Biosynthesis of Polyunsaturated Fatty Acids in Marine Invertebrates: Recent Advances in Molecular Mechanisms


{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Pathways of PUFA Synthesis in Primary Producers
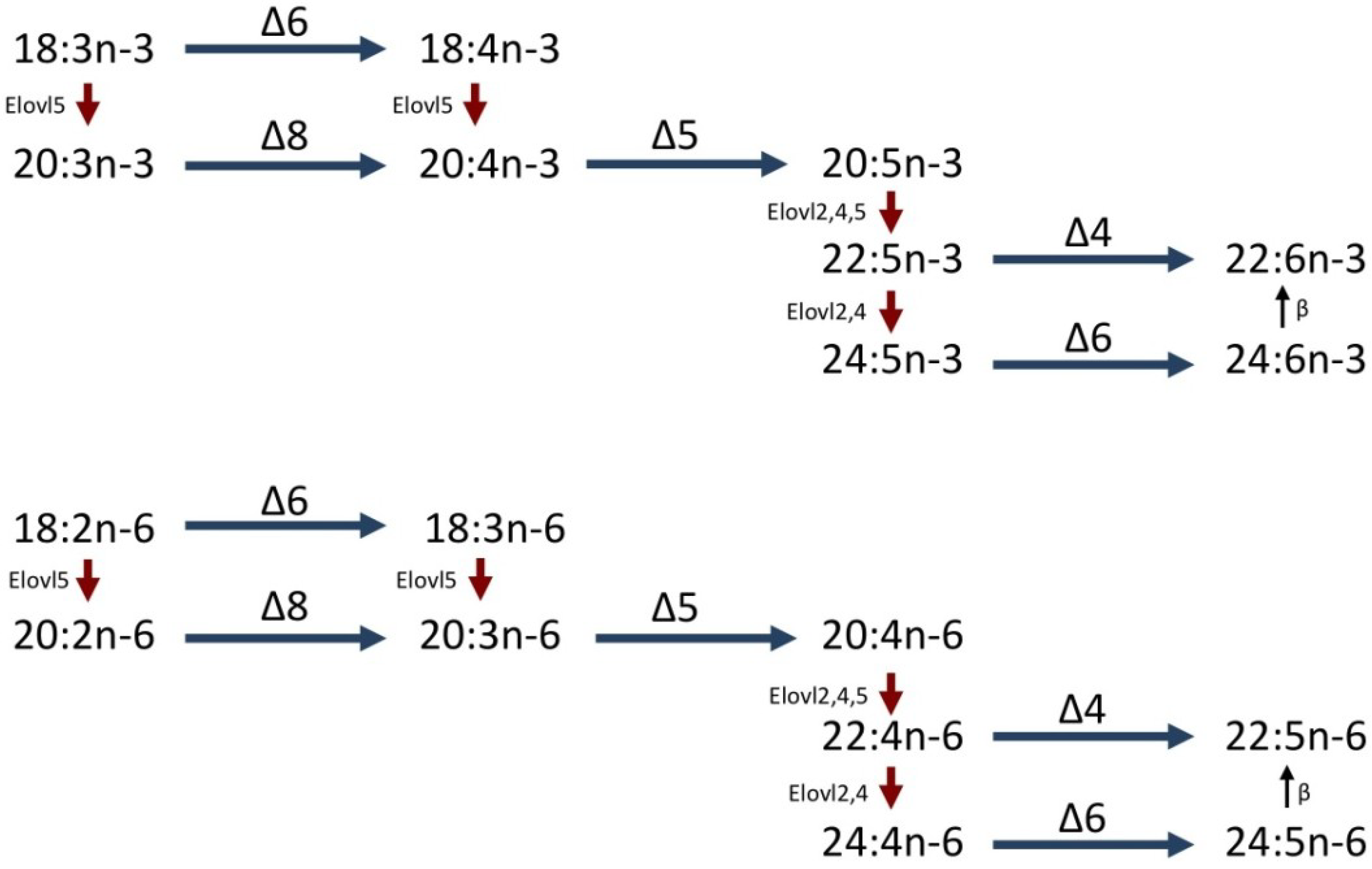
1.2. LC-PUFA Biosynthesis in Fish
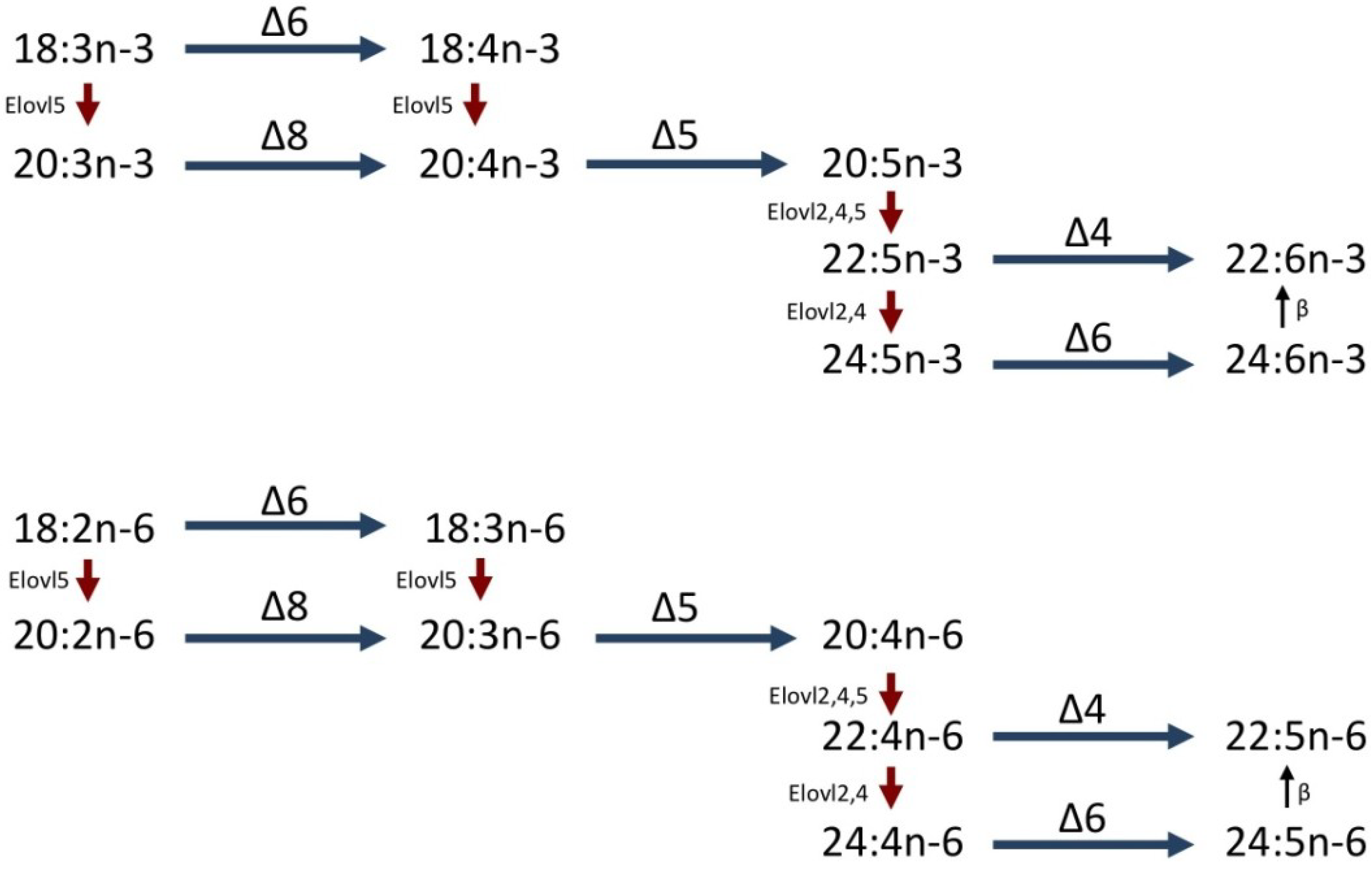
1.3. PUFA Production in Marine Invertebrates
2. Molluscs Can Biosynthesize PUFA
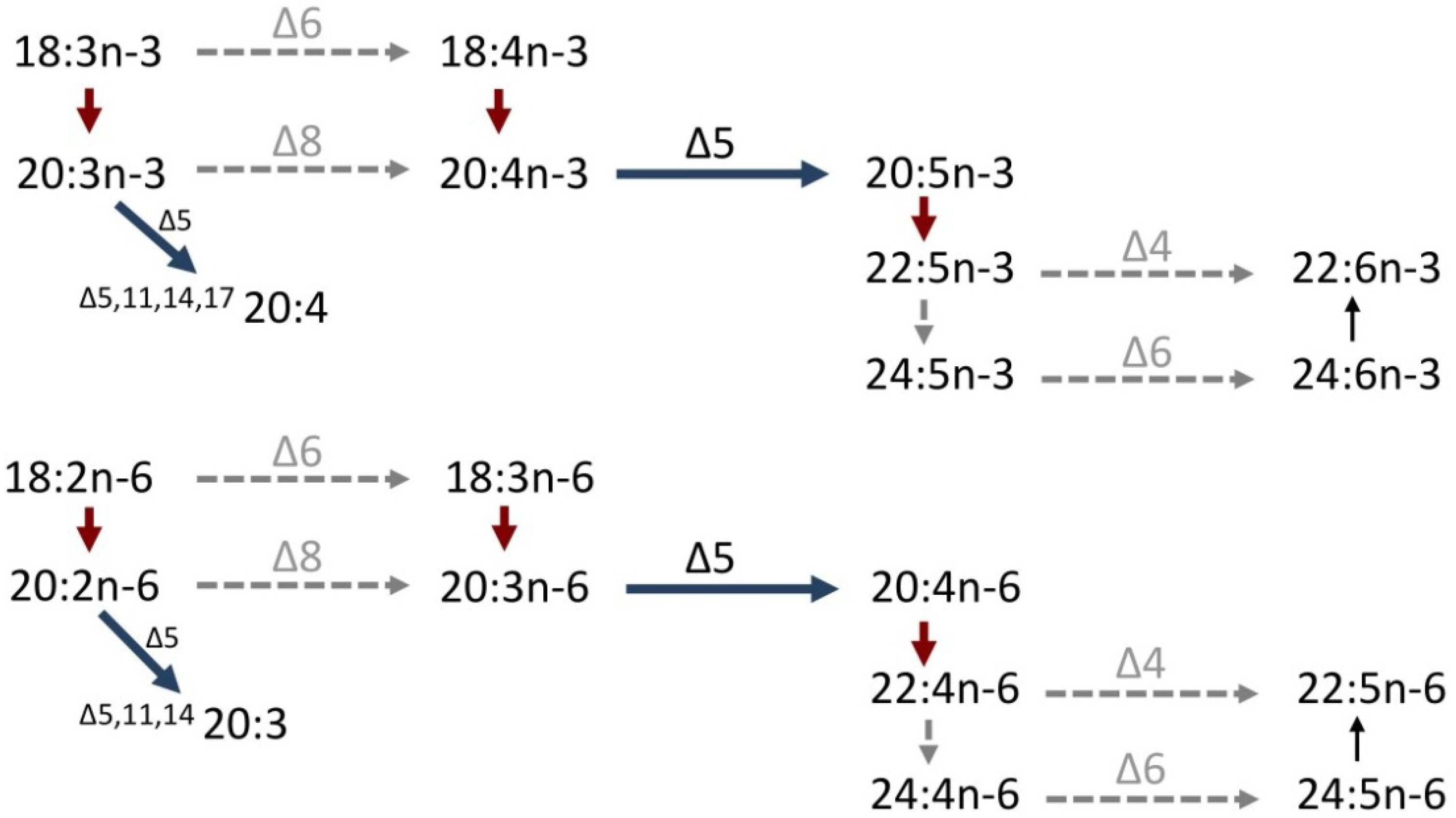
3. Molecular Studies on the Biosynthesis of PUFA in Cephalopods
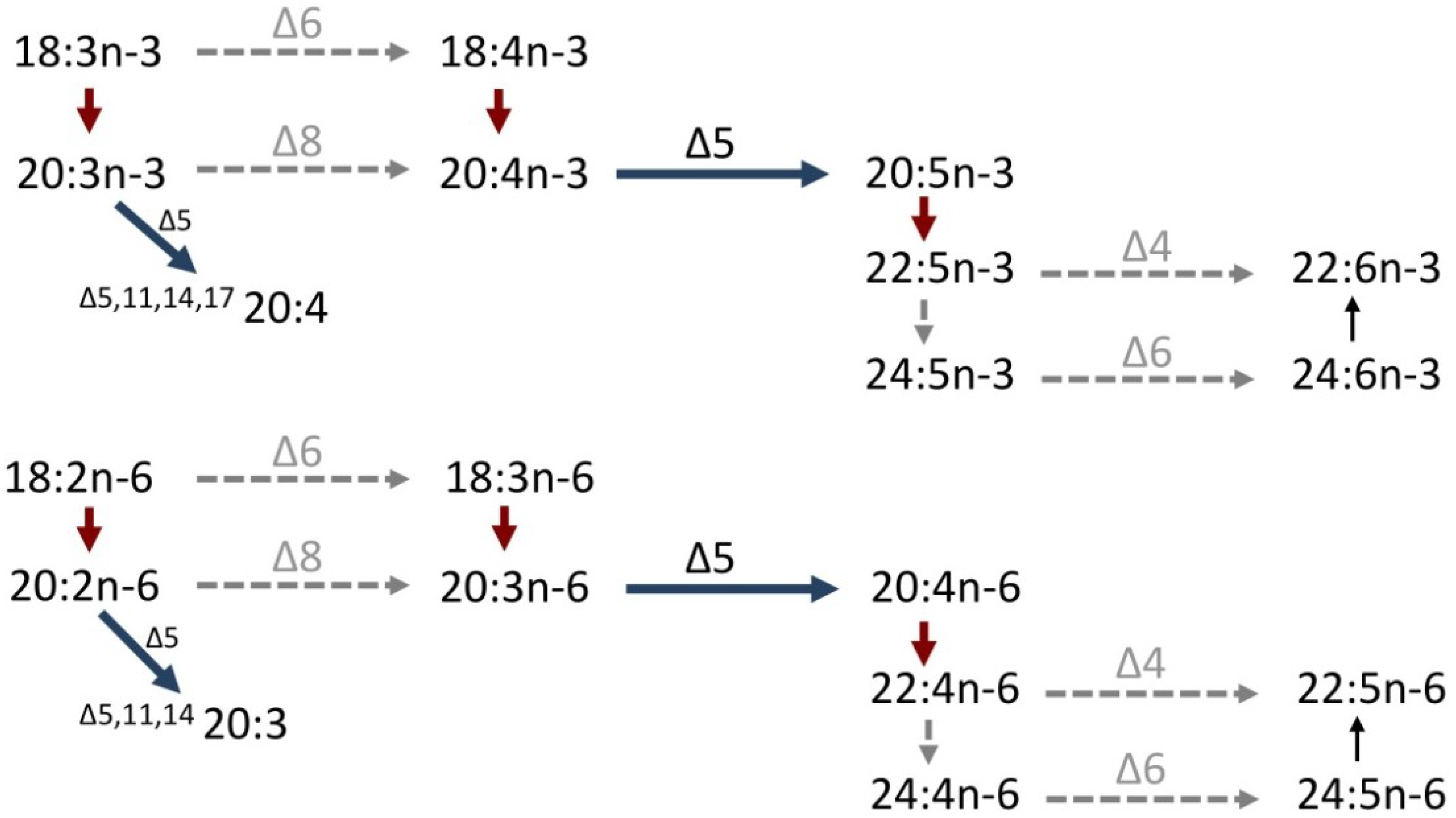
4. Molecular Mechanisms of Non-Methylene-Interrupted Fatty Acid Biosynthesis
5. Biosynthesis of PUFA in Other Marine Invertebrates
5.1. Sponges
5.2. Crustaceans
5.3. Cnidarians
5.4. Other Non-vertebrate Groups
6. Concluding Remarks
Acknowledgments
Conflicts of Interest
References
- Pereira, H.; Barreira, L.; Figueiredo, F.; Custódio, L.; Vizetto-Duarte, C.; Polo, C.; Rešek, E.; Engelen, A.; Varela, J. Polyunsaturated fatty acids of marine macroalgae: Potential for nutritional and pharmaceutical applications. Mar. Drugs 2012, 10, 1920–1935. [Google Scholar] [CrossRef]
- Tur, J.A.; Bibiloni, M.M.; Sureda, A.; Pons, A. Dietary sources of omega 3 fatty acids: Public health risks and benefits. Br. J. Nutr. 2012, 107, S23–S52. [Google Scholar] [CrossRef]
- Ahlgren, G.; Vrede, T.; Goedkoop, W. Fatty acid ratios in freshwater fish, zooplankton and zoobenthos. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M., Kainz, M., Eds.; Springer-Verlag: New York, NY, USA, 2009; pp. 147–178. [Google Scholar]
- Brett, M.T.; Muller-Navarra, D.C.; Persson, J. Crustacean zooplankton fatty acid composition. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M., Kainz, M., Eds.; Springer-Verlag: New York, NY, USA, 2009; pp. 115–146. [Google Scholar]
- Desvilettes, C.; Bec, A. Formation and transfer of fatty acids in aquatic microbial food webs: Role of heterotrophic protists. In Lipids in Aquatic Ecosystems; Arts, M.T., Brett, M., Kainz, M., Eds.; Springer-Verlag: New York, NY, USA, 2009; pp. 25–42. [Google Scholar]
- Guschina, I.A.; Harwood, J.L. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res. 2006, 45, 160–186. [Google Scholar] [CrossRef]
- Bell, M.V.; Dick, J.R.; Pond, D.W. Octadecapentaenoic acid in a raphidophyte alga, Heterosigma akashiwo. Phytochemistry 1997, 45, 303–306. [Google Scholar] [CrossRef]
- Metz, J.G.; Roessler, P.; Facciotti, D.; Levering, C.; Dittrich, F.; Lassner, M.; Valentine, R.; Lardizabel, K.; Domergue, F.; Yamada, A.; et al. Production of polyunsaturated fatty acids by polyketide synthases in both prokaryotes and eukaryotes. Science 2001, 293, 290–293. [Google Scholar] [CrossRef]
