Astaxanthin Treatment Reduced Oxidative Induced Pro-Inflammatory Cytokines Secretion in U937: SHP-1 as a Novel Biological Target




 ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Cell Viability

{kind=link}
{kind=link}
{kind=link}
| Before Incubation (%) | After Incubation (%) | |
|---|---|---|
| CTRL cells | 96.9 ± 2.4 | 96.0 ± 2.8 |
| H2O2 100 µM | 61.5 ± 2.8 * | |
| H2O2 100 µM + ASTA 10 µM | 78.1 ± 1.9 # | |
| ASTA 10 µM | 97.0 ± 2.2 |
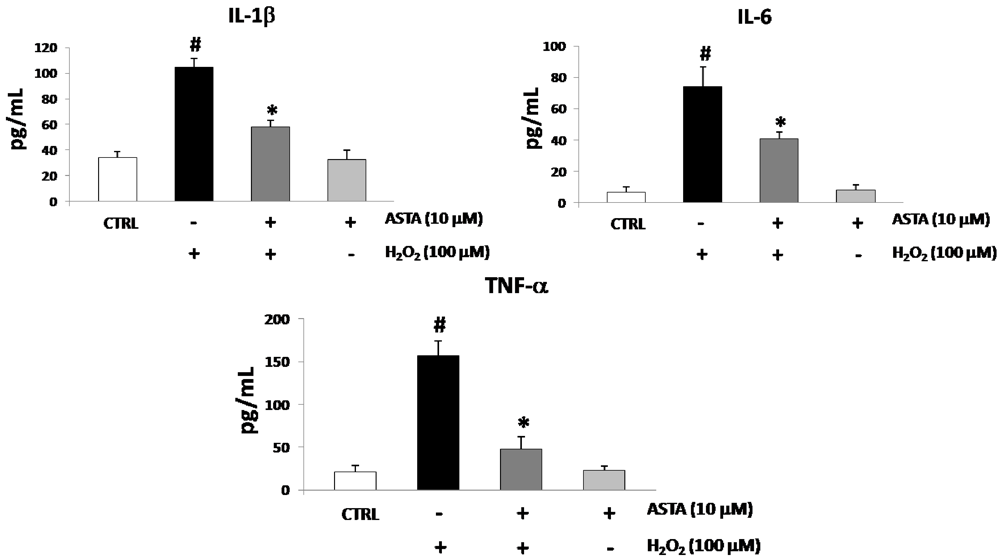
2.2. Cytokines Elisa
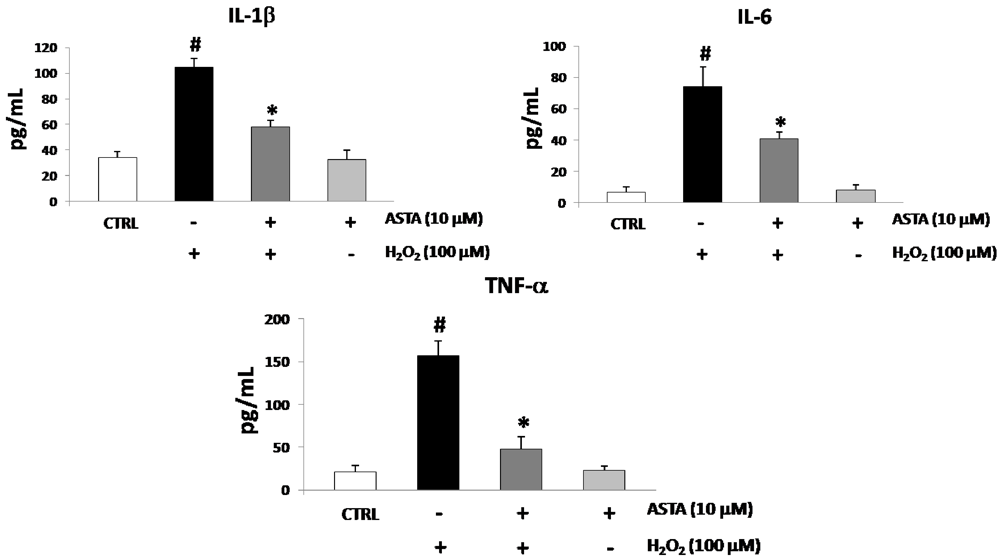
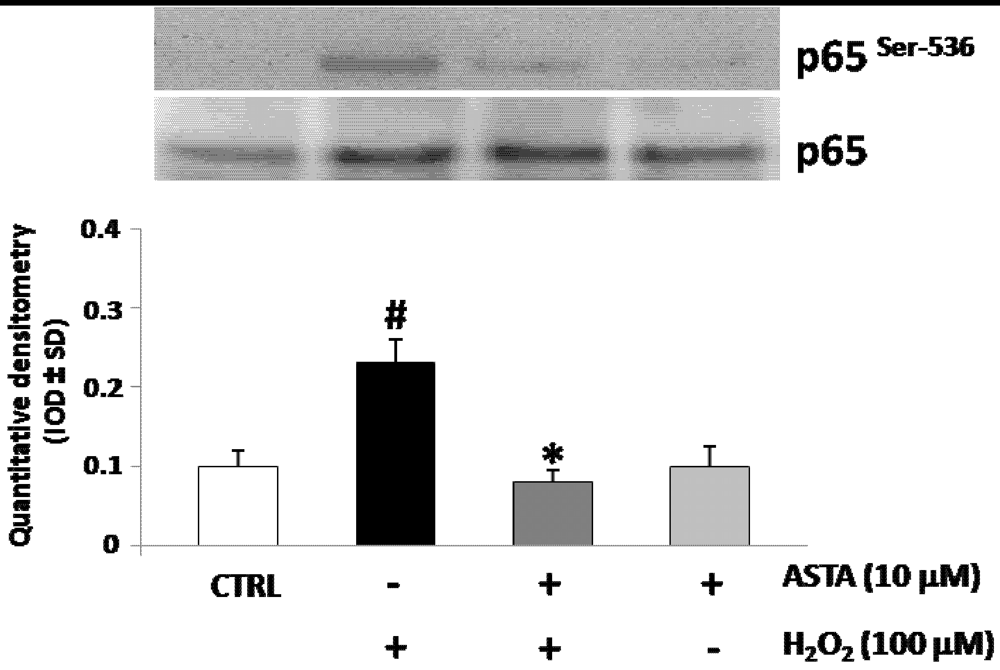
2.3. Effect of H2O2 and ASTA on the NF-κB Activity in U937 Cells
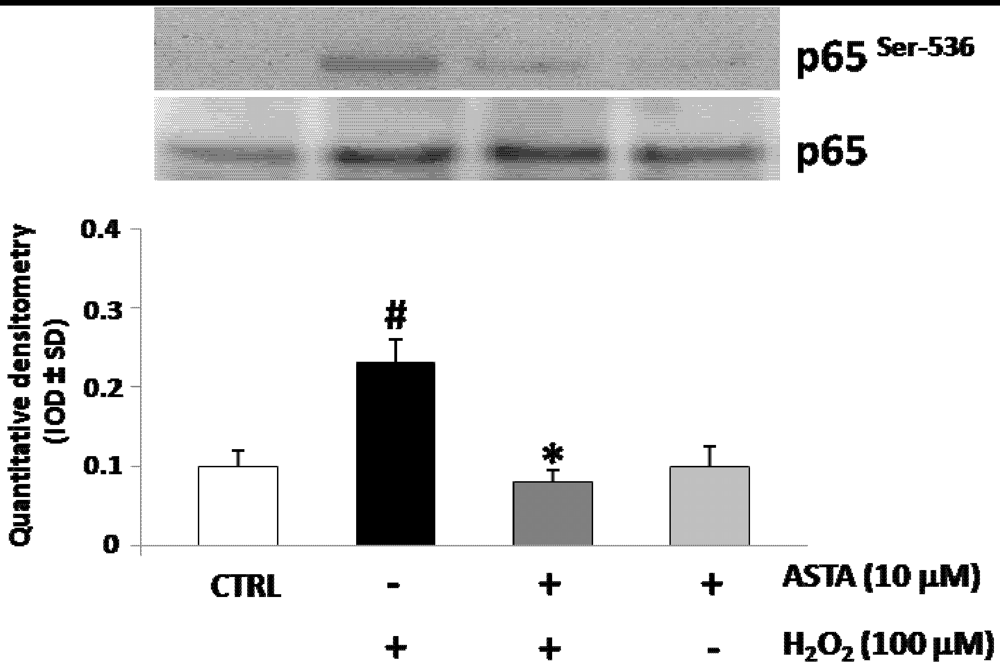
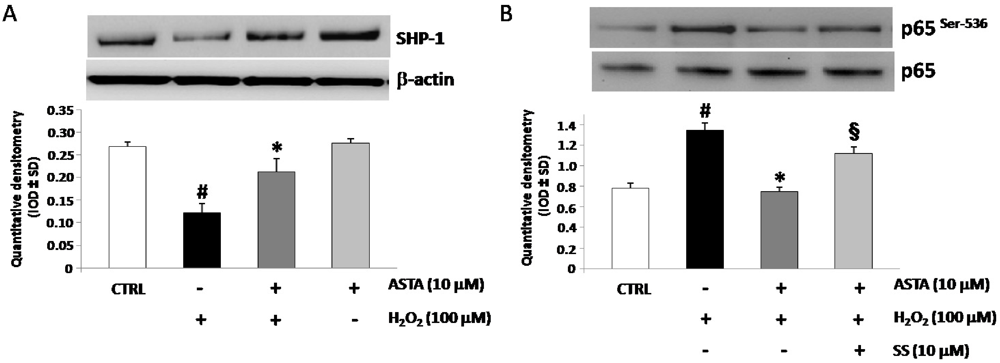
2.4. Effect of H2O2 and ASTA on SHP-1 Expression and Phosphatase Activity in U937 Cells
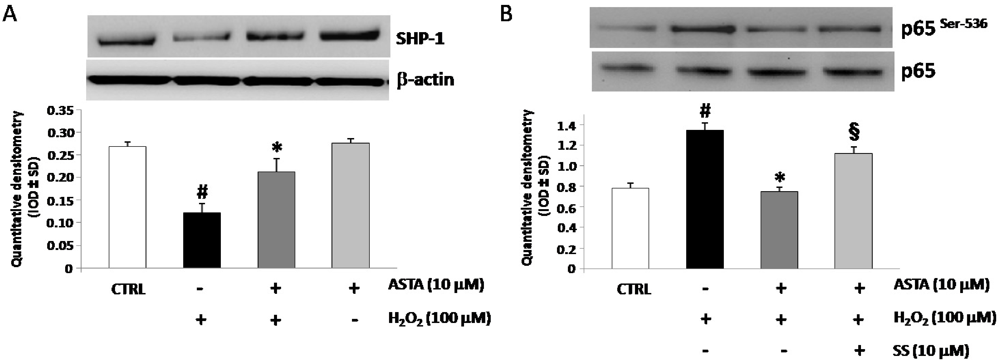
| IL-1β (pg/mL) | IL-6 (pg/mL) | TNF-α (pg/mL) | |
|---|---|---|---|
| CTRL cells | 34.2 ± 4.9 | 7.1 ± 2.9 | 20.4 ± 8.6 |
| H2O2 100 µM | 90.5 ± 4.5 | 66.7 ± 6.0 | 121.4 ± 11.1 |
| H2O2 100 µM + ASTA 10 µM | 56.9 ± 8.3 | 41.5 ± 9.4 | 48.9 ± 12.2 |
| H2O2 100 µM + ASTA 10µM + SS 10 µM | 88.9 ± 9.5 * | 65.2 ± 4.2 * | 122.9 ± 12.5 * |
3. Experimental Section
3.1. Cell Culture
3.2. Reagents
3.3. Cytokines ELISA
3.4. Western Blotting
3.5. Statistical Analysis
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Bartsch, H.; Nair, J. Chronic inflammation and oxidative stress in the genesis and perpetuation of cancer: Role of lipid peroxidation, DNA damage, and repair. Langenbecks Arch. Surg. 2006, 391, 499–510. [Google Scholar] [CrossRef]
- Libby, P. Inflammation and cardiovascular disease mechanisms. Am. J. Clin. Nutr. 2006, 83, 456S–460S. [Google Scholar]
