
Role of Elevated Serum TGF-β1 and the Common Promoter TGFB1-509C/T Polymorphism in the Development and Progression of Primary Glial Tumors and Brain Metastases


 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Blood Samples and DNA Extraction
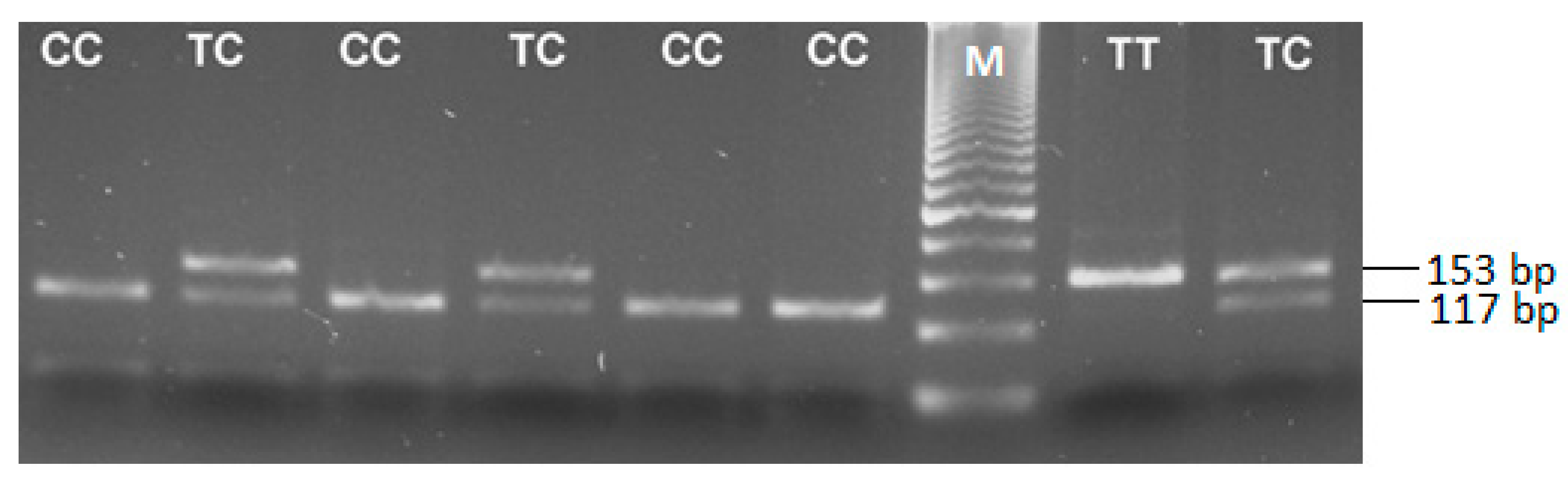
2.3. Genotyping for the TGFB1 -509 C/T Polymorphism (rs1800469)
2.4. Quantification of Serum TGF-β1 Levels
2.5. Immunohistochemistry
2.6. Statistical Analyses
3. Results
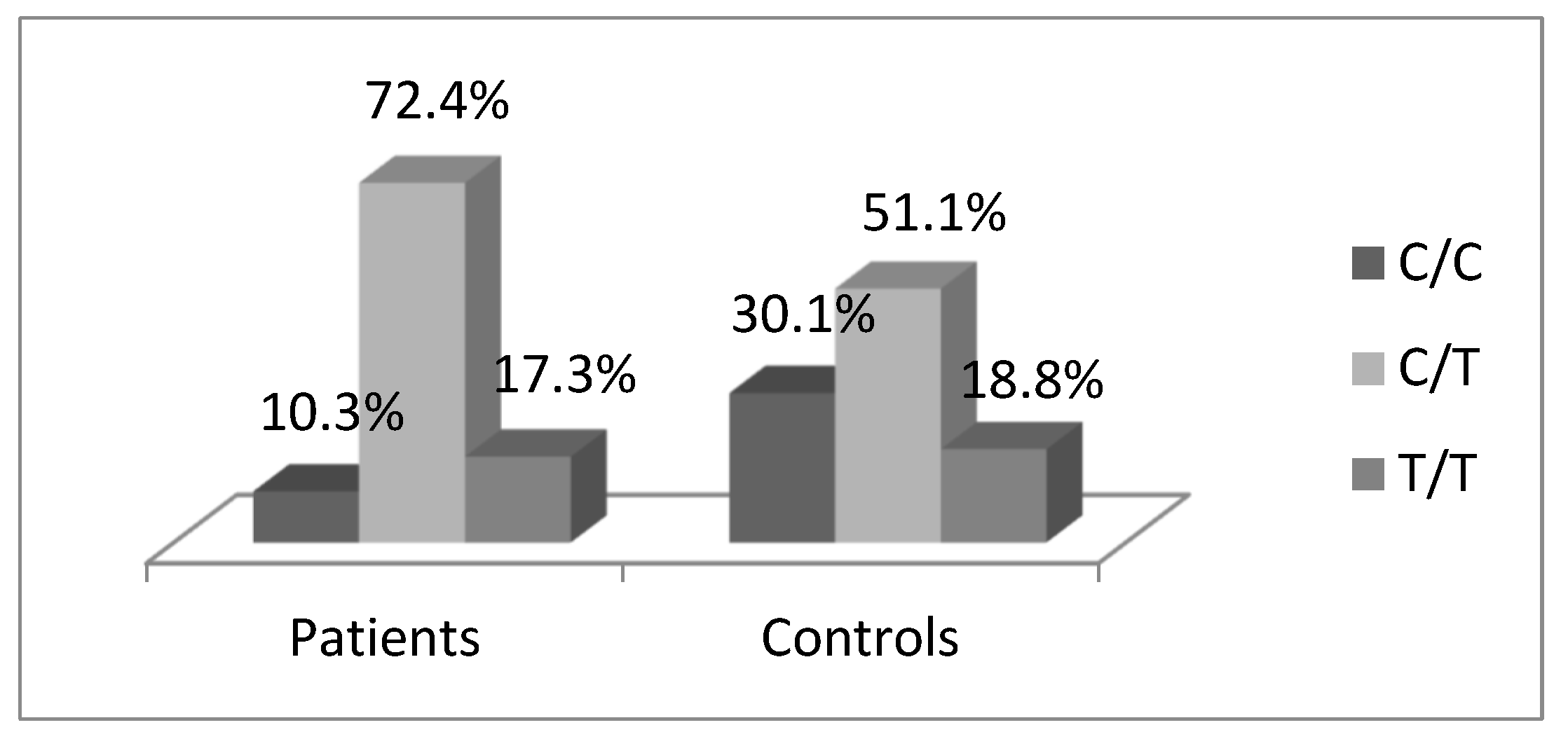
3.1. Distribution of TGFB1-509C/T Polymorphism
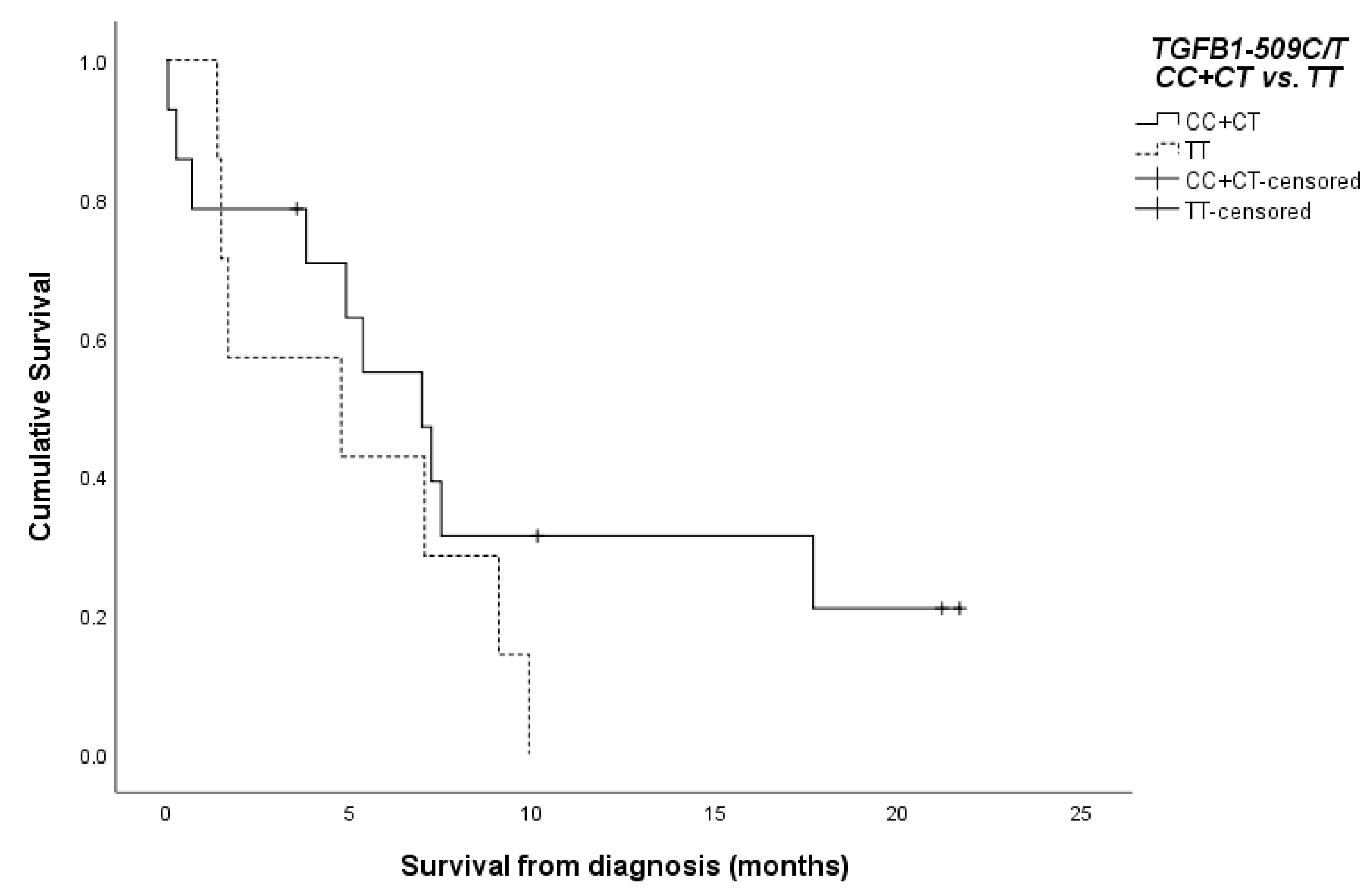
3.2. Survival of the Patients According to the TGFB1-509C/T SNP
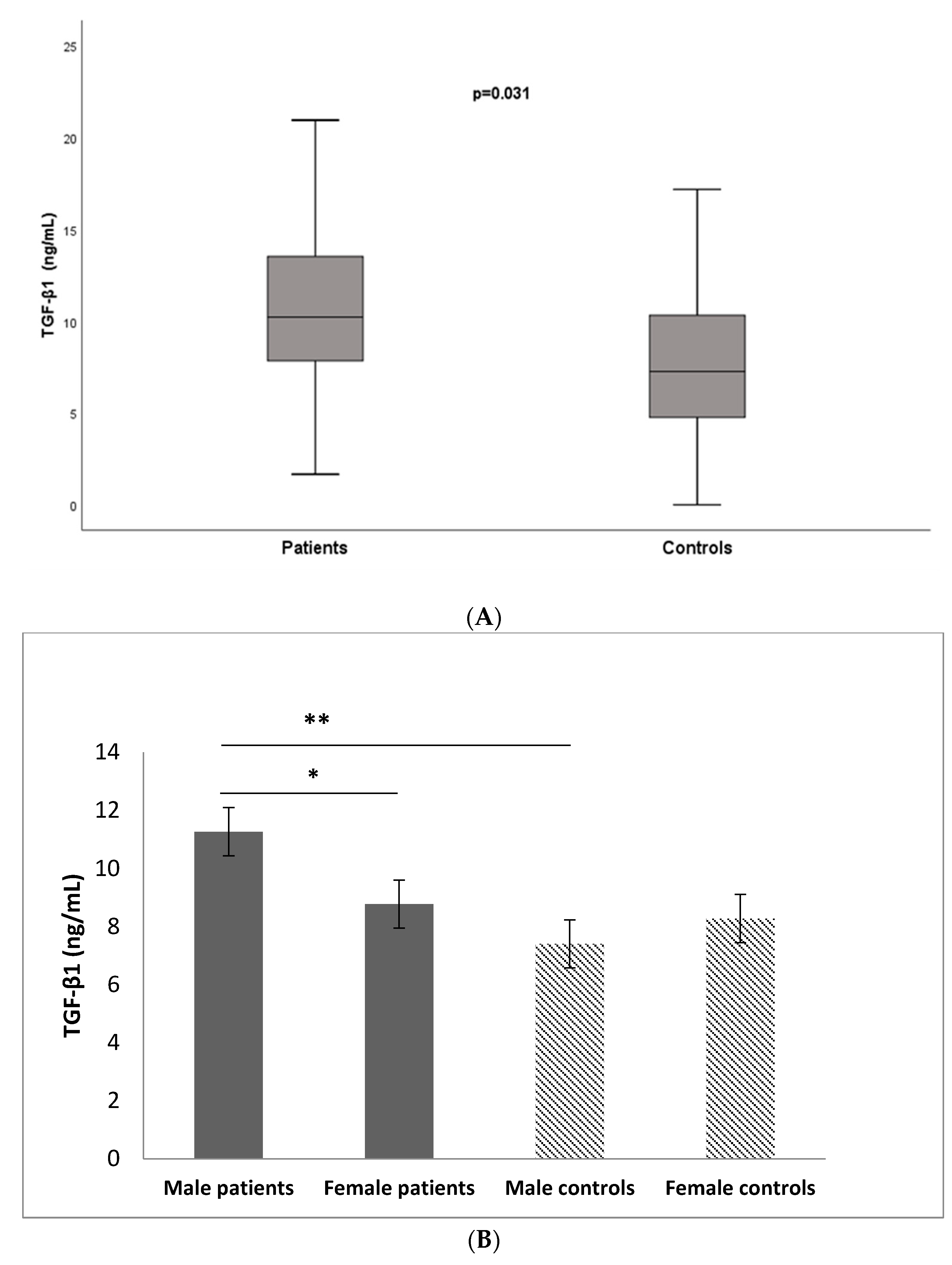
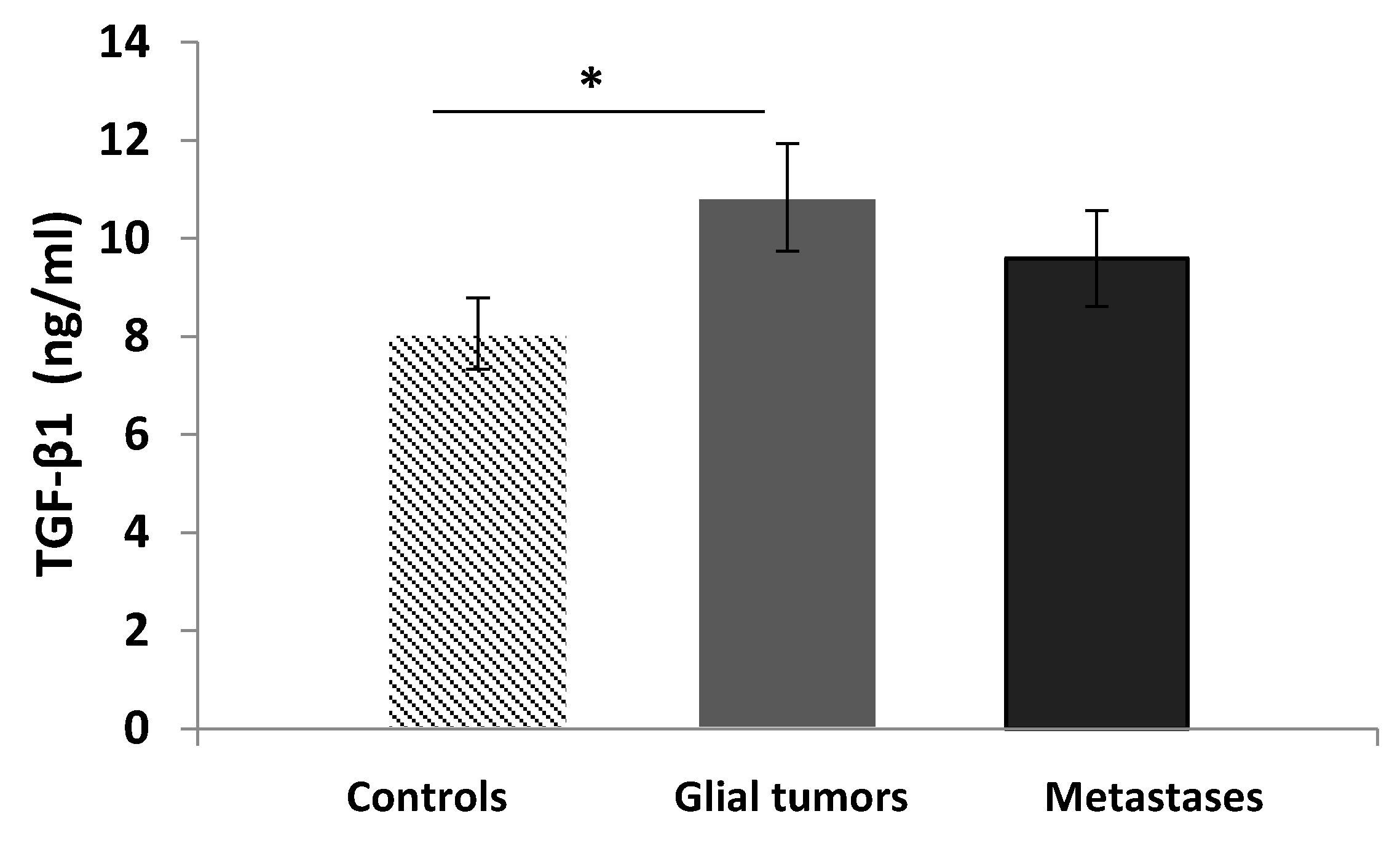
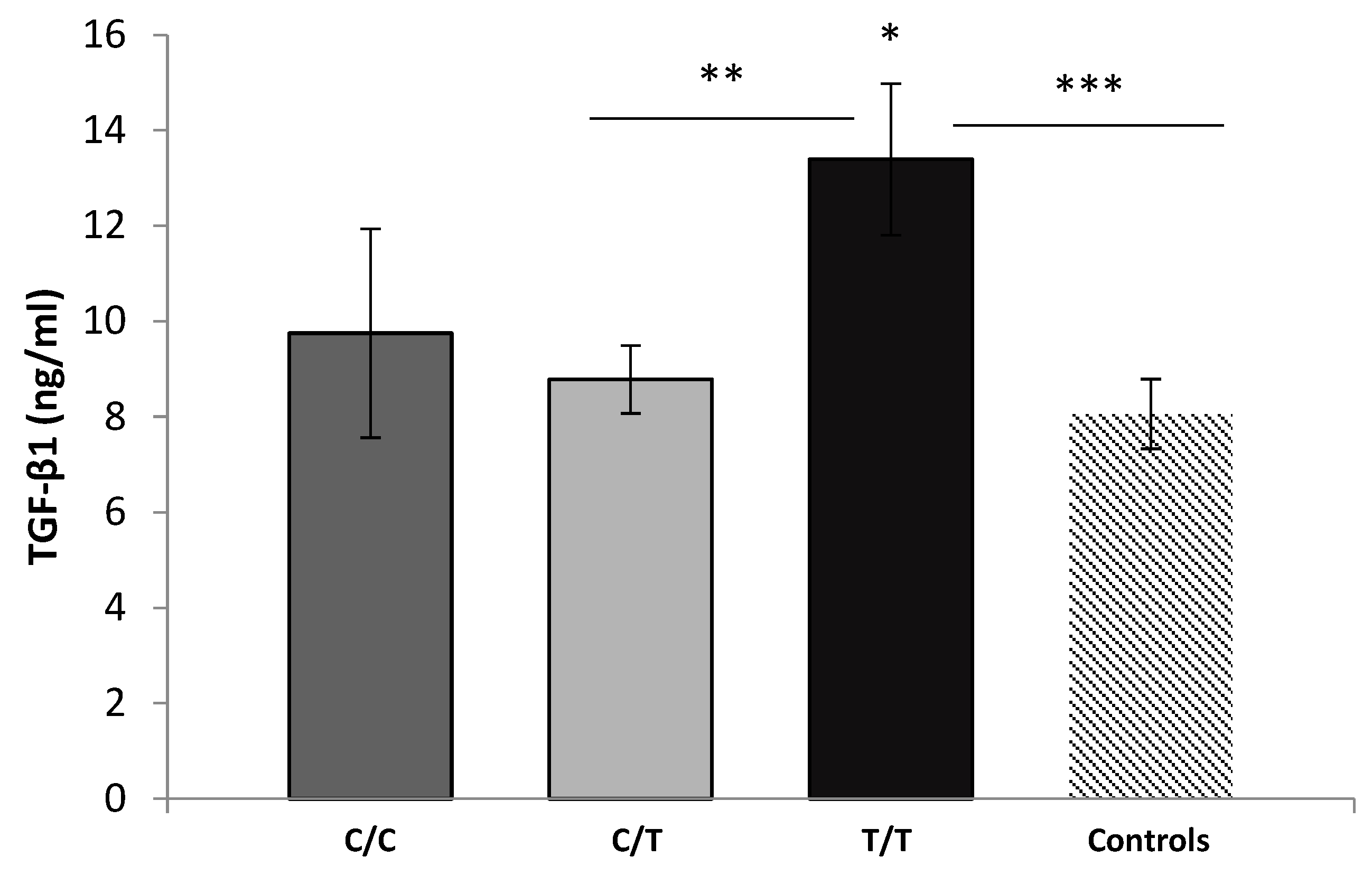
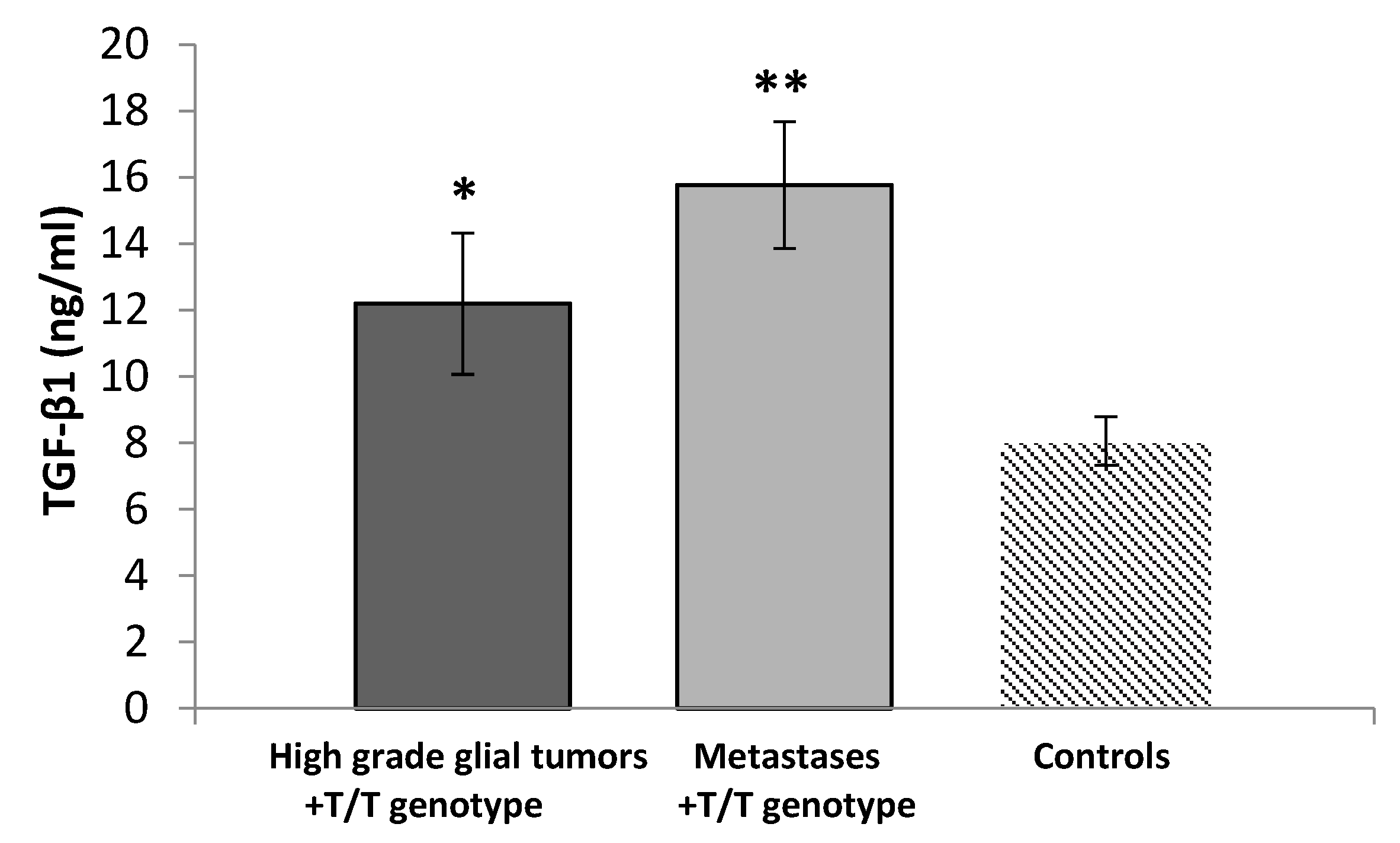
3.3. Serum Levels of TGF-β1
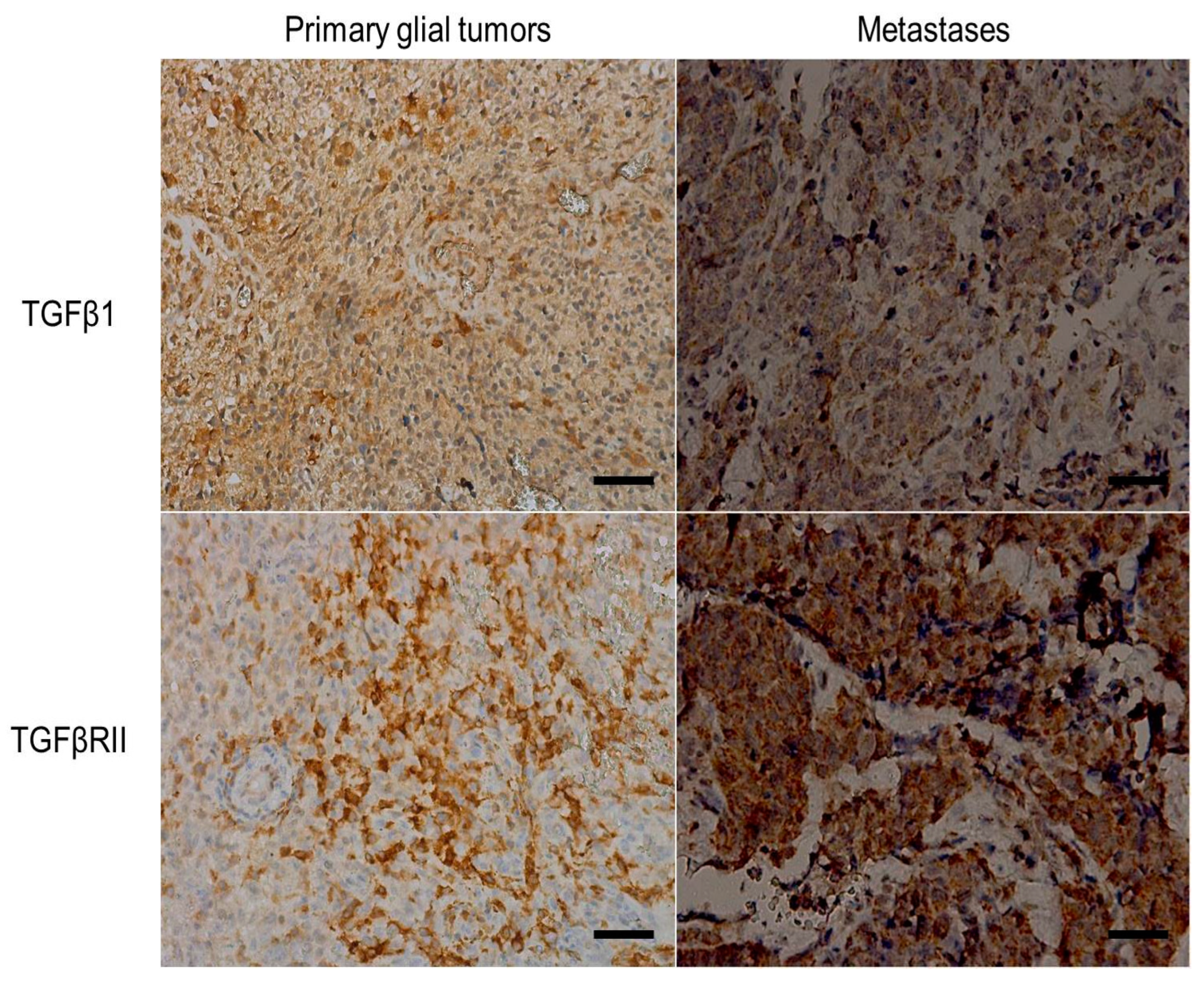
3.4. Immunohistochemistry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaminska, B.; Wesolowska, A.; Danilkiewicz, M. TGF beta signalling and its role in tumour pathogenesis. Acta Biochim. Pol. 2005, 52, 329–337. [Google Scholar] [CrossRef]
- Li, M.O.; Wan, Y.Y.; Sanjabi, S.; Robertson, A.K.; Flavell, R.A. Transforming growth factor-beta regulation of immune responses. Annu. Rev. Immunol. 2006, 24, 99–146. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.M. Transforming growth factor beta (TGF-beta) in inflammation: A cause and a cure. J. Clin. Immunol. 1992, 12, 61–74. [Google Scholar] [CrossRef]