- Russell, N.J.; Nichols, D.S. Polyunsaturated fatty acids in marine bacteria—a dogma rewritten. Microbiology 1999, 145, 765–779. [Google Scholar]
- Barclay, W.R.; Meager, K.M.; Abril, J.R. Heterotrophic production of long chain omega-3 fatty acids utilizing algae and algae-like microorganisms. J. Appl. Phycol. 1994, 6, 123–129. [Google Scholar] [CrossRef]
- Hauvermale, A.; Kuner, J.; Rosenzweig, B.; Guerra, D.; Diltz, S.; Metz, J.G. Fatty acid production in Schizochytrium sp.: Involvement of a polyunsaturated fatty acid synthase and a type 1 fatty acid synthase. Lipids 2006, 41, 739–747. [Google Scholar] [CrossRef]
- Qiu, X.; Hong, H.P.; Mackenzie, S.L. Identification of a Δ4 fatty acid desaturase from Thraustochytrium sp. involved in biosynthesis of docosahexaenoic acid by heterologous expression in Saccharomyces cervisiae and Brassica juncea. J. Biol. Chem. 2001, 276, 31561–31566. [Google Scholar] [CrossRef]
- Bell, M.V.; Tocher, D.R. Biosynthesis of fatty acids; general principles and new directions. Arts, M.T., Brett, M., Kainz, M., Eds.; Springer-Verlag: New York, NY, USA, 2009; pp. 211–236. [Google Scholar]
- Mansour, M.P.; Volkman, J.K.; Holdsworth, D.G.; Jackson, A.E.; Blackburn, S.I. Very-long chain (C28) highly unsaturated fatty acids in marine dinoflagellates. Phytochemistry 1999, 50, 541–548. [Google Scholar] [CrossRef]
- Harwood, J.L.; Guschina, I.A. The versatility of algae and their lipid metabolism. Biochimie 2009, 91, 679–684. [Google Scholar] [CrossRef]
- Jøstensen, J.P.; Landfald, B. High prevalence of polyunsaturated-fatty-acid producing bacteria in Arctic invertebrates. FEMS Microbiol. Lett. 1997, 151, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Vrinten, P.; Mavraganis, I.; Qiu, X.; Senger, T. Biosynthesis of long chain polyunsaturated fatty acids in the marine ichthyosporean Sphaeroforma arctica. Lipids 2013, 48, 263–274. [Google Scholar] [CrossRef]
- Tocher, D.R. Metabolism and functions of lipids and fatty acids in teleost fish. Rev. Fish. Sci. 2003, 11, 107–184. [Google Scholar] [CrossRef]
- Cook, H.W.; McMaster, R.C.R. Fatty acid desaturation and chain elongation in eukaryotes. In Biochemistry of Lipids, Lipoproteins and Membranes; Vance, D.E., Vance, J.E., Eds.; Elsevier: Amsterdam, The Netherlands, 2004; pp. 181–204. [Google Scholar]
- Monroig, Ó.; Li, Y.; Tocher, D.R. Delta-8 desaturation activity varies among fatty acyl desaturases of teleost fish: high activity in delta-6 desaturases of marine species. Comp. Biochem. Physiol. Part B 2011, 159, 206–213. [Google Scholar] [CrossRef]
- Sprecher, H. Metabolism of highly unsaturated n-3 and n-6 fatty acids. Biochim. Biophys. Acta 2000, 1486, 219–231. [Google Scholar] [CrossRef]
