
Suramin Exerts an Ameliorative Effect on Acetic Acid-Induced Acute Colitis in Rats by Demonstrating Potent Antioxidant and Anti-Inflammatory Properties


Abstract

1. Introduction
2. Materials and Methods
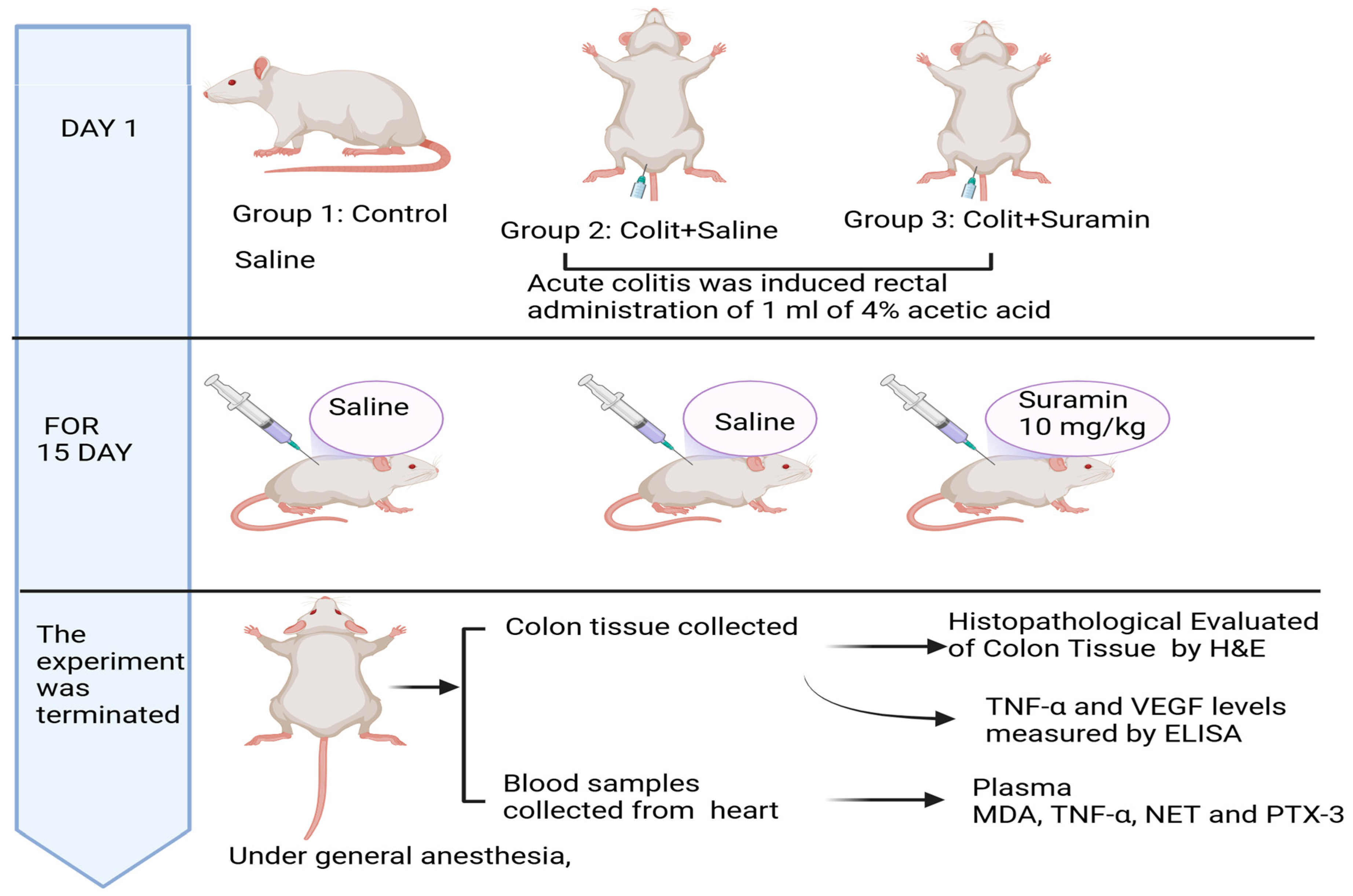
2.1. Animals
2.2. Experimental Model of Acute Colitis
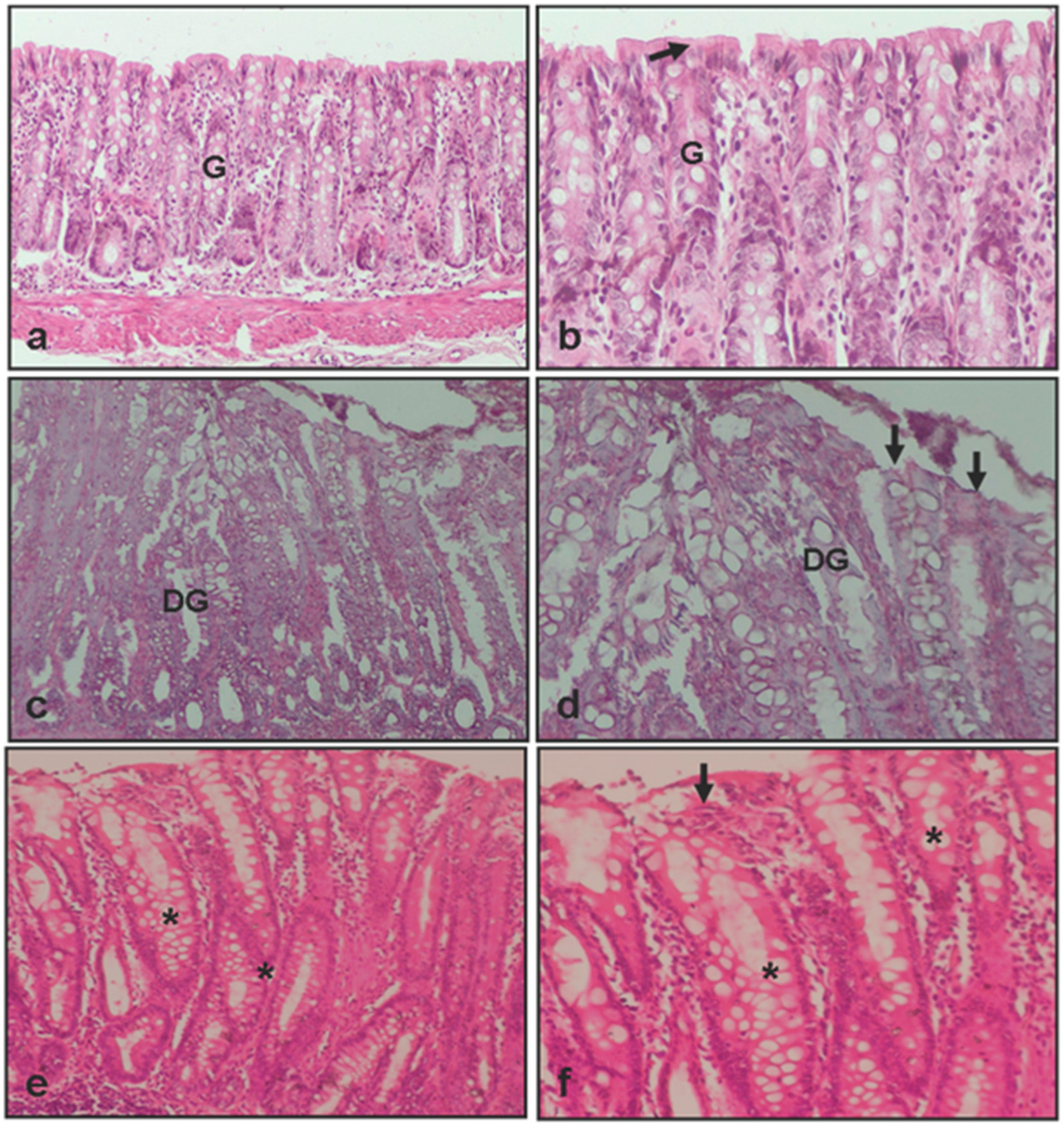
2.3. Histopathological Evaluation of Colon Tissue
- 0:
- Intact epithelium; no leukocyte infiltration or hemorrhage
- 1:
- <25% epithelial damage with focal leukocyte infiltration and hemorrhage
- 2:
- 25% epithelial disruption with focal infiltrates and hemorrhage
- 3:
- 50% epithelial damage with widespread leukocyte infiltration and hemorrhage
- 4:
- >50% epithelial damage with extensive infiltration and hemorrhage
2.4. Measurement of Plasma TNF-α and NETs Levels
2.5. Measurement of Plasma PTX3
2.6. Detection of TNF-α and VEGF in Colon Tissue
2.7. Plasma Lipid Peroxidation Measurement
2.8. Statistical Analysis
3. Results
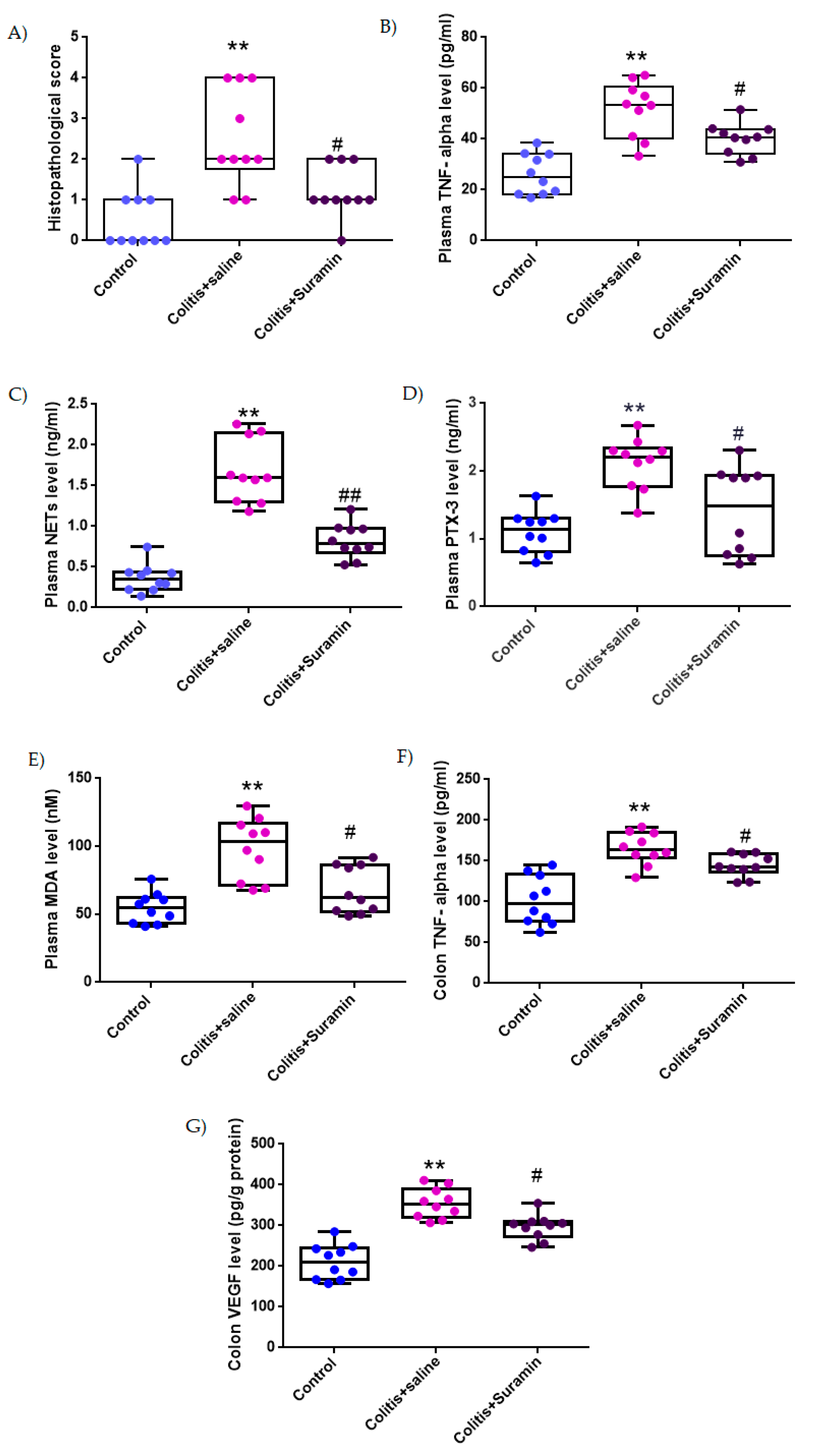
3.1. Histopathological Evaluation
3.2. Plasma TNF-α Concentrations
3.3. Plasma NETs Levels
3.4. Plasma PTX3 Concentrations
3.5. Plasma MDA Levels
3.6. Colonic TNF-α Levels
3.7. Colonic VEGF Levels
4. Discussion
5. Conclusions
6. Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, R.; Weber, M.C.; Hu, X.; Neumann, P.A.; Kamaly, N. Annexin A1 based inflammation re-solving mediators and nanomedicines for inflammatory bowel disease therapy. Semin Immunol. 2022, 61–64, 101664. [Google Scholar] [CrossRef] [PubMed]
- Zaghloul, M.S.; Elshal, M.; Abdelmageed, M.E. Preventive empagliflozin activity on acute acetic acid-induced ulcerative colitis in rats via modulation of SIRT-1/PI3K/AKT pathway and improving colon barrier. Environ. Toxicol. Pharmacol. 2022, 91, 103833. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Sandborn, W.J. Inflammatory bowel disease: Clinical aspects and established and evolving therapies. Lancet 2007, 369, 1641–1657. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef] [PubMed]
