
The Known Unknowns: An Enigmatic Pathway of C17-Polyacetylenic Oxylipins in Carrot (Daucus carota L.)




Abstract
1. Introduction
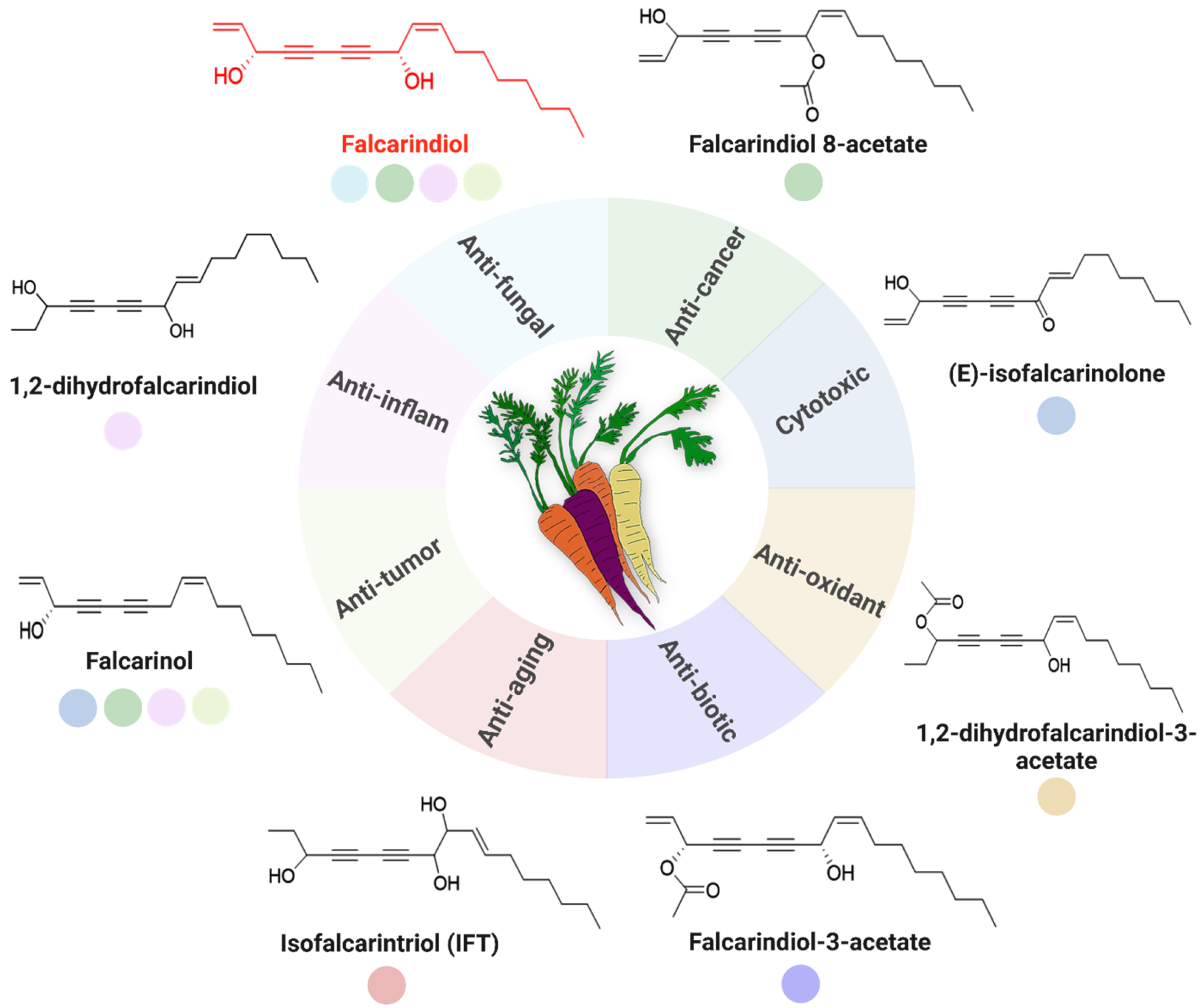
2. Chemical Nature and Biological Importance of C17-Polyacetylenes

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C17-Polyacetylene | Structural Features | Structural Genes | Transcription Factors | References |
|---|---|---|---|---|
| Falcarinol | One hydroxyl group at C3, two double bonds (C1/2, C9/10), two triple bonds (C4/5, C6/7) | DcFAD2-6 (Δ12 acetylenase), DcFAD2-11 (Δ12 desaturase), DcFAD2-13, DcFAD2-14 (bifunctional 12/Δ14 desaturases) | None specifically identified; potential MYB/bHLH involvement speculative | [2,20,28] |
| Falcarindiol | Hydroxyl groups at C3 and C8, two double bonds (C1/2, C9/10), two triple bonds (C4/5, C6/7) | DcFAD2-6, DcFAD2-11, DcFAD2-13, DcFAD2-14 | None specifically identified; potential MBW complex (MYB, bHLH, WD40) involvement speculative | [2,20,28] |
| Falcarindiol-3-acetate | Acetyl group at C3, hydroxyl at C8, two double bonds (C1/2, C9/10), two triple bonds (C4/5, C6/7) | DcFAD2-6, DcFAD2-11, DcFAD2-13, DcFAD2-14 | None specifically identified; potential MBW complex involvement speculative | [2,20,28] |
| (E)-Isofalcarinolone | Ketone group, two double bonds, two triple bonds | Likely derived from FAD2-mediated pathways; specific genes not confirmed | None specifically identified | [2,20,28] |
| Falcarindiol-8-acetate | Acetyl group at C8, hydroxyl at C3, two double bonds, two triple bonds | Likely derived from FAD2-mediated pathways; specific genes not confirmed | None specifically identified | [2,20,28] |
| 1,2-Dihydrofalcarindiol-3-acetate | Dihydro at C1/2, acetyl group at C3, hydroxyl at C8, one double bond, two triple bonds | Likely derived from FAD2-mediated pathways; specific genes not confirmed | None specifically identified | [2,20,28] |
| (E)-Falcarindiolone-8-acetate | Ketone group, acetyl at C8, two double bonds, two triple bonds | Likely derived from FAD2-mediated pathways; specific genes not confirmed | None specifically identified | [2,20,28] |