- Buzzi, M.; Henderson, R.J.; Sargent, J.R. Biosynthesis of docosahexaenoic acid in trout hepatocytes proceeds via 24-carbon intermediates. Comp. Biochem. Physiol. Part B 1997, 116, 263–267. [Google Scholar] [CrossRef]
- Li, Y.; Monroig, Ó.; Zhang, L.; Wang, S.; Zheng, X.; Dick, J.R.; You, C.; Tocher, D.R. Vertebrate fatty acyl desaturase with Δ4 activity. Proc. Natl. Acad. Sci. USA 2010, 107, 16840–16845. [Google Scholar]
- Morais, S.; Castanheira, F.; Martínez-Rubio, L.; Conceição, L.E.C.; Tocher, D.R. Long-chain polyunsaturated fatty acid synthesis in a marine vertebrate: Ontogenetic and nutritional regulation of a fatty acyl desaturase with Δ4 activity. Biochim. Biophys. Acta 2012, 1821, 660–671. [Google Scholar] [CrossRef]
- Tocher, D.R. Fatty acid requirements in ontogeny of marine and freshwater fish. Aquacul. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Hastings, N.; Agaba, M.K.; Tocher, D.R.; Zheng, X.; Dickson, C.A.; Dick, J.R.; Teale, A.J. Molecular cloning and functional characterization of fatty acyl desaturase and elongase cDNAs involved in the production of eicosapentaenoic and docosahexaenoic acids from α-linolenic acid in Atlantic salmon (Salmo salar). Mar. Biotechnol. 2005, 6, 463–474. [Google Scholar]
- Hastings, N.; Agaba, M.; Tocher, DR.; Leaver, M.J.; Dick, J.R.; Sargent, J.R.; Teale, A.J. A vertebrate fatty acid desaturase with Δ5 and Δ6 activities. Proc. Natl. Acad. Sci. USA 2001, 98, 14304–14309. [Google Scholar] [CrossRef]
- Leaver, M.J.; Bautista, J.M.; Björnsson, T.; Jönsson, E.; Krey, G.; Tocher, D.R.; Torstensen, B.E. Towards fish lipid nutrigenomics: Current state and prospects for fin-fish aquaculture. Rev. Fish. Sci. 2008, 16, 71–92. [Google Scholar]
- Jakobsson, A.; Westerberg, R.; Jacobsson, A. Fatty acid elongases in mammals: Their regulation and roles in metabolism. Prog. Lipid Res. 2006, 45, 237–249. [Google Scholar] [CrossRef]
- Agbaga, M.P.; Brush, R.S.; Mandal, M.N.A.; Henry, K.; Elliott, M.H.; Anderson, R.E. Role of Stargardt-3 macular dystrophy protein (ELOVL4) in the biosynthesis of very long chain fatty acids. Proc. Nat. Acad. Sci. USA 2008, 105, 12843–12848. [Google Scholar]
- Monroig, Ó.; Webb, K.; Ibarra-Castro, L.; Holt, G.J.; Tocher, D.R. Biosynthesis of long-chain polyunsaturated fatty acids in marine fish: Characterisation of an Elovl4-like elongase from cobia Rachycentron canadum and activation of the pathway during early life stages. Aquaculture 2011, 312, 145–153. [Google Scholar] [CrossRef]
