TZAP Mutation Leads to Poor Prognosis of Patients with Breast Cancer †


Abstract
1. Introduction
2. Materials and Methods
2.1. Patients and DNA Extraction
2.2. Telomeric Zinc Finger-Associated Protein (TZAP) Mutation Analysis
2.3. Telomere Length (TL) Analysis
2.4. Statistical Analyses
3. Results
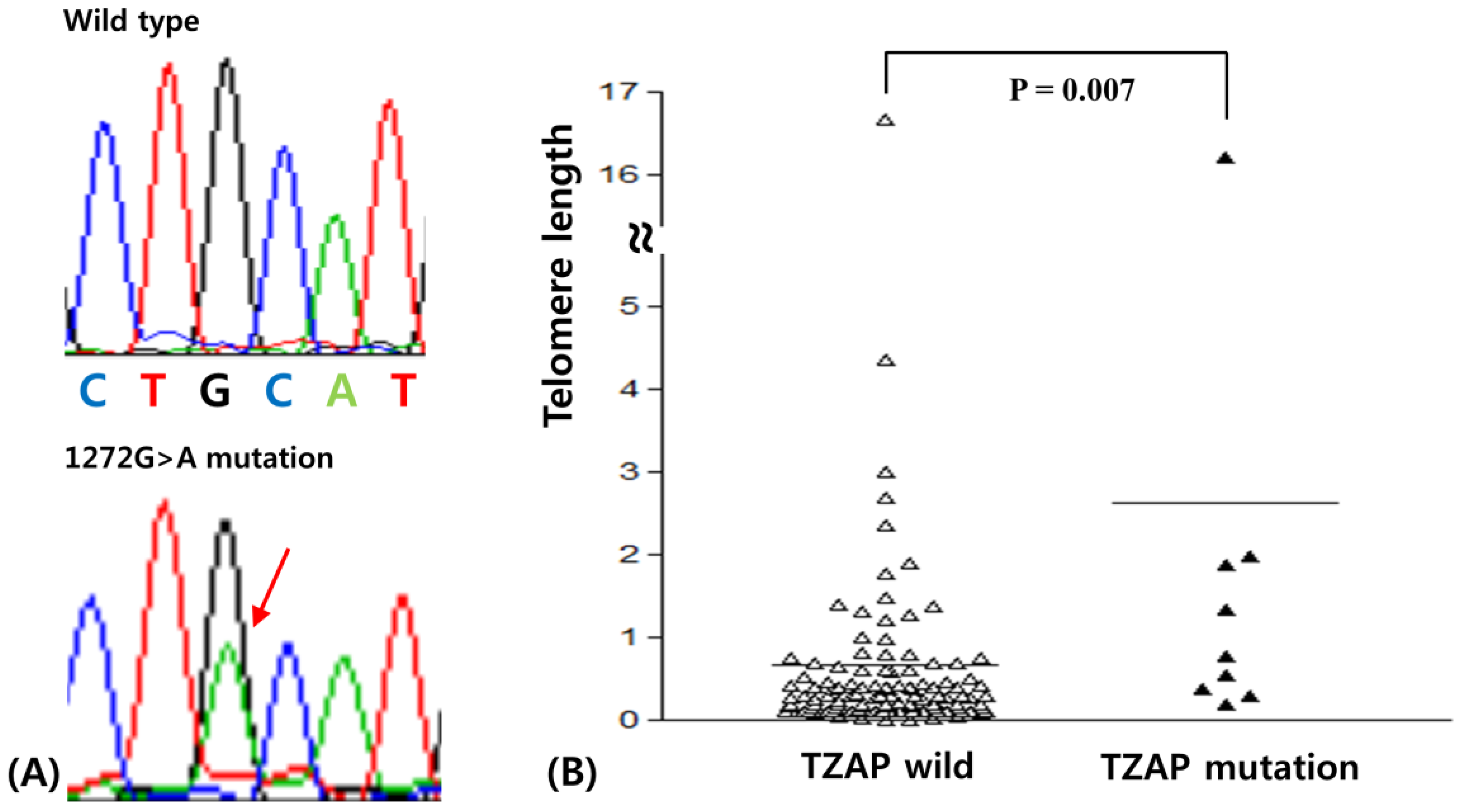
3.1. The TZAP Mutation in Breast Cancer
3.2. TL in Breast Cancer
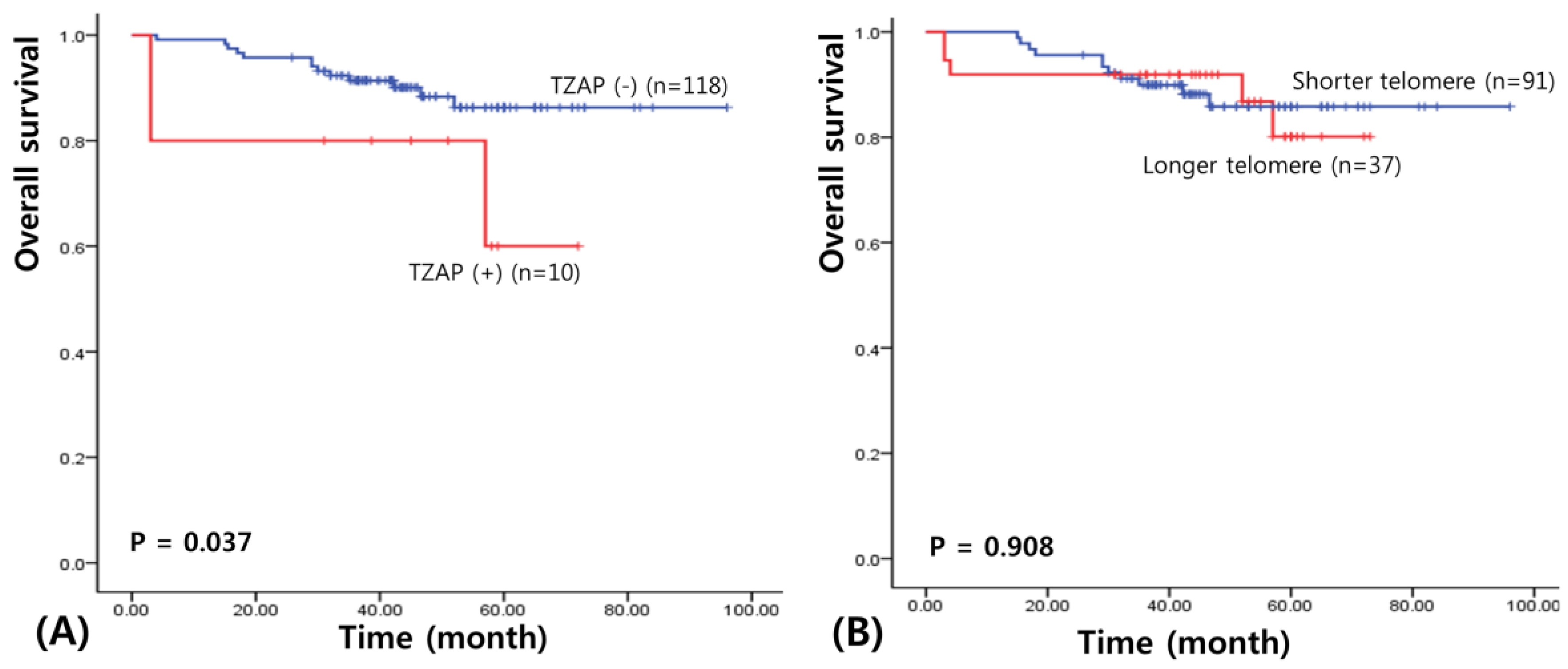
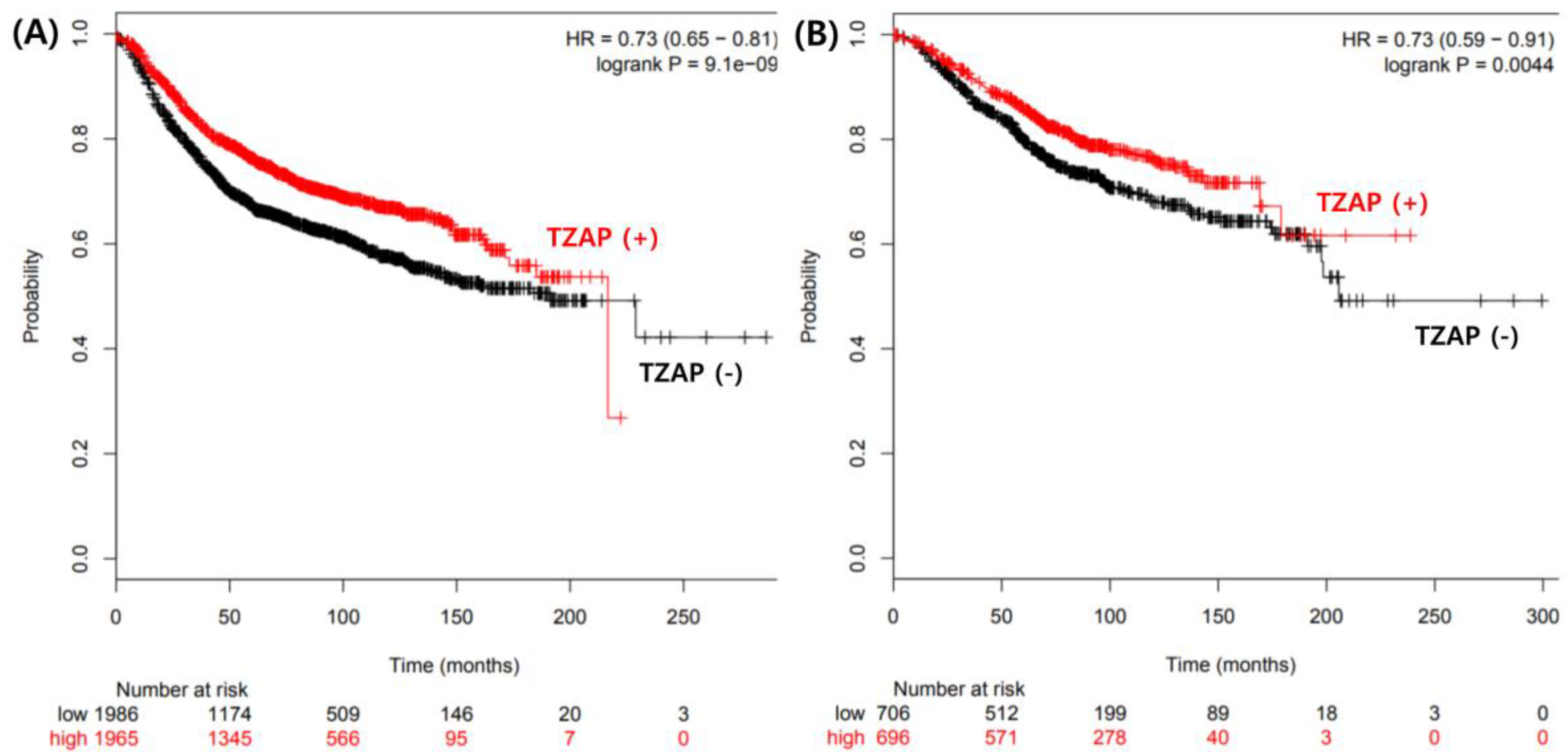
3.3. Prognostic Value of the TZAP Mutation in BC
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular portraits of human breast tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Petruccelli, N.; Daly, M.B.; Feldman, G.L. Hereditary breast and ovarian cancer due to mutations in BRCA1 and BRCA2. Genet. Med. 2010, 12, 245–259. [Google Scholar] [CrossRef] [PubMed]
- Reis-Filho, J.S.; Lakhani, S.R. The diagnosis and management of preinvasive breast disease: Genetic alterations in pre-invasive lesions. Breast Cancer Res. 2003, 5, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Fatah, T.M.; Powe, D.G.; Hodi, Z.; Reis-Filho, J.S.; Lee, A.H.; Ellis, I.O. Morphologic and molecular evolutionary pathways of low nuclear grade invasive breast cancers and their putative precursor lesions: Further evidence to support the concept of low nuclear grade breast neoplasia family. Am. J. Surg. Pathol. 2008, 32, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Rakha, E.A.; Reis-Filho, J.S.; Ellis, I.O. Basal-like breast cancer: A critical review. J. Clin. Oncol. 2008, 26, 2568–2581. [Google Scholar] [CrossRef] [PubMed]
