
Influence of Maternal Diet and Intergenerational Change in Diet Type on Ovarian and Adipose Tissue Morphology in Female Rat Offspring

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
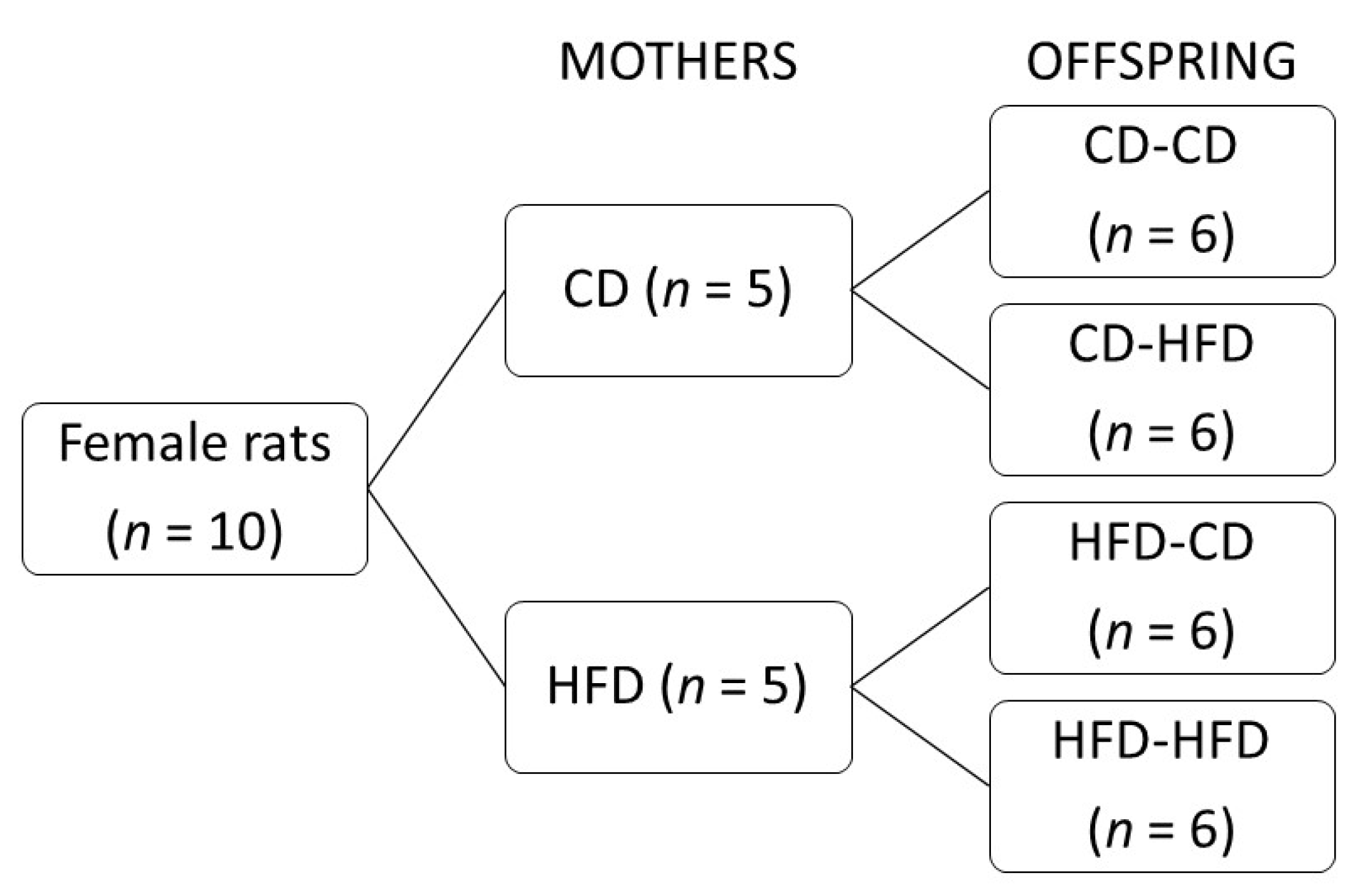
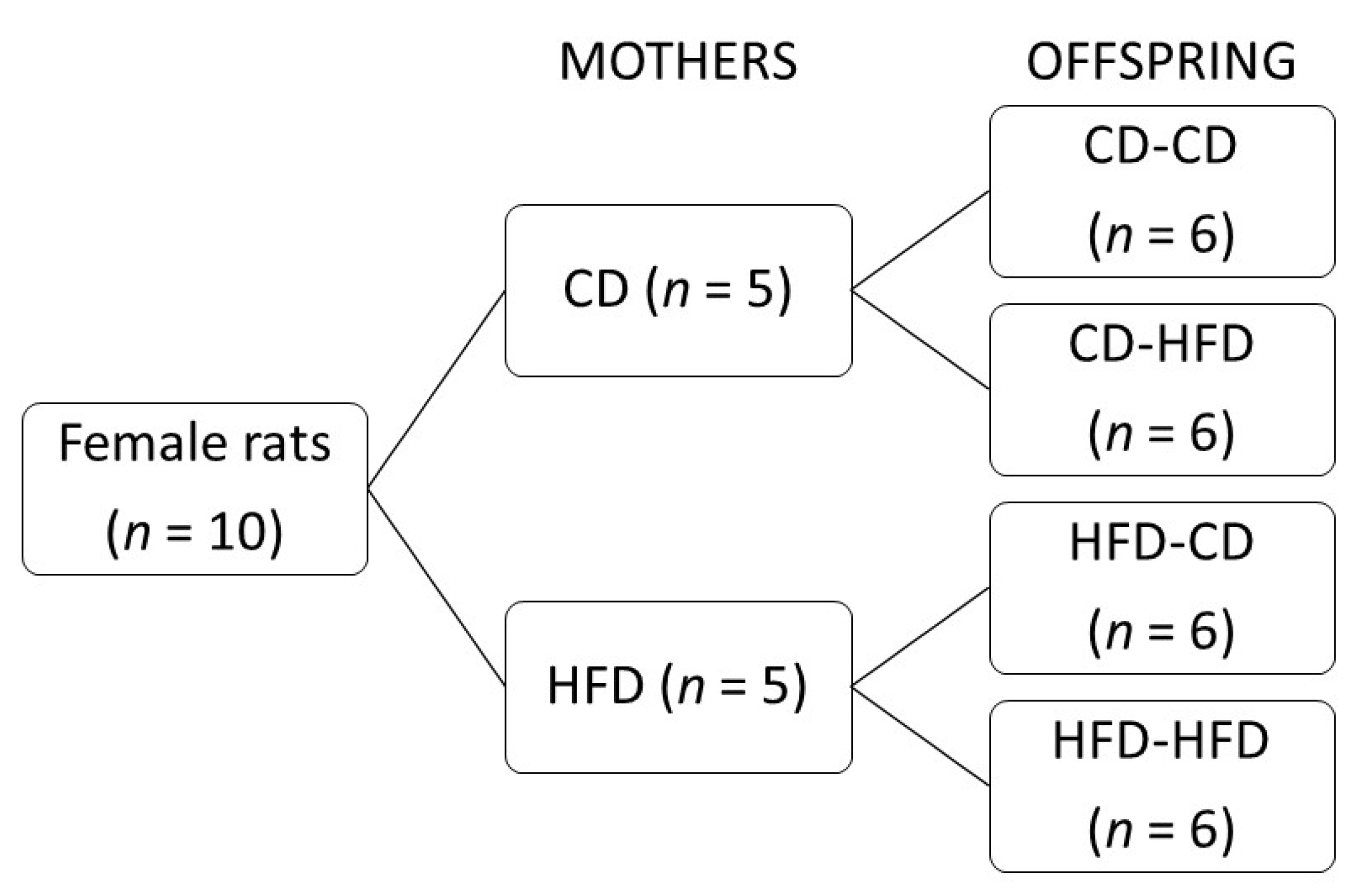
2.1. Study Design
2.2. Tissue Preparation and Histological Examination
2.3. Statistical Analysis
3. Results
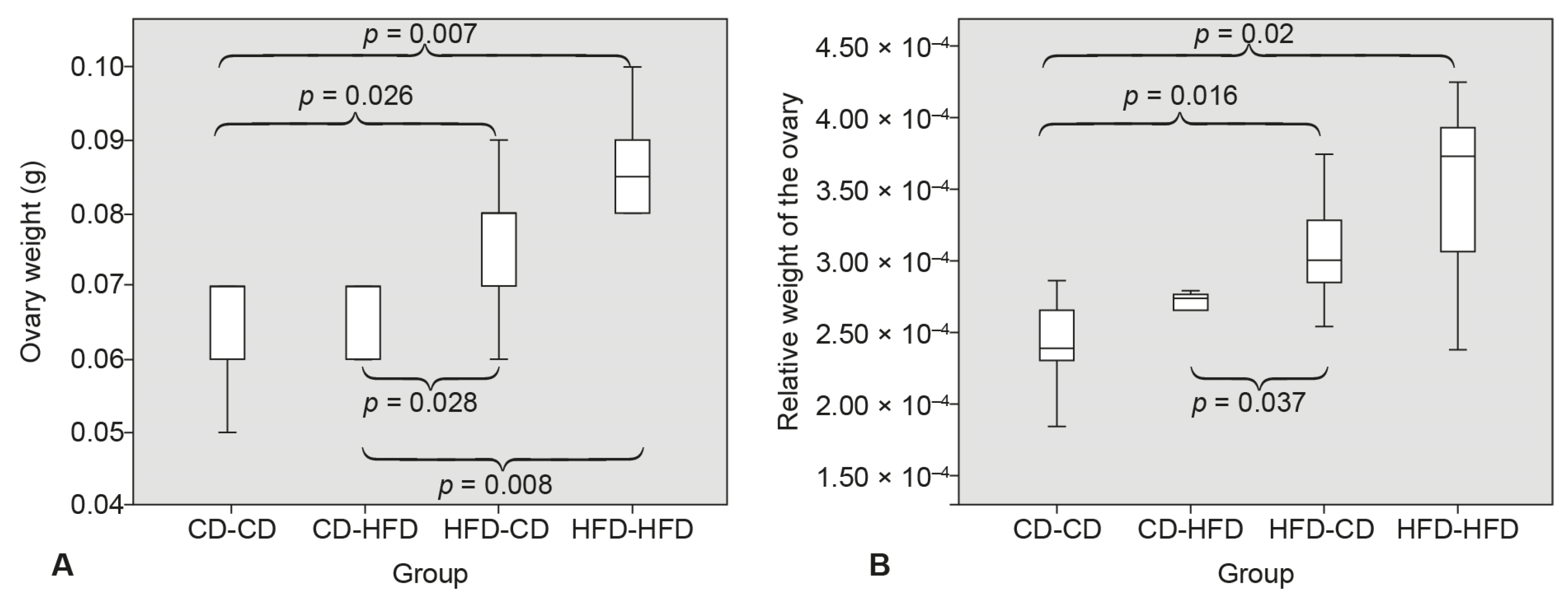
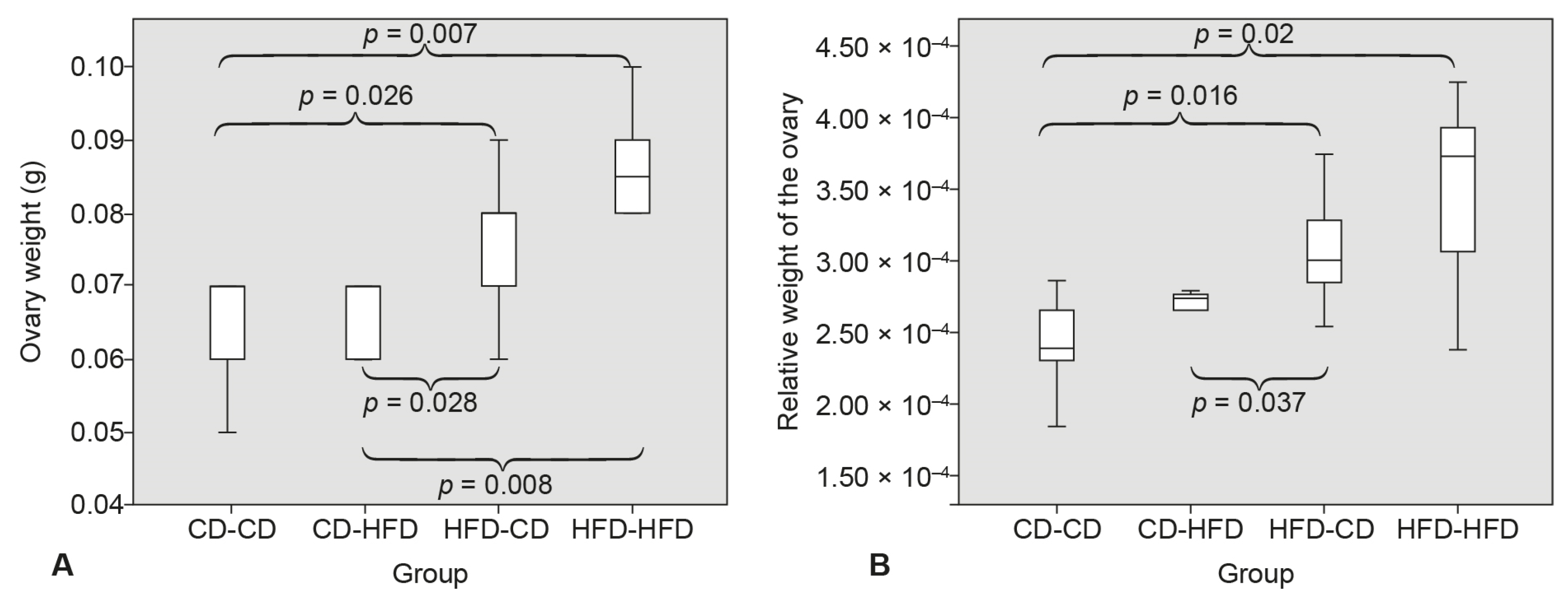
3.1. Ovary Weight
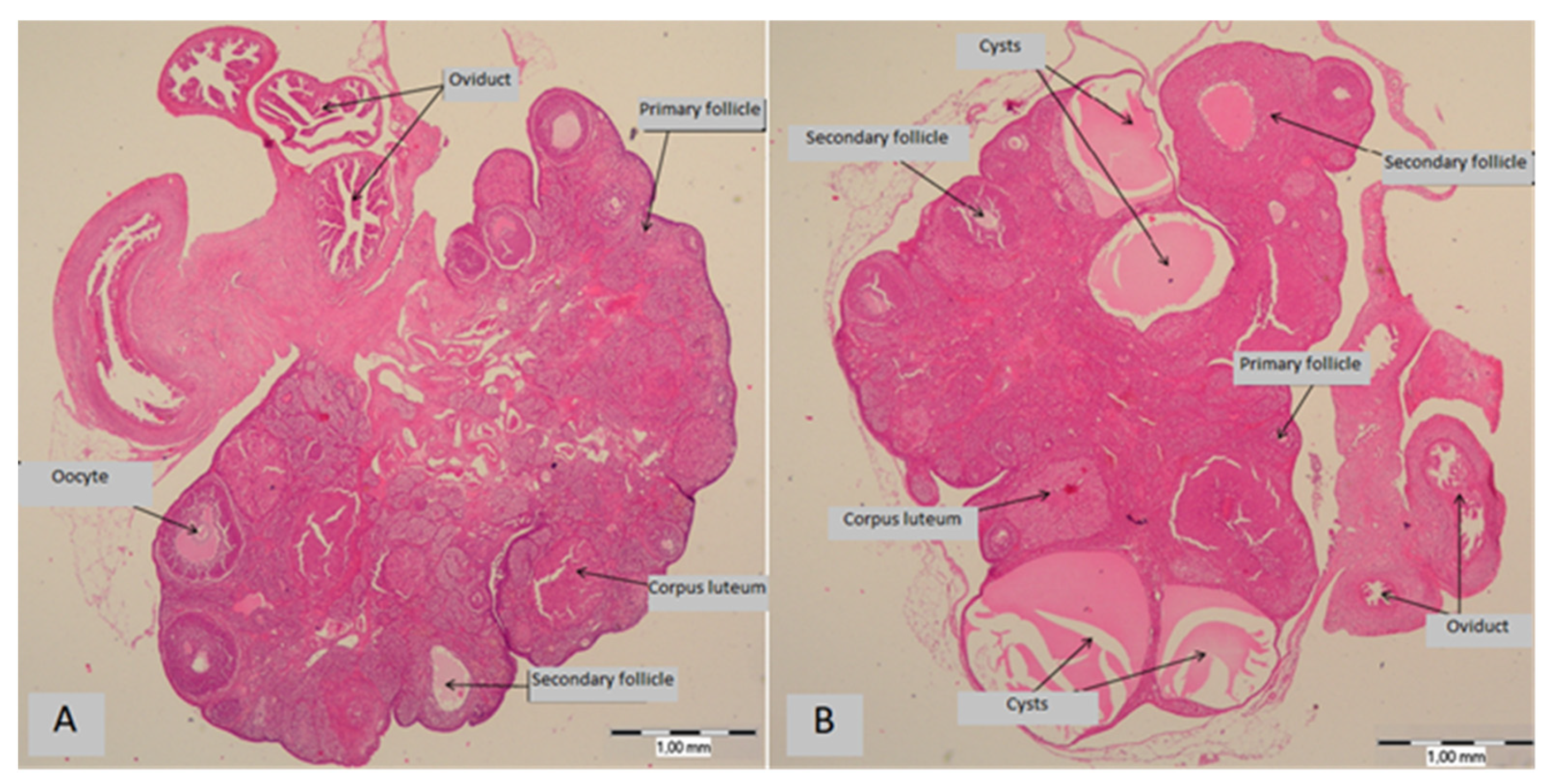
3.2. Ovarian Cysts
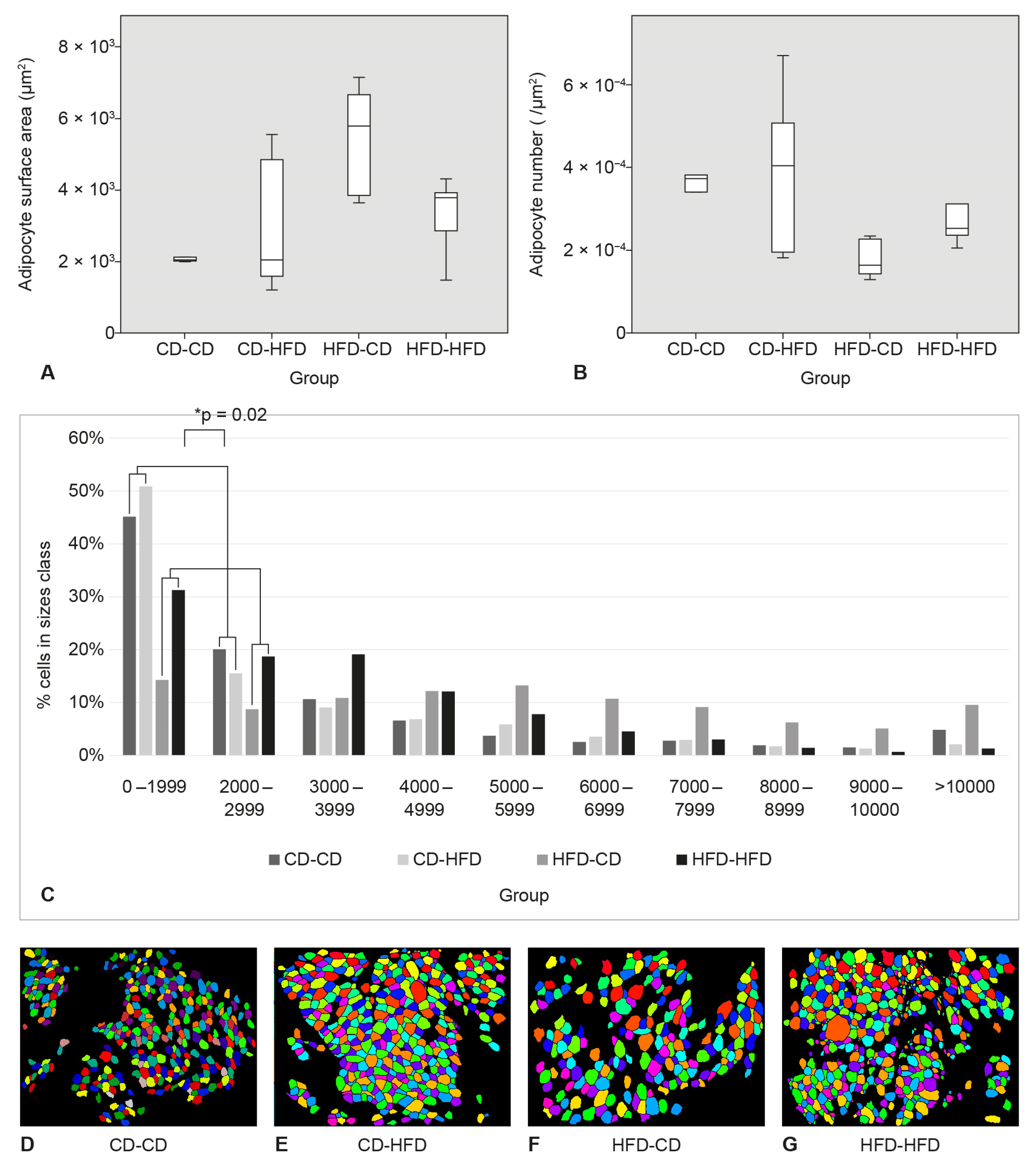
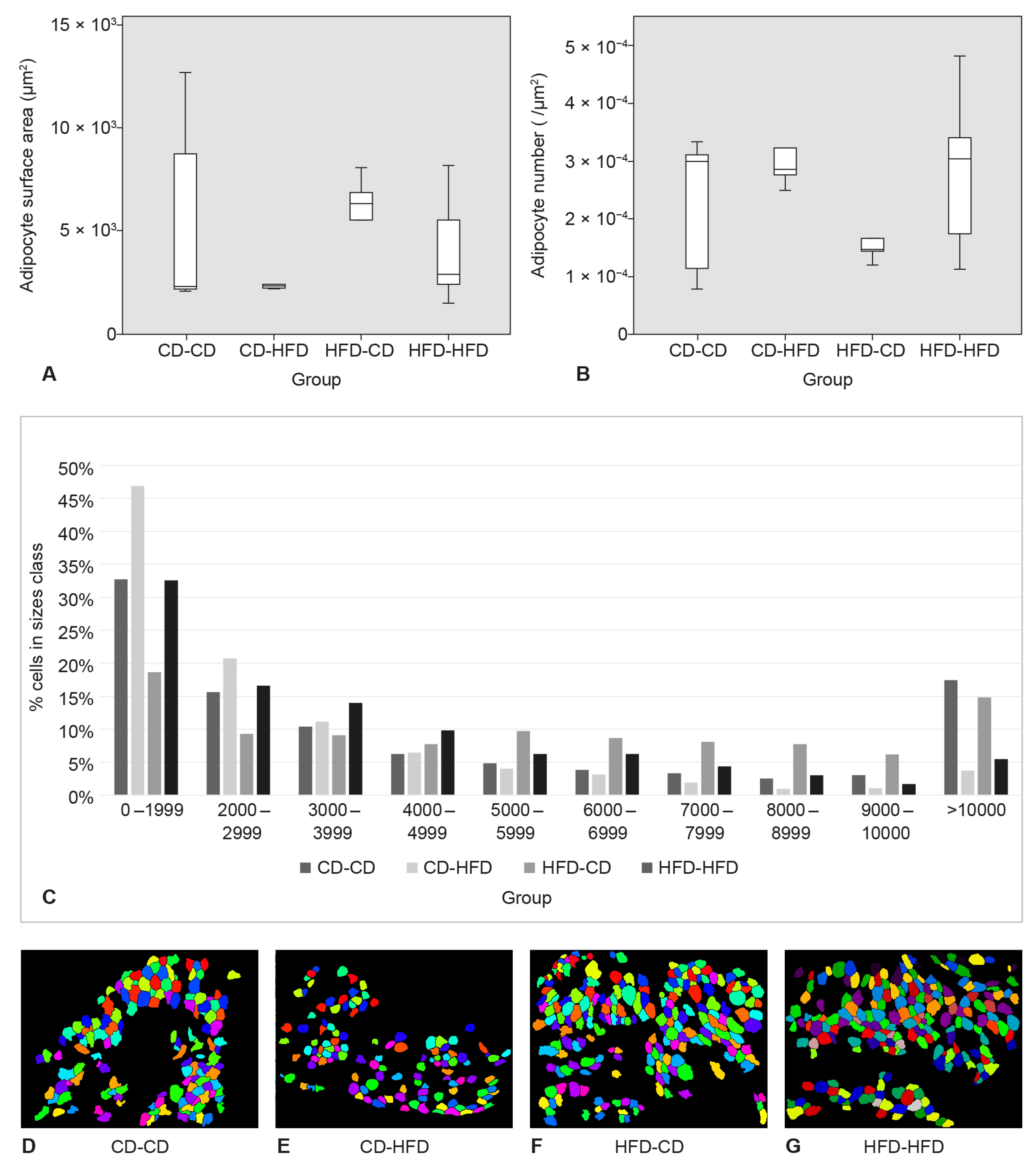
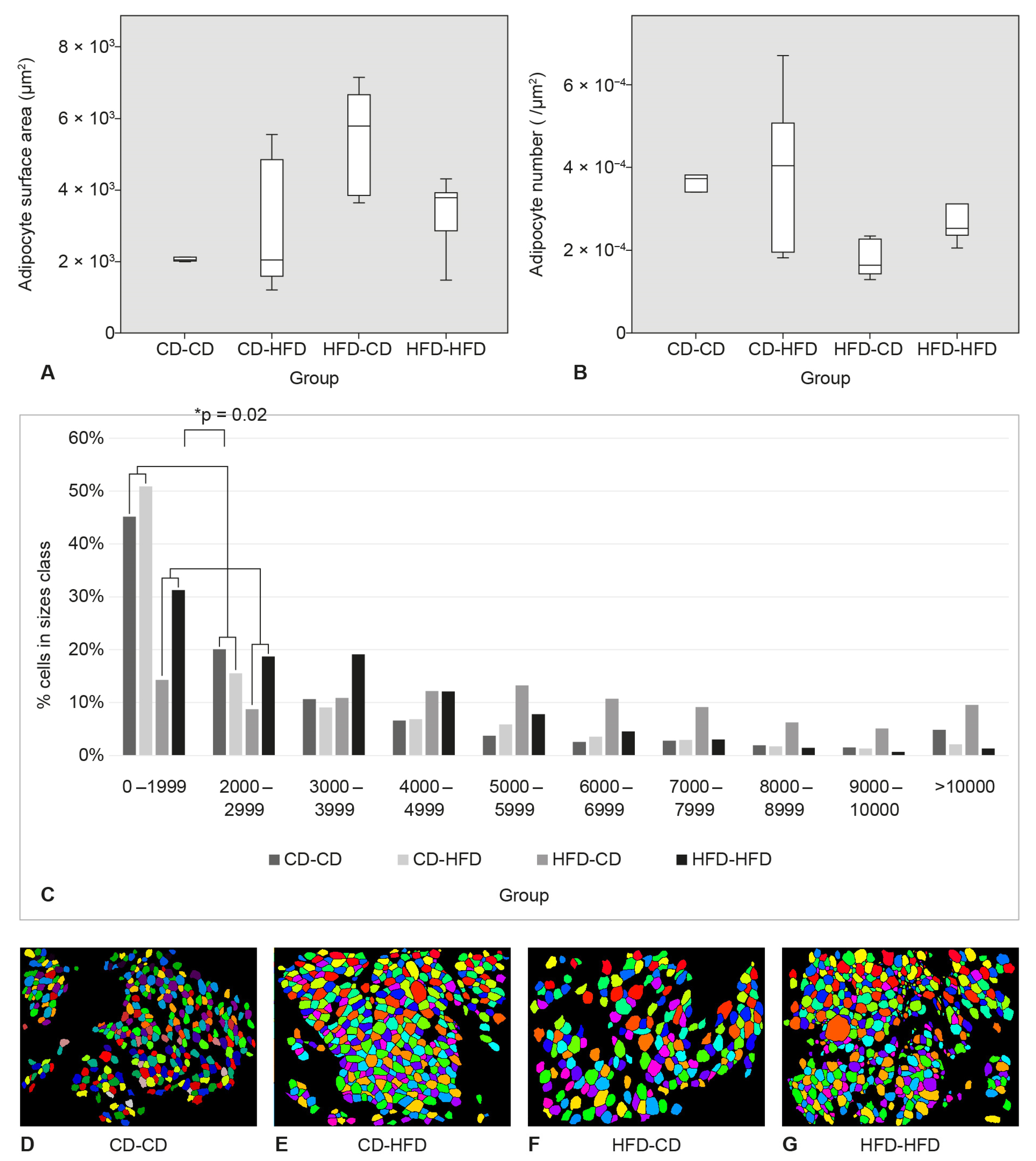
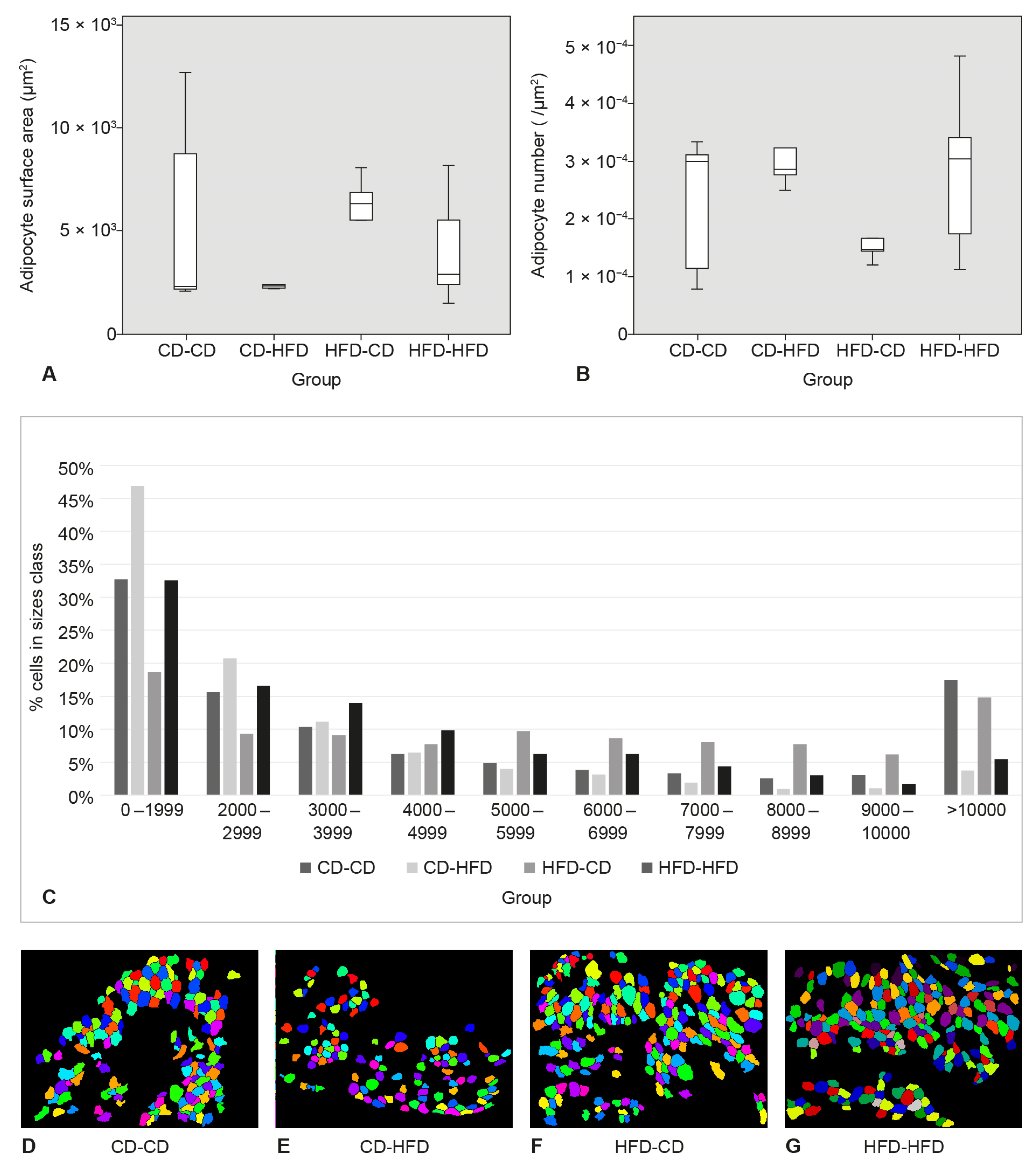
3.3. Surface Area and Number of Adipocytes in SAT
3.4. Area and Number of Adipocytes in Perigonadal Adipose Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Jaacks, L.M.; Vandevijvere, S.; Pan, A.; McGowan, C.J.; Wallace, C.; Imamura, F.; Mozaffarian, D.; Swinburn, B.; Ezzati, M. The obesity transition: Stages of the global epidemic. lancet Diabetes Endocrinol. Lancet Diabetes Endocrinol. 2019, 7, 231–240. [Google Scholar] [CrossRef]
- Vuik, S.; Lerouge, A.; Guillemette, Y.; Feigl, A.; Aldea, A. The Economic Burden of Obesity; OECD: Paris, France, 2019; pp. 74–100. [Google Scholar]