| (E)-Falcarindiolone-9-acetate | Ketone group, acetyl at C9, two double bonds, two triple bonds | Likely derived from FAD2-mediated pathways; specific genes not confirmed | None specifically identified | [2,20,28] |
| 1,2-Dihydrofalcarindiol | Dihydro at C1/2, hydroxyls at C3 and C8, one double bond, two triple bonds | Likely derived from FAD2-mediated pathways; specific genes not confirmed | None specifically identified | [2,20,28] |
| (E)-1-Methoxy-falcarindiolone-8-acetate | Methoxy group at C1, ketone, acetyl at C8, two double bonds, two triple bonds | Likely derived from FAD2-mediated pathways; specific genes not confirmed | None specifically identified | [2,20,28] |
| (E)-1-Methoxy-falcarindiolone-9-acetate | Methoxy group at C1, ketone, acetyl at C9, two double bonds, two triple bonds | Likely derived from FAD2-mediated pathways; specific genes not confirmed | None specifically identified | [2,20,28] |
| Panaxydiol | Two hydroxyl groups, two double bonds, two triple bonds | Likely derived from FAD2-mediated pathways; specific genes not confirmed | None specifically identified | [2,20,28] |
3. Bioactive Potential of C17-Polyacetylenes
3.1. Anticancer Activities of C17-Polyacetylenes
3.2. Anti-Inflammatory Properties of C17-Polyacetylenes
3.3. Antimicrobial and Antifungal Activities of C17-Polyacetylenes
3.4. Neuroprotective and Neuromodulatory Effects of C17-Polyacetylenes
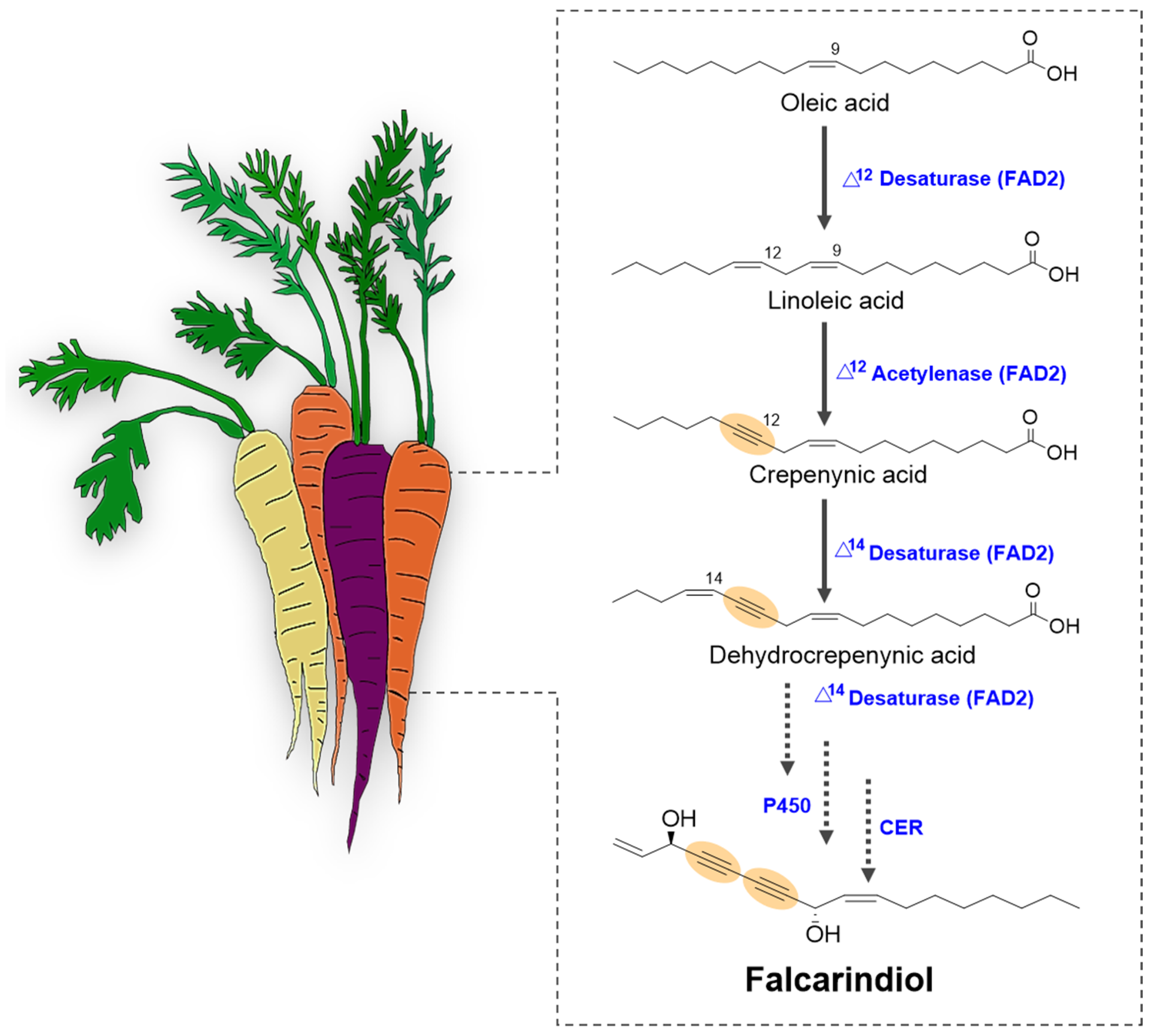
4. Current Understanding of the C17-Polyacetylene Biosynthetic Pathway in Carrot
4.1. Enzymatic Mechanisms
4.2. Genetic and Tissue-Specific Regulation
4.3. Environmental and Microbial Elicitaions
4.4. Knowledge Gaps in Carrot Polyacetylene Biosynthetic Pathway
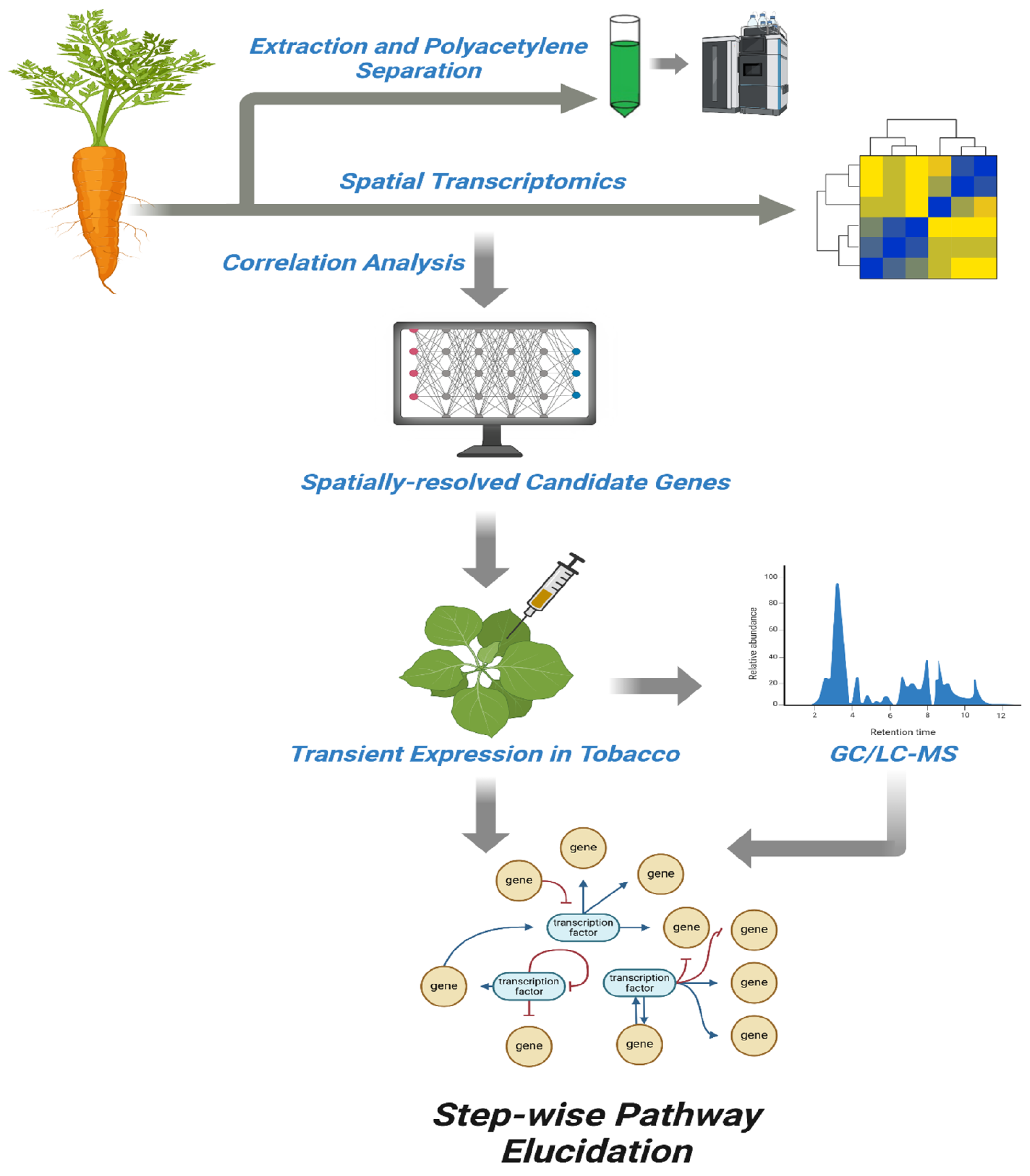
5. Methodological Challenges and Strategic Advances in Polyacetylene Pathway Elucidation
5.1. Methodological and Analytical Approaches
5.2. Comparative Insights from Other Apiaceae
5.3. Human Bioavailability and Pharmacokinetics
5.4. Next-Generation-Aided AI/ML Strategies
6. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Alfurayhi, R.; Huang, L.; Brandt, K. Pathways affected by falcarinol-type polyacetylenes and implications for their anti-inflammatory function and potential in cancer chemoprevention. Foods 2023, 12, 1192. [Google Scholar] [CrossRef] [PubMed]