- Danpanichkul, P.; Duangsonk, K.; Lopimpisuth, C.; Ho, A.H.-Y.; Fangsaard, P.; Sukphutanan, B.; Pang, Y.; Chaisrimaneepan, N.; Dejvajara, D.; Suenghataiphorn, T.; et al. Geographical and sociodemographic epidemiology of inflammatory bowel disease in young females from 2010 to 2019. Dig. Liver Dis. 2025, 57, 190–197. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N. Environmental Risk Factors for Inflammatory Bowel Diseases: A Review. Dig. Dis. Sci. 2015, 60, 290–298. [Google Scholar] [CrossRef]
- Tripathi, K.; Feuerstein, J.D. New developments in ulcerative colitis: Latest evidence on management, treatment, and maintenance. Drugs Context 2019, 8, 212572. [Google Scholar] [CrossRef]
- Yamamoto-Furusho, J.K.; Gutierrez-Herrera, F.D. Molecular Mechanisms and Clinical Aspects of Colitis-Associated Cancer in Ulcerative Colitis. Cells 2025, 14, 162. [Google Scholar] [CrossRef]
- Bhat, M.A.; Usman, I.; Dhaneshwar, S. Application of Drug Repurposing Approach for Therapeutic Intervention of Inflammatory Bowel Disease. Curr. Rev. Clin. Exp. Pharmacol. Former. Curr. Clin. Pharmacol. 2024, 19, 234–249. [Google Scholar] [CrossRef]
- Kobayashi, T.; Siegmund, B.; Le Berre, C.; Wei, S.C.; Ferrante, M.; Shen, B.; Bernstein, C.N.; Danese, S.; Peyrin-Biroulet, L.; Hibi, T. Ulcerative colitis. Nat. Rev. Dis. Prim. 2020, 6, 74. [Google Scholar] [CrossRef]
- Li, M.-X.; Li, M.-Y.; Lei, J.-X.; Wu, Y.-Z.; Li, Z.-H.; Chen, L.-M.; Zhou, C.-L.; Su, J.-Y.; Huang, G.-X.; Huang, X.-Q.; et al. Huangqin decoction ameliorates DSS-induced ulcerative colitis: Role of gut microbiota and amino acid metabolism, mTOR pathway and intestinal epithelial barrier. Phytomedicine 2022, 100, 154052. [Google Scholar] [CrossRef] [PubMed]
- Loh, J.T.; Lee, K.-G.; Lee, A.P.; Teo, J.K.H.; Lim, H.L.; Kim, S.S.-Y.; Tan, A.H.-M.; Lam, K.-P. DOK3 maintains intestinal homeostasis by suppressing JAK2/STAT3 signaling and S100a8/9 production in neutrophils. Cell Death Dis. 2021, 12, 1054. [Google Scholar] [CrossRef]