- Rubtsov, Y.P.; Rudensky, A.Y. TGFbeta signalling in control of T-cell-mediated self-reactivity. Nat. Rev. Immunol. 2007, 7, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Reibman, J.; Meixler, S.; Lee, T.C.; Gold, L.I.; Cronstein, B.N.; Haines, K.A.; Kolasinski, S.L.; Weissmann, G. Transforming growth factor beta 1, a potent chemoattractant for human neutrophils, bypasses classic signal-transduction pathways. Proc. Natl. Acad. Sci. USA 1991, 88, 6805–6809. [Google Scholar] [CrossRef]
- Bettelli, E.; Oukka, M.; Kuchroo, V.K. T(H)-17 cells in the circle of immunity and autoimmunity. Nat Immunol. 2007, 8, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chu, X.; Meng, X.; Zou, F. Association of TGF-β1-509C/T polymorphisms with breast cancer risk: Evidence from an updated meta-analysis. Tumor Biol. 2014, 35, 935–942. [Google Scholar] [CrossRef]
- Shah, R.; Hurley, C.K.; Posch, P.E. A molecular mechanism for the differential regulation of TGF-β1 expression due to the common SNP-509C-T (c.-1347C > T). Hum Genet. 2006, 120, 461–469. [Google Scholar] [CrossRef]
- Fleming, N.I.; Jorissen, R.N.; Mouradov, D.; Christie, M.; Sakthianandeswaren, A.; Palmieri, M.; Day, F.; Li, S.; Tsui, C.; Lipton, L.; et al. SMAD2, SMAD3 and SMAD4 mutations in colorectal cancer. Cancer Res. 2013, 73, 725–735. [Google Scholar] [CrossRef]
- Biswas, S.; Chytil, A.; Washington, K.; Romero-Gallo, J.; Gorska, A.E.; Wirth, P.S.; Gautam, S.; Moses, H.L.; Grady, W.M. Transforming growth factor beta receptor type II inactivation promotes the establishment and progression of colon cancer. Cancer Res. 2004, 64, 4687–4692. [Google Scholar] [CrossRef]