- Dawid, C.; Dunemann, F.; Schwab, W.; Nothnagel, T.; Hofmann, T. Bioactive C17-polyacetylenes in carrots (Daucus carota L.): Current knowledge and future perspectives. J. Agric. Food Chem. 2015, 63, 9211–9222. [Google Scholar] [CrossRef]
- Christensen, L.P. Bioactive C17 and C18 acetylenic oxylipins from terrestrial plants as potential lead compounds for anticancer drug development. Molecules 2020, 25, 2568. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.; Cawood, M.; Iqbal, Q.; Ariño, A.; Batool, A.; Tariq, R.M.S.; Azam, M.; Akhtar, S. Phytochemicals in Daucus carota and their health benefits. Foods 2019, 8, 424. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, U.; Kobæk-Larsen, M.; Kjøller, K.D.; Antonsen, S.; Baatrup, G.; Trelle, M.B. Quantification of the anti-neoplastic polyacetylene falcarinol from carrots in human serum by LC-MS/MS. J. Chromatogr. B 2022, 1210, 123440. [Google Scholar] [CrossRef]
- Kobaek-Larsen, M.; Baatrup, G.K.; Notabi, M.; El-Houri, R.B.; Pipó-Ollé, E.; Christensen Arnspang, E.; Christensen, L.P. Dietary polyacetylenic oxylipins falcarinol and falcarindiol prevent inflammation and colorectal neoplastic transformation: A mechanistic and dose-response study in a rat model. Nutrients 2019, 11, 2223. [Google Scholar]
- Xie, Q.; Wang, C. Polyacetylenes in herbal medicine: A comprehensive review of its occurrence, pharmacology, toxicology, and pharmacokinetics (2014–2021). Phytochemistry 2022, 201, 113288. [Google Scholar] [CrossRef]
- Guaâdaoui, A. Recent Advances in Bioactivities of Common Food Biocompounactives. In Fruit and Vegetable Phytochemicals: Chemistry and Human Health, 2nd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; pp. 541–594. [Google Scholar]
- Kumar, S.; Saini, R.; Suthar, P.; Kumar, V.; Sharma, R. Plant secondary metabolites: Their food and therapeutic importance. In Plant Secondary Metabolites: Physico-Chemical Properties and Therapeutic Applications; Springer: Berlin/Heidelberg, Germany, 2022; pp. 371–413. [Google Scholar]
- Bouvet, J.; Maraval, V.; Ballereau, S.; Bernardes-Génisson, V.; Génisson, Y. Natural and Bioinspired Lipidic Alkynylcarbinols as Leishmanicidal, Antiplasmodial, Trypanocidal, Fungicidal, Antibacterial, and Antimycobacterial Agents. J. Nat. Prod. 2024, 87, 2550–2566. [Google Scholar] [CrossRef]