- Strober, W.; Fuss, I.J. Proinflammatory Cytokines in the Pathogenesis of Inflammatory Bowel Diseases. Gastroenterology 2011, 140, 1756–1767.e1. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, M. Oxidative Stress-Induced Gastrointestinal Diseases: Biology and Nanomedicines—A Review. BioChem 2024, 4, 189–216. [Google Scholar] [CrossRef]
- McGeary, R.P.; Bennett, A.J.; Tran, Q.B.; Cosgrove, K.L.; Ross, B.P. Suramin: Clinical Uses and Structure-Activity Relationships. Mini-Rev. Med. Chem. 2008, 8, 1384–1394. [Google Scholar] [CrossRef] [PubMed]
- Di Prospero, N.A.; Zhou, X.-R.; Meiners, S.; Mcauliffe, W.G.; Ho, S.-Y.; Geller, H.M. Suramin disrupts the gliotic response following a stab wound injury to the adult rat brain. J. Neurocytol. 1998, 27, 491–506. [Google Scholar] [CrossRef]
- Alyoussef, A. Suramin Attenuated Inflammation and Reversed Skin Tissue Damage in Experimentally Induced Atopic Dermatitis in Mice. Inflamm. Allergy-Drug Targets 2015, 13, 406–410. [Google Scholar] [CrossRef]
- Bademci, R.; Erdoğan, M.A.; Kara, A.Y.; Yiğittürk, G.; Erbaş, O. Therapeutic effects of vitamin D on acetic acid-induced colitis in rats. Acta Cir. Bras. 2020, 35, e202000404. [Google Scholar] [CrossRef]
- Ercan, G.; Yigitturk, G.; Erbas, O. Therapeutic effect of adenosine on experimentally induced acute ulcerative colitis model in rats. Acta Cir. Bras. 2020, 34, e201901204. [Google Scholar] [CrossRef]
- MacPherson, B.; Pfeiffer, C.J. Experimental colitis. Digestion 1976, 14, 424–452. [Google Scholar] [CrossRef]
- Melo, N.M.d.C.; Almeida, M.V.S.; Campos, D.M.d.O.; de Oliveira, C.B.S.; Oliveira, J.I.N. Animal models for inducing inflammatory bowel diseases: Integrative review. HSJ 2021, 11, 80–87. [Google Scholar] [CrossRef]
- Hagar, H.H.; El Medany, A.; El Eter, E.; Arafa, M. Ameliorative effect of pyrrolidinedithiocarbamate on acetic acid-induced colitis in rats. Eur. J. Pharmacol. 2007, 554, 69–77. [Google Scholar] [CrossRef]
- El-Akabawy, G.; El-Sherif, N.M. Zeaxanthin exerts protective effects on acetic acid-induced colitis in rats via modulation of pro-inflammatory cytokines and oxidative stress. Biomed. Pharmacother. 2019, 111, 841–851. [Google Scholar] [CrossRef]