- Speranza, L.; Franceschelli, S.; Pesce, M.; Reale, M.; Menghini, L.; Vinciguerra, I.; de Lutiis, M.A.; Felaco, M.; Grilli, A. Antiinflammatory effects in THP-1 cells treated with verbascoside. Phytother. Res. 2010, 24, 1398–1404. [Google Scholar]
- Levine, B.; Mizushima, N.; Virgin, H.W. Autophagy in immunity and inflammation. Nature 2011, 469, 323–335. [Google Scholar]
- Riccioni, G.; D’Orazio, N.; Franceschelli, S.; Speranza, L. Marine carotenoids and cardiovascular risk markers. Mar. Drugs 2011, 9, 1166–1175. [Google Scholar]
- Wolf, A.M.; Asoh, S.; Hiranuma, H.; Ohsawa, I.; Lio, K.; Satou, A.; Ishikura, M.; Ohta, S. Astaxanthin protects mitochondrial redox state and functional integrity against oxidative stress. J. Nutr. Biochem. 2010, 21, 381–389. [Google Scholar]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar]
- Iwamoto, T.; Hosoda, K.; Hirano, R.; Kurata, H.; Matsumoto, A.; Miki, W.; Kamiyama, M.; Itakura, H.; Yamamoto, S.; Kondo, K. Inhibition of low-density lipoprotein oxidation by astaxanthin. J. Atheroscler. Thromb. 2000, 7, 216–222. [Google Scholar]
- Park, J.S.; Chyun, J.H.; Kim, Y.K.; Line, L.L.; Chew, B.P. Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutr. Metab. (Lond.) 2010, 7. [Google Scholar] [CrossRef]
- Kobayashi, M.; Sakamoto, Y. Singlet oxygen quenching ability of astaxanthin esters from the green alga Haematococcus pluvialis. Biotechnol. Lett. 1999, 21, 265–269. [Google Scholar]
- Yoshida, H.; Yanai, H.; Ito, K.; Tomono, Y.; Koikeda, T.; Tsukahara, H.; Tada, N. Administration of natural astaxanthin increases serum HDL-cholesterol and adiponectin in subjects with mild hyperlipidemia. Atherosclerosis 2010, 209, 520–523. [Google Scholar]