- He, W. Uncovering the Role Of Falcarin-Type Polyacetylenes in Carrots for Enhanced Resistance; Georg-August-Universität Göttingen: Göttingen, Germany, 2024. [Google Scholar]
- Savchenko, T.; Degtyaryov, E.; Radzyukevich, Y.; Buryak, V. Therapeutic potential of plant oxylipins. Int. J. Mol. Sci. 2022, 23, 14627. [Google Scholar] [CrossRef]
- Santos, P.; Busta, L.; Yim, W.C.; Cahoon, E.B.; Kosma, D.K. Structural diversity, biosynthesis, and function of plant falcarin-type polyacetylenic lipids. J. Exp. Bot. 2022, 73, 2889–2904. [Google Scholar] [CrossRef]
- Crozier, A.; Yokota, T.; Jaganath, I.B.; Marks, S.; Saltmarsh, M.; Clifford, M.N. Secondary metabolites in fruits, vegetables, beverages and other plant based dietary components. In Plant Secondary Metabolites: Occurrence, Structure and Role in the Human Diet; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 208–302. [Google Scholar]
- P Christensen, L. Aliphatic C17-polyacetylenes of the falcarinol type as potential health promoting compounds in food plants of the Apiaceae family. Recent Pat. Food Nutr. Agric. 2011, 3, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Erni, R.; Wu, J.Y.; Fischer, F.; Lamers, G.; Grigolon, G.; Mitchell, S.J.; Zarse, K.; Carreira, E.M.; Ristow, M. A naturally occurring polyacetylene isolated from carrots promotes health and delays signatures of aging. Nat. Commun. 2023, 14, 8142. [Google Scholar] [CrossRef] [PubMed]
- Minto, R.E.; Blacklock, B.J. Biosynthesis and function of polyacetylenes and allied natural products. Prog. Lipid Res. 2008, 47, 233–306. [Google Scholar] [CrossRef]
- Feng, T.; Yang, Y.; Busta, L.; Cahoon, E.B.; Wang, H.; Lü, S. FAD2 gene radiation and positive selection contributed to polyacetylene metabolism evolution in Campanulids. Plant Physiol. 2019, 181, 714–728. [Google Scholar] [CrossRef]
- Schmiech, L.; Alayrac, C.; Witulski, B.; Hofmann, T. Structure determination of bisacetylenic oxylipins in carrots (Daucus carota L.) and enantioselective synthesis of falcarindiol. J. Agric. Food Chem. 2009, 57, 11030–11040. [Google Scholar] [CrossRef]
