
The RANK–RANKL–OPG System: A Multifaceted Regulator of Homeostasis, Immunity, and Cancer
 , , ,
, , , 
 , and
, and 
{kind=link}
{kind=link}
Abstract
1. Introduction to the RANK–RANKL–OPG System
2. The RANK–RANKL–OPG System
3. Functions of RANK–RANKL
3.1. Osteoclast Differentiation and Activation
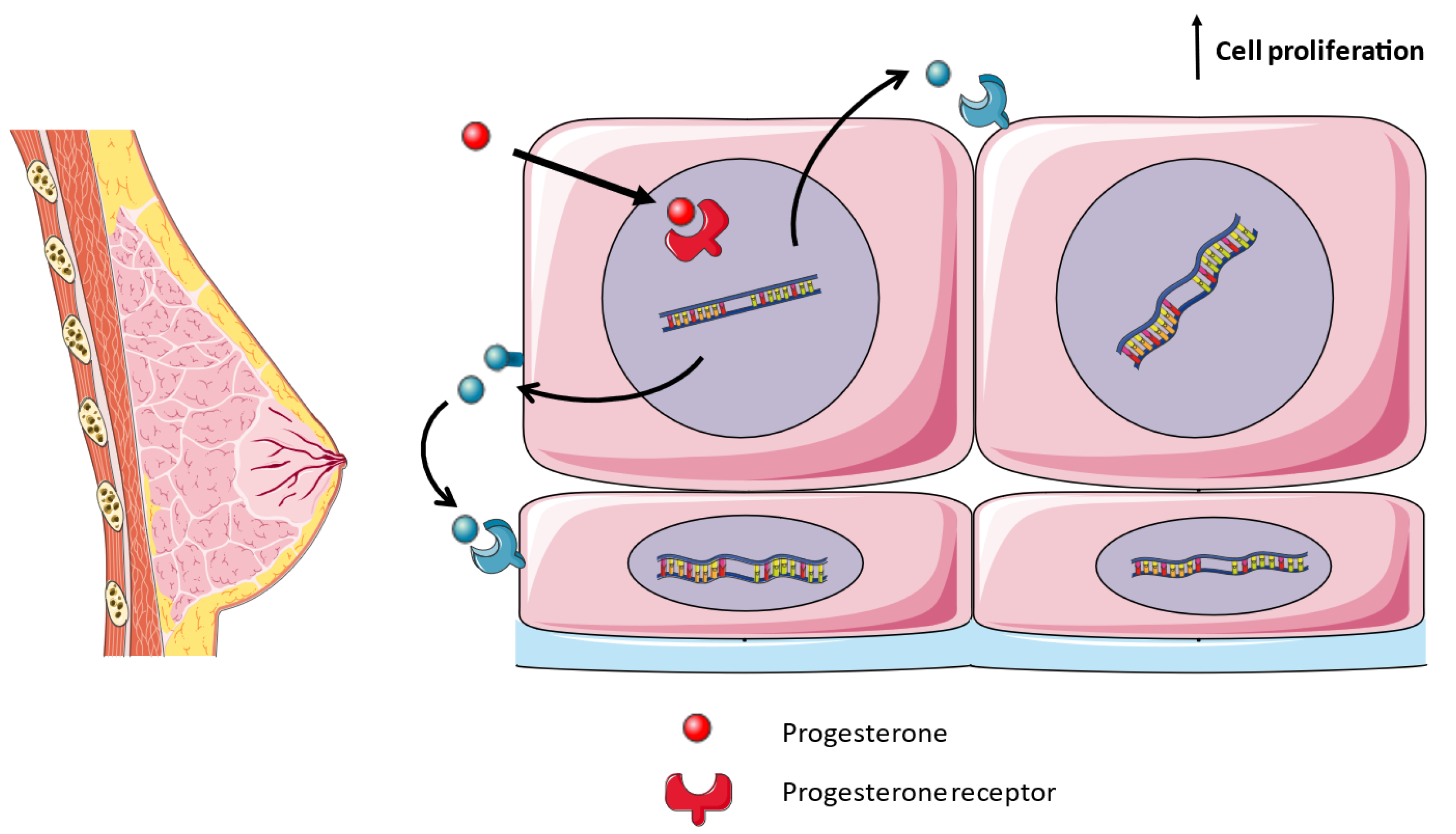
3.2. Formation of Lactating Mammary Glands
3.3. Roles in the Immune System
3.3.1. Organogenesis of Lymphoid Organs
3.3.2. Regulation of Immune Cells
3.3.3. Other Immune Functions in the Skin, Central Nervous, and Skeletal Systems
4. RANK–RANKL and Tumor Growth
4.1. Bone Metastasis
4.2. Multiple Myeloma
4.3. Breast Cancer
4.4. Urologic Cancers
4.5. Lung Cancer
5. Therapeutic Implications in Cancer
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lacey, D.L.; Timms, E.; Tan, H.; Kelley, M.J.; Dunstan, C.R.; Burgess, T.; Elliott, R.; Colombero, A.; Scully, G.E.S.; Hsu, H.; et al. Osteoprotegerin Ligand Is a Cytokine That Regulates Osteoclast Differentiation and Activation. Cell 1998, 93, 165–176. [Google Scholar] [CrossRef]
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Mochizuki, S.-I.; Tomoyasu, A.; Yano, K.; Goto, M.; Murakami, A.; et al. Osteoclast Differentiation Factor Is a Ligand for Osteoprotegerin/Osteoclastogenesis-Inhibitory Factor and Is Identical to TRANCE/RANKL. Proc. Natl. Acad. Sci. USA 1998, 95, 3597–3602. [Google Scholar] [CrossRef]
- Li, B.; Wang, P.; Jiao, J.; Wei, H.; Xu, W.; Zhou, P. Roles of the RANKL–RANK Axis in Immunity—Implications for Pathogenesis and Treatment of Bone Metastasis. Front. Immunol. 2022, 13, 824117. [Google Scholar] [CrossRef]
- Rao, S.; Cronin, S.J.F.; Sigl, V.; Penninger, J.M. RANKL and RANK: From Mammalian Physiology to Cancer Treatment. Trends Cell Biol. 2018, 28, 213–223. [Google Scholar] [CrossRef]
- Akiyama, T.; Shimo, Y.; Yanai, H.; Qin, J.; Ohshima, D.; Maruyama, Y.; Asaumi, Y.; Kitazawa, J.; Takayanagi, H.; Penninger, J.M.; et al. The Tumor Necrosis Factor Family Receptors RANK and CD40 Cooperatively Establish the Thymic Medullary Microenvironment and Self-Tolerance. Immunity 2008, 29, 423–437. [Google Scholar] [CrossRef]
- Gonzalez-suarez, E.; Jacob, A.P.; Jones, J.; Miller, R.; Roudier-meyer, M.P.; Erwert, R.; Pinkas, J.; Branstetter, D.; Dougall, W.C. RANK Ligand Mediates Progestin-Induced Mammary Epithelial Proliferation and Carcinogenesis. Nature 2010, 468, 103–109. [Google Scholar] [CrossRef]
- Hanada, R.; Leibbrandt, A.; Hanada, T.; Kitaoka, S.; Furuyashiki, T.; Fujihara, H.; Trichereau, J.; Paolino, M.; Qadri, F.; Plehm, R.; et al. Central Control of Fever and Female Body Temperature by RANKL/RANK. Nature 2009, 462, 505–509. [Google Scholar] [CrossRef]
- Duheron, V.; Hess, E.; Duval, M.; Decossas, M.; Castaned, B.; Klöpper, J.E.; Amoas, L.; Barbaroux, J.B.; Williams, I.R.; Yagita, H.; et al. Receptor Activator of NF-κB (RANK) Stimulates the Proliferation of Epithelial Cells of the Epidermo-Pilosebaceous Unit. Proc. Natl. Acad. Sci. USA 2011, 108, 5342–5347. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, J.; Liu, P.; Wang, Q.; Liu, L.; Zhao, H. The RANK/RANKL/OPG System and Tumor Bone Metastasis: Potential Mechanisms and Therapeutic Strategies. Front. Endocrinol. 2022, 13, 3320. [Google Scholar] [CrossRef]
- Pfitzner, B.M.; Branstetter, D.; Loibl, S.; Denkert, C.; Lederer, B.; Schmitt, W.D.; Dombrowski, F.; Werner, M.; Rüdiger, T.; Dougall, W.C.; et al. RANK Expression as a Prognostic and Predictive Marker in Breast Cancer. Breast Cancer Res. Treat. 2014, 145, 307–315. [Google Scholar] [CrossRef]
- Casimiro, S.; Vilhais, G.; Gomes, I.; Costa, L. The Roadmap of RANKL/RANK Pathway in Cancer. Cells 2021, 10, 1978. [Google Scholar] [CrossRef] [PubMed]
- Ono, T.; Hayashi, M.; Sasaki, F.; Nakashima, T. RANKL Biology: Bone Metabolism, the Immune System, and Beyond. Inflamm. Regen. 2020, 40, 2. [Google Scholar] [CrossRef]
- Martin, J.T. Historically Significant Events in the Discovery of RANK/RANKL/OPG. World J. Orthop. 2013, 4, 186–197. [Google Scholar] [CrossRef]
