
Response of Soil Net Nitrogen Mineralization to a Litter in Three Subalpine Forests

Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
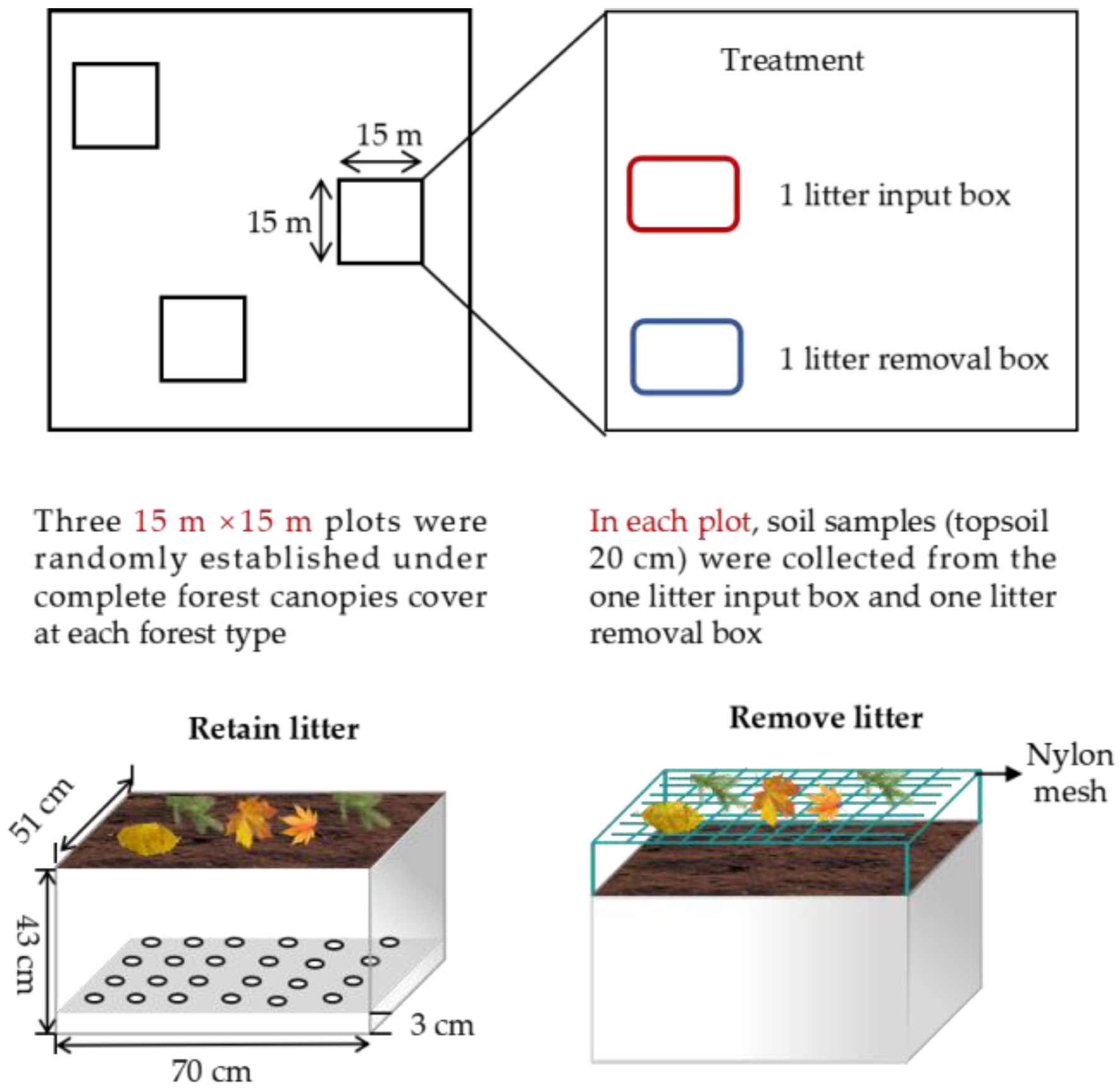
2.2. Experimental Design
2.3. Samples Collection
2.4. Microclimate and Soil Biochemical Analyses
2.5. Data Calculations and Statistical Analysis
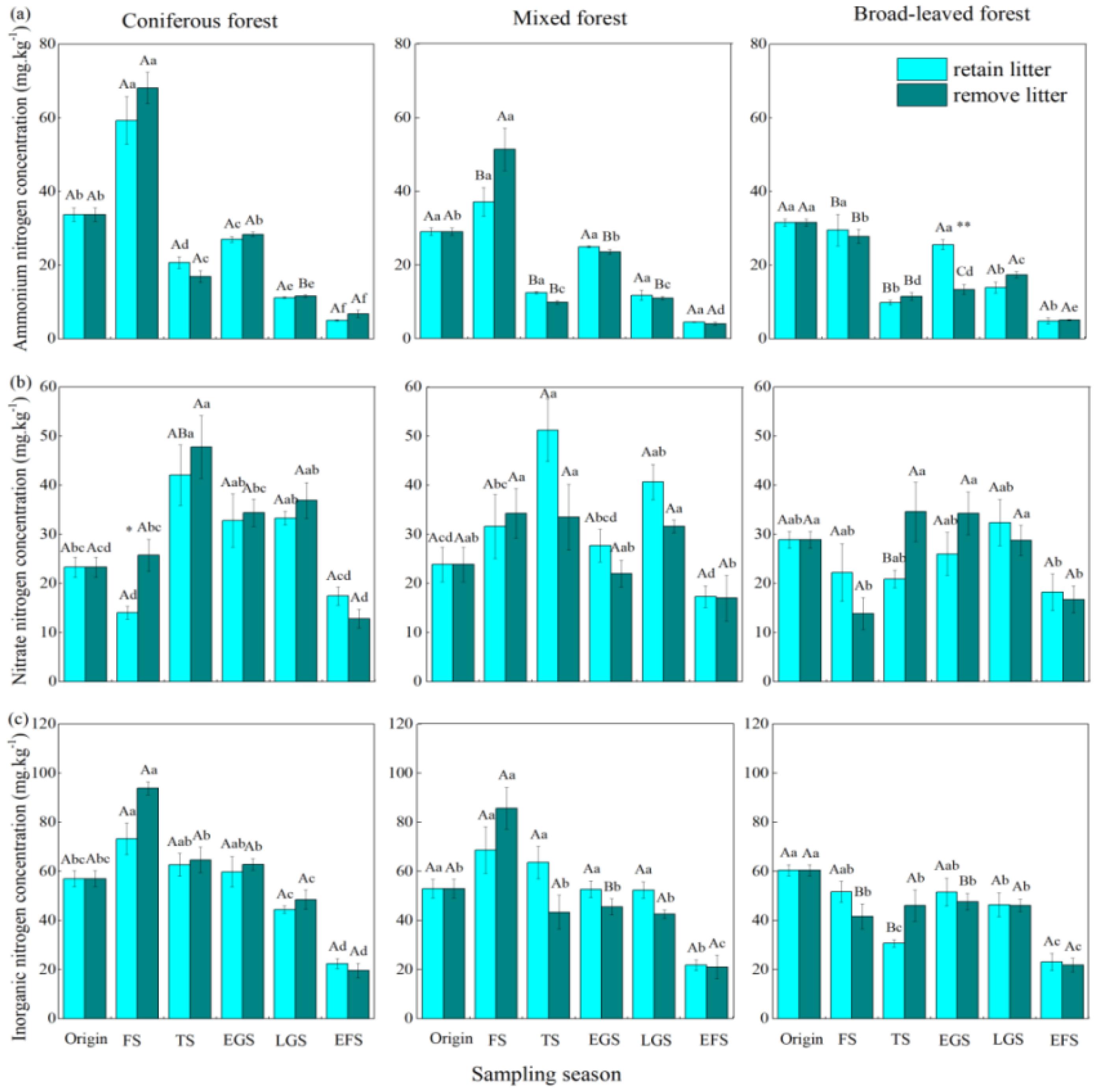
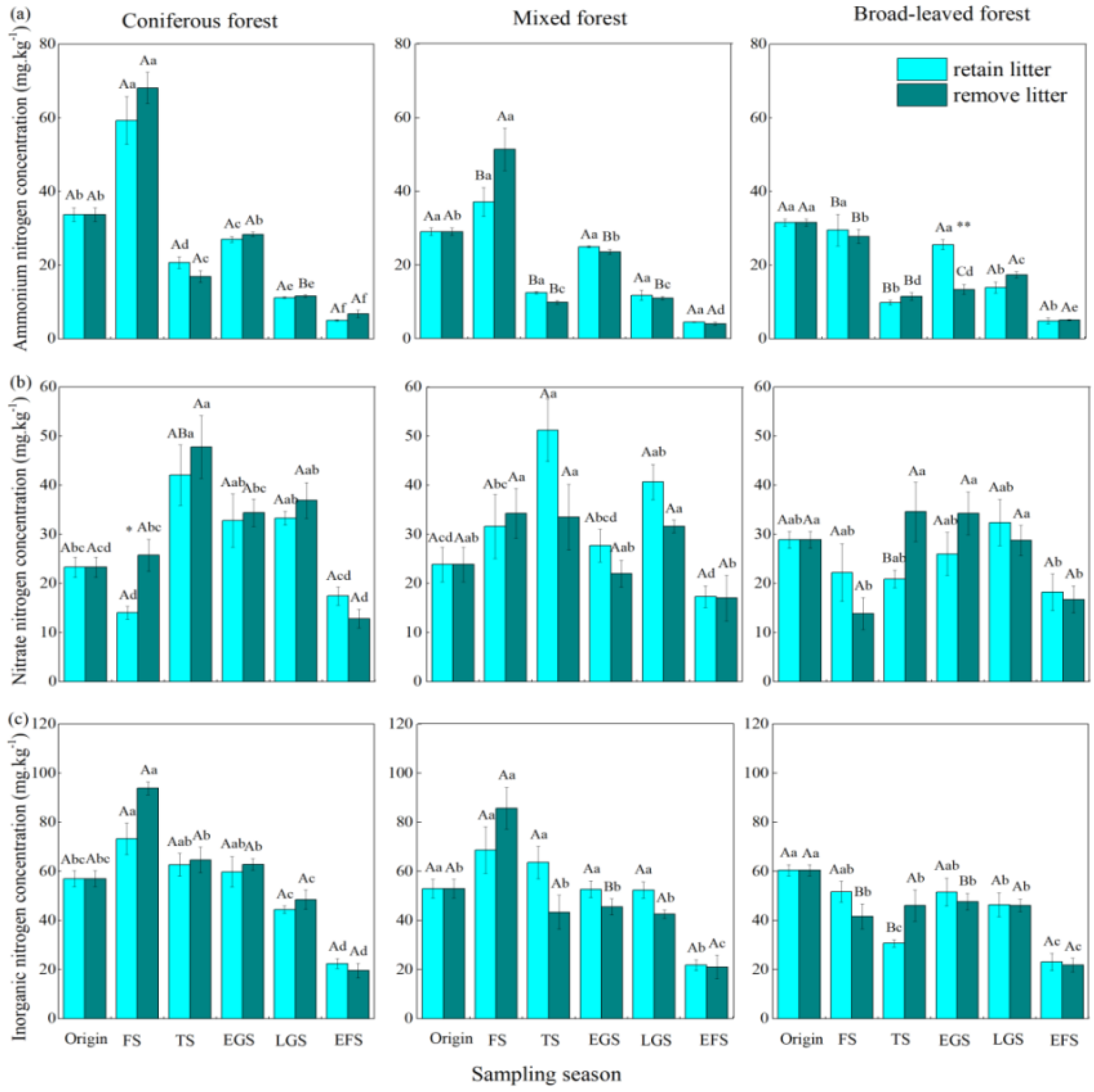
3. Results
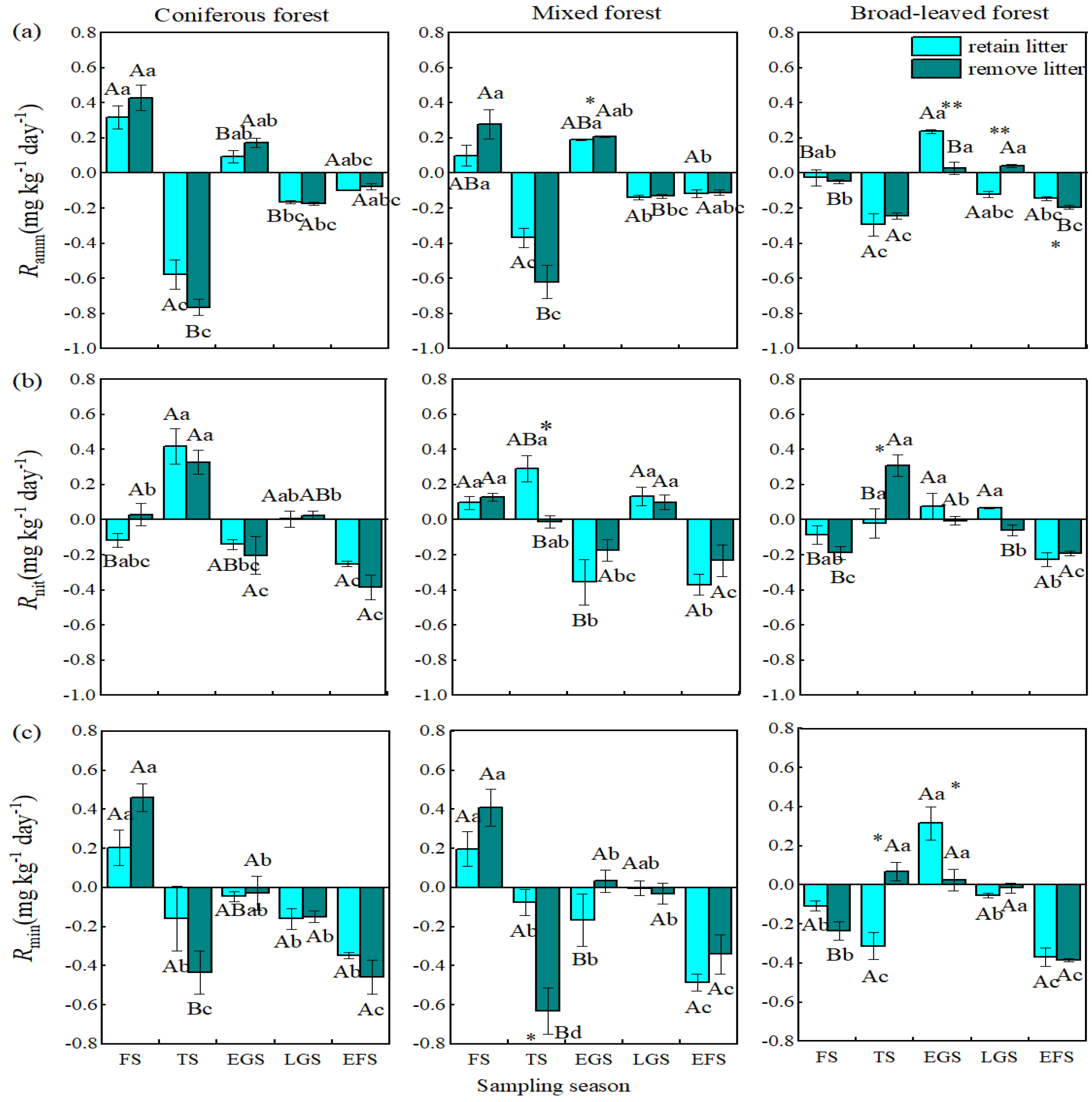
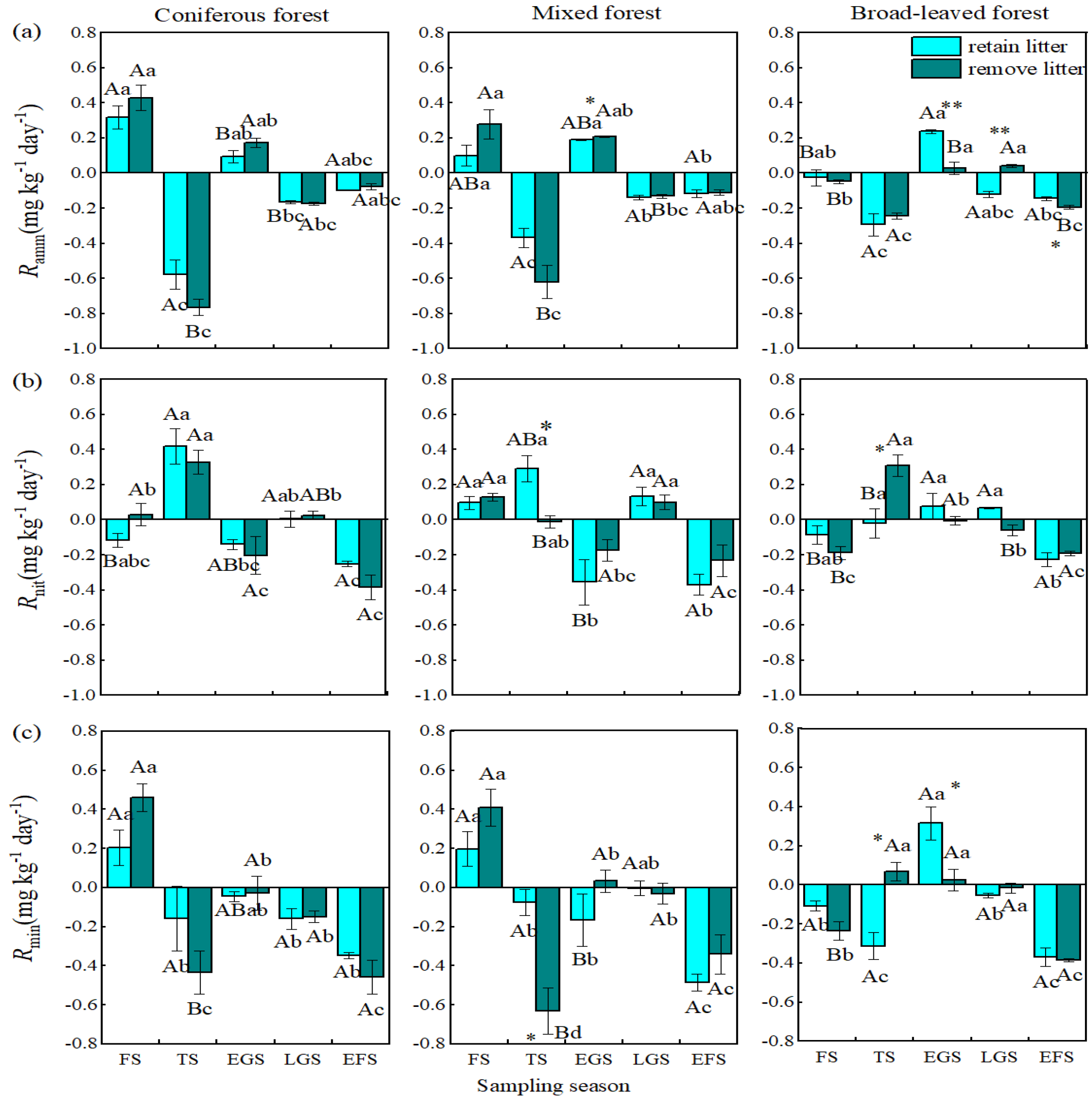
3.1. Net Ammonification Rate
3.2. Net Nitrification Rate
3.3. Net Nitrogen Mineralization Rate
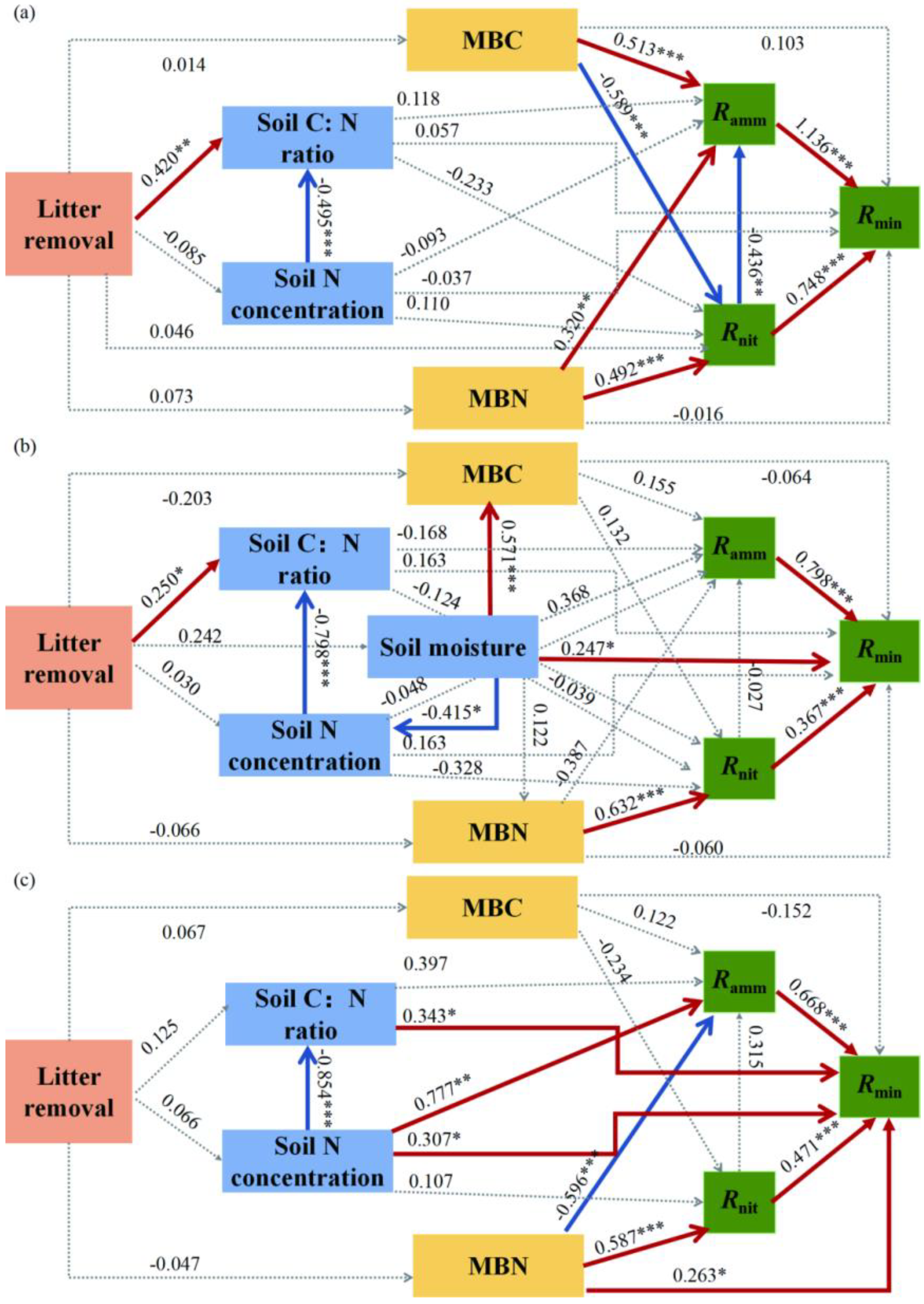
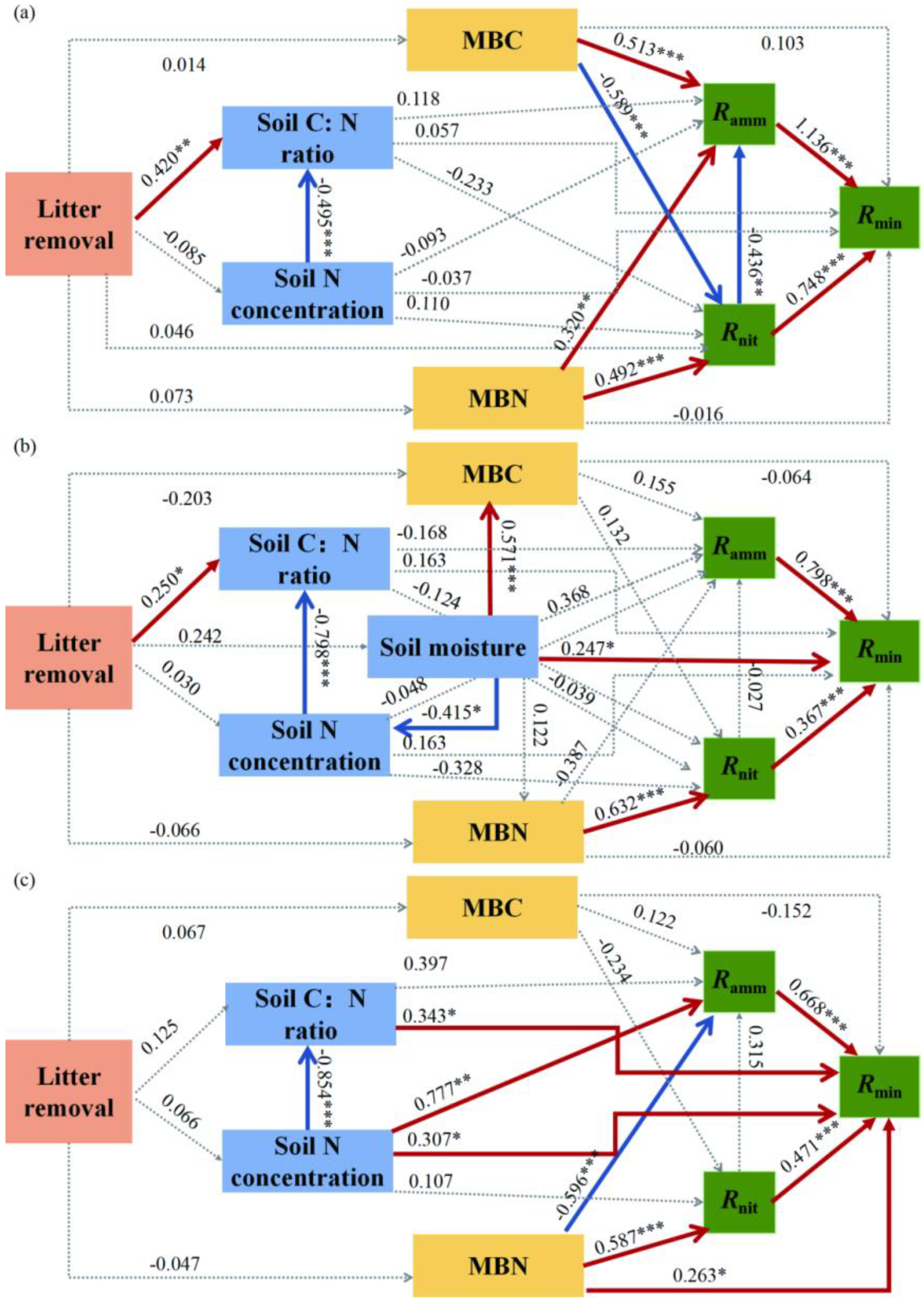
3.4. Driving Factors Difference among Different Forest Types
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hosokawa, N.; Isobe, K.; Urakawa, R.; Tateno, R.; Fukuzawa, K.; Watanabe, T.; Shibata, H. Effect of root litter addition on nitrogen mineralization rate under laboratory low-temperature conditions in soil from a Japanese northern hardwood forest. Ecol. Res. 2020, 35, 888–899. [Google Scholar] [CrossRef]
- Elrys, A.S.; Ali, A.; Zhang, H.; Cheng, Y.; Zhang, J.; Cai, Z.C.; Müller, C.; Chang, S.X. Patterns and drivers of global gross nitrogen mineralizaton in soils. Glob. Chang. Biol. 2021, 27, 5950–5962. [Google Scholar] [CrossRef] [PubMed]
- Keuper, F.; Dorrepaal, E.; van Bodegom, P.M.; van Logtestijn, R.; Venhuizen, G.; van Hal, J.; Aerts, R. Experimentally increased nutrient availability at the permafrost thaw front selectively enhances biomass production of deep-rooting subarctic peatland species. Glob. Chang. Biol. 2017, 23, 4257–4266. [Google Scholar] [PubMed]
- Yan, Y.; Fang, S.; Tian, Y.; Deng, S.; Tang, L.; Chuong, D. Influence of tree spacing on soil nitrogen mineralization and availability in hybrid poplar plantations. Forests 2015, 6, 636–649. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Li, Y.; Wang, M.; Zhou, C.; Cao, G.; Shi, P.; Song, M. Litter species traits, but not richness, contribute to carbon and nitrogen dynamics in an alpine meadow on the Tibetan Plateau. Plant Soil 2013, 373, 931–941. [Google Scholar]
- Berg, B.; McClaugherty, C.A. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Buckeridge, K.M.; Zufelt, E.; Chu, H.; Grogan, P. Soil nitrogen cycling rates in low arctic shrub tundra are enhanced by litter feedbacks. Plant Soil 2010, 330, 407–421. [Google Scholar] [CrossRef]