- Harley, C.B.; Futcher, A.B.; Greider, C.W. Telomeres shorten during ageing of human fibroblasts. Nature 1990, 345, 458–460. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, E.H.; Greider, C.W.; Szostak, J.W. Telomeres and telomerase: The path from maize, Tetrahymena and yeast to human cancer and aging. Nat. Med. 2006, 12, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Liu, D.; Songyang, Z. The telosome/shelterin complex and its functions. Genome Biol. 2008, 9, 232. [Google Scholar] [CrossRef] [PubMed]
- Shay, J.W.; Wright, W.E. Role of telomeres and telomerase in cancer. Semin. Cancer Biol. 2011, 21, 349–353. [Google Scholar] [CrossRef] [PubMed]
- Jafri, M.A.; Ansari, S.A.; Alqahtani, M.H.; Shay, J.W. Roles of telomeres and telomerase in cancer, and advances in telomerase-targeted therapies. Genome Med. 2016, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Pickett, H.A.; Cesare, A.J.; Johnston, R.L.; Neumann, A.A.; Reddel, R.R. Control of telomere length by a trimming mechanism that involves generation of t-circles. EMBO J. 2009, 28, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Li, J.S.; Miralles, F.J.; Simavorian, T.; Bartocci, C.; Tsai, J.; Karlseder, J.; Lazzerini, D.E. TZAP: A telomere-associated protein involved in telomere length control. Science 2017, 355, 638–641. [Google Scholar] [CrossRef] [PubMed]
- Maris, J.M.; Jensen, S.J.; Sulman, E.P.; Beltinger, C.P.; Gates, K.; Allen, C.; Biegel, J.A.; Brodeur, G.M.; White, P.S. Cloning, chromosomal localization, physical mapping, and genomic characterization of HKR3. Genomics 1996, 35, 289–298. [Google Scholar] [CrossRef] [PubMed]