- Takayanagi, H. Osteoimmunology: Shared Mechanisms and Crosstalk between the Immune and Bone Systems. Nat. Rev. Immunol. 2007, 7, 292–304. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, G.K.; Kasi, A. Denosumab. In StatPearls—NCBI Bookshelf; StatPearls Publishing: Trasure Island, FL, USA, 2022. [Google Scholar]
- Liu, C.; Walter, T.S.; Huang, P.; Zhang, S.; Zhu, X.; Wu, Y.; Wedderburn, L.R.; Tang, P.; Owens, R.J.; Stuart, D.I.; et al. Structural and Functional Insights of RANKL–RANK Interaction and Signaling. J. Immunol. 2010, 184, 6910–6919. [Google Scholar] [CrossRef] [PubMed]
- Sigl, V.; Jones, L.P.; Penninger, J.M. RANKL/RANK: From Bone Loss to the Prevention of Breast Cancer. Open Biol. 2016, 6, 160230. [Google Scholar] [CrossRef] [PubMed]
- Renema, N.; Navet, B.; Heymann, M.F.; Lezot, F.; Heymann, D. RANK-RANKL Signalling in Cancer. Biosci. Rep. 2016, 36, e00366. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.C.; Choi, Y. Biology of the RANKL-RANK-OPG System in Immunity, Bone, and Beyond. Front. Immunol. 2014, 5, 511. [Google Scholar] [CrossRef]
- Nelson, C.A.; Warren, J.T.; Wang, M.W.H.; Teitelbaum, S.L.; Fremont, D.H. RANKL Employs Distinct Binding Modes to Engage RANK and the Osteoprotegerin Decoy Receptor. Structure 2012, 20, 1971–1982. [Google Scholar] [CrossRef]
- O’Brien, C.A. Control of RANKL Gene Expression. Bone 2010, 46, 911–919. [Google Scholar] [CrossRef]
- Elango, J.; Bao, B.; Wu, W. The Hidden Secrets of Soluble RANKL in Bone Biology. Cytokine 2021, 144, 155559. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Cawley, K.; Piemontese, M.; Fujiwara, Y.; Zhao, H.; Goellner, J.J.; O’Brien, C.A. Soluble RANKL Contributes to Osteoclast Formation in Adult Mice but Not Ovariectomy-Induced Bone Loss. Nat. Commun. 2018, 9, 2909. [Google Scholar] [CrossRef] [PubMed]
- Munasinghe, A.; Lin, P.; Colina, C.M. Unraveling Binding Interactions between Human RANKL and Its Decoy Receptor Osteoprotegerin. J. Phys. Chem. B 2017, 121, 9141–9148. [Google Scholar] [CrossRef]
- Tat, S.K.; Padrines, M.; Theoleyre, S.; Couillaud-Battaglia, S.; Heymann, D.; Redini, F.; Fortun, Y. OPG/Membranous-RANKL Complex Is Internalized via the Clathrin Pathway before a Lysosomal and a Proteasomal Degradation. Bone 2006, 39, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Yang, Z.; Ma, Y.; Yue, Z.; Lin, H.; Qu, G.; Huang, J.; Dai, W.; Li, C.; Zheng, C.; et al. LGR4 Is a Receptor for RANKL and Negatively Regulates Osteoclast Differentiation and Bone Resorption. Nat. Med. 2016, 22, 539–546. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, J.A.; Partridge, N.C. Physiological Bone Remodeling: Systemic Regulation and Growth Factor Involvement. Physiology 2016, 31, 233–245. [Google Scholar] [CrossRef]
- Wang, L.; You, X.; Zhang, L.; Zhang, C.; Zou, W. Mechanical Regulation of Bone Remodeling. Bone Res. 2022, 10, 16. [Google Scholar] [CrossRef]
- Rowe, P.; Koller, A.; Sharma, S. Physiology, Bone Remodeling. In StatPearls; StatPearls Publishing: Trasure Island, FL, USA, 2023. [Google Scholar]
- Jimi, E.; Akiyama, S.; Tsurukai, T.; Okahashi, N.; Kobayashi, K.; Udagawa, N.; Nishihara, T.; Takahashi, N.; Suda, T. Osteoclast Differentiation Factor Acts as a Multifunctional Regulator in Murine Osteoclast Differentiation and Function. J. Immunol. 1999, 163, 434–442. [Google Scholar] [CrossRef]
- Udagawa, N.; Koide, M.; Nakamura, M.; Nakamichi, Y.; Yamashita, T.; Uehara, S.; Kobayashi, Y.; Furuya, Y.; Yasuda, H.; Fukuda, C.; et al. Osteoclast Differentiation by RANKL and OPG Signaling Pathways. J. Bone Miner. Metab. 2021, 39, 19–26. [Google Scholar] [CrossRef]
- Takegahara, N.; Kim, H.; Choi, Y. RANKL Biology. Bone 2022, 159, 116353. [Google Scholar] [CrossRef]
- Nakashima, T.; Hayashi, M.; Fukunaga, T.; Kurata, K.; Oh-Hora, M.; Feng, J.Q.; Bonewald, L.F.; Kodama, T.; Wutz, A.; Wagner, E.F.; et al. Evidence for Osteocyte Regulation of Bone Homeostasis through RANKL Expression. Nat. Med. 2011, 17, 1231–1234. [Google Scholar] [CrossRef] [PubMed]