- Mager, L.F.; Wasmer, M.H.; Rau, T.T.; Krebs, P. Cytokine-Induced Modulation of Colorectal Cancer. Front. Oncol. 2016, 6, 96. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Yuan, X.Q.; Zhang, C.Y.; Ye, F.; Zhou, H.F.; Li, W.L.; Liu, Z.Y.; Zhang, Y.Q.; Pan, X.; Li, G.C. High TGF-β1 expression predicts poor disease prognosis in hepatocellular carcinoma patients. Oncotarget 2017, 8, 34387–34397. [Google Scholar] [CrossRef] [PubMed]
- Merzak, A.; McCrea, S.; Koocheckpour, S.; Pilkington, G.J. Control of human glioma cell growth, migration and invasion in vitro by transforming growth factor beta 1. Br. J. Cancer 1994, 70, 199–203. [Google Scholar] [CrossRef] [PubMed]
- Saltzman, B.S.; Yamamoto, J.F.; Decker, R.; Yokochi, L.; Theriault, A.G.; Vogt, T.M.; Le Marchand, L. Association of genetic variation in the transforming growth factor beta-1 gene with serum levels and risk of colorectal neoplasia. Cancer Res. 2008, 68, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Moustakas, A.; Heldin, C.H. Signaling networks guiding epithelial-mesenchymal transitions during embryogenesis and cancer progression. Cancer Sci. 2007, 98, 1512–1520. [Google Scholar] [CrossRef]
- Kalluri, R. EMT: When epithelial cells decide to become mesenchymal-like cells. J. Clin. Investig. 2009, 119, 1417–1419. [Google Scholar] [CrossRef]
- Burghardt, I.; Schroeder, J.J.; Weiss, T.; Gramatzki, D.; Weller, M. A tumor-promoting role for soluble TβRIII in glioblastoma. Mol. Cell. Biochem. 2021, 476, 2963–2973. [Google Scholar] [CrossRef]
- Wrana, J.L.; Attisano, L.; Wieser, R.; Ventura, F.; Massague, J. Mechanism of activation of the TGF-b receptor. Nature 1994, 370, 341–347. [Google Scholar] [CrossRef]
- Dobolyi, A.; Vincze, C.; Pál, G.; Lovas, G. The neuroprotective functions of transforming growth factor beta proteins. Int. J. Mol. Sci. 2012, 13, 8219–8258. [Google Scholar] [CrossRef]
