Impact of Activation of EGFL7 within Microenvironment of High Grade Ovarian Serous Carcinoma on Infiltration of CD4+ and CD8+ Lymphocytes

 ,
,
Abstract
:1. Background
2. Material and Methods
2.1. Human Samples
2.2. Tissue Samples
2.3. Immunohistochemistry
2.4. IHC Evaluation of ICAM-1
2.5. Evaluation and Classification of CD4+ and CD8+ Cells
2.6. qPCR in Samples Obtained with Microdissection
2.7. Statistical Analysis
3. Results
3.1. Patient Population
3.2. EGFL7 Expression at mRNA Level
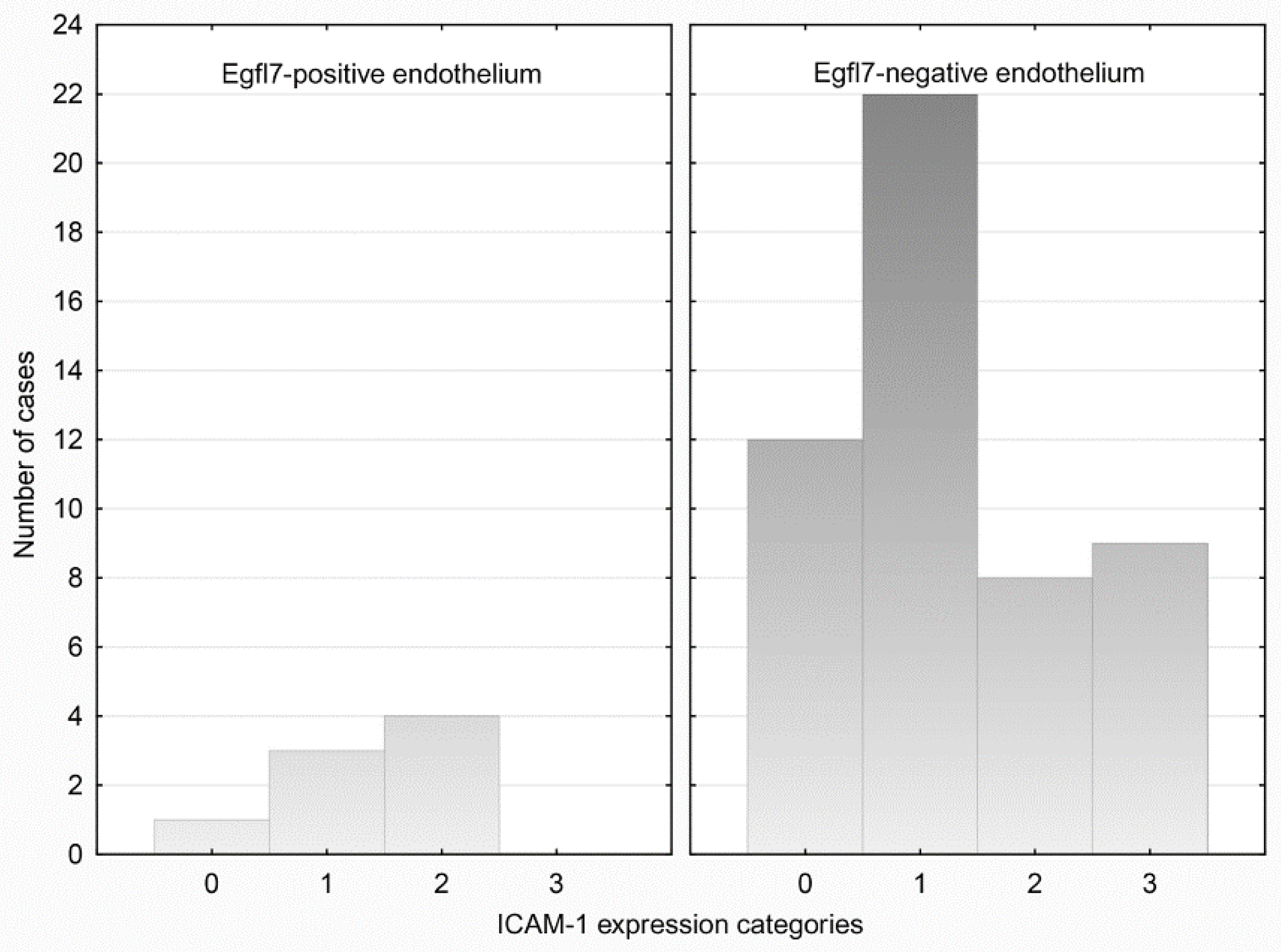
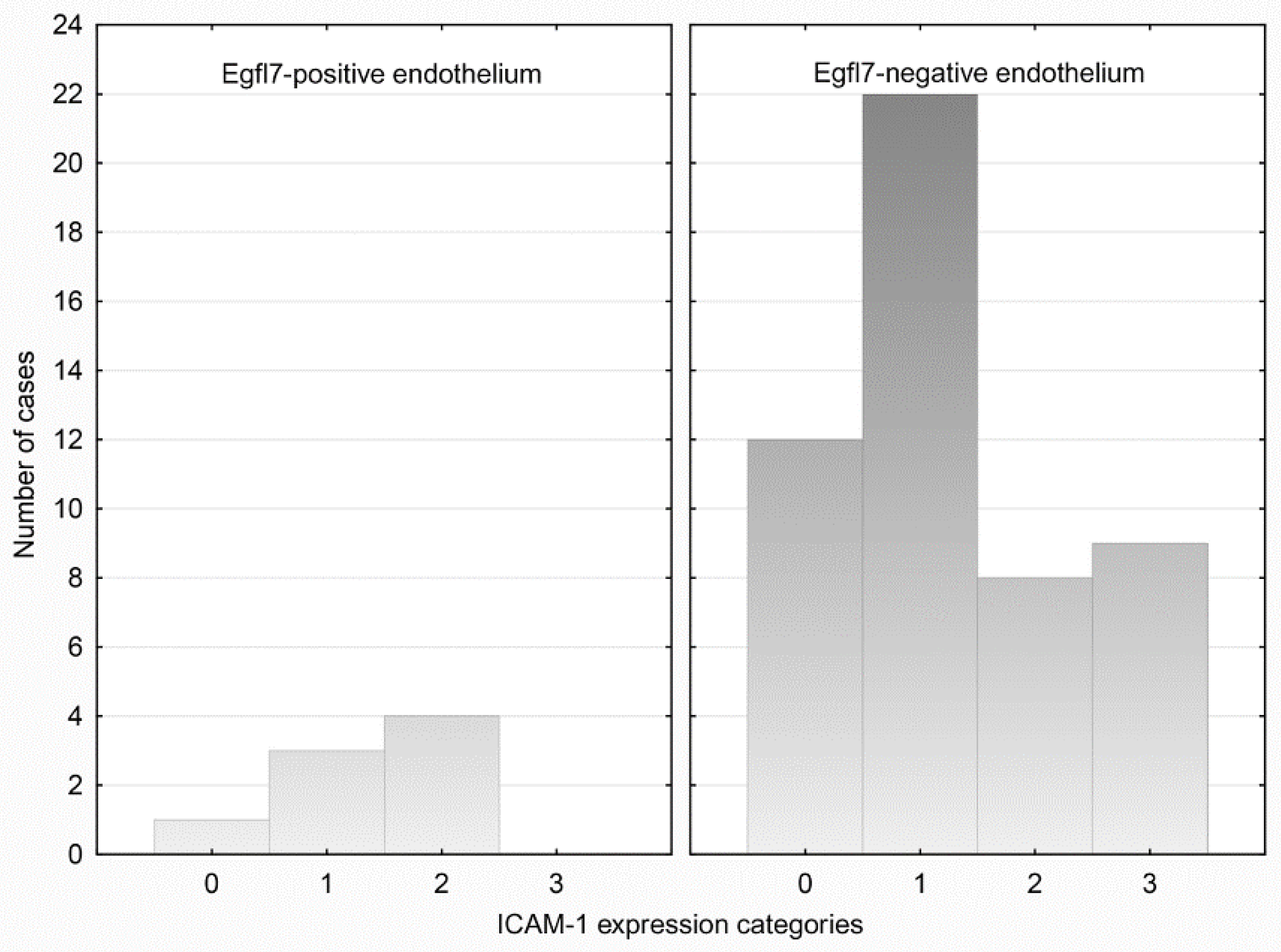
3.3. ICAM-1 Expression
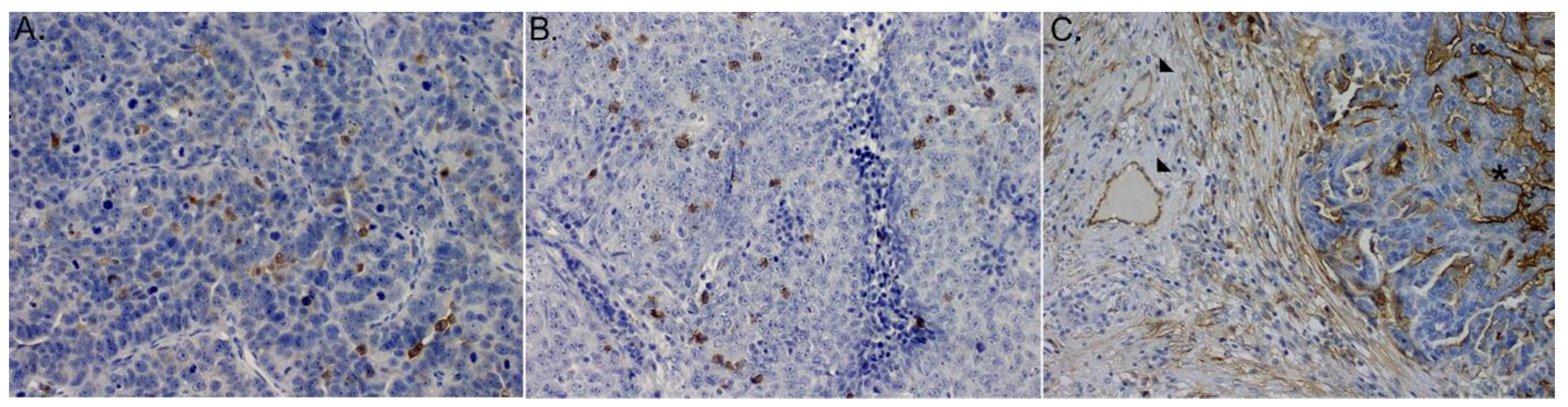
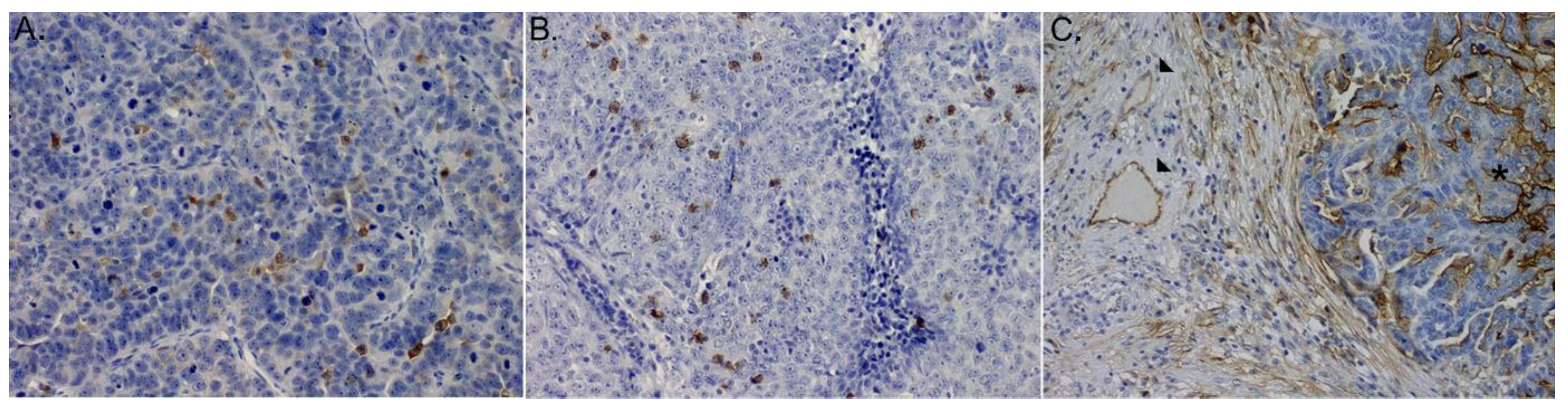
3.4. Immunohistochemistry for CD4+ and CD8+Cells
3.5. Association of mRNA Egfl7 Status with (IE)CD4+ and (IE)CD8+ Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Egfl7 | Epidermal Growth Factor like domain 7 |
| OvCa | Ovarian cancer |
| ICAM-1 | Intercellular adhesion molecule-1 |
| VCAM-1 | Vascular cell adhesion molecule-1 |
| LCM | Laser Capture Microdissection |
| HGSOC | High Grade Ovarian Serous Carcinoma |
| PDS | Primary debulking surgery |
| FIGO | The International Federation of Gynecology and Obstetrics |
| FFPE | Formalin-fixed paraffin-embedded |
| IHC | Immunohistochemical |
| DFS | Disease free survival |
| PFS | Progression free survival |
| OS | Overall survival |
| HPF | High-power field |
| IE | Intraepithelial |
References
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yigit, R.; Massuger, L.F.; Figdor, C.; Torensma, R. Ovarian cancer creates a suppressive microenvironment to escape immune elimination. Gynecol. Oncol. 2010, 117, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Ogino, S.; Galon, J.; Fuchs, C.S.; Dranoff, G. Cancer immunology—Analysis of host and tumor factors for personalized medicine. Nat. Rev. Clin. Oncol. 2011, 8, 711–719. [Google Scholar] [CrossRef] [PubMed]