- Cercato, C.; Fonseca, F.A. Cardiovascular risk and obesity. Diabetol. Metab. Syndr. 2019, 11, 74. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Metabolically Healthy Obesity. Endocarp. Rev. 2020, 41, bnaa004. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.A.; Abogresha, N.M.; Hassan, R.; Tamany, D.A.; Lotfy, M. Effect of feeding a high-fat diet independently of caloric intake on reproductive function in diet-induced obese female rats. Arch. Med. Sci. 2016, 12, 906–914. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Park, N.-Y.; Valacchi, G.; Lim, Y. Calorie restriction with a high-fat diet effectively attenuated inflammatory response and oxidative stress-related markers in obese tissues of the high diet fed rats. Mediat. Inflamm. 2012, 2012, 984643. [Google Scholar] [CrossRef] [Green Version]
- de Almeida-Souza, C.B.; Antunes, M.M.; Carbonera, F.; Godoy, G.; da Silva, M.A.R.C.P.; Masi, L.N.; Visentainer, J.V.; Curi, R.; Bazotte, R.B. A High-Fat Diet Induces Lower Systemic Inflammation than a High-Carbohydrate Diet in Mice. Metab. Syndr. Relat. Disord. 2021, 19, 296–304. [Google Scholar] [CrossRef] [PubMed]
- Licholai, J.A.; Nguyen, K.P.; Fobbs, W.C.; Schuster, C.J.; Ali, M.A.; Kravitz, A.V. Why Do Mice Overeat High-Fat Diets? How High-Fat Diet Alters the Regulation of Daily Caloric Intake in Mice. Obesity 2018, 26, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Garman, T.S.; Setlow, B.; Orsini, C.A. Effects of a high-fat diet on impulsive choice in rats. Physiol. Behav. 2021, 229, 113260. [Google Scholar] [CrossRef]