- Guo, S.K.; Shen, M.F.; Yao, H.W.; Liu, Y.S. Enhanced Expression of TGFBI Promotes the Proliferation and Migration of Glioma Cells. Cell. Physiol. Biochem. 2018, 49, 1097–1109. [Google Scholar] [CrossRef]
- Peñuelas, S.; Anido, J.; Prieto-Sanchez, R.M.; Folch, G.; Barba, I.; Cuartas, I.; Garcia-Dorado, D.; Poca, M.A.; Sahuquillo, J.; Baselga, J.; et al. TGF-β increases glioma-initiating cell self-renewal through the induction of LIF in human glioblastoma. Cancer Cell 2009, 15, 315–327. [Google Scholar] [CrossRef]
- Pickup, M.; Novitskiy, S.; Moses, H.L. The roles of TGFβ in the tumour microenvironment. Nat. Rev. Cancer. 2013, 13, 788–799. [Google Scholar] [CrossRef] [PubMed]
- Calon, A.; Tauriello, D.V.; Batlle, E. TGF-beta in CAF-mediated tumor growth and metastasis. Semin. Cancer Biol. 2014, 25, 15–22. [Google Scholar] [CrossRef]
- Gulubova, M.; Aleksandrova, E.; Vlaykova, T. Promoter polymorphisms in TGFB1 and IL10 genes influence tumor dendritic cells infiltration, development and prognosis of colorectal cancer. J. Gene Med. 2018, 20, e3005. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B.; Song, W.G.; Liu, H.Q.; Fang, F.; Xiao, Y. Role of TGFB1 polymorphism in the development of metastatic brain tumors in non-small cell lung cancer patients. Genet. Mol. Res. 2015, 14, 3545–3550. [Google Scholar] [CrossRef]
- Vieira de Castro, J.; Gonçalves, C.S.; Costa, S.; Linhares, P.; Vaz, R.; Nabiço, R.; Amorim, J.; Viana-Pereira, M.; Reis, R.M.; Costa, B.M. Impact of TGF-β1-509C/T and 869T/C polymorphisms on glioma risk and patient prognosis. Tumor Biol. 2015, 36, 6525–6532. [Google Scholar] [CrossRef]