- Morrell, A.E.; Brown, G.N.; Robinson, S.T.; Sattler, R.L.; Baik, A.D.; Zhen, G.; Cao, X.; Bonewald, L.F.; Jin, W.; Kam, L.C.; et al. Mechanically Induced Ca2+ Oscillations in Osteocytes Release Extracellular Vesicles and Enhance Bone Formation. Bone Res. 2018, 6, 6. [Google Scholar] [CrossRef]
- Bolamperti, S.; Villa, I.; Rubinacci, A. Bone Remodeling: An Operational Process Ensuring Survival and Bone Mechanical Competence. Bone Res. 2022, 10, 48. [Google Scholar] [CrossRef] [PubMed]
- Honma, M.; Ikebuchi, Y.; Kariya, Y.; Hayashi, M.; Hayashi, N.; Aoki, S.; Suzuki, H. RANKL Subcellular Trafficking and Regulatory Mechanisms in Osteocytes. J. Bone Miner. Res. 2013, 28, 1936–1949. [Google Scholar] [CrossRef]
- Al-Dujaili, S.A.; Lau, E.; Al-Dujaili, H.; Tsang, K.; Guenther, A.; You, L. Apoptotic Osteocytes Regulate Osteoclast Precursor Recruitment and Differentiation in Vitro. J. Cell. Biochem. 2011, 112, 2412–2423. [Google Scholar] [CrossRef]
- Honma, M.; Ikebuchi, Y.; Suzuki, H. Mechanisms of RANKL Delivery to the Osteoclast Precursor Cell Surface. J. Bone Miner. Metab. 2021, 39, 27–33. [Google Scholar] [CrossRef]
- Chen, X.; Zhi, X.; Wang, J.; Su, J. RANKL Signaling in Bone Marrow Mesenchymal Stem Cells Negatively Regulates Osteoblastic Bone Formation. Bone Res. 2018, 6, 34. [Google Scholar] [CrossRef] [PubMed]
- Ikebuchi, Y.; Aoki, S.; Honma, M.; Hayashi, M.; Sugamori, Y.; Khan, M.; Kariya, Y.; Kato, G.; Tabata, Y.; Penninger, J.M.; et al. Coupling of Bone Resorption and Formation by RANKL Reverse Signalling. Nature 2018, 561, 195–200. [Google Scholar] [CrossRef]
- Alex, A.; Bhandary, E.; McGuire, K.P. Anatomy and Physiology of the Breast during Pregnancy and Lactation. In Diseases of the Breast during Pregnancy and Lactation. Advances in Experimental Medicine and Biology; Alipour, S., Omranipour, R., Eds.; Springer: Cham, Switzerland, 2020; Volume 1252, pp. 3–7. ISBN 9783030415969. [Google Scholar]
- Fata, J.E.; Kong, Y.Y.; Li, J.; Sasaki, T.; Irie-Sasaki, J.; Moorehead, R.A.; Elliott, R.; Scully, S.; Voura, E.B.; Lacey, D.L.; et al. The Osteoclast Differentiation Factor Osteoprotegerin-Ligand Is Essential for Mammary Gland Development. Cell 2000, 103, 41–50. [Google Scholar] [CrossRef]
- Azim, H.A.; Peccatori, F.A.; Brohée, S.; Branstetter, D.; Loi, S.; Viale, G.; Piccart, M.; Dougall, W.C.; Pruneri, G.; Sotiriou, C. RANK-Ligand (RANKL) Expression in Young Breast Cancer Patients and during Pregnancy. Breast Cancer Res. 2015, 17, 24. [Google Scholar] [CrossRef]
- Joshi, P.A.; Jackson, H.W.; Beristain, A.G.; Di Grappa, M.A.; Mote, P.A.; Clarke, C.L.; Stingl, J.; Waterhouse, P.D.; Khokha, R. Progesterone Induces Adult Mammary Stem Cell Expansion. Nature 2010, 465, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Asselin-Labat, M.L.; Vaillant, F.; Sheridan, J.M.; Pal, B.; Wu, D.; Simpson, E.R.; Yasuda, H.; Smyth, G.K.; Martin, T.J.; Lindeman, G.J.; et al. Control of Mammary Stem Cell Function by Steroid Hormone Signalling. Nature 2010, 465, 798–802. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Bonizzi, G.; Seagroves, T.N.; Greten, F.R.; Johnson, R.; Schmidt, E.V.; Karin, M. IKKα Provides an Essential Link between RANK Signaling and Cyclin D1 Expression during Mammary Gland Development. Cell 2001, 107, 763–775. [Google Scholar] [CrossRef] [PubMed]
- Joshi, P.A.; Waterhouse, P.D.; Kannan, N.; Narala, S.; Fang, H.; Di Grappa, M.A.; Jackson, H.W.; Penninger, J.M.; Eaves, C.; Khokha, R. RANK Signaling Amplifies WNT-Responsive Mammary Progenitors through R-SPONDIN1. Stem Cell Rep. 2015, 5, 31–44. [Google Scholar] [CrossRef]
- Wunderle, M.; Ruebner, M.; Häberle, L.; Schwenke, E.; Hack, C.C.; Bayer, C.M.; Koch, M.C.; Schwitulla, J.; Schulz-Wendtland, R.; Kozieradzki, I.; et al. RANKL and OPG and Their Influence on Breast Volume Changes during Pregnancy in Healthy Women. Sci. Rep. 2020, 10, 5171. [Google Scholar] [CrossRef]
- Dougall, W.C.; Glaccum, M.; Charrier, K.; Rohrbach, K.; Brasel, K.; De Smedt, T.; Daro, E.; Smith, J.; Tometsko, M.E.; Maliszewski, C.R.; et al. RANK Is Essential for Osteoclast and Lymph Node Development. Genes Dev. 1999, 13, 2412–2424. [Google Scholar] [CrossRef]