- Lee, S.M.; Kim, E.J.; Suk, K.; Lee, W.H. Synthetic peptides containing ITIM-like sequences of IREM-1 inhibit BAFF-mediated regulation of interleukin-8 expression and phagocytosis through SHP-1 and/or PI3K. Immunology 2011, 134, 224–233. [Google Scholar]
- López-Ruiz, P.; Rodriguez-Ubreva, J.; Cariaga, A.E.; Cortes, M.A.; Colás, B. SHP-1 incell-cycle regulation. Anti-Cancer Agents Med. Chem. 2011, 11, 89–98. [Google Scholar]
- Poole, A.W.; Jones, M.L. A SHPing tale: Perspectives on the regulation of SHP-1 and SHP-2 tyrosine phosphatases by the C-terminal tail. Cell. Signal. 2005, 17, 1323–1332. [Google Scholar]
- Scott, L.M.; Lawrence, H.R.; Sebti, S.M.; Lawrence, N.J.; Wu, J. Targeting protein tyrosine phosphatases for anticancer drug discovery. Curr. Pharm. Des. 2010, 16, 1843–1862. [Google Scholar]
- Yang, J.; Liang, X.; Niu, T.; Meng, W.; Zhao, Z.; Zhou, G.W. Crystal structure of the catalytic domain of protein-tyrosine phosphatase SHP-1. J. Biol. Chem. 1998, 273, 28199–28207. [Google Scholar]
- Lawrence, H.R.; Pireddu, R.; Chen, L.; Luo, Y.; Sung, S.S.; Szymanski, A.M.; Yip, M.L.; Guida, W.C.; Sebti, S.M.; Wu, J.; et al. Inhibitors of Src homology-2 domain containing protein tyrosine phosphatase-2 (Shp2) based on oxindole scaffolds. J. Med. Chem. 2008, 51, 4948–4956. [Google Scholar]
- Christophi, G.P.; Panos, M.; Hudson, C.A.; Tsikkou, C.; Mihai, C.; Mejico, L.J.; Jubelt, B.; Massa, P.T. Interferon-beta treatment in multiple sclerosis attenuates inflammatorygene expression through inducible activity of the phosphatase SHP-1. Clin. Immunol. 2009, 133, 27–44. [Google Scholar]
- Christophi, G.P.; Panos, M.; Hudson, C.A.; Christophi, R.L.; Gruber, R.C.; Mersich, A.T.; Blystone, S.D.; Jubelt, B.; Massa, P.T. Macrophages of multiple sclerosis patients display deficient SHP-1 expression and enhanced inflammatory phenotype. Lab. Invest. 2009, 89, 742–759. [Google Scholar]