- Quesnel, B. Tumor dormancy and immunoescape. APMIS 2008, 116, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Muller, W.A. Mechanisms of Transendothelial Migration of Leukocytes. Circ. Res. 2009, 105, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.Z.; Klitzman, B.; Dodge, R.; Dewhirst, M.W. Diminished leukocyte-endothelium interaction in tumor microvessels. Cancer Res. 1992, 52, 4265–4268. [Google Scholar]
- Castermans, K.; Griffioen, A. Tumor blood vessels, a difficult hurdle for infiltrating leukocytes. Biochim. Biophys. Acta 2007, 1776, 160–174. [Google Scholar] [CrossRef]
- Liu, Z.; Guo, B.; Lopez, R.D. Expression of intercellular adhesion molecule (ICAM)-1 or ICAM-2 is critical in determining sensitivity of pancreatic cancer cells to cytolysis by human γδ-T cells: Implications in the design of γδ-T-cell-based immunotherapies for pancreatic cancer. J. Gastroenterol. Hepatol. 2009, 24, 900–911. [Google Scholar] [CrossRef]
- Piali, L.; Fichtel, A.; Terpe, H.J.; Imhof, B.A.; Gisler, R.H. Endothelial vascular cell adhesion molecule 1 expression is suppressed by melanoma and carcinoma. J. Exp. Med. 1995, 181, 811–816. [Google Scholar] [CrossRef] [Green Version]
- Kuzu, I.; Bicknell, R.; Fletcher, C.D.; Gatter, K.C. Expression of adhesion molecules on the endothelium of normal tissue vessels and vascular tumors. Lab. Investig. 1993, 69, 322–328. [Google Scholar]
- Griffioen, A.W.; Damen, C.A.; Martinotti, S.; Blijham, G.H.; Groenewegen, G. Endothelial intercellular adhesion molecule-1 expression is suppressed in human malignancies: The role of angiogenic factors. Cancer Res. 1996, 56, 1111–1117. [Google Scholar] [PubMed]
- Dirkx, A.E.; Oude Egbrink, M.G.; Kuijpers, M.J.; van der Niet, S.T.; Heijnen, V.V.; Bouma-ter Steege, J.C.; Wagstaff, J.; Griffioen, A.W. Tumor angiogenesis modulates leukocyte-vessel wall interactions in vivo by reducing endothelial adhesion molecule expression. Cancer Res. 2003, 63, 2322–2329. [Google Scholar] [PubMed]