- Perlot, T.; Penninger, J.M. Development and Function of Murine B Cells Lacking RANK. J. Immunol. 2012, 188, 1201–1205. [Google Scholar] [CrossRef]
- Kanaya, T.; Sakakibara, S.; Jinnohara, T.; Hachisuka, M.; Tachibana, N.; Hidano, S.; Kobayashi, T.; Kimura, S.; Iwanaga, T.; Nakagawa, T.; et al. Development of Intestinal M Cells and Follicle-Associated Epithelium Is Regulated by TRAF6-Mediated NF-κB Signaling. J. Exp. Med. 2018, 215, 501–519. [Google Scholar] [CrossRef]
- Kim, D.; Mebius, R.E.; MacMicking, J.D.; Jung, S.; Cupedo, T.; Castellanos, Y.; Rho, J.; Wong, B.R.; Josien, R.; Kim, N.; et al. Regulation of Peripheral Lymph Node Genesis by the Tumor Necrosis Factor Family Member TRANCE. J. Exp. Med. 2000, 192, 1467–1478. [Google Scholar] [CrossRef]
- Mueller, C.G.; Hess, E. Emerging Functions of RANKL in Lymphoid Tissues. Front. Immunol. 2012, 3, 261. [Google Scholar] [CrossRef]
- Cupedo, T.; Mebius, R.E. Cellular Interactions in Lymph Node Development. J. Immunol. 2005, 174, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Pei, X.Y.; Shen, H.; Gao, Y.N.; Sun, X.Y.; Wang, W.; Ge, Q.; Zhang, Y. Thymic Homing of Activated CD4+ T Cells Induces Degeneration of the Thymic Epithelium through Excessive RANK Signaling. Sci. Rep. 2017, 7, 2421. [Google Scholar] [CrossRef] [PubMed]
- Irla, M. RANK Signaling in the Differentiation and Regeneration of Thymic Epithelial Cells. Front. Immunol. 2021, 11, 623265. [Google Scholar] [CrossRef] [PubMed]
- Knoop, K.A.; Kumar, N.; Butler, B.R.; Sakthivel, S.K.; Taylor, R.T.; Nochi, T.; Akiba, H.; Yagita, H.; Kiyono, H.; Williams, I.R. RANKL Is Necessary and Sufficient to Initiate Development of Antigen-Sampling M Cells in the Intestinal Epithelium. J. Immunol. 2009, 183, 5738–5747. [Google Scholar] [CrossRef]
- Akiyama, T.; Shinzawa, M.; Akiyama, N. RANKL-RANK Interaction in Immune Regulatory Systems. World J. Orthop. 2012, 3, 142–150. [Google Scholar] [CrossRef]
- Walsh, M.C.; Choi, Y. Regulation of T Cell-Associated Tissues and T Cell Activation by RANKL-RANK-OPG. J. Bone Miner. Metab. 2021, 39, 54–63. [Google Scholar] [CrossRef]
- Ma, D.Y.; Clark, E.A. The Role of CD40 and CD154/CD40L in Dendritic Cells. Semin. Immunol. 2009, 21, 265–272. [Google Scholar] [CrossRef]
- Knoop, K.A.; Butler, B.R.; Kumar, N.; Newberry, R.D.; Williams, I.R. Distinct Developmental Requirements for Isolated Lymphoid Follicle Formation in the Small and Large Intestine: RANKL Is Essential Only in the Small Intestine. Am. J. Pathol. 2011, 179, 1861–1871. [Google Scholar] [CrossRef]
- Mota, R.F.; Cavalcanti De Araújo, P.H.; Cezine, M.E.R.; Matsuo, F.S.; Metzner, R.J.M.; Oliveira De Biagi Junior, C.A.; Peronni, K.C.; Hayashi, H.; Shimamura, M.; Nakagami, H.; et al. RANKL Impairs the TLR4 Pathway by Increasing TRAF6 and RANK Interaction in Macrophages. Biomed Res. Int. 2022, 2022, 7740079. [Google Scholar] [CrossRef]
- Loser, K.; Mehling, A.; Loeser, S.; Apelt, J.; Kuhn, A.; Grabbe, S.; Schwarz, T.; Penninger, J.M.; Beissert, S. Epidermal RANKL Controls Regulatory T-Cell Numbers via Activation of Dendritic Cells. Nat. Med. 2006, 12, 1372–1379. [Google Scholar] [CrossRef]
- Guerrini, M.M.; Okamoto, K.; Komatsu, N.; Sawa, S.; Danks, L.; Penninger, J.M.; Nakashima, T.; Takayanagi, H. Inhibition of the TNF Family Cytokine RANKL Prevents Autoimmune Inflammation in the Central Nervous System. Immunity 2015, 43, 1174–1185. [Google Scholar] [CrossRef] [PubMed]
- Shimamura, M.; Nakagami, H.; Osako, M.K.; Kurinami, H.; Koriyama, H.; Zhengda, P.; Tomioka, H.; Tenma, A.; Wakayama, K.; Morishita, R. OPG/RANKL/RANK Axis Is a Critical Inflammatory Signaling System in Ischemic Brain in Mice. Proc. Natl. Acad. Sci. USA 2014, 111, 8191–8196. [Google Scholar] [CrossRef] [PubMed]
- Biscetti, F.; Giovannini, S.; Straface, G.; Bertucci, F.; Angelini, F.; Porreca, C.; Landolfi, R.; Flex, A. RANK/RANKL/OPG Pathway: Genetic Association with History of Ischemic Stroke in Italian Population. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 4574–4580. [Google Scholar]