- Ungaro, R.; Mehandru, S.; Allen, P.B.; Peyrin-Biroulet, L.; Colombel, J.-F. Ulcerative colitis. Lancet 2017, 389, 1756–1770. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Ye, B.; Ren, Y. ABIN1 Inhibits Inflammation through Necroptosis-Dependent Pathway in Ulcerative Colitis. Genet. Res. 2022, 2022, 9313559. [Google Scholar] [CrossRef]
- Nakase, H.; Sato, N.; Mizuno, N.; Ikawa, Y. The influence of cytokines on the complex pathology of ulcerative colitis. Autoimmun. Rev. 2022, 21, 103017. [Google Scholar] [CrossRef] [PubMed]
- Mao, L.; Kitani, A.; Strober, W.; Fuss, I.J. The Role of NLRP3 and IL-1β in the Pathogenesis of Inflammatory Bowel Disease. Front. Immunol. 2018, 9, 2566. [Google Scholar] [CrossRef]
- Al-Rejaie, S.S.; Abuohashish, H.M.; Al-Enazi, M.M.; Al-Assaf, A.H.; Parmar, M.Y.; Ahmed, M.M. Protective effect of naringenin on acetic acid-induced ulcerative colitis in rats. World J. Gastroenterol. WJG 2013, 19, 5633. [Google Scholar] [CrossRef] [PubMed]
- Rossaint, J.; Herter, J.M.; Van Aken, H.; Napirei, M.; Döring, Y.; Weber, C.; Soehnlein, O.; Zarbock, A. Synchronized integrin engagement and chemokine activation is crucial in neutrophil extracellular trap–mediated sterile inflammation. Blood J. Am. Soc. Hematol. 2014, 123, 2573–2584. [Google Scholar] [CrossRef]
- Szatmary, P.; Huang, W.; Criddle, D.; Tepikin, A.; Sutton, R. Biology, role and therapeutic potential of circulating histones in acute inflammatory disorders. J. Cell. Mol. Med. 2018, 22, 4617–4629. [Google Scholar] [CrossRef]
- Angelidou, I.; Chrysanthopoulou, A.; Mitsios, A.; Arelaki, S.; Arampatzioglou, A.; Kambas, K.; Ritis, D.; Tsironidou, V.; Moschos, I.; Dalla, V.; et al. REDD1/autophagy pathway is associated with neutrophil-driven IL1b Inflammatory response in active ulcerative colitis. J. Immunol. 2018, 200, 3950–3961. [Google Scholar] [CrossRef] [PubMed]