- Vangoitsenhoven, R.; van der Ende, M.; Corbeels, K.; Monteiro Carvalho Mori Cunha, J.P.; Lannoo, M.; Bedossa, P.; van der Merwe, S.; Mertens, A.; Gesquiere, I.; Meulemans, A. At similar weight loss, dietary composition determines the degree of glycemic improvement in diet-induced obese C57BL/6 mice. PLoS ONE 2018, 13, e0200779. [Google Scholar] [CrossRef]
- Coelho, M.; Oliveira, T.; Fernandes, R. Biochemistry of adipose tissue: An endocrine organ. Arch. Med. Sci. 2013, 9, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zoico, E.; Rubele, S.; De Caro, A.; Nori, N.; Mazzali, G.; Fantin, F.; Rossi, A.; Zamboni, M. Brown and Beige Adipose Tissue and Aging. Front. Endocrinol. 2019, 10, 368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chusyd, D.E.; Wang, D.; Huffman, D.M.; Nagy, T.R. Relationships between Rodent White Adipose Fat Pads and Human White Adipose Fat Depots. Front. Nutr. 2016, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Fuster, J.J.; Ouchi, N.; Gokce, N.; Walsh, K. Obesity-Induced Changes in Adipose Tissue Microenvironment and Their Impact on Cardiovascular Disease. Circ. Res. 2016, 118, 1786–1807. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, J.; Heslehurst, N.; Hall, J.; Schoenaker, D.A.J.M.; Hutchinson, J.; Cade, J.E.; Poston, L.; Barrett, G.; Crozier, S.R.; Barker, M.; et al. Before the beginning: Nutrition and lifestyle in the preconception period and its importance for future health. Lancet 2018, 391, 1830–1841. [Google Scholar] [CrossRef]