- Park, M.S.; Park, J.H.; Joo, A.; Chang, Y.; Song, T.J. The Association of Plasma Osteoprotegerin Levels and Functional Outcomes Post Endovascular Thrombectomy in Acute Ischemic Stroke Patients: A Retrospective Observational Study. PeerJ 2022, 10, e13327. [Google Scholar] [CrossRef]
- Song, T.J.; Kim, J.; Yang, S.H.; Park, J.H.; Lee, H.S.; Nam, C.M.; Lee, O.H.; Kim, Y.D.; Nam, H.S.; Heo, J.H. Association of Plasma Osteoprotegerin Levels with Stroke Severity and Functional Outcome in Acute Ischaemic Stroke Patients. Biomarkers 2012, 17, 738–744. [Google Scholar] [CrossRef]
- Nagy, V.; Penninger, J.M. The RANKL-RANK Story. Gerontology 2015, 61, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Baud’huin, M.; Lamoureux, F.; Duplomb, L.; Rédini, F.; Heymann, D. RANKL, RANK, Osteoprotegerin: Key Partners of Osteoimmunology and Vascular Diseases. Cell. Mol. Life Sci. 2007, 64, 2334–2350. [Google Scholar] [CrossRef]
- Nakashima, T.; Takayanagi, H. Osteoimmunology: Crosstalk between the Immune and Bone Systems. J. Clin. Immunol. 2009, 29, 555–567. [Google Scholar] [CrossRef]
- Okamoto, K.; Nakashima, T.; Shinohara, M.; Negishi-Koga, T.; Komatsu, N.; Terashima, A.; Sawa, S.; Nitta, T.; Takayanagi, H. Osteoimmunology: The Conceptual Framework Unifying the Immune and Skeletal Systems. Physiol. Rev. 2017, 97, 1295–1349. [Google Scholar] [CrossRef]
- Guo, Q.; Wang, Y.; Xu, D.; Nossent, J.; Pavlos, N.J.; Xu, J. Rheumatoid Arthritis: Pathological Mechanisms and Modern Pharmacologic Therapies. Bone Res. 2018, 6, 15. [Google Scholar] [CrossRef]
- Geusens, P. The Role of RANK Ligand/Osteoprotegerin in Rheumatoid Arthritis. Ther. Adv. Musculoskelet. Dis. 2012, 4, 225–233. [Google Scholar] [CrossRef]
- Danks, L.; Komatsu, N.; Guerrini, M.M.; Sawa, S.; Armaka, M.; Kollias, G.; Nakashima, T.; Takayanagi, H. RANKL Expressed on Synovial Fibroblasts Is Primarily Responsible for Bone Erosions during Joint Inflammation. Ann. Rheum. Dis. 2016, 75, 1187–1195. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, N.; Okamoto, K.; Sawa, S.; Nakashima, T.; Oh-Hora, M.; Kodama, T.; Tanaka, S.; Bluestone, J.A.; Takayanagi, H. Pathogenic Conversion of Foxp3 + T Cells into TH17 Cells in Autoimmune Arthritis. Nat. Med. 2014, 20, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, M.; Rinotas, V.; Violitzi, F.; Thireou, T.; Panayotou, G.; Samiotaki, M.; Douni, E. New Insights for RANKL as a Proinflammatory Modulator in Modeled Inflammatory Arthritis. Front. Immunol. 2019, 10, 97. [Google Scholar] [CrossRef]
- Tanaka, S. RANKL Is a Therapeutic Target of Bone Destruction in Rheumatoid Arthritis. F1000Research 2019, 8, Rev-533. [Google Scholar] [CrossRef] [PubMed]
- Jayarangaiah, A.; Kemp, A.K.; Kariyanna, P.T. Bone Metastasis. In StatPearls—NCBI Bookshelf; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Macedo, F.; Ladeira, K.; Pinho, F.; Saraiva, N.; Bonito, N.; Pinto, L.; Gonçalves, F. Bone Metastases: An Overview. Oncol. Rev. 2017, 11, 321. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.H.; Nakashima, T.; Sanchez, O.H.; Kozieradzki, I.; Komarova, S.V.; Sarosi, I.; Morony, S.; Rubin, E.; Sarao, R.; Hojilla, C.V.; et al. Regulation of Cancer Cell Migration and Bone Metastasis by RANKL. Nature 2006, 440, 8–12. [Google Scholar] [CrossRef]
- Ricarte, F.R.; Le Henaff, C.; Kolupaeva, V.G.; Gardella, T.J.; Partridge, N.C. Parathyroid Hormone(1–34) and Its Analogs Differentially Modulate Osteoblastic Rankl Expression via PKA/SIK2/SIK3 and PP1/PP2A–CRTC3 Signaling. J. Biol. Chem. 2018, 293, 20200–20213. [Google Scholar] [CrossRef]
- Othman, A.; Winogradzki, M.; Lee, L.; Tandon, M.; Blank, A.; Pratap, J. Bone Metastatic Breast Cancer: Advances in Cell Signaling and Autophagy Related Mechanisms. Cancers 2021, 13, 4310. [Google Scholar] [CrossRef]
- Ming, J.; Cronin, S.J.F.; Penninger, J.M. Targeting the RANKL/RANK/OPG Axis for Cancer Therapy. Front. Oncol. 2020, 10, 1283. [Google Scholar] [CrossRef]