- Parker, L.H.; Schmidt, M.; Jin, S.-W.; Gray, A.M.; Beis, D.; Pham, T.; Frantz, G.; Palmieri, S.; Hillan, K.; Stainier, D.; et al. The endothelial-cell-derived secreted factor Egfl7 regulates vascular tube formation. Nature 2004, 428, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Soncin, F.; Mattot, V.; Lionneton, F.; Spruyt, N.; Lepretre, F.; Bégué, A.; Stéhelin, D. VE-statin, an endothelial repressor of smooth muscle cell migration. EMBO J. 2003, 22, 5700–5711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lelièvre, E.; Hinek, A.; Lupu, F.; Buquet, C.; Soncin, F.; Mattot, V. VE-statin/egfl7 regulates vascular elastogenesis by interacting with lysyl oxidases. EMBO J. 2008, 27, 1658–1670. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Yang, L.-Y.; Wu, F.; Tao, Y.-M.; Liu, L.-S.; Zhang, J.-F.; He, Y.-N.; Tang, L.-L.; Chen, G.-D.; Guo, L. The Expression of Egfl7 in Human Normal Tissues and Epithelial Tumors. Int. J. Biol. Mark. 2013, 28, 71–83. [Google Scholar] [CrossRef]
- Delfortrie, S.; Pinte, S.; Mattot, V.; Samson, C.; Villain, G.; Caetano, B.; Lauridant-Philippin, G.; Baranzelli, M.-C.; Bonneterre, J.; Trottein, F.; et al. Egfl7 Promotes Tumor Escape from Immunity by Repressing Endothelial Cell Activation. Cancer Res. 2011, 71, 7176–7186. [Google Scholar] [CrossRef] [Green Version]
- Pannier, D.; Philippin-Lauridant, G.; Baranzelli, M.-C.; Bertin, D.; Bogart, E.; Delprat, V.; Villain, G.; Mattot, V.; Bonneterre, J.; Soncin, F. High expression levels of egfl7 correlate with low endothelial cell activation in peritumoral vessels of human breast cancer. Oncol. Lett. 2016, 12, 1422–1428. [Google Scholar] [CrossRef] [Green Version]
- Espina, V.; Wulfkuhle, J.D.; Calvert, V.S.; VanMeter, A.; Zhou, W.; Coukos, G.; Geho, D.H.; Petricoin, E.F.; Liotta, L.A. Laser-capture microdissection. Nat. Protoc. 2006, 1, 586–603. [Google Scholar] [CrossRef]
- Prat, J.; FIGO Committee on Gynecologic Oncology. Staging classification for cancer of the ovary, fallopian tube, and peritoneum. Int. J. Gynecol. Obstet. 2014, 124, 1–5. [Google Scholar] [CrossRef]