- Breton, C.V.; Landon, R.; Kahn, L.G.; Enlow, M.B.; Peterson, A.K.; Bastain, T.; Braun, J.; Comstock, S.S.; Duarte, C.S.; Hipwell, A.; et al. Exploring the evidence for epigenetic regulation of environmental influences on child health across generations. Commun. Biol. 2021, 4, 769. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wang, F.; Shi, Q. The effect of maternal high-fat-diet mediated oxidative stress on ovarian function in mice offspring. Exp. Ther. Med. 2020, 20, 135. [Google Scholar] [CrossRef]
- Zhou, Z.; Lin, Q.; Xu, X.; Illahi, G.S.; Dong, C.; Wu, X. Maternal high-fat diet impairs follicular development of offspring through intraovarian kisspeptin/GPR54 system. Reprod. Biol. Endocrinol. 2019, 17, 13. [Google Scholar] [CrossRef]
- Andreas, E.; Reid, M.; Zhang, W.; Moley, K.H. The effect of maternal high-fat/high-sugar diet on offspring oocytes and early embryo development. Mol. Hum. Reprod. 2019, 25, 717–728. [Google Scholar] [CrossRef]
- Yildiz, B.O.; Azziz, R.; Androgen Excess and PCOS Society. Ovarian and adipose tissue dysfunction in polycystic ovary syndrome: Report of the 4th special scientific meeting of the Androgen Excess and PCOS Society. Fertil. Steril. 2010, 94, 690–693. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Yang, M.; Ericsson, A.C. Function of Macrophages in Disease: Current Understanding on Molecular Mechanisms. Front. Immunol. 2021, 12, 635. [Google Scholar] [CrossRef] [PubMed]