- Busta, L.; Yim, W.C.; LaBrant, E.W.; Wang, P.; Grimes, L.; Malyszka, K.; Cushman, J.C.; Santos, P.; Kosma, D.K.; Cahoon, E.B. Identification of genes encoding enzymes catalyzing the early steps of carrot polyacetylene biosynthesis. Plant Physiol. 2018, 178, 1507–1521. [Google Scholar] [CrossRef]
- Zidorn, C.; Jöhrer, K.; Ganzera, M.; Schubert, B.; Sigmund, E.M.; Mader, J.; Greil, R.; Ellmerer, E.P.; Stuppner, H. Polyacetylenes from the Apiaceae vegetables carrot, celery, fennel, parsley, and parsnip and their cytotoxic activities. J. Agric. Food Chem. 2005, 53, 2518–2523. [Google Scholar] [CrossRef] [PubMed]
- Christensen, L.P.; Brandt, K. Bioactive polyacetylenes in food plants of the Apiaceae family: Occurrence, bioactivity and analysis. J. Pharm. Biomed. Anal. 2006, 41, 683–693. [Google Scholar] [CrossRef]
- Bentley, R.; Bhattacharjee, D.; Jones, E.; Thaller, V. C17-Polyacetylenic alcohols from the umbellifer Daucus carota (carrot): Alkylation of benzene by acetylenyl (vinyl) carbinols in the presence of p-toluenesulfonic acid. J. Chem. Soc. C 1969, 685–688. [Google Scholar] [CrossRef]
- Christensen, L.P.; Kreutzmann, S. Determination of polyacetylenes in carrot roots (Daucus carota L.) by high-performance liquid chromatography coupled with diode array detection. J. Sep. Sci. 2007, 30, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Czepa, A.; Hofmann, T. Quantitative studies and sensory analyses on the influence of cultivar, spatial tissue distribution, and industrial processing on the bitter off-taste of carrots (Daucus carota L.) and carrot products. J. Agric. Food Chem. 2004, 52, 4508–4514. [Google Scholar] [CrossRef] [PubMed]
- Kidmose, U.; Hansen, S.L.; Christensen, L.P.; Edelenbos, M.; Larsen, E.; Nørbæk, R. Effects of genotype, root size, storage, and processing on bioactive compounds in organically grown carrots (Daucus carota L.). J. Food Sci. 2004, 69, S388–S394. [Google Scholar] [CrossRef]
- Pferschy-Wenzig, E.-M.; Getzinger, V.; Kunert, O.; Woelkart, K.; Zahrl, J.; Bauer, R. Determination of falcarinol in carrot (Daucus carota L.) genotypes using liquid chromatography/mass spectrometry. Food Chem. 2009, 114, 1083–1090. [Google Scholar] [CrossRef]