- Lipponen, P.K.; Colon, Y. Simple quantitation of immunohistochemical staining positive in microscopy for histopathology rountine. Acta Stereol. 1992, 11, 125–132. [Google Scholar]
- Oh, J.; Park, S.H.; Lee, T.S.; Oh, H.K.; Choi, J.-H.; Choi, Y.S. High expression of epidermal growth factor-like domain 7 is correlated with poor differentiation and poor prognosis in patients with epithelial ovarian cancer. J. Gynecol. Oncol. 2014, 25, 334–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.-H.; Li, X.-J.; Zhou, Y.-Z.; Luo, Y.; Li, C.; Yuan, X.-R. Expression and clinical significance of EGFL7 in malignant glioma. J. Cancer Res. Clin. Oncol. 2010, 136, 1737–1743. [Google Scholar] [CrossRef] [PubMed]
- Díaz, R.; Silva, J.; García, J.M.; Lorenzo, Y.; García, V.; Peña, C.; Rodríguez, R.; Muñoz, C.; García, F.; Bonilla, F.; et al. Deregulated expression of miR-106a predicts survival in human colon cancer patients. Genes, Chromosom. Cancer 2008, 47, 794–802. [Google Scholar] [CrossRef]
- Wu, F.; Yang, L.-Y.; Li, Y.-F.; Ou, D.-P.; Chen, D.-P.; Fan, C. Novel role for epidermal growth factor-like domain 7 in metastasis of human hepatocellular carcinoma. Hepatology 2009, 50, 1839–1850. [Google Scholar] [CrossRef]
- Philippin-Lauridant, G.; Baranzelli, M.-C.; Samson, C.; Fournier, C.; Pinte, S.; Mattot, V.; Bonneterre, J.; Soncin, F. Expression of Egfl7 correlates with low-grade invasive lesions in human breast cancer. Int. J. Oncol. 2013, 42, 1367–1375. [Google Scholar] [CrossRef] [Green Version]
- Santin, A.D.; Rose, G.S.; Hiserodt, J.C.; Fruehauf, J.; Eck, L.M.; Garcia, R.I.; Schranz, V.; Disaia, P.J.; Pecorelli, S.; Granger, G.A. Effects of cytokines combined with high-dose gamma irradiation on the expression of major histocompatibility complex molecules and intercellular adhesion molecule-1 in human ovarian cancers. Int. J. Cancer 1996, 65, 688–694. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Shun, C.