- Raje, N.S.; Bhatta, S.; Terpos, E. Role of the RANK/RANKL Pathway in Multiple Myeloma. Clin. Cancer Res. 2019, 25, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Terpos, E.; Ntanasis-Stathopoulos, I.; Gavriatopoulou, M.; Dimopoulos, M.A. Pathogenesis of Bone Disease in Multiple Myeloma: From Bench to Bedside. Blood Cancer J. 2018, 8, 7. [Google Scholar] [CrossRef]
- Kassen, D.; Lath, D.; Lach, A.; Evans, H.; Chantry, A.; Rabin, N.; Croucher, P.; Yong, K.L. Myeloma Impairs Mature Osteoblast Function but Causes Early Expansion of Osteo-Progenitors: Temporal Changes in Bone Physiology and Gene Expression in the KMS12BM Model. Br. J. Haematol. 2016, 172, 64–79. [Google Scholar] [CrossRef] [PubMed]
- Sfiridaki, K.; Pappa, C.A.; Tsirakis, G.; Kanellou, P.; Kaparou, M.; Stratinaki, M.; Sakellaris, G.; Kontakis, G.; Alexandrakis, M.G. Angiogenesis-Related Cytokines, RANKL, and Osteoprotegerin in Multiple Myeloma Patients in Relation to Clinical Features and Response to Treatment. Mediators Inflamm. 2011, 2011, 867576. [Google Scholar] [CrossRef][Green Version]
- Ney, J.; Fehm, T.; Juhasz-Boess, I.; Solomayer, E. RANK, RANKL and OPG Expression in Breast Cancer—Influence on Osseous Metastasis. Geburtshilfe Frauenheilkd. 2012, 72, 385–391. [Google Scholar] [CrossRef]
- Trabert, B.; Sherman, M.E.; Kannan, N.; Stanczyk, F.Z. Progesterone and Breast Cancer. Endocr. Rev. 2019, 41, 320–344. [Google Scholar] [CrossRef]
- Kiesel, L.; Kohl, A. Role of the RANK/RANKL Pathway in Breast Cancer. Maturitas 2016, 86, 10–16. [Google Scholar] [CrossRef]
- Fu, X.; Tan, W.; Song, Q.; Pei, H.; Li, J. BRCA1 and Breast Cancer: Molecular Mechanisms and Therapeutic Strategies. Front. Cell Dev. Biol. 2022, 10, 813457. [Google Scholar] [CrossRef] [PubMed]
- Widschwendter, M.; Rosenthal, A.N.; Philpott, S.; Rizzuto, I.; Fraser, L.; Hayward, J.; Intermaggio, M.P.; Edlund, C.K.; Ramus, S.J.; Gayther, S.A.; et al. The Sex Hormone System in Carriers of BRCA1/2 Mutations: A Case-Control Study. Lancet Oncol. 2013, 14, 1226–1232. [Google Scholar] [CrossRef]
- Sigl, V.; Owusu-boaitey, K.; Joshi, P.A.; Kavirayani, A.; Wirnsberger, G.; Novatchkova, M.; Kozieradzki, I.; Schramek, D.; Edokobi, N.; Hersl, J.; et al. RANKL/RANK Control Brca1 Mutation-Driven Mammary Tumors. Cell Res. 2016, 26, 761–774. [Google Scholar] [CrossRef]
- Ye, X.; Weinberg, R.A. Epithelial-Mesenchymal Plasticity: A Central Regulator of Cancer Progression. Trends Cell Biol. 2015, 25, 675–686. [Google Scholar] [CrossRef]
- Palafox, M.; Ferrer, I.; Pellegrini, P.; Vila, S.; Hernandez-Ortega, S.; Urruticoechea, A.; Climent, F.; Soler, M.T.; Muñoz, P.; Viñals, F.; et al. RANK Induces Epithelial-Mesenchymal Transition and Stemness in Human Mammary Epithelial Cells and Promotes Tumorigenesis and Metastasis. Cancer Res. 2012, 72, 2879–2888. [Google Scholar] [CrossRef]
- Tsubaki, M.; Komai, M.; Fujimoto, S.I.; Itoh, T.; Imano, M.; Sakamoto, K.; Shimaoka, H.; Takeda, T.; Ogawa, N.; Mashimo, K.; et al. Activation of NF-κB by the RANKL/RANK System up-Regulates Snail and Twist Expressions and Induces Epithelial-to-Mesenchymal Transition in Mammary Tumor Cell Lines. J. Exp. Clin. Cancer Res. 2013, 32, 62. [Google Scholar] [CrossRef]
- Benítez, S.; Cordero, A.; Santamaría, P.G.; Redondo-Pedraza, J.; Rocha, A.S.; Collado-Solé, A.; Jimenez, M.; Sanz-Moreno, A.; Yoldi, G.; Santos, J.C.; et al. RANK Links Senescence to Stemness in the Mammary Epithelia, Delaying Tumor Onset but Increasing Tumor Aggressiveness. Dev. Cell 2021, 56, 1727–1741.e7. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Zhang, W.; Strasner, A.; Grivennikov, S.; Cheng, J.Q.; Hoffman, R.M.; Karin, M. Tumour-Infiltrating Regulatory T Cells Stimulate Mammary Cancer Metastasis through RANKL-RANK Signalling. Nature 2011, 470, 548–553. [Google Scholar] [CrossRef]