- Bai, X.; Dippold, M.A.; An, S.; Wang, B.; Zhang, H.; Loeppmann, S. Extracellular enzyme activity and stoichiometry: The effect of soil microbial element limitation during leaf litter decomposition. Ecol. Indic. 2021, 121, 107200. [Google Scholar] [CrossRef]
- Shi, L.; Feng, W.; Jing, X.; Zang, H.; Mortimer, P.; Zou, X. Contrasting responses of soil fungal communities and soil respiration to the above- and below-ground plant C inputs in a subtropical forest. Eur. J. Soil Sci. 2018, 70, 751–764. [Google Scholar] [CrossRef]
- Brant, J.B.; Sulzman, E.W.; Myrold, D.D. Microbial community utilization of added carbon substrates in response to long-term carbon input manipulation. Soil Biol. Biochem. 2006, 38, 2219–2232. [Google Scholar] [CrossRef]
- Song, Q.; Ouyang, M.; Yang, Q.; Lu, H.; Yang, G.; Chen, F.; Shi, J. Degradation of litter quality and decline of soil nitrogen mineralization after moso bamboo (Phyllostachys pubscens) expansion to neighboring broadleaved forest in subtropical China. Plant Soil 2016, 404, 113–124. [Google Scholar] [CrossRef]
- Song, Y.; Zou, Y.; Wang, G.; Yu, X. Altered soil carbon and nitrogen cycles due to the freeze-thaw effect: A meta-analysis. Soil Biol. Biochem. 2017, 109, 35–49. [Google Scholar] [CrossRef]
- Sorensen, P.O.; Beller, H.R.; Bill, M.; Bouskill, N.J.; Hubbard, S.S.; Karaoz, U.; Polussa, A.; Steltzer, H.; Wang, S.; Williams, K.H.; et al. The snowmelt niche differentiates three microbial life strategies that influence soil nitrogen availability during and after winter. Front. Microbiol. 2020, 11, 871. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Xie, Z.; Xu, W.; Zhao, C. Controls over leaf litter decomposition in a mixed evergreen and deciduous broad-leaved forest, Central China. Plant Soil 2017, 412, 345–355. [Google Scholar] [CrossRef]
- Xia, S.; Chen, J.; Schaefer, D.; Goodale, U.M. Effect of topography and litterfall input on fine-scale patch consistency of soil chemical properties in a tropical rainforest. Plant Soil 2016, 404, 385–398. [Google Scholar] [CrossRef]
- Wei, X.; Yang, Y.; Shen, Y.; Chen, Z.; Dong, Y.; Wu, F.; Zhang, L. Effects of litterfall on the accumulation of extracted soil humic substances in subalpine forests. Front. Plant Sci. 2020, 11, 254. [Google Scholar] [CrossRef]