- Yuan, X.; Wei, Q.; Komaki, R.; Liu, Z.; Yang, J.; Tucker, S.L.; Xu, T.; Heymach, J.V.; Lu, C.; Cox, J.D.; et al. TGFβ1 Polymorphisms Predict Distant Metastasis-Free Survival in Patients with Inoperable Non-Small-Cell Lung Cancer after Definitive Radiotherapy. PLoS ONE 2013, 8, e65659. [Google Scholar] [CrossRef] [PubMed]
- Ananiev, J.; Manolova, I.; Aleksandrova, E.; Gulubova, M. Impact of TGF-β1 expression and -509C>T polymorphism in the TGF-β1 gene on the progression and survival of gastric cancer. Pol. J. Pathol. 2017, 68, 234–240. [Google Scholar] [CrossRef]
- Grainger, D.J.; Heathcote, K.; Chiano, M.; Snieder, H.; Kemp, P.R.; Metcalfe, J.C.; Carter, N.D.; Spector, T.D. Genetic control of the circulating concentration of transforming growth factor type beta1. Hum. Mol. Genet. 1999, 8, 93–97. [Google Scholar] [CrossRef]
- Plaks, V.; Kong, N.; Werb, Z. The cancer stem cell niche: How essential is the niche in regulating stemness of tumor cells? Cell Stem Cell 2015, 16, 225–238. [Google Scholar] [CrossRef]
- Zhao, J.; Liang, Y.; Yin, Q.; Liu, S.; Wang, Q.; Tang, Y.; Cao, C. Clinical and prognostic significance of serum transforming growth factor-β1 levels in patients with pancreatic ductal adenocarcinoma. Braz. J. Med. Biol. Res. 2016, 49, e5485. [Google Scholar] [CrossRef]
- Lin, Y.; Kikuchi, S.; Obata, Y.; Yagyu, K. Serum levels of transforming growth factor beta1 are significantly correlated with venous invasion in patients with gastric cancer. J. Gastroenterol. Hepatol. 2006, 21, 432–437. [Google Scholar] [CrossRef]