-T.; Wu, M.-S.; Chen, C.-C. A Novel Anticancer Effect of Thalidomide: Inhibition of Intercellular Adhesion Molecule-1–Mediated Cell Invasion and Metastasis through Suppression of Nuclear Factor-κB. Clin. Cancer Res. 2006, 12, 7165–7173. [Google Scholar] [CrossRef] [Green Version]
- Roland, C.L.; Harken, A.H.; Sarr, M.G.; Barnett, C.C. ICAM-1 expression determines malignant potential of cancer. Surgery 2007, 141, 705–707. [Google Scholar] [CrossRef]
- Brooks, K.J.; Coleman, E.J.; Vitetta, E.S. The antitumor activity of an anti-CD54 antibody in SCID mice xenografted with human breast, prostate, non-small cell lung, and pancreatic tumor cell lines. Int. J. Cancer 2008, 123, 2438–2445. [Google Scholar] [CrossRef]
- Miele, M.E.; Bennett, C.; Miller, B.E.; Welch, D.R. Enhanced Metastatic Ability of TNF-α-Treated Malignant Melanoma Cells Is Reduced by Intercellular Adhesion Molecule-1 (ICAM-1, CD54) Antisense Oligonucleotides. Exp. Cell Res. 1994, 214, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Coleman, E.J.; Pop, L.M.; Brooks, K.J.; Vitetta, E.S.; Niederkorn, J.Y. Effect of an anti-CD54 (ICAM-1) monoclonal antibody (UV3) on the growth of human uveal melanoma cells transplanted heterotopically and orthotopically in SCID mice. Int. J. Cancer 2006, 118, 932–941. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Antibody | Nr. Cat | Dilution | Expression | |
|---|---|---|---|---|
| EGFL7 | Santa Cruz | Santa Cruz sc-101349 | 1:200 | Nuclear-cytoplasmic |
| ICAM | Abcam | [MEM-111] (ab2213) | 1:400 | Membranous |
| CD4 | Millipore | IHCR2110-6 | RTU | cytoplasmic |
| CD8 | Millipore | IHCR2114-6 | RTU | cytoplasmic |
| Feature | Value |
|---|---|
| Age (years) | median 59 (range 39–86) |
| Preoperative CA125 (U/mL) | median 849 (range 6.8–25,000) |
| Ascites Yes/No (number) | 38/21 |
| Preoperative peripheral blood neutrocytes (G/L) | median 6.15 (range 3.27–10.99) |
| Preoperative peripheral blood lymphocytes (G/L) | median 1.5 (range 0.43–3.02) |