- Monroig, Ó.; Navarro, J.C.; Tocher, D.R. Long-chain polyunsaturated fatty acids in fish: Recent advances on desaturases and elongases involved in their biosynthesis. In Proceedings of the XI International Symposium on Aquaculture Nutrition; Cruz-Suarez, L.E., Ricque-Marie, D., Tapia-Salazar, M., Nieto-López, M.G., Villarreal-Cavazos, D.A., Gamboa-Delgado, J., Hernández-Hernández, L.H., Eds.; Universidad Autónoma de Nuevo León, Monterrey: Nuevo León, México, 2011; pp. 257–282. [Google Scholar]
- Venegas-Calerón, M.; Sayanova, O.; Napier, J.A. An alternative to fish oils: Metabolic engineering of oil-seed crops to produce omega-3 long chain polyunsaturated fatty acids. Prog. Lipid Res. 2010, 49, 108–119. [Google Scholar] [CrossRef]
- Lu, C.; Napier, J.A.; Clemente, T.E.; Cahoon, E.B. New frontiers in oilseed biotechnology: Meeting the global demand for vegetable oils for food, feed, biofuel, and industrial applications. Curr. Opin. Biotechnol. 2011, 22, 252–259. [Google Scholar] [CrossRef]
- Pereira, S.L.; Leonard, A.E.; Mukerji, P. Recent advances in the study of fatty acid desaturases from animals and lower eukaryotes. Prostag. Leukotr. Ess. 2003, 68, 97–106. [Google Scholar] [CrossRef]
- Joseph, J.D. Lipid composition of marine and estuarine invertebrates. Part II: Mollusca. Prog. Lipid Res. 1982, 21, 109–153. [Google Scholar] [CrossRef]
- Joseph, J.D. Distribution and composition of lipids in marine invertebrates. In Marine Biogenic Lipids, Fats, and Oils. Vol. II; Ackman, R.G., Ed.; CRC Press, Inc.: Boca Raton, Florida, FL, USA, 1989; pp. 49–143. [Google Scholar]
- van der Horst, D.J. Biosynthesis of saturated and unsaturated fatty acids in the pulmonate land snail Cepaea nemoralis (L.). Comp. Biochem. Physiol. Part B 1973, 46, 551–560. [Google Scholar]
- van der Horst, D.J. In vivo biosynthesis of fatty acids in the pulmonate land snail Cepaea nemoralis (L.) under anoxic conditions. Comp. Biochem. Physiol. Part B 1974, 47, 181–187. [Google Scholar] [CrossRef]
- Weinert, J.; Blomquist, G.J.; Borgeson, C.E. De novo biosynthesis of linoleic acid in two non-insect invertebrates: The land slug and the garden snail. Experientia 1993, 49, 919–921. [Google Scholar] [CrossRef]
- Zhu, N.; Dai, X.; Lin, D.S.; Connor, W.E. The lipids of slugs and snails: Evolution, diet and biosynthesis. Lipids 1994, 29, 869–875. [Google Scholar] [CrossRef]
- De Moreno, J.E.A.; Moreno, V.J.; Brenner, R.R. Lipid metabolism of the yellow clam, Mesodesma mactroides: 2-polyunsaturated fatty acid metabolism. Lipids 1976, 11, 561–566. [Google Scholar] [CrossRef]
- Waldock, M.J.; Holland, D.L. Fatty acid metabolism in young oysters, Crassostrea gigas: Polyunsaturated fatty acids. Lipids 1984, 19, 332–336. [Google Scholar] [CrossRef]