- Christophi, G.P.; Hudson, C.A.; Panos, M.; Gruber, R.C.; Massa, P.T. Modulation of macrophage infiltration and inflammatory activity by the phosphatase SHP-1 invirus-induced demyelinating disease. J. Virol. 2009, 83, 522–539. [Google Scholar]
- Verhasselt, V.; Goldman, M.; Willems, F. Oxidative stress up-regulates IL-8 and TNF-alpha synthesis by human dendritic cells. Eur. J. Immunol. 1998, 28, 3886–3890. [Google Scholar]
- Josse, C.; Boelaert, J.R.; Best-Belpomme, M.; Piette, J. Importance of post-transcriptional regulation of chemokine genes by oxidative stress. Biochem. J. 2001, 360, 321–333. [Google Scholar]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radical Bio. Med. 2010, 49, 1603–1616. [Google Scholar]
- Osborn, L.; Kunkel, S.; Nabel, G.J. Tumor necrosis factor alpha and interleukin 1 stimulate the human immunodeficiency virus enhancer by activation of the nuclear factor kappa B. Proc. Natl. Acad. Sci. USA 1989, 86, 2336–2340. [Google Scholar]
- Zhong, H.; May, M.J.; Jimi, E.; Ghosh, S. The phosphorylation status of nuclear NF-kappa B determines its association with CBP/p300 or HDAC-1. Mol. Cell. 2002, 9, 625–636. [Google Scholar]
- Yang, F.; Tang, E.; Guan, K.; Wang, C.Y. IKK beta plays an essential role in the phosphorylation of RelA/p65 on serine 536 induced by ipopolysaccharide. J. Immunol. 2003, 170, 5630–5635. [Google Scholar]
- Wrigley, B.J.; Lip, G.Y.; Shantsila, E. The role of monocytes and inflammation in the pathophysiology of heart failure. Eur. J. Heart Failure 2011, 13, 1161–1171. [Google Scholar]
- Tsui, H.W.; Hasselblatt, K.; Martin, A.; Mok, S.C.; Tsui, F.W. Molecular mechanisms underlying SHP-1 gene expression. Eur. J. Biochem. 2002, 269, 3057–3064. [Google Scholar]