- Yang, W.Q.; Wang, K.Y.; Kellomäki, S.; Gong, H.D. Litter dynamics of three subalpine forests in western Sichuan. Pedosphere 2005, 15, 653–659. [Google Scholar]
- Vervaet, H.; Massart, B.; Boeckx, P.; Van Cleemput, O.; Hofman, G. Use of principal component analysis to assess factors controlling net N mineralization in deciduous and coniferous forest soils. Biol. Fertil. Soils 2002, 36, 93–101. [Google Scholar] [CrossRef]
- Li, H.; Wu, F.; Yang, W.; Xu, L.; Ni, X.; He, J.; Tan, B.; Hu, Y. Effects of forest gaps on litter lignin and cellulose dynamics vary seasonally in an Alpine forest. Forests 2016, 7, 27. [Google Scholar] [CrossRef] [Green Version]
- Zhan, P.; Liu, Y.; Wang, H.; Wang, C.; Xia, M.; Wang, N.; Cui, W.; Xiao, D.; Wang, H. Plant litter decomposition in wetlands is closely associated with phyllospheric fungi as revealed by microbial community dynamics and co-occurrence network. Sci. Total Environ. 2021, 753, 142194. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Wu, F.; Yang, W.; Zhang, J.; Deng, R. Litter decomposition in two subalpine forests during the freeze-thaw season. Acta Oecol. 2010, 36, 135–140. [Google Scholar] [CrossRef]
- Chen, Z.; Shen, Y.; Tan, B.; Li, H.; You, C.; Xu, Z.; Wei, X.; Ni, X.; Yang, Y.; Zhang, L. Decreased Soil Organic Carbon under Litter Input in Three Subalpine Forests. Forests 2021, 12, 1479. [Google Scholar] [CrossRef]
- Yang, W.; Wang, K.; Kellomäk, S.; Zhang, J. Annual and monthly variations in litter macronutrients of three subalpine forests in western China. Pedosphere 2006, 16, 788–798. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, L.; Wei, X.; Chen, Y.; Yang, W.; Tan, B.; Yue, K.; Ni, X.; Wu, F. Litter removal reduced soil nitrogen mineralization in repeated freeze-thaw cycles. Sci. Rep. 2019, 9, 2052. [Google Scholar] [CrossRef] [PubMed]
- International World Resource Base. World Reference Base for Soil Resources 2014, update 2015 International soil classification system for naming soils and creating legends for soil maps. In World Soil Resources Reports No. 106; FAO: Rome, Italy, 2015. [Google Scholar]
- Tan, B.; Wu, F.; Yang, W.Q.; Yu, S.; Liu, L.; Wang, A.; Yang, Y.L. Activities of soil oxidordeuctase and their response to seasonal freeze-thaw in the subalpine/alpine forests of western Sichuan. Acta Ecol. Sin. 2012, 32, 6670–6678. [Google Scholar] [CrossRef]