- Dinallo, V.; Marafini, I.; Di Fusco, D.; Laudisi, F.; Franzè, E.; Di Grazia, A.; Figliuzzi, M.M.; Caprioli, F.; Stolfi, C.; Monteleone, I.; et al. Neutrophil Extracellular Traps Sustain Inflammatory Signals in Ulcerative Colitis. J. Crohn’s Colitis 2019, 13, 772–784. [Google Scholar] [CrossRef]
- El Hafez, A.A.; Mohamed, A.S.; Shehta, A.; Sheta, H.A.E.A.S. Neutrophil extracellular traps-associated protein peptidyl arginine deaminase 4 immunohistochemical expression in ulcerative colitis and its association with the prognostic predictors. Pathol. Res. Pract. 2020, 216, 153102. [Google Scholar] [CrossRef] [PubMed]
- Bennike, T.B.; Carlsen, T.G.; Ellingsen, T.; Bonderup, O.K.; Glerup, H.; Bøgsted, M.; Christiansen, G.; Birkelund, S.; Stensballe, A.; Andersen, V. Neutrophil extracellular traps in ulcerative colitis: A proteome analysis of intestinal biopsies. Inflamm. Bowel Dis. 2015, 21, 2052–2067. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Wang, C.; Liu, Y.; Li, B.; Zhang, W.; Wang, L.; Yu, M.; Zhao, X.; Du, J.; Zhang, J.; et al. Neutrophil Extracellular Traps Induce Intestinal Damage and Thrombotic Tendency in Inflammatory Bowel Disease. J. Crohn’s Colitis 2020, 14, 240–253. [Google Scholar] [CrossRef]
- Xu, C.; Ye, Z.; Jiang, W.; Wang, S.; Zhang, H. Cyclosporine A alleviates colitis by inhibiting the formation of neutrophil extracellular traps via the regulating pentose phosphate pathway. Mol. Med. 2023, 29, 169. [Google Scholar] [CrossRef]
- Keshari, R.S.; Jyoti, A.; Dubey, M.; Kothari, N.; Kohli, M.; Bogra, J.; Barthwal, M.K.; Dikshit, M. Cytokines Induced Neutrophil Extracellular Traps Formation: Implication for the Inflammatory Disease Condition. PLoS ONE 2012, 7, e48111. [Google Scholar] [CrossRef]
- Long, D.; Mao, C.; Xu, Y.; Zhu, Y. The emerging role of neutrophil extracellular traps in ulcerative colitis. Front. Immunol. 2024, 15, 1425251. [Google Scholar] [CrossRef]
- Alzani, R.; Cozzi, E.; Corti, A.; Temponi, M.; Trizio, D.; Gigli, M.; Rizzo, V. Mechanism of suramin-induced deoligomerization of tumor necrosis factor alpha. Biochemistry 1995, 34, 6344–6350. [Google Scholar] [CrossRef]