| FIGO I (number/percentage) | 8/13.6% |
| FIGO II (number/percentage) | 1/1.7% |
| FIGO III (number/percentage) | 48/81.4% |
| FIGO IV (number/percentage) | 2/3.3% |
| Residual disease: 0 cm (number/percentage) | 8/13.6% |
| Residual disease: <1 cm (number/percentage) | 22/37.3% |
| Residual disease: >1 cm (number/percentage) | 29/49.1% |
| OS (months). | median 26.99 (range 1.07–78.07) |
| Feature | Cancer Egfl7(+) | Cancer Egfl7(−) | p | Cancer ICAM-1(+) | Cancer ICAM-1(−) | p |
|---|---|---|---|---|---|---|
| Age (years) median (range) | 60 (40–84) | 58.5 (39–86) | 0.626 | 58 (39–84) | 62 (43–86) | 0.309 |
| Preoperative CA125 (U/mL) median (range) | 660 (6.8–3000) | 1000.8 (19.56–25,000) | 0.204 | 837 (6.8–25,000) | 1304 (174–8910) | 0.337 |
| Ascites Y/N number | 6/3 | 32/18 | 1 | 32/15 | 6/6 | 0.315 |
| Preoperative neutrocytes (G/L) median (range) | 7.17 (5.38–8.19) | 6.02 (3.27–10.99) | 0.446 | 6.15 (3.27–10.99) | 5.64 (4.99–8.55) | 0.929 |
| Preoperative lymphocytes (G/L) median (range) | 1.465 (1.28–2.99) | 1.5 (0.43–3.02) | 0.732 | 1.49 (0.43–3.02) | 1.8 (1.12–2.55) | 0.244 |
| FIGO I | 1 | 7 | 0.882 | 6 | 2 | 0.830 |
| FIGO II | 0 | 1 | 1 | 0 | ||
| FIGO III | 8 | 40 | 38 | 10 | ||
| FIGO IV | 0 | 2 | 2 | 0 | ||
| Residual disease: 0 cm | 1 | 7 | 0.962 | 7 | 1 | 0.311 |
| Residual disease: <1 cm | 4 | 18 | 19 | 3 | ||
| Residual disease: >1 cm | 4 | 25 | 21 | 8 | ||
| Platin sensitive | 7 | 27 | 0.081 | 16 | 2 | 0.462 |
| Platin refractor/resistance | 0 | 18 | 26 | 8 |
| Feature | Endothelium Egfl7 (+) | Endothelium Egfl7 (−) | p | Endothelium ICAM-1 (+) | Endothelium ICAM-1 (−) | p |
|---|---|---|---|---|---|---|
| Age (years) median (range) | 59 (40–80) | 59 (39–68) | 0.625 | 59.5 (39–86) | 57 (43–80) | 0.752 |
| Preoperative CA125 (U/mL) median (range) | 1026 (19.56–3012) | 837 (6.8–25,000) | 0.795 | 900 (6.8–25,000) | 570 (88–9935) | 0.605 |
| Ascites Y/N number | 3/5 | 36/16 | 0.119 | 29/17 | 9/4 | 0.754 |