- Pirini, M.; Manuzzi, M.P.; Pagliarani, A.; Trombetti, F.; Borgatti, A.R.; Ventrella, V. Changes in fatty acid composition of Mytilus galloprovincialis (Lmk) fed on microalgal and wheat germ diets. Comp. Biochem. Physiol. Part B 2007, 147, 616–626. [Google Scholar]
- Zhukova, N.V. Biosynthesis of non-methylene-interrupted dienoic fatty acids for [C-14] acetate in mollusks. Biochim. Biophys. Acta 1986, 878, 131–133. [Google Scholar] [CrossRef]
- Zhukova, N.V. The pathway of the biosynthesis of non-methylene-interrupted dienoic fatty acids in mollusks. Comp. Biochem. Physiol. Part B 1991, 100, 801–804. [Google Scholar] [CrossRef]
- Kornprobst, J.M.; Barnathan, G. Demospongic acids revisited. Mar. Drugs 2010, 8, 2569–2577. [Google Scholar] [CrossRef]
- Barnathan, G. Non-methylene-interrupted fatty acids from marine invertebrates: Occurrence, characterization and biological properties. Biochimie 2009, 91, 671–678. [Google Scholar] [CrossRef]
- Castro, L.F.C.; Wilson, J.M.; Gonçalves, O.; Galante-Oliveira, S.; Rocha, E.; Cunha, I. The evolutionary history of the stearoyl-CoA desaturase gene family in vertebrates. BMC Evol. 2011, 11, 132. [Google Scholar] [CrossRef]
- David, E.; Tanguy, A.; Pichavant, K.; Moraga, D. Response of the Pacific oyster Crassostrea gigas to hypoxia exposure under experimental conditions. FEBS J. 2005, 272, 5635–5652. [Google Scholar] [CrossRef]
- David, E.; Boudry, R.; Degremont, L.; Tanguy, A.; Quéré, N.; Samain, J.-F.; Moraga, D. Genetic polymorphism of glutamine synthetase and delta-9 desaturase in families of Pacific oyster Crassostrea gigas and susceptibility to summer mortality. J. Exp. Mar. Biol. Ecol. 2007, 349, 272–283. [Google Scholar] [CrossRef]
- David, E.; Tanguy, A.; Riso, R.; Quiniou, L.; Laroche, J.; Moraga, D. Responses of Pacific oyster Crassostrea gigas populations to abiotic stress in environmentally contrasted estuaries along the Atlantic coast of France. Aquat Toxicol. 2012, 109, 70–79. [Google Scholar] [CrossRef]
- Guillou, H.; Zadravec, D.; Martin, P.G.P.; Jacobsson, A. The key roles of elongases and desaturases in mammalian fatty acid metabolism: insights from transgenic mice. Prog. Lipid Res. 2010, 49, 186–199. [Google Scholar] [CrossRef]
- Li, M.; Mai, K.; Ai, Q.; He, G.; Xu, W.; Zhang, W.; Zhang, Y.; Zhou, H. Effects of dietary grape seed oil and linseed oil on growth, muscle fatty acid composition and expression of putative Δ5 fatty acyl desaturase in abalone Haliotis discus hannai Ino. Aquaculture 2013, 406–407, 105–114. [Google Scholar] [CrossRef]
- Li, M.; Mai, K.; He, G.; Ai, Q.; Zhang, W.; Xu, W.; Wang, J.; Liufu, Z.; Zhang, Y.; Zhou, H. Characterization of Δ5 fatty acyl desaturase in abalone Haliotis discus hannai Ino. Aquaculture 2013, 416–417, 48–56. [Google Scholar] [CrossRef]