- Dong, M. Survey, Observation and Analysis of Terrestrial Biocommunities; Standards Press of China: Beijing, China, 1997. [Google Scholar]
- LY/T 1211-1999; Field sampling and preparation of forest plant and forest floor samples. China Academy of Forestry, Forest Research Institute: Beijing, China, 1999.
- Inglett, P.W.; Inglett, K.S. Biogeochemical changes during early development of restored calcareous wetland soils. Geoderma 2013, 192, 132–141. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Lu, R.K. Soil and Agro-Chemical Analytical Methods; China Agricultural Science and Technology: Beijing, China, 1999. [Google Scholar]
- Ojeda, C.B.; Rojas, F.S. Recent developments in derivative ultraviolet/visible absorption spectrophotometry. Anal. Chim. Acta 2004, 518, 1–24. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, Q.; Yin, H. Effects of temperature on soil net nitrogen mineralisation in two contrasting forests on the eastern Tibetan Plateau, China. Soil Res. 2014, 52, 562. [Google Scholar] [CrossRef]
- You, C.; Wu, F.; Yang, W.; Xu, Z.; Tan, B.; Li, Z.; Kai, Y.; Ni, X.; Li, H.; Chang, C. Does foliar nutrient resorption regulate the coupled relationship between nitrogen and phosphorus in plant leaves in response to nitrogen deposition? Sci. Total Environ. 2018, 645, 733–742. [Google Scholar] [CrossRef]
- Yue, K.; Peng, Y.; Fornara, D.A.; Meerbeek, K.V.; Vesterdal, L.; Yang, W.; Peng, C.; Tan, B.; Zhou, W.; Xu, Z. Responses of nitrogen concentrations and pools to multiple environmental change drivers: A meta-analysis across terrestrial ecosystems. Glob. Ecol. Biogeogr. 2019, 28, 690–724. [Google Scholar] [CrossRef]
- Hatton, P.J.; Castanha, C.; Torn, M.S.; Bird, J.A. Litter type control on soil C and N stabilization dynamics in a temperate forest. Glob. Chang. Biol. 2015, 21, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Zhao, Y.; Xiao, D.; Xu, Z.; Zhang, W.; Xiao, J.; Wang, K. Dynamics of soil nitrogen availability following vegetation restoration along a climatic gradient of a subtropical karst region in China. J. Soil Sediments 2021, 21, 2167–2178. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, C.; He, N.; Wen, X.; Gao, Y.; Li, S.; Niu, S.; Butterbach-Bahl, K.; Luo, Y.; Yu, G. A global synthesis of the rate and temperature sensitivity of soil nitrogen mineralization: Latitudinal patterns and mechanisms. Glob. Chang. Biol. 2017, 23, 455–464. [Google Scholar] [CrossRef]