- Xu, S.; Yang, S.; Sun, G.; Huang, W.; Zhang, Y. Transforming growth factor-beta polymorphisms and serum level in the development of osteosarcoma. DNA Cell Biol. 2014, 33, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-García, A.; Samsó, P.; Fontova, P.; Simon-Molas, H.; Manzano, A.; Castaño, E.; Rosa, J.L.; Martinez-Outshoorn, U.; Ventura, F.; Navarro-Sabaté, À.; et al. TGF-β1 targets Smad, p38 MAPK, and PI3K/Akt signaling pathways to induce PFKFB3 gene expression and glycolysis in glioblastoma cells. FEBS J. 2017, 284, 3437–3454. [Google Scholar] [CrossRef] [PubMed]
- Jurisic, D.; Erjavec, I.; Trkulja, V.; Dumic-Cule, I.; Hadzibegovic, I.; Kovacevic, L.; Svagusa, T.; Stanec, Z.; Vukicevic, S.; Grgurevic, L.; et al. Soluble type III TGF β receptor in diagnosis and follow-up of patients with breast cancer. Growth Factors 2015, 33, 200–209. [Google Scholar] [PubMed]
- Roberts, A.B.; Wakefield, L.M. The two faces of transforming growth factor β in carcinogenesis. Proc. Natl. Acad. Sci. USA 2003, 100, 8621–8623. [Google Scholar] [CrossRef]
- Principe, D.R.; DeCant, B.; Staudacher, J.; Vitello, D.; Mangan, R.J.; Wayne, E.A.; Mascariñas, E.; Diaz, A.M.; Bauer, J.; McKinney, R.D.; et al. Loss of TGFβ signaling promotes colon cancer progression and tumor-associated inflammation. Oncotarget 2017, 8, 3826–3839. [Google Scholar] [CrossRef]
- Oshima, H.; Nakayama, M.; Han, T.S.; Naoi, K.; Ju, X.; Maeda, Y.; Robine, S.; Tsuchiya, K.; Sato, T.; Sato, H.; et al. Suppressing TGFβ signaling in regenerating epithelia in an inflammatory microenvironment is sufficient to cause invasive intestinal cancer. Cancer Res. 2015, 75, 766–776. [Google Scholar] [CrossRef]