- Lima, P.D.A.; Nivet, A.-L.; Wang, Q.; Chen, Y.-A.; Leader, A.; Cheung, A.; Tzeng, C.H.; Tsang, B.K. Polycystic ovary syndrome: Possible involvement of androgen-induced, chemerin-mediated ovarian recruitment of monocytes/macrophages. Biol. Reprod. 2018, 99, 838–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hohos, N.M.; Skaznik-Wikiel, M.E. High-Fat Diet and Female Fertility. Endocrinology 2017, 158, 2407–2419. [Google Scholar] [CrossRef] [PubMed]
- Marić, A.; Kačarević, Ž.P.; Čekić, N.; Šerić, V.; Radić, R. Effects of between generations changes in nutrition type on vaginal smear and serum lipids in Sprague-Dawley rats. J. Matern. Fetal Neonatal Med. 2016, 29, 1491–1497. [Google Scholar] [CrossRef] [PubMed]
- Manneras, L.; Cajander, S.; Holmäng, A.; Seleskovic, Z.; Lystig, T.; Lönn, M.; Stener-Victorin, E. A New Rat Model Exhibiting Both Ovarian and Metabolic Characteristics of Polycystic Ovary Syndrome. Endocrinology 2007, 148, 3781–3791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley-Evans, S.C. Developmental programming of health and disease. Proc. Nutr. Soc. 2006, 65, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Kalem, Z.; Namlı