- Mancini, F.; Toro, C.M.; Mabilia, M.; Giannangeli, M.; Pinza, M.; Milanese, C. Inhibition of tumor necrosis factor-α (TNF-α)/TNF-α receptor binding by structural analogues of suramin. Biochem. Pharmacol. 1999, 58, 851–859. [Google Scholar] [CrossRef]
- Peyrin-Biroulet, L. Anti-TNF therapy in inflammatory bowel diseases: A huge review. Minerva Gastroenterol. e dietologica 2010, 56, 233–243. [Google Scholar]
- Villalba, N.; Sackheim, A.M.; A Lawson, M.; Haines, L.; Chen, Y.-L.; Sonkusare, S.K.; Ma, Y.-T.; Li, J.; Majumdar, D.; Bouchard, B.A.; et al. The Polyanionic Drug Suramin Neutralizes Histones and Prevents Endotheliopathy. J. Immunol. 2023, 211, 648–657. [Google Scholar] [CrossRef]
- Bottazzi, B.; Garlanda, C.; Cotena, A.; Moalli, F.; Jaillon, S.; Deban, L.; Mantovani, A. The long pentraxin PTX3 as a prototypic humoral pattern recognition receptor: Interplay with cellular innate immunity. Immunol. Rev. 2009, 227, 9–18. [Google Scholar] [CrossRef]
- Kato, S.; Ochiai, M.; Sakurada, T.; Ohno, S.; Miyamoto, K.; Sagara, M.; Ito, M.; Takeuchi, K.; Imaki, J.; Itoh, K.; et al. Increased Expression of Long Pentraxin PTX3 in Inflammatory Bowel Diseases. Dig. Dis. Sci. 2008, 53, 1910–1916. [Google Scholar] [CrossRef]
- Woo, J.M.; Kwon, M.-Y.; Shin, D.-Y.; Kang, Y.-H.; Hwang, N.; Chung, S.W. Human retinal pigment epithelial cells express the long pentraxin PTX3. Mol. Vis. 2013, 19, 303–310. [Google Scholar]
- Souza, D.G.; Amaral, F.A.; Fagundes, C.T.; Coelho, F.M.; Arantes, R.M.; Sousa, L.P.; Matzuk, M.M.; Garlanda, C.; Mantovani, A.; Dias, A.A.; et al. The Long Pentraxin PTX3 Is Crucial for Tissue Inflammation after Intestinal Ischemia and Reperfusion in Mice. Am. J. Pathol. 2009, 174, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Jaillon, S.; Peri, G.; Delneste, Y.; Frémaux, I.; Doni, A.; Moalli, F.; Mantovani, A. The humoral pattern recognition receptor PTX3 is stored in neutrophil granules and localizes in extracellular traps. J. Exp. Med. 2007, 204, 793–804. [Google Scholar] [CrossRef]