- Ohtaka, M.; Kawahara, T.; Mochizuki, T.; Takamoto, D.; Hattori, Y.; Teranishi, J.-i.; Miyoshi, Y.; Yumura, Y.; Hasumi, H.; Yokomizo, Y.; et al. RANK/RANKL Expression in Prostate Cancer. Int. J. Surg. Case Rep. 2017, 30, 106–107. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Sircar, K.; Aprikian, A.; Potti, A.; Goltzman, D.; Rabbani, S.A. Expression of RANKL/RANK/OPG in Primary and Metastatic Human Prostate Cancer as Markers of Disease Stage and Functional Regulation. Cancer 2006, 107, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Casimiro, S.; Mohammad, K.S.; Pires, R.; Tato-Costa, J.; Alho, I.; Teixeira, R.; Carvalho, A.; Ribeiro, S.; Lipton, A.; Guise, T.A.; et al. RANKL/RANK/MMP-1 Molecular Triad Contributes to the Metastatic Phenotype of Breast and Prostate Cancer Cells In Vitro. PLoS ONE 2013, 8, e63153. [Google Scholar] [CrossRef]
- Mikami, S.; Katsube, K.; Oya, M.; Ishida, M.; Kosaka, T.; Mizuno, R.; Mochizuki, S.; Ikeda, T.; Mukai, M.; Okada, Y. Increased RANKL Expression Is Related to Tumour Migration and Metastasis of Renal Cell Carcinomas. J. Pathol. 2009, 218, 530–539. [Google Scholar] [CrossRef]
- Ohtaka, M.; Kawahara, T.; Ishiguro, Y.; Sharma, M.; Yao, M.; Miyamoto, H.; Uemura, H. Expression of Receptor Activator of Nuclear Factor Kappa B Ligand in Bladder Cancer. Int. J. Urol. 2018, 25, 901–902. [Google Scholar] [CrossRef]
- de Souza Sasaki, B.R.; Reis, I.B.; Oliveira, G.; Duran, N.; Fávaro, W.J. Modulation of the RANK/RANKL/OPG System and FOXP3+ Regulatory T Cells in the Tumor Microenvironment of Noninvasive Bladder Cancer after Intravesical Oncotherad Immunotherapy Associated with Platelet-Rich Plasma. J. Clin. Oncol. 2021, 39, 462. [Google Scholar] [CrossRef]
- Faget, J.; Contat, C.; Zangger, N.; Peters, S.; Meylan, E. RANKL Signaling Sustains Primary Tumor Growth in Genetically Engineered Mouse Models of Lung Adenocarcinoma. J. Thorac. Oncol. 2018, 13, 387–398. [Google Scholar] [CrossRef]
- Rao, S.; Sigl, V.; Wimmer, R.A.; Novatchkova, M.; Jais, A.; Wagner, G.; Handschuh, S.; Uribesalgo, I.; Hagelkruys, A.; Kozieradzki, I.; et al. RANK Rewires Energy Homeostasis in Lung Cancer Cells and Drives Primary Lung Cancer. Genes Dev. 2017, 31, 2099–2111. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Lara, V.; Hernandez-Martinez, J.M.; Arrieta, O. Influence of Estrogen in Non-Small Cell Lung Cancer and Its Clinical Implications. J. Thorac. Dis. 2018, 10, 482–497. [Google Scholar] [CrossRef]
- Peng, X.; Guo, W.; Ren, T.; Lou, Z.; Lu, X.; Zhang, S.; Lu, Q.; Sun, Y. Differential Expression of the RANKL/RANK/OPG System Is Associated with Bone Metastasis in Human Non-Small Cell Lung Cancer. PLoS ONE 2013, 8, e58361. [Google Scholar] [CrossRef]
- Scagliotti, G.V.; Hirsh, V.; Siena, S.; Henry, D.H.; Woll, P.J.; Manegold, C.; Solal-Celigny, P.; Rodriguez, G.; Krzakowski, M.; Mehta, N.D.; et al. Overall Survival Improvement in Patients with Lung Cancer and Bone Metastases Treated with Denosumab versus Zoledronic Acid: Subgroup Analysis from a Randomized Phase 3 Study. J. Thorac. Oncol. 2012, 7, 1823–1829. [Google Scholar] [CrossRef] [PubMed]
- Body, J.J.; Greipp, P.; Coleman, R.E.; Facon, T.; Geurs, F.; Fermand, J.P.; Harousseau, J.L.; Lipton, A.; Mariette, X.; Williams, C.D.; et al. A Phase I Study of AMGN-0007, a Recombinant Osteoprotegerin Construct, in Patients with Multiple Myeloma or Breast Carcinoma Related Bone Metastases. Cancer 2003, 97, 887–892. [Google Scholar] [CrossRef]
- Lipton, A.; Goessl, C. Clinical Development of Anti-RANKL Therapies for Treatment and Prevention of Bone Metastasis. Bone 2011, 48, 96–99. [Google Scholar] [CrossRef]
- Lacey, D.L.; Boyle, W.J.; Simonet, W.S.; Kostenuik, P.J.; Dougall, W.C.; Sullivan, J.K.; Martin, J.S.; Dansey, R. Bench to Bedside: Elucidation of the OPG-RANK-RANKL Pathway and the Development of Denosumab. Nat. Rev. Drug Discov. 2012, 11, 401–419. [Google Scholar] [CrossRef]
- Kearns, A.E.; Khosla, S.; Kostenuik, P.J. Receptor Activator of Nuclear Factor κB Ligand and Osteoproteger in Regulation of Bone Remodeling in Health and Disease. Endocr. Rev. 2008, 29, 155–192. [Google Scholar] [CrossRef]
- Lu, J.; Hu, D.; Zhang, Y.; Ma, C. Current Comprehensive Understanding of Denosumab (the RANKL Neutralizing Antibody) in the Treatment of Bone Metastasis of Malignant Tumors, Including Pharmacological Mechanism and Clinical Trials. Front. Oncol. 2023, 13, 1133828. [Google Scholar] [CrossRef] [PubMed]