Kalem, M.; Anadol, E.; Bakırarar, B.; Yılmaz, C.; Elmas, Ç.; Yalçınkaya, P.; Ruso, H.; Gürgan, T. Maternal nutrition and reproductive functions of female and male offspring. Reproduction 2018, 156, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Léveillé, P.; Tarrade, A.; Dupont, C.; Larcher, T.; Dahirel, M.; Poumerol, E.; Cordier, A.-G.; Picone, O.; Mandon-Pepin, B.; Jolivet, G.; et al. Maternal high-fat diet induces follicular atresia but does not affect fertility in adult rabbit offspring. J. Dev. Orig. Health Dis. 2014, 5, 88–97. [Google Scholar] [CrossRef]
- Xu, M.; Che, L.; Yang, Z.; Zhang, P.; Shi, J.; Li, J.; Lin, Y.; Fang, Z.; Che, L.; Feng, B.; et al. Effect of High Fat Dietary Intake during Maternal Gestation on Offspring Ovarian Health in a Pig Model. Nutrients 2016, 8, 498. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; der Hoek, K.H.; Ryan, N.K.; Norman, R.J.; Robker, R.L. Macrophage contributions to ovarian function. Hum. Reprod. Update 2004, 10, 119–133. [Google Scholar] [CrossRef] [Green Version]
- Hirosumi, J.; Tuncman, G.; Chang, L.; Görgün, C.Z.; Uysal, K.T.; Maeda, K.; Karin, M.; Hotamisligi, G.S. A central role for JNK in obesity and insulin resistance. Nature 2002, 420, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Makki, K.; Froguel, P.; Wolowczuk, I. Adipose tissue in obesity-related inflammation and insulin resistance: Cells, cytokines, and chemokines. ISRN Inflamm. 