- Imamura, M.; Kawasaki, T.; Savchenko, A.S.; Ohashi, R.; Jiang, S.; Miyamoto, K.; Ito, Y.; Iwanari, H.; Sagara, M.; Tanaka, T.; et al. Lipopolysaccharide induced expression of pentraxin 3 in human neutrophils and monocyte-derived macrophages. Cell. Immunol. 2007, 248, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Tian, W.-F.; Zeng, S.; Sheng, Q.; Chen, J.-L.; Weng, P.; Zhang, X.-T.; Yuan, J.-J.; Pang, Q.-F.; Wang, Z.-Q. Methylene Blue Protects the Isolated Rat Lungs from Ischemia–Reperfusion Injury by Attenuating Mitochondrial Oxidative Damage. Lung 2018, 196, 73–82. [Google Scholar] [CrossRef]
- Zeng, Y.-S.; Peng, J.; Gao, X.-F.; Tian, D.; Zhan, W.; Liu, J.; Hu, X.-J.; Huang, S.; Tian, S.-T.; Qiu, L.; et al. A novel gut-restricted RIPK1 inhibitor, SZ-15, ameliorates DSS-induced ulcerative colitis. Eur. J. Pharmacol. 2022, 937, 175381. [Google Scholar] [CrossRef]
- Hwang, N.; Kwon, M.-Y.; Woo, J.M.; Chung, S.W. Oxidative Stress-Induced Pentraxin 3 Expression Human Retinal Pigment Epithelial Cells Is Involved in the Pathogenesis of Age-Related Macular Degeneration. Int. J. Mol. Sci. 2019, 20, 6028. [Google Scholar] [CrossRef] [PubMed]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2018, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Betto, T.; Amano, H.; Ito, Y.; Eshima, K.; Yoshida, T.; Matsui, Y.; Yamane, S.; Inoue, T.; Otaka, F.; Kobayashi, K.; et al. Vascular endothelial growth factor receptor 1 tyrosine kinase signaling facilitates healing of DSS-induced colitis by accumulation of Tregs in ulcer area. Biomed. Pharmacother. 2019, 111, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Tolstanova, G.; Khomenko, T.; Deng, X.; Chen, L.; Tarnawski, A.; Ahluwalia, A.; Szabo, S.; Sandor, Z. Neutralizing Anti-Vascular Endothelial Growth Factor (VEGF) Antibody Reduces Severity of Experimental Ulcerative Colitis in Rats: Direct Evidence for the Pathogenic Role of VEGF. J. Pharmacol. Exp. Ther. 2009, 328, 749–757. [Google Scholar] [CrossRef]