- Deng, J.; Zhu, W.; Zhou, Y.; Yin, Y. Soil organic carbon chemical functional groups under different revegetation types are coupled with changes in the microbial community composition and the functional genes. Forests 2019, 10, 240. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Zhang, J.; Meng, M.; Guo, X.; Wu, Y.; Liu, X.; Zhao, K.; Ding, L.; Shao, Y.; Fu, W. Forest-type shift and subsequent intensive management affected soil organic carbon and microbial community in southeastern China. Eur. J. For. Res. 2017, 136, 689–697. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, W.; Chen, H.; Mo, J. Impacts of nitrogen deposition on soil nitrogen cycle in forest ecosystems: A review. Acta Ecol. Sin. 2015, 35, 35–43. [Google Scholar] [CrossRef]
- Strickland, M.S.; Rousk, J. Considering fungal:bacterial dominance in soils-methods, controls, and ecosystem implications. Soil Biol. Biochem. 2010, 42, 1385–1395. [Google Scholar] [CrossRef]
- Edwards, K.A.; McCulloch, J.; Peter Kershaw, G.; Jefferies, R.L. Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring. Soil Biol. Biochem. 2006, 38, 2843–2851. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, N.; Xiao, J.; Zhao, C.; Zou, T.; Li, D.; Liu, Q.; Yin, H. Changes in plant nitrogen acquisition strategies during the restoration of spruce plantations on the eastern Tibetan Plateau, China. Soil Biol. Biochem. 2018, 119, 50–58. [Google Scholar] [CrossRef]
- Xu, B.; Wang, J.; Wu, N.; Wu, Y.; Shi, F. Seasonal and interannual dynamics of soil microbial biomass and available nitrogen in an alpine meadow in the eastern part of Qinghai-Tibet Plateau, China. Biogeosciences 2018, 15, 567–579. [Google Scholar] [CrossRef] [Green Version]
- Walker, V.K.; Palmer, G.R.; Voordouw, G. Freeze-thaw tolerance and clues to the winter survival of a soil community. Appl. Environ. Microb. 2006, 72, 1784–1792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Wang, J.; Chang, S.X.; Cai, Z.; Müller, C. Nitrogen deposition affects both net and gross soil nitrogen transformations in forest ecosystem: A review. Environ. Pollut. 2019, 244, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Churchland, C.; Mayo-Bruinsma, L.; Ronson, A.; Grogan, P. Soil microbial and plant community responses to single large carbon and nitrogen additions in low arctic tundra. Plant Soil 2010, 334, 409–421. [Google Scholar] [CrossRef]