- Czepa, A.; Hofmann, T. Structural and sensory characterization of compounds contributing to the bitter off-taste of carrots (Daucus carota L.) and carrot puree. J. Agric. Food Chem. 2003, 51, 3865–3873. [Google Scholar] [CrossRef]
- Baranska, M.; Schulz, H.; Baranski, R.; Nothnagel, T.; Christensen, L.P. In situ simultaneous analysis of polyacetylenes, carotenoids and polysaccharides in carrot roots. J. Agric. Food Chem. 2005, 53, 6565–6571. [Google Scholar] [CrossRef]
- Lecomte, M.; Berruyer, R.; Hamama, L.; Boedo, C.; Hudhomme, P.; Bersihand, S.; Arul, J.; N’guyen, G.; Gatto, J.; Guilet, D. Inhibitory effects of the carrot metabolites 6-methoxymellein and falcarindiol on development of the fungal leaf blight pathogen Alternaria dauci. Physiol. Mol. Plant Pathol. 2012, 80, 58–67. [Google Scholar] [CrossRef]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef]
- Tan, K.W.; Killeen, D.P.; Li, Y.; Paxton, J.W.; Birch, N.P.; Scheepens, A. Dietary polyacetylenes of the falcarinol type are inhibitors of breast cancer resistance protein (BCRP/ABCG2). Eur. J. Pharmacol. 2014, 723, 346–352. [Google Scholar] [CrossRef]
- Warner, S.R. Carrots and Cancer: The Bioavailability of Polyacetylene from Carrots and Their Assocation with Biomarkers of Cancer Risk. Ph.D. Thesis, Newcastle University, Newcastle upon Tyne, UK, 2019. [Google Scholar]
- Lu, T.; Gu, M.; Zhao, Y.; Zheng, X.; Xing, C. Autophagy contributes to falcarindiol-induced cell death in breast cancer cells with enhanced endoplasmic reticulum stress. PLoS ONE 2017, 12, e0176348. [Google Scholar] [CrossRef]
- Tsolmon, S.; Kurita, Y.; Yamada, P.; Shigemori, H.; Isoda, H. Indoleacetic acid falcarindiol ester induces granulocytic differentiation of the human leukemia cell line HL-60. Planta Medica 2009, 75, 49–54. [Google Scholar] [CrossRef]
- Yamazoe, S.; Hasegawa, K.; Shigemori, H. Growth inhibitory indole acetic acid polyacetylenic ester from Japanese ivy (Hedera rhombea Bean). Phytochemistry 2007, 68, 1706–1711. [Google Scholar] [CrossRef]
- Bae, K.-E.; Choi, Y.-W.; Kim, S.-T.; Kim, Y.-K. Components of rhizome extract of Cnidium officinale Makino and their in vitro biological effects. Molecules 2011, 16, 8833–8847. [Google Scholar] [CrossRef]