- Anogeianaki, A.; Angelucci, D.; Cianchetti, E.; D’Alessandro, M.; Maccauro, G.; Saggini, A.; Salini, V.; Caraffa, A.; Tete, S.; Conti, F.; et al. Atherosclerosis: A classic inflammatory disease. Int. J. Immunopathol. Pharmacol. 2011, 24, 817–825. [Google Scholar]
- Fassett, R.G.; Coombes, J.S. Astaxanthin, oxidative stress, inflammation and cardiovascular disease. Future Cardiol. 2009, 5, 333–342. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Speranza, L.; Pesce, M.; Patruno, A.; Franceschelli, S.; Lutiis, M.A.d.; Grilli, A.; Felaco, M. Astaxanthin Treatment Reduced Oxidative Induced Pro-Inflammatory Cytokines Secretion in U937: SHP-1 as a Novel Biological Target. Mar. Drugs 2012, 10, 890-899. https://doi.org/10.3390/md10040890
Speranza L, Pesce M, Patruno A, Franceschelli S, Lutiis MAd, Grilli A, Felaco M. Astaxanthin Treatment Reduced Oxidative Induced Pro-Inflammatory Cytokines Secretion in U937: SHP-1 as a Novel Biological Target. Marine Drugs. 2012; 10(4):890-899. https://doi.org/10.3390/md10040890
Chicago/Turabian StyleSperanza, Lorenza, Mirko Pesce, Antonia Patruno, Sara Franceschelli, Maria Anna de Lutiis, Alfredo Grilli, and Mario Felaco. 2012. "Astaxanthin Treatment Reduced Oxidative Induced Pro-Inflammatory Cytokines Secretion in U937: SHP-1 as a Novel Biological Target" Marine Drugs 10, no. 4: 890-899. https://doi.org/10.3390/md10040890
APA StyleSperanza, L., Pesce, M., Patruno, A., Franceschelli, S., Lutiis, M. A. d., Grilli, A., & Felaco, M. (2012). Astaxanthin Treatment Reduced Oxidative Induced Pro-Inflammatory Cytokines Secretion in U937: SHP-1 as a Novel Biological Target. Marine Drugs, 10(4), 890-899. https://doi.org/10.3390/md10040890