- Peddi, P.; Lopez-Olivo, M.A.; Pratt, G.F.; Suarez-Almazor, M.E. Denosumab in Patients with Cancer and Skeletal Metastases: A Systematic Review and Meta-Analysis. Cancer Treat. Rev. 2013, 39, 97–104. [Google Scholar] [CrossRef] [PubMed]
- de Groot, A.F.; Appelman-Dijkstra, N.M.; van der Burg, S.H.; Kroep, J.R. The Anti-Tumor Effect of RANKL Inhibition in Malignant Solid Tumors—A Systematic Review. Cancer Treat. Rev. 2018, 62, 18–28. [Google Scholar] [CrossRef] [PubMed]
- De Castro, J.; García, R.; Garrido, P.; Isla, D.; Massuti, B.; Blanca, B.; Vázquez, J. Therapeutic Potential of Denosumab in Patients with Lung Cancer: Beyond Prevention of Skeletal Complications. Clin. Lung Cancer 2015, 16, 431–446. [Google Scholar] [CrossRef][Green Version]
- Terpos, E.; Heath, D.J.; Rahemtulla, A.; Zervas, K.; Chantry, A.; Anagnostopoulos, A.; Pouli, A.; Katodritou, E.; Verrou, E.; Vervessou, E.C.; et al. Bortezomib Reduces Serum Dickkopf-1 and Receptor Activator of Nuclear Factor-κB Ligand Concentrations and Normalises Indices of Bone Remodelling in Patients with Relapsed Multiple Myeloma. Br. J. Haematol. 2006, 135, 688–692. [Google Scholar] [CrossRef]
- Terpos, E.; Politou, M.; Szydlo, R.; Nadal, E.; Avery, S.; Olavaria, E.; Kanfer, E.; Goldman, J.M.; Apperley, J.F.; Rahemtulla, A. Autologous Stem Cell Transplantation Normalizes Abnormal Bone Remodeling and SRANKL/Osteoprotegerin Ratio in Patients with Multiple Myeloma. Leukemia 2004, 18, 1420–1426. [Google Scholar] [CrossRef][Green Version]
- Peters, S.; Clézardin, P.; Márquez-Rodas, I.; Niepel, D.; Gedye, C. The RANK–RANKL Axis: An Opportunity for Drug Repurposing in Cancer? Clin. Transl. Oncol. 2019, 21, 977–991. [Google Scholar] [CrossRef]
- Ciscar, M.; Trinidad, E.M.; Perez-Chacon, G.; Alsaleem, M.; Jimenez, M.; Jimenez-Santos, M.J.; Perez-Montoyo, H.; Sanz-Moreno, A.; Vethencourt, A.; Toss, M.; et al. RANK Is a Poor Prognosis Marker and a Therapeutic Target in ER -negative Postmenopausal Breast Cancer. EMBO Mol. Med. 2023, 15, e16715. [Google Scholar] [CrossRef]
- Ferreira, A.; Alho, I.; Vendrell, I.; Melo, M.; Brás, R.; Costa, A.L.; Sousa, A.R.; Mansinho, A.; Abreu, C.; Pulido, C.; et al. The Prognostic Role of RANK SNP Rs34945627 in Breast Cancer Patients with Bone Metastases. Oncotarget 2016, 7, 41380–41389. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Leon-Oliva, D.; Barrena-Blázquez, S.; Jiménez-Álvarez, L.; Fraile-Martinez, O.; García-Montero, C.; López-González, L.; Torres-Carranza, D.; García-Puente, L.M.; Carranza, S.T.; Álvarez-Mon, M.Á.; et al. The RANK–RANKL–OPG System: A Multifaceted Regulator of Homeostasis, Immunity, and Cancer. Medicina 2023, 59, 1752. https://doi.org/10.3390/medicina59101752
De Leon-Oliva D, Barrena-Blázquez S, Jiménez-Álvarez L, Fraile-Martinez O, García-Montero C, López-González L, Torres-Carranza D, García-Puente LM, Carranza ST, Álvarez-Mon MÁ, et al. The RANK–RANKL–OPG System: A Multifaceted Regulator of Homeostasis, Immunity, and Cancer. Medicina. 2023; 59(10):1752. https://doi.org/10.3390/medicina59101752
Chicago/Turabian StyleDe Leon-Oliva, Diego, Silvestra Barrena-Blázquez, Laura Jiménez-Álvarez, Oscar Fraile-Martinez, Cielo García-Montero, Laura López-González, Diego Torres-Carranza, Luis M. García-Puente, Sara T. Carranza, Miguel Ángel Álvarez-Mon, and et al. 2023. "The RANK–RANKL–OPG System: A Multifaceted Regulator of Homeostasis, Immunity, and Cancer" Medicina 59, no. 10: 1752. https://doi.org/10.3390/medicina59101752
APA StyleDe Leon-Oliva, D., Barrena-Blázquez, S., Jiménez-Álvarez, L., Fraile-Martinez, O., García-Montero, C., López-González, L., Torres-Carranza, D., García-Puente, L. M., Carranza, S. T., Álvarez-Mon, M. Á., Álvarez-Mon, M., Diaz, R., & Ortega, M. A. (2023). The RANK–RANKL–OPG System: A Multifaceted Regulator of Homeostasis, Immunity, and Cancer. Medicina, 59(10), 1752. https://doi.org/10.3390/medicina59101752