- De la Cruz, J.; Kim, D.-H.; Hwang, S.G. Anti cancer effects of Cnidium officinale Makino extract mediated through apoptosis and cell cycle arrest in the HT-29 human colorectal cancer cell line. Asian Pac. J. Cancer Prev. 2014, 15, 5117–5121. [Google Scholar] [CrossRef] [PubMed]
- Stefanson, A.L.; Bakovic, M. Falcarinol Is a Potent Inducer of Heme Oxygenase-1 and Was More Effective than Sulforaphane in Attenuating Intestinal Inflammation at Diet-Achievable Doses. Oxidative Med. Cell. Longev. 2018, 2018, 3153527. [Google Scholar] [CrossRef] [PubMed]
- Ohnuma, T.; Nakayama, S.; Anan, E.; Nishiyama, T.; Ogura, K.; Hiratsuka, A. Activation of the Nrf2/ARE pathway via S-alkylation of cysteine 151 in the chemopreventive agent-sensor Keap1 protein by falcarindiol, a conjugated diacetylene compound. Toxicol. Appl. Pharmacol. 2010, 244, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and-independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef]
- Ohnuma, T.; Komatsu, T.; Nakayama, S.; Nishiyama, T.; Ogura, K.; Hiratsuka, A. Induction of antioxidant and phase 2 drug-metabolizing enzymes by falcarindiol isolated from Notopterygium incisum extract, which activates the Nrf2/ARE pathway, leads to cytoprotection against oxidative and electrophilic stress. Arch. Biochem. Biophys. 2009, 488, 34–41. [Google Scholar] [CrossRef]
- Kong, C.-S.; Um, Y.R.; Im Lee, J.; Kim, Y.A.; Yea, S.S.; Seo, Y. Constituents isolated from Glehnia littoralis suppress proliferations of human cancer cells and MMP expression in HT1080 cells. Food Chem. 2010, 120, 385–394. [Google Scholar] [CrossRef]
- Cai, J.; Shi, S.-Y.; Cheng, F.; Wei, M.; Zou, K.; Yu, X.-Q.; Chen, J.-F. Polyacetylene isomers isolated from Bidens pilosa L. suppress the metastasis of gastric Cancer cells by inhibiting Wnt/β-Catenin and Hippo/YAP signaling pathways. Molecules 2023, 28, 1837. [Google Scholar] [CrossRef]
- Resetar, M.; Liu, X.; Herdlinger, S.; Kunert, O.; Pferschy-Wenzig, E.-M.; Latkolik, S.; Steinacher, T.; Schuster, D.; Bauer, R.; Dirsch, V.M. Polyacetylenes from Oplopanax horridus and Panax ginseng: Relationship between structure and PPARγ activation. J. Nat. Prod. 2020, 83, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Kobaek-Larsen, M.; El-Houri, R.B