2013, 2013, 139239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Gao, Z.; Yin, J.; He, Q. Hypoxia is a potential risk factor for chronic inflammation and adiponectin reduction in adipose tissue of ob/ob and dietary obese mice. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1118–E1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Z.; Rosen, E.D.; Brun, R.; Hauser, S.; Adelmant, G.; Troy, A.E.; McKeon, C.; Darlington, G.J.; Spiegelman, B.M. Cross-regulation of C/EBP alpha and PPAR gamma controls the transcriptional pathway of adipogenesis and insulin sensitivity. Mol. Cell. 1999, 3, 151–158. [Google Scholar] [CrossRef]
- Koh, Y.J.; Park, B.-H.; Park, J.-H.; Han, J.; Lee, I.-K.; Park, J.W.; Koh, G.Y. Activation of PPAR gamma induces profound multilocularization of adipocytes in adult mouse white adipose tissues. Exp. Mol. Med. 2009, 41, 880–895. [Google Scholar] [CrossRef]
- Rudnicki, M.; Abdifarkosh, G.; Rezvan, O.; Nwadozi, E.; Roudier, E.; Haas, T.L. Female Mice Have Higher Angiogenesis in Perigonadal Adipose Tissue Than Males in Response to High-Fat Diet. Front. Physiol. 2018, 9, 1452. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Percent of Food Component (%) | CD | HFD | |
|---|---|---|---|
| Before 14 weeks of age | After 14 weeks of age | ||
| Complex carbohydrates | 53.7 | 66.5 | 30 |
| Plant proteins | 30.5 | 18.2 | 27.2 |
| Animal proteins | 4.7 | 3.5 | |