- White, P.S.; Maris, J.M.; Sulman, E.P.; Jensen, S.J.; Kyemba, S.M.; Beltinger, C.P.; Allen, C.; Kramer, D.L.; Biegel, J.A.; Brodeur, G.M. Molecular analysis of the region of distal 1p commonly deleted in neuroblastoma. Eur. J. Cancer 1997, 33, 1957–1961. [Google Scholar] [CrossRef]
- Jahn, A.; Rane, G.; Paszkowski-Rogacz, M.; Sayols, S.; Bluhm, A.; Han, C.T.; Draškovič, I.; Londoño-Vallejo, J.A.; Kumar, A.P.; Buchholz, F. ZBTB48 is both a vertebrate telomere-binding protein and a transcriptional activator. EMBO Rep. 2017, 18, 929–946. [Google Scholar] [CrossRef] [PubMed]
- The COSMIC, the Catalogue of Somatic Mutations in Cancer. Available online: http://cancer.sanger.ac.uk/cosmic/mutation/overview?id=3705990/ (accessed on 01 February 2019).
- Thriveni, K.; Raju, A.; Kumar, R.V.; Krishnamurthy, S.; Chaluvarayaswamy, R. Patterns of Relative Telomere Length in Associated with hTERT Gene Expression in the Tissue of Breast Cancer Patients. Clin. Breast Cancer 2018, 19, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Kim, H.; Verhaak, R.G.W. Silent Mutations Make Some Noise. Cell 2014, 156, 1129–1131. [Google Scholar] [CrossRef] [PubMed]
- Lossaint, G.; Lingner, J. TZAP or not to zap telomeres. Science 2017, 355, 578–579. [Google Scholar] [CrossRef] [PubMed]
- Zlotorynski, E. Chromosome biology: TZAP trims telomeres. Nat. Rev. Mol. Cell Biol. 2017, 18, 140. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Primer Sequences | Exon | |
|---|---|---|
| TZAP 1 | F: CCAGACCTCAACAGCACAGA R: CACAGCCCACGAACCTAGTG | 1 |
| TZAP 2 | F: ATCCCATTGGCCGTTCTCT R: CCGGCACAGTGAGAGGAT | 2 |