- Gu, Y.H.; Feng, Y.G. Down-regulation of TGF-β RII expression is correlated with tumor growth and invasion in non-functioning pituitary adenomas. J. Clin. Neurosci. 2018, 47, 264–268. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Number (%) |
|---|---|
| Patients | n = 61 |
| Gender | |
| Male | 40 (65.6) |
| Female | 21 (34.4) |
| Age at diagnosis (years) | |
| Range | 25–82 |
| Mean (±SD) | 62.1 (±11.76) |
| <62.1 | 27 (44.3) |
| >62.1 | 34 (55.7) |
| Primary/Metastases | |
| Primary glial tumors | 31(50.8) |
| Grade III | 2 (3.2) |
| Grade IV | 29 (47.6) |
| Metastases | 30 (49.2) |
| Chemotherapy | |
| yes | 52 (85.2) |
| no | 9 (14.8) |
| Radiotherapy | |
| yes | 28 (45.9) |
| no | 33 (54.1) |
| Survival status | n = 48 |
| alive | 15 (31.2) |
| deceased | 33 (68.8) |
| TGF-beta1 expression | n = 39 |
| positive | 19 (48.7) |
| negative | 20 (51.3) |
| TGF-beta RII expression | n = 39 |
| positive | 27 (69.2) |
| negative | 12 (30.8) |
| TGFB1-509 C/T SNP | Patients | Controls | OR (95% CI), p-Value | OR * (95% CI), p-Value | ||
|---|---|---|---|---|---|---|
| n | Frequency | n | Frequency | |||
| 61 | 176 | |||||
| Genotype | ||||||
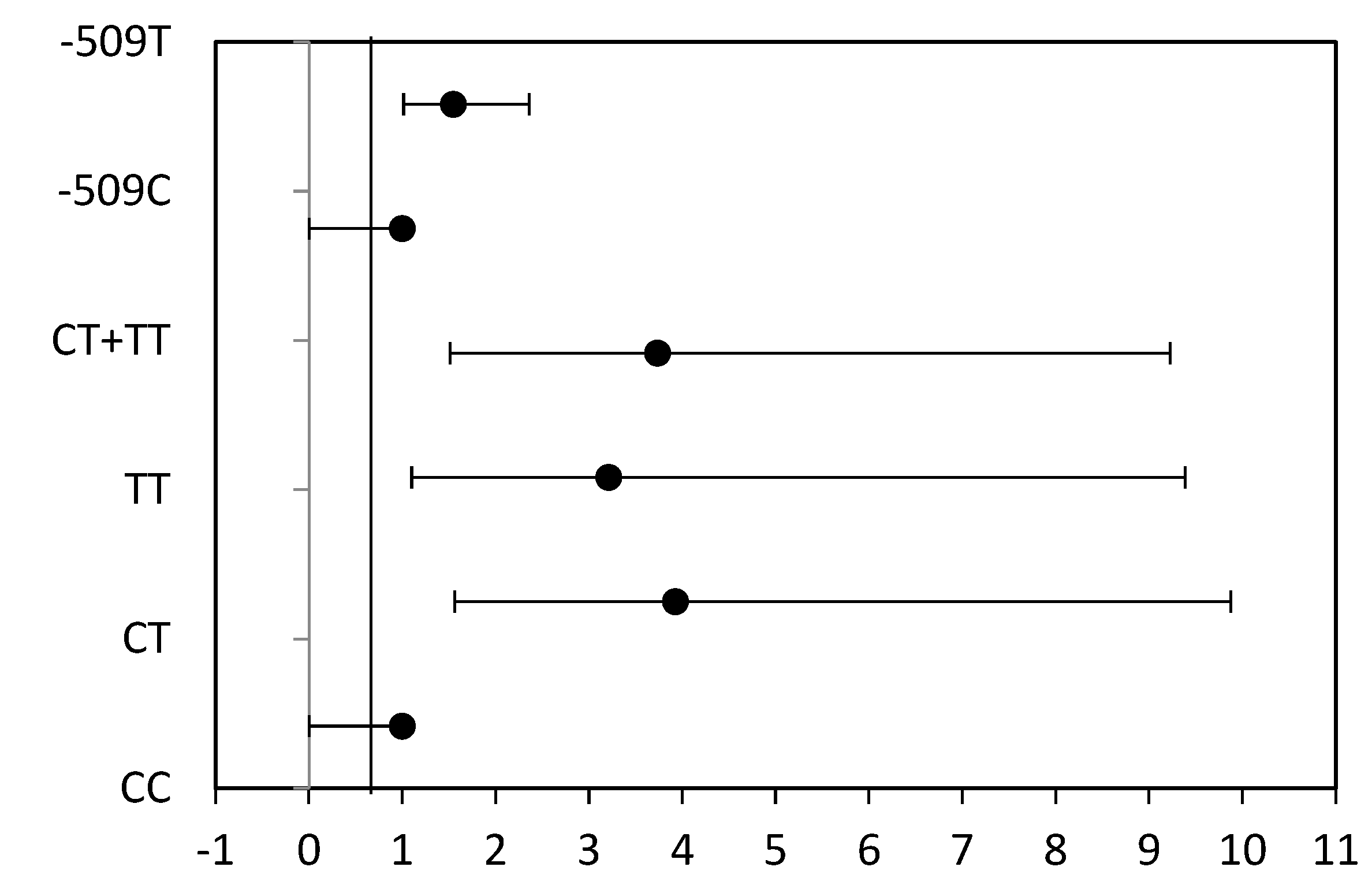
| CC | 7 | 0.10 | 53 | 0.30 | 1.0 (Ref.) | 1.0 (Ref.) |
| CT | 41 | 0.73 | 90 | 0.51 | 3.926 (1.560–9.878), p = 0.002 | 4.292 (1.092–9.685), p = 0.002 |
| TT | 13 | 0.17 | 33 | 0.19 | 3.212 (1.100–9.384), p = 0.037 | 3.215 (1.060–9.110), p = 0.033 |
| CT + TT | 54 | 0.90 | 123 | 0.70 | 3.734 (1.512–9.225), p = 0.003 | 3.968 (1.010–9.624), p = 0.003 |
| Alleles | ||||||
| -509C | 35 | 0.47 | 196 | 0.56 | 1.0 (Ref.) | … |
| -509T | 46 | 0.53 | 156 | 0.44 | 1.546 (1.014–2.358), p = 0.053 | … |