- Wu, H.; Cai, A.; Xing, T.; Huai, S.; Zhu, P.; Xu, M.; Lu, C. Fertilization enhances mineralization of soil carbon and nitrogen pools by regulating the bacterial community and biomass. J. Soil Sediments 2021, 21, 1633–1643. [Google Scholar] [CrossRef]
- Pandey, C.B.; Rai, R.B.; Singh, L. Seasonal dynamics of mineral N pools and N-mineralization in soils under homegarden trees in South Andaman, India. Agrofor. Syst. 2007, 71, 57–66. [Google Scholar] [CrossRef]
- Tang, H.L.; Wang, J.Y.; Huang, S.; Gong, W.; Zhou, Y.B. Responses of soil nitrogen mineralization of evergreen broad-leaved forest in rainy area of western China to moisture and temperature. J. Gansu Agric. Univ. 2019, 2, 124–131. [Google Scholar]
- Ros, G.H.; Hanegraaf, M.C.; Hoffland, E.; van Riemsdijk, W.H. Predicting soil N mineralization: Relevance of organic matter fractions and soil properties. Soil Biol. Biochem. 2011, 43, 1714–1722. [Google Scholar] [CrossRef]
- Niu, S.; Classen, A.T.; Dukes, J.S.; Kardol, P.; Liu, L.; Luo, Y.; Rustad, L.; Sun, J.; Tang, J.; Templer, P.H.; et al. Global patterns and substrate-based mechanisms of the terrestrial nitrogen cycle. Ecol. Lett. 2016, 19, 697–709. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, L.; Zhao, W.; Hu, H.; Feng, X.; Müller, C.; Cai, Z. Soil gross nitrogen transformations along the Northeast China Transect (NECT) and their response to simulated rainfall events. Sci. Rep. 2016, 6, 22830. [Google Scholar] [CrossRef] [Green Version]
- Janssen, B.H. Nitrogen mineralization in relation to C/N ratio and decomposability of organic materials. Plant Soil 1996, 181, 39–45. [Google Scholar] [CrossRef]
- Bengtsson, G.; Bengtson, P.; Månsson, K.F. Gross nitrogen mineralization-, immobilization-, and nitrification rates as a function of soil C/N ratio and microbial activity. Soil Biol. Biochem. 2003, 35, 143–154. [Google Scholar] [CrossRef]
- Bonito, G.M.; Coleman, D.C.; Haines, B.L.; Cabrera, M.L. Can nitrogen budgets explain differences in soil nitrogen mineralization rates of forest stands along an elevation gradient? For. Ecol. Manag. 2003, 176, 563–574. [Google Scholar] [CrossRef]