| TZAP 3 | F: TAGAGGCCAACTTCCCGTTT R: CCTGGGCACAGTACCTCATT | 3 |
| TZAP 4 | F: CCTGCTGATTCATTTGGTGA R: GGAATGGCAGACAGGAAAAG | 4 |
| TZAP 5 | F: GGAGGTGAGGAAGTTGACCA R: CCCTTCTAAGGGGAACAAGTG | 5 |
| TZAP 6 | F: GCTTGTCCCTGCACCTTAAC R: GGAGAGGGCAACACATAACC | 5 |
| TZAP 7 | F: AGTCTGTCTGGGCCTGAGAA R: CCCTCCCTGTCACTTACTGC | 5 |
| TZAP 8 | F: CCCTTCCCTGCTCTCACC R: AAGAGAGAACGGGCGACAC | 6 |
| TZAP 9 | F: GTCACTTCCCTTGGTGATGG R: GAGGGGACCAGTGGTTTACA | 7 |
| TZAP 10 | F: CTGGGTGGCACTGGAGAG R: CACGGGAACAGACTGTCAGG | 8 |
| Telomere length | F: CGGTTTGTTTGGGTTTGGGTTTGGGTTTGGGTTTGGGTT R: GGCTTGCCTTACCCTTACCCTTACCCTTACCCTTACCCT | |
| β-globin | F: TGTGCTGGCCCATCACTTTG R: ACCAGCCA-CCACTTTCTGATAGG |
| TZAP | |||
|---|---|---|---|
| Wild (N, %) | Mutant (N, %) | p | |
| Total | 118 (92.2) | 10 (7.8) | |
| Age | 0.253 | ||
| <65 | 80 (94.1) | 5 (5.9) | |
| ≥65 | 38 (88.4) | 5 (11.6) | |
| BMI | 0.494 | ||
| <25 | 44 (91.7) | 4 (8.3) | |
| ≥25 | 32 (86.5) | 5 (13.5) | |
| T stage | 0.706 | ||
| T1 | 35 (89.7) | 4 (10.3) | |
| T2 | 77 (93.9) | 5 (6.1) | |
| T3 | 5 (83.3) | 1 (16.7) | |
| T4 | 1 (100) | 0 (0) | |
| N stage | 0.035 | ||
| N0 | 69 (87.3) | 10 (12.7) | |
| N1 | 33 (100) | 0 (0) | |
| N2 | 16 (100) | 0 (0) | |
| Subtype | 0.259 | ||
| Luminal A | 43 (87.8) | 6 (12.2) | |
| Luminal B | 38 (97.4) | 1 (2.6) | |
| HER2-enriched | 11 (100) | 0 (0) | |
| TN | 26 (89.7) | 3 (10.6) | |
| Telomere length | 0.064 | ||
| Short | 86 (95.6) | 4 (4.4) | |
| Long | 32 (84.2) | 6 (15.8) | |
| Telomere Length | |||
|---|---|---|---|
| Short (N, %) | Long (N, %) | p | |