- Mateos, H.T.; Lewandowski, P.A.; Su, X.Q. Dietary fish oil supplements increase tissue n-3 fatty acid composition and expression of Delta-6 desaturase and elongase-2 in Jade Tiger hybrid abalone. Lipids 2011, 46, 741–751. [Google Scholar] [CrossRef]
- Mateos, H.T.; Lewandowski, P.A.; Su, X.Q. Effects of dietary fish oil replacement with flaxseed oil on tissue fatty acid composition and expression of desaturase and elongase genes. J. Sci. Food Agric. 2012, 92, 418–426. [Google Scholar] [CrossRef]
- Mateos, H.T.; Lewandowski, P.A.; Su, X.Q. The effect of replacing dietary fish oil with canola oil on fatty acid composition and expression of desaturase and elongase genes in Jade Tiger hybrid abalone. Food Chem. 2012, 131, 1217–1222. [Google Scholar] [CrossRef]
- Durazo-Beltran, E.; D’Abramo, L.R.; Toro-Vazquez, J.F.; Vasquez-Pelaez, C; Viana, M.T. Effect of triacylglycerols in formulated diets on growth and fatty acid composition in tissue of green abalone (Haliotis fulgens). Aquaculture 2003, 224, 257–270. [Google Scholar] [CrossRef]
- Kawashima, H.; Ohnishi, M. Identification of minor fatty acids and various nonmethylene-interrupted diene isomers in mantle, muscle, and viscera of the marine bivalve Megangulus zyonoensis. Lipids 2004, 39, 265–271. [Google Scholar] [CrossRef]
- Monroig, Ó.; Monroig, Ó.; Guinot, D.; Hontoria, F.; Tocher, D.R.; Navarro, J.C. Biosynthesis of essential fatty acids in Octopus vulgaris (Cuvier, 1797): Molecular cloning, functional characterisation and tissue distribution of a fatty acyl elongase. Aquaculture 2012, 360–361, 45–53. [Google Scholar] [CrossRef]
- Monroig, Ó.; Navarro, J.C.; Dick, J.R.; Alemany, F.; Tocher, D.R. Identification of a Δ5-like fatty acyl desaturase from the cephalopod Octopus vulgaris (Cuvier 1797) involved in the biosynthesis of essential fatty acids. Mar. Biotechnol. 2012, 14, 411–422. [Google Scholar] [CrossRef]
- Navarro, J.C.; Villanueva, R. Lipid and fatty acid composition of early stages of cephalopods: an approach to their lipid requirements. Aquaculture 2000, 183, 161–177. [Google Scholar] [CrossRef]
- Iglesias, J.; Sánchez, F.J.; Bersano, J.G.F.; Carrasco, J.F.; Dhont, J.; Fuentes, L.; Linares, F.; Muñoz, J.L.; Okumura, S.; Roo, J.; et al. Rearing of Octopus vulgaris paralarvae: Present status, bottlenecks and trends. Aquaculture 2007, 266, 1–15. [Google Scholar] [CrossRef]
- Monroig, Ó.; Hontoria, F.; Varó, I.; Tocher, D.R.; Navarro, J.C. Investigating the essential fatty acid requirements of the cephalopod Sepia officinalis: A molecular approach. In Proceedings of Larvi 2013 6th Fishand Shellfish Symposium, Ghent, Belgium, 2–5 September 2013; pp. 286–289.