- Chapman, L.Y.; McNulty, S.G.; Sun, G.; Zhang, Y. Net nitrogen mineralization in natural ecosystems across the conterminous US. Int. J. Geosci. 2013, 4, 1300–1312. [Google Scholar] [CrossRef] [Green Version]
- Zaman, M.; Chang, S.X. Substrate type, temperature, and moisture content affect gross and net N mineralization and nitrification rates in agroforestry systems. Biol. Fertil. Soils 2004, 39, 269–279. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Inorganic N | Ramm | Rnit | Rmin | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | P | F | P | F | P | F | P | F | P | F | P | |
| LT | 0.44 | 0.52 | 0.01 | 0.93 | 0.02 | 0.90 | 3.05 | 0.10 | 0.23 | 0.64 | 1.34 | 0.26 |
| FT | 42.68 | 0.00 *** | 0.99 | 0.39 | 6.99 | 0.01 * | 3.95 | 0.04 * | 0.89 | 0.43 | 0.09 | 0.91 |
| SS | 304.55 | 0.00 *** | 43.86 | 0.00 *** | 115.49 | 0.00 *** | 193.82 | 0.00 *** | 46.83 | 0.00 *** | 42.22 | 0.00 *** |
| LT × FT | 1.587 | 0.24 | 0.98 | 0.40 | 0.83 | 0.46 | 1.49 | 0.265 | 2.36 | 0.137 | 0.43 | 0.66 |
| LT × SS | 4.845 | 0.03 * | 0.72 | 0.55 | 2.26 | 0.06 | 4.94 | 0.03 * | 0.30 | 0.804 | 1.96 | 0.15 |
| FT × SS | 22.88 | 0.00 *** | 7.94 | 0.00 *** | 12.79 | 0.00 *** | 20.70 | 0.00 *** | 7.25 | 0.00 *** | 8.48 | 0.00 *** |
| LT × FT × SS | 2.673 | 0.08 | 4.29 | 0.00 *** | 4.55 | 0.00 *** | 3.95 | 0.03 * | 4.58 | 0.00 ** | 6.41 | 0.00 *** |
| Coniferous Forest | Mixed Forest | Broad-Leaved Forest | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Ramm | Rnit | Rmin | Ramm | Rnit | Rmin | Ramm | Rnit | Rmin | ||
| Litter treatment | Total | 0.112 | −0.043 | 0.125 | 0.047 | −0.056 | 0.122 | 0.104 | −0.036 | 0.074 |
| Direct | 0.000 | 0.046 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Indirect | 0.112 | −0.089 | 0.125 | 0.047 | −0.056 | 0.122 | 0.104 | −0.036 | 0.074 | |
| Soil moisture | Total | - | - | - | 0.368 | 0.208 | 0.559 | - | - | - |
| Direct | - | - | - | 0.368 | −0.039 | 0.247 | - | - | - | |
| Indirect | - | - | - | 0.000 | 0.247 | 0.313 | - | - | - | |
| Soil N concentration | Total | −0.250 | 0.225 | −0.181 | 0.092 | −0.228 | 0.022 | 0.471 | 0.107 | 0.380 |
| Direct | −0.093 | 0.110 | −0.037 | −0.048 | −0.328 | 0.163 | 0.777 | 0.107 | 0.307 | |
| Indirect | −0.157 | 0.115 | −0.144 | 0.140 | 0.099 | −0.141 | −0.306 | 0.000 | 0.073 | |