- Tolstanova, G.; Khomenko, T.; Deng, X.; Szabo, S.; Sandor, Z. New molecular mechanisms of the unexpectedly complex role of VEGF in ulcerative colitis. Biochem. Biophys. Res. Commun. 2010, 399, 613–616. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Group | Colitis + Saline | Colitis + Suramin | |
|---|---|---|---|
| Histopathological score | 0.5 ± 0.2 | 2.5 ± 0.4 ** | 1.2 ± 0.2 # |
| Plasma TNF-α level (pg/mL) | 26.1 ± 2.5 | 51.6 ± 3.4 ** | 40.1 ± 1.9 # |
| Plasma NETs level (ng/mL) | 0.36 ± 0.05 | 1.67 ± 0.12 ** | 0.82 ± 0.06 ## |
| Plasma PTX3 level (ng/mL) | 1.1 ± 0.09 | 2.1 ± 0.1 ** | 1.4 ± 0.2 # |
| Plasma MDA level (nM) | 54.6 ± 3.5 | 98.2 ± 7.1 ** | 67.8 ± 5.5 # |
| Colon TNF-α level (pg/mg tissue) | 101.7 ± 9.3 | 165.08 ± 6.2 ** | 144.6 ± 4.3 # |
| Colon VEGF level (pg/g protein) | 210.6 ± 13.5 | 354.8 ± 11.6 ** | 295.9 ± 9.7 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Published by MDPI on behalf of the Lithuanian University of Health Sciences. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ercan, G.; Aygün, H.; Akbaş, A.; Çınaroğlu, O.S.; Erbas, O. Suramin Exerts an Ameliorative Effect on Acetic Acid-Induced Acute Colitis in Rats by Demonstrating Potent Antioxidant and Anti-Inflammatory Properties. Medicina 2025, 61, 829. https://doi.org/10.3390/medicina61050829
Ercan G, Aygün H, Akbaş A, Çınaroğlu OS, Erbas O. Suramin Exerts an Ameliorative Effect on Acetic Acid-Induced Acute Colitis in Rats by Demonstrating Potent Antioxidant and Anti-Inflammatory Properties. Medicina. 2025; 61(5):829. https://doi.org/10.3390/medicina61050829
Chicago/Turabian StyleErcan, Gulcin, Hatice Aygün, Ahmet Akbaş, Osman Sezer Çınaroğlu, and Oytun Erbas. 2025. "Suramin Exerts an Ameliorative Effect on Acetic Acid-Induced Acute Colitis in Rats by Demonstrating Potent Antioxidant and Anti-Inflammatory Properties" Medicina 61, no. 5: 829. https://doi.org/10.3390/medicina61050829
APA StyleErcan, G., Aygün, H., Akbaş, A., Çınaroğlu, O. S., & Erbas, O. (2025). Suramin Exerts an Ameliorative Effect on Acetic Acid-Induced Acute Colitis in Rats by Demonstrating Potent Antioxidant and Anti-Inflammatory Properties. Medicina, 61(5), 829. https://doi.org/10.3390/medicina61050829