- Hashimoto, K.; Yoshizawa, A.C.; Okuda, S.; Kuma, K.; Goto, S.; Kanehisa, M. The repertoire of desaturases and elongases reveals fatty acid variations in 56 eukaryotic genomes. J. Lipid Res. 2008, 49, 183–191. [Google Scholar] [CrossRef]
- Monroig, Ó.; Rotllant, J.; Cerdá-Reverter, J.M.; Dick, J.R.; Figueras, A.; Tocher, D.R. Expression and role of Elovl4 elongases in biosynthesis of very long-chain fatty acids during zebrafish Danio rerio early embryonic development. Biochim. Biophys. Acta 2011, 1801, 1145–1154. [Google Scholar]
- Carmona-Antoñanzas, G.; Monroig, Ó.; Dick, J.R.; Davie, A.; Tocher, D.R. Biosynthesis of very long-chain fatty acids (C > 24) in Atlantic salmon: Cloning, functional characterisation, and tissue distribution of an Elovl4 elongase. Comp. Biochem. Physiol. Part B 2011, 159, 122–129. [Google Scholar]
- Monroig, Ó.; Wang, S.; Zhang, L.; You, C.; Tocher, D.R.; Li, Y. Elongation of long-chain fatty acids in rabbitfish Siganus canaliculatus: Cloning, functional characterisation and tissue distribution of Elovl5- and Elovl4-like elongases. Aquaculture 2012, 350–353, 63–70. [Google Scholar] [CrossRef]
- Zhukova, N.V. Lipid classes and fatty acid composition of the tropical nudibranch mollusks Chromodoris sp. and Phyllidia coelestis. Lipids 2007, 42, 1169–1175. [Google Scholar] [CrossRef]
- Klingensmith, J. Distribution of methylene and nonmethylene-interrupted dienoic fatty acids in polar lipids and triacylglycerols of selected tissues of the hardshell clam (Mercenaria mercenaria). Lipids 1982, 17, 976–981. [Google Scholar] [CrossRef]
- Ojea, J.; Pazos, A.J.; Martinez, D.; Novoa-Sánchez, S.J.L.; Abad, M. Seasonal variation in weight and biochemical composition of the tissues of Ruditapes decussatus in relation to the gametogenic cycle. Aquaculture 2004, 238, 451–468. [Google Scholar] [CrossRef]
- Litchfield, C.; Morales, R.W. Are demospongiae membranes unique among living organisms? In Aspects of Sponge Biology; Harrison, F.W., Cowden, R.R., Eds.; Academic Press: New York, NY, USA, 1976; pp. 183–200. [Google Scholar]
- Litchfield, C.; Tyszkiewicz, J.; Dato, V. 5,9,23-Triacontatrienoic acid, principal fatty acid of the marine sponge Chondrilla nucula. Lipids 1980, 15, 200–202. [Google Scholar] [CrossRef]
- Abad, M.; Ruiz, C.; Martinez, D.; Mosquera, G.; Sanchez, J.L. Seasonal variations of lipid classes and fatty acids in flat oysters, Ostrea edulis, from San Cibran (Galicia, Spain). Comp. Biochem. Physiol. Part C 1995, 110, 109–118. [Google Scholar]
- Monroig, Ó.; Varó, I.; Tocher, D.R.; Navarro, J.C. Isolation and functional characterisation of a stearoyl-CoA desaturase from the marine invertebrate Octopus vulgaris. Comp. Physiol. Biochem. Part A 2012, 163, S46–S47. [Google Scholar]
- Kawashima, H. Unusual minor nonmethylene-interrupted di-, tri-, and tetraenoic fatty acids in limpet gonads. Lipids 2005, 40, 627–630. [Google Scholar] [CrossRef]
- Rod’kina, S.A. Fatty acids and other lipids of marine sponges. Russ. J. Mar. Biol. 2005, 31, S49–S60. [Google Scholar] [CrossRef]
- Carballeira, N.M.; Maldonado, M.E. On the isolation of the new fatty acid 6,11-eicosadienoic (20:2) and related 6,11-dienoic acids from the sponge Euryspongia rosea. Lipids 1989, 24, 665–665. [Google Scholar] [CrossRef]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E. A and 28 other authors. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [Google Scholar] [CrossRef]
- Norsker, N.H.; Støttrup, J.G. The importance of dietary HUFAs for fecundity and HUFA content in the harpacticoid, Tisbe holothuriae Humes. Aquaculture 1994, 125, 155–166. [Google Scholar] [CrossRef]
- Nanton, D.A.; Castell, J.D. The effects of dietary fatty acids on the fatty acid composition of the harpacticoid copepod, Tisbe sp., for use in live food for marine fish larvae. Aquaculture 1998, 163, 251–261. [Google Scholar] [CrossRef]
- Støttrup, J.G. The elusive copepods: their production and suitability in marine aquaculture. Aquacult. Res. 2000, 31, 703–711. [Google Scholar] [CrossRef]
- Rhodes, A.; Boyd, L. Formulated feeds for harpacticoid copepods: implications for population growth and fatty acid composition. In Copepods in Aquaculture; Lee, C.S., O’Bryen, P.J., Marcus, N.H., Eds.; Blackwell Publishing: Oxford, UK, 2005; pp. 61–73. [Google Scholar]
- De Troch, M.; Boeckx, P.; Cnudde, C.; Van Gansbeke, D.; Vanreusel, A.; Vincx, M.; Caramujo, M.J. Bioconversion of fatty acids at the basis of marine food webs: insights from a compound-specific stable isotope analysis. Mar. Ecol. Prog. Ser. 2012, 465, 53–67. [Google Scholar] [CrossRef]
- Desvilettes, C.; Bourdier, G.; Breton, J.C. On the occurrence of a possible bioconversion of linolenic acid into docosahexaenoic acid by the copepod Eucyclops serrulatus fed on microalgae. J. Plankton Res. 1997, 19, 273–278. [Google Scholar] [CrossRef]
- Parrish, C.C.; French, V.M.; Whiticar, M.J. Lipid class and fatty acid composition of copepods (Calanus finmarchicus, C. glacialis, Pseudocalanus sp., Tisbe furcata and Nitokra lacustris) fed various combinations of autotrophic and heterotrophic protists. J. Plankton Res. 2012, 34, 356–375. [Google Scholar] [CrossRef]
- Bell, M.V.; Dick, J.R.; Anderson, T.R.; Pond, D.W. Application of liposome and stable isotope tracer techniques to study polyunsaturated fatty acid biosynthesis in marine zooplankton. J. Plankton Res. 2007, 29, 417–422. [Google Scholar] [CrossRef]