| Soil C:N ratio | Total | 0.220 | −0.233 | 0.133 | −0.164 | −0.124 | −0.013 | 0.397 | 0.000 | 0.609 |
| Direct | 0.118 | −0.233 | 0.057 | −0.168 | −0.124 | 0.163 | 0.397 | 0.000 | 0.343 | |
| Indirect | 0.102 | 0.000 | 0.076 | 0.004 | 0.000 | −0.176 | 0.000 | 0.000 | 0.266 | |
| MBC | Total | 0.770 | −0.589 | 0.538 | 0.151 | 0.132 | 0.105 | 0.049 | −0.234 | −0.229 |
| Direct | 0.513 | −0.589 | 0.103 | 0.155 | 0.132 | −0.064 | 0.122 | −0.234 | −0.152 | |
| Indirect | 0.257 | 0.000 | 0.435 | −0.004 | 0.000 | 0.169 | −0.073 | 0.000 | −0.077 | |
| MBN | Total | 0.105 | 0.492 | 0.471 | −0.404 | 0.632 | −0.150 | −0.411 | 0.587 | 0.265 |
| Direct | 0.320 | 0.492 | −0.016 | −0.387 | 0.632 | −0.060 | −0.596 | 0.587 | 0.263 | |
| Indirect | −0.215 | 0.000 | 0.487 | −0.017 | 0.000 | −0.090 | 0.185 | 0.000 | 0.002 | |
| Ramm | Total | - | - | 1.136 | - | - | 0.798 | - | - | 0.668 |
| Direct | - | - | 1.136 | - | - | 0.798 | - | - | 0.668 | |
| Indirect | - | - | 0.000 | - | - | 0.000 | - | - | 0.000 | |
| Rnit | Total | −0.436 | - | 0.252 | −0.027 | - | 0.345 | 0.315 | - | 0.682 |
| Direct | −0.436 | - | 0.748 | −0.027 | - | 0.367 | 0.315 | - | 0.471 | |
| Indirect | 0.000 | - | −0.496 | 0.000 | - | −0.021 | 0.000 | - | 0.210 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Yang, Y.; Jiao, Z.; Chen, Z.; Shen, Y.; Liu, Y.; Zhang, L.; Wang, L.; Liu, S.; Wu, Q.; et al. Response of Soil Net Nitrogen Mineralization to a Litter in Three Subalpine Forests. Forests 2022, 13, 597. https://doi.org/10.3390/f13040597
Zhang L, Yang Y, Jiao Z, Chen Z, Shen Y, Liu Y, Zhang L, Wang L, Liu S, Wu Q, et al. Response of Soil Net Nitrogen Mineralization to a Litter in Three Subalpine Forests. Forests. 2022; 13(4):597. https://doi.org/10.3390/f13040597
Chicago/Turabian StyleZhang, Li, Yulian Yang, Zebin Jiao, Zihao Chen, Ya Shen, Yao Liu, Linhui Zhang, Lixia Wang, Sining Liu, Qinggui Wu, and et al. 2022. "Response of Soil Net Nitrogen Mineralization to a Litter in Three Subalpine Forests" Forests 13, no. 4: 597. https://doi.org/10.3390/f13040597
APA StyleZhang, L., Yang, Y., Jiao, Z., Chen, Z., Shen, Y., Liu, Y., Zhang, L., Wang, L., Liu, S., Wu, Q., & Li, H. (2022). Response of Soil Net Nitrogen Mineralization to a Litter in Three Subalpine Forests. Forests, 13(4), 597. https://doi.org/10.3390/f13040597