| Preoperative neutrocytes (G/l) median (range) | 7.665 (3.69–8.51) | 6.02 (3.27–10.99) | 0.502 | 5.925 (3.27–10.99) | 6.15 (5.3–8.89) | 0.221 |
| Preoperative lymphocytes (G/l) median (range) | 1.63 (1.18–2.11) | 1.43 (0.43–3.02) | 0.394 | 1.5 (0.43–3.02) | 1.4 (0.64–2.27) | 0.991 |
| FIGO I | 1 | 7 | 0.914 | 5 | 3 | 0.456 |
| FIGO II | 0 | 1 | 1 | 0 | ||
| FIGO III | 7 | 41 | 39 | 9 | ||
| FIGO IV | 0 | 2 | 1 | 1 | ||
| Residual disease: 0 cm | 1 | 2 | 0.705 | 6 | 2 | 0.659 |
| Residual disease: <1 cm | 2 | 20 | 19 | 3 | ||
| Residual disease: >1 cm | 5 | 24 | 21 | 8 | ||
| Platin sensitive | 6 | 28 | 0.081 | 15 | 3 | 1.000 |
| Platin refractor/resistance | 0 | 18 | 27 | 7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sznurkowski, J.J.; Żawrocki, A.; Krawczyńska, N.; Bieńkowski, M.; Wasąg, B.; Biernat, W. Impact of Activation of EGFL7 within Microenvironment of High Grade Ovarian Serous Carcinoma on Infiltration of CD4+ and CD8+ Lymphocytes. Medicina 2022, 58, 588. https://doi.org/10.3390/medicina58050588
Sznurkowski JJ, Żawrocki A, Krawczyńska N, Bieńkowski M, Wasąg B, Biernat W. Impact of Activation of EGFL7 within Microenvironment of High Grade Ovarian Serous Carcinoma on Infiltration of CD4+ and CD8+ Lymphocytes. Medicina. 2022; 58(5):588. https://doi.org/10.3390/medicina58050588
Chicago/Turabian StyleSznurkowski, Jacek J., Anton Żawrocki, Natalia Krawczyńska, Michał Bieńkowski, Bartosz Wasąg, and Wojciech Biernat. 2022. "Impact of Activation of EGFL7 within Microenvironment of High Grade Ovarian Serous Carcinoma on Infiltration of CD4+ and CD8+ Lymphocytes" Medicina 58, no. 5: 588. https://doi.org/10.3390/medicina58050588
APA StyleSznurkowski, J. J., Żawrocki, A., Krawczyńska, N., Bieńkowski, M., Wasąg, B., & Biernat, W. (2022). Impact of Activation of EGFL7 within Microenvironment of High Grade Ovarian Serous Carcinoma on Infiltration of CD4+ and CD8+ Lymphocytes. Medicina, 58(5), 588. https://doi.org/10.3390/medicina58050588