid identification of the paralytic shellfish poison producing dinoflagellate, Gymnodinium catenatum. New Z. J. Mar. Freshw. Res. 2007, 41, 385–390. [Google Scholar] [CrossRef]
- Wyatt, T.; Jenkinson, I.R. Notes on Alexandrium population dynamics. J. Plankton Res. 1997, 19, 551–575. [Google Scholar] [CrossRef]
- Ritson-Williams, R.; Yotsu-Yamashita, M.; Paul, V.J. Ecological functions of tetrodotoxin in a deadly polyclad flatworm. Proc. Natl. Acad. Sci. USA 2006, 103, 3176–3179. [Google Scholar] [CrossRef]
- Cembella, A.D. Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar] [CrossRef]
- Anderson, D.M.; Cheng, T.P.-O. Intracellular localization of saxitoxins in the dinofllagellate Gonyaulax tamarensis. J. Phycol. 1988, 24, 17–22. [Google Scholar] [CrossRef]
- Cembella, A.D. Ecophysiology and Metabolism of Paralytic Shellfish Toxins in Marine Microalgae. In Physiological Ecology of Harmful Algal Blooms; Anderson, D.M., Cembella, A.D., Hallegraeff, G., Eds.; Springer-Verlag: Berlin, Germany, 1998; pp. 381–403. [Google Scholar]
- Zimmer, R.K.; Ferrer, R.P. Neuroecology, chemical defense, and the keystone species concept. Biol. Bull. 2007, 213, 208–225. [Google Scholar] [CrossRef]
- Oikawa, H.; Satomi, M.; Watabe, S.; Yano, Y. Accumulation and depuration rates of paralytic shellfish poisoning toxins in the shore crab Telmessus acutidens by feeding toxic mussels under laboratory controlled conditions. Toxicon 2005, 45, 163–169. [Google Scholar] [CrossRef]
- Robineau, B.; Gagné, J.A.; Fortier, L.; Cembella, A.D. Potential impact of a toxic dinoflagellate (Alexandrium excavatum) bloom on survival of fish and crustacean larvae. Mar. Biol. 1991, 108, 293–301. [Google Scholar] [CrossRef]
- Selander, E.; Thor, P.; Toth, G.; Pavia, H. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc. R. Soc. B 2006, 273, 1673–1680. [Google Scholar] [CrossRef]
- Bergkvist, J.; Selander, E.; Pavia, H. Induction of toxin production in dinoflagellates: The grazer makes a difference. Oecologia 2008, 156, 147–154. [Google Scholar] [CrossRef]
- Teegarden, G.J. Copepod grazing selection and particle discrimination on the basis of PSP toxin content. Mar. Ecol. Prog. Ser. 1999, 181, 163–176. [Google Scholar] [CrossRef]
- Da Costa, R.M.; Franco, J.; Cacho, E.; Fernandez, F. Toxin content and toxic effects of the dinoflagellate Gyrodinium corsicum (Paulmier) on the ingestion and survival rates of the copepods Acartia grani and Euterpina acutifrons. J. Exp. Mar. Biol. Ecol. 2005, 322, 177–183. [Google Scholar] [CrossRef]
- Frangoulos, M.; Guisande, C.; Maneiro, I.; Riveiro, I.; Franco, J. Short-term and long-term effects of the toxic dinoflagellate Alexandrium minutum on the copepod Acartia clausi. Mar. Ecol. Prog. Ser. 2000, 203, 161–169. [Google Scholar] [CrossRef]
- Barreiro, A.; Guisande, C.; Frangopulos, M.; Gonzalez-Fernandez, A.; Munoz, S.; Perez, D.; Magadan, S.; Maneiro, I.; Riveiro, I.; Iglesias, P. Feeding strategies of the copepod Acartia clausi on single and mixed diets of toxic and non-toxic strains of the dinoflagellate Alexandrium minutum. Mar. Ecol. Prog. Ser. 2006, 316, 115–125. [Google Scholar] [CrossRef]
- Turner, J.T.; Tester, P.A. Toxic marine phytoplankton, zooplankton grazers, and pelagic food webs. Limnol. Oceanogr. 1997, 42, 1203–1214. [Google Scholar] [CrossRef]
- Solé, J.; Estrada, M.; Garcia-Ladona, E. Biological control of harmful algal blooms: A modelling study. J. Mar. Syst. 2006, 61, 165–179. [Google Scholar] [CrossRef]
- Tillmann, U.; John, U. Toxic effects of Alexandrium spp. on heterotrophic dinoflagellates: An allelochemical defence mechanism independent of PSP-toxin content. Mar. Ecol. Prog. Ser. 2006, 61, 47–59. [Google Scholar]
- Juhl, A.R.; Martins, C.A.; Anderson, D.M. Toxicity of Alexandrium lusitanicum to gastropod larvae is not caused by paralytic-shellfish-poisoning toxins. Harmful Algae 2008, 7, 567–573. [Google Scholar] [CrossRef]
- Ianora, A.; Bentley, M.G.; Caldwell, G.S.; Casotti, R.; Cembella, A.D.; Engstrom-Ost, J.; Halsband, C.; Sonnenschein, E.; Legrand, C.; Llewellyn, C.A.; et al. The relevance of marine chemical ecology to plankton and ecosystem function: An emerging field. Mar. Drugs 2011, 9, 1625–1648. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cusick, K.D.; Sayler, G.S. An Overview on the Marine Neurotoxin, Saxitoxin: Genetics, Molecular Targets, Methods of Detection and Ecological Functions. Mar. Drugs 2013, 11, 991-1018. https://doi.org/10.3390/md11040991
Cusick KD, Sayler GS. An Overview on the Marine Neurotoxin, Saxitoxin: Genetics, Molecular Targets, Methods of Detection and Ecological Functions. Marine Drugs. 2013; 11(4):991-1018. https://doi.org/10.3390/md11040991
Chicago/Turabian StyleCusick, Kathleen D., and Gary S. Sayler. 2013. "An Overview on the Marine Neurotoxin, Saxitoxin: Genetics, Molecular Targets, Methods of Detection and Ecological Functions" Marine Drugs 11, no. 4: 991-1018. https://doi.org/10.3390/md11040991
APA StyleCusick, K. D., & Sayler, G. S. (2013). An Overview on the Marine Neurotoxin, Saxitoxin: Genetics, Molecular Targets, Methods of Detection and Ecological Functions. Marine Drugs, 11(4), 991-1018. https://doi.org/10.3390/md11040991