| TGF-β1 Expression | TGFβ-RII Expression | TGF-β1/TGFβ-RII Co-Expression | p | |
|---|---|---|---|---|
| Positive | 19 | 27 | 17 | 0.006 |
| Negative | 20 | 12 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aleksandrova, E.; Mindov, I.; Petrov, B.; Dimitrova, I.; Petrov, N.; Ananiev, J.; Vlaykova, T.; Valkanov, S. Role of Elevated Serum TGF-β1 and the Common Promoter TGFB1-509C/T Polymorphism in the Development and Progression of Primary Glial Tumors and Brain Metastases. Medicina 2024, 60, 146. https://doi.org/10.3390/medicina60010146
Aleksandrova E, Mindov I, Petrov B, Dimitrova I, Petrov N, Ananiev J, Vlaykova T, Valkanov S. Role of Elevated Serum TGF-β1 and the Common Promoter TGFB1-509C/T Polymorphism in the Development and Progression of Primary Glial Tumors and Brain Metastases. Medicina. 2024; 60(1):146. https://doi.org/10.3390/medicina60010146
Chicago/Turabian StyleAleksandrova, Elina, Ivan Mindov, Bozhidar Petrov, Ivelina Dimitrova, Nikolay Petrov, Julian Ananiev, Tatyana Vlaykova, and Stefan Valkanov. 2024. "Role of Elevated Serum TGF-β1 and the Common Promoter TGFB1-509C/T Polymorphism in the Development and Progression of Primary Glial Tumors and Brain Metastases" Medicina 60, no. 1: 146. https://doi.org/10.3390/medicina60010146
APA StyleAleksandrova, E., Mindov, I., Petrov, B., Dimitrova, I., Petrov, N., Ananiev, J., Vlaykova, T., & Valkanov, S. (2024). Role of Elevated Serum TGF-β1 and the Common Promoter TGFB1-509C/T Polymorphism in the Development and Progression of Primary Glial Tumors and Brain Metastases. Medicina, 60(1), 146. https://doi.org/10.3390/medicina60010146