| Total | 90 (70.3) | 38 (29.7) | |
| Age | 0.613 | ||
| <65 | 61 (71.8) | 24 (28.2) | |
| ≥65 | 29 (67.4) | 14 (32.6) | |
| BMI | 0.127 | ||
| <25 | 36 (75.0) | 12 (25.0) | |
| ≥25 | 22 (59.5) | 15 (40.5) | |
| T stage | 0.647 | ||
| T1 | 28 (71.8) | 11 (28.2) | |
| T2 | 58 (70.7) | 24 (29.3) | |
| T3 | 3 (50.0) | 3 (50.0) | |
| T4 | 1 (100) | 0 (0) | |
| N stage | 0.314 | ||
| N0 | 63 (79.7) | 16 (20.3) | |
| N1 | 23 (69.7) | 10 (30.3) | |
| N2 | 14 (87.5) | 2 (12.5) | |
| Subtype | <0.001 | ||
| Luminal A | 28 (57.1) | 21 (42.9) | |
| Luminal B | 39 (100) | 0 (0) | |
| HER2-enriched | 11 (100) | 0 (0) | |
| TN | 22 (75.9) | 7 (24.1) | |
| TZAP mutation | 0.064 | ||
| Wild | 86 (72.9) | 32 (27.1) | |
| Mutant | 4 (40.0) | 6 (60.0) | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heo, Y.-R.; Lee, M.-H.; Kwon, S.-Y.; Cho, J.; Lee, J.-H. TZAP Mutation Leads to Poor Prognosis of Patients with Breast Cancer †. Medicina 2019, 55, 748. https://doi.org/10.3390/medicina55110748
Heo Y-R, Lee M-H, Kwon S-Y, Cho J, Lee J-H. TZAP Mutation Leads to Poor Prognosis of Patients with Breast Cancer †. Medicina. 2019; 55(11):748. https://doi.org/10.3390/medicina55110748
Chicago/Turabian StyleHeo, Yu-Ran, Moo-Hyun Lee, Sun-Young Kwon, Jihyoung Cho, and Jae-Ho Lee. 2019. "TZAP Mutation Leads to Poor Prognosis of Patients with Breast Cancer †" Medicina 55, no. 11: 748. https://doi.org/10.3390/medicina55110748
APA StyleHeo, Y.-R., Lee, M.-H., Kwon, S.-Y., Cho, J., & Lee, J.-H. (2019). TZAP Mutation Leads to Poor Prognosis of Patients with Breast Cancer †. Medicina, 55(11), 748. https://doi.org/10.3390/medicina55110748