| Fodder mix | 3.0 | 7.5 | 9.7 |
| Vitamins and minerals | 4.1 | 3.2 | 3.9 |
| Fats (soybean oil) | 1.4 | 0.4 | 28 |
| Amino acids | 0.1 | 0.1 | 0.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Čekić, N.; Grgić, A.; Kokot, A.; Mujkić, R.; Šnajder Mujkić, D.; Bijelić, N.; Sablić, M. Influence of Maternal Diet and Intergenerational Change in Diet Type on Ovarian and Adipose Tissue Morphology in Female Rat Offspring. Medicina 2022, 58, 854. https://doi.org/10.3390/medicina58070854
Čekić N, Grgić A, Kokot A, Mujkić R, Šnajder Mujkić D, Bijelić N, Sablić M. Influence of Maternal Diet and Intergenerational Change in Diet Type on Ovarian and Adipose Tissue Morphology in Female Rat Offspring. Medicina. 2022; 58(7):854. https://doi.org/10.3390/medicina58070854
Chicago/Turabian StyleČekić, Nenad, Anđela Grgić, Antonio Kokot, Robert Mujkić, Darija Šnajder Mujkić, Nikola Bijelić, and Marko Sablić. 2022. "Influence of Maternal Diet and Intergenerational Change in Diet Type on Ovarian and Adipose Tissue Morphology in Female Rat Offspring" Medicina 58, no. 7: 854. https://doi.org/10.3390/medicina58070854
APA StyleČekić, N., Grgić, A., Kokot, A., Mujkić, R., Šnajder Mujkić, D., Bijelić, N., & Sablić, M. (2022). Influence of Maternal Diet and Intergenerational Change in Diet Type on Ovarian and Adipose Tissue Morphology in Female Rat Offspring. Medicina, 58(7), 854. https://doi.org/10.3390/medicina58070854