- Guo, Z.; Yang, Z.; Cheng, Y.; Ji, L.; Que, Y.; Liu, Z.; Zeng, Q. Molecular characterization, tissue expression of acyl-CoA Delta 9-desaturase-like gene, and effects of dietary lipid levels on its expression in the hepatopancreas of the Chinese mitten crab (Eriocheir sinensis). Aquaculture 2013, 402, 58–65. [Google Scholar]
- Yang, Z.; Guo, Z.; Ji, L.; Zeng, Q.; Wang, Y.; Yang, X.; Cheng, Y. Cloning and tissue distribution of a fatty acyl Δ6-desaturase-like gene and effects of dietary lipid levels on its expression in the hepatopancreas of Chinese mitten crab (Eriocheir sinensis). Comp. Biochem. Physiol. Part B 2013, 165, 99–105. [Google Scholar] [CrossRef]
- Schlechtriem, C.; Arts, M.T.; Zellmer, I.D. Effect of temperature on the fatty acid composition and temporal trajectories of fatty acids in fasting Daphnia pulex (Crustacea, Cladocera). Lipids 2006, 41, 397–400. [Google Scholar] [CrossRef]
- Schauer, P.S.; Simpson, K.L. Bioaccumulation and bioconversion of dietary labeled fatty acid in Artemia and winter flounder (Pseudopleurunectes americanus). Can. J. Fish. Aquat. Sci. 1985, 42, 1430–1438. [Google Scholar] [CrossRef]
- Ito, M.K.; Simpson, K.L. The biosynthesis of omega 3 fatty acids from 18:2 omega 6 in Artemia spp. Comp. Biochem. Physiol. Part B 1996, 115, 69–76. [Google Scholar] [CrossRef]
- Spychalla, J.P.; Kinney, A.J.; Browse, J. Identification of an animal omega-3 fatty acid desaturase by heterologous expression in Arabidopsis. Proc. Natl. Acad. Sci. USA 1997, 94, 1142–1147. [Google Scholar] [CrossRef]
- Zhou, X.R.; Green, A.G.; Singh, S.P. Caenorhabditis elegans Δ12-desaturase FAT-2 is a bifunctional desaturase able to desaturate a diverse range of fatty acid substrates at the Δ12 and Δ15 positions. J. Biol. Chem. 2011, 286, 43644–43650. [Google Scholar] [CrossRef]
- Douglas, A.E. The symbiotic habit; Princeton University Press: Princeton, NJ, USA, 2010. [Google Scholar]
- Johnston, M.; Yellowlees, D.; Gilmour, I. Carbon isotopic analysis of the free fatty acids in a tridacnid-algal symbiosis: Interpretation and implications for the symbiotic association. Proc. Roy. Soc. Ser. B 1995, 260, 293–297. [Google Scholar] [CrossRef]
- Muscatine, L.; Gates, R.D.; LaFontaine, I. Do symbiotic dinoflagellates secrete lipid droplets? Limnol Oceanogr. 1994, 39, 925–929. [Google Scholar]
- Mortillaro, J.M.; Pitt, K.A.; Lee, S.Y.; Meziane, T. Light intensity influences the production and translocation of fatty acids by zooxanthellae in the jellyfish Cassiopea sp. J. Exp. Mar. Biol. Ecol. 2009, 378, 22–30. [Google Scholar] [CrossRef]
- Harland, A.D.; Navarro, J.C.; Spencer Davies, P.; Fixter, L.M. Lipids of some Caribbean and Red Sea corals: total lipid, wax esters, triglycerides and fatty acids. Mar. Biol. 1993, 117, 113–117. [Google Scholar] [CrossRef]
- Al-Moghrabi, S.; Allemand, D.; Couret, J.M.; Jaubert, J. Fatty acids of the scleractinian coral Galaxea fascicularis: effect of light and feeding. J. Comp. Physiol. Part B 1995, 165, 183–195. [Google Scholar]
- Imbs, A.B.; Demidkova, D.A.; Dautova, T.N.; Latyshev, N.A. Fatty acid biomarkers of symbionts and unusual inhibition of tetracosapolyenoic acid biosynthesis in corals (Octocorallia). Lipids 2009, 44, 325–335. [Google Scholar] [CrossRef]
- Imbs, A.B.; Yakovleva, I.M.; Latyshev, N.A.; Pham, L.Q. Biosynthesis of polyunsaturated fatty acids in zooxanthellae and polyps of corals. Russ. J. Mar. Biol. 2010, 36, 452–457. [Google Scholar] [CrossRef]
- Imbs, A.B.; Latyshev, N.A.; Zhukova, N.V.; Dautova, T.N. Comparison of fatty acid compositions of azooxanthellate Dendronephthya and zooxanthellate soft coral species. Comp. Biochem. Physiol. Part B 2007, 148, 314–321. [Google Scholar] [CrossRef]
- Dunn, S.R.; Thomas, M.C.; Nette, G.W.; Dove, S.G. A lipidomic approach to understanding free fatty acid lipogenesis derived from dissolved inorganic carbon within cnidarian-dinoflagellate symbiosis. PLoS One 2012, 7, e46801. [Google Scholar]
- Meyer, A.; Kirsch, H.; Domergue, F.; Abbadi, A.; Sperling, P.; Bauer, J.; Cirpus, P.; Zank, T.K.; Moreau, H.; Roscoe, T.J.; Zähringer, U.; Heinz, E. Novel fatty acid elongases and their use for the reconstitution of docosahexaenoic acid biosynthesis. J. Lipid Res. 2004, 45, 1899–1909. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Monroig, Ó.; Tocher, D.R.; Navarro, J.C. Biosynthesis of Polyunsaturated Fatty Acids in Marine Invertebrates: Recent Advances in Molecular Mechanisms. Mar. Drugs 2013, 11, 3998-4018. https://doi.org/10.3390/md11103998
Monroig Ó, Tocher DR, Navarro JC. Biosynthesis of Polyunsaturated Fatty Acids in Marine Invertebrates: Recent Advances in Molecular Mechanisms. Marine Drugs. 2013; 11(10):3998-4018. https://doi.org/10.3390/md11103998
Chicago/Turabian StyleMonroig, Óscar, Douglas R. Tocher, and Juan C. Navarro. 2013. "Biosynthesis of Polyunsaturated Fatty Acids in Marine Invertebrates: Recent Advances in Molecular Mechanisms" Marine Drugs 11, no. 10: 3998-4018. https://doi.org/10.3390/md11103998
APA StyleMonroig, Ó., Tocher, D. R., & Navarro, J. C. (2013). Biosynthesis of Polyunsaturated Fatty Acids in Marine Invertebrates: Recent Advances in Molecular Mechanisms. Marine Drugs, 11(10), 3998-4018. https://doi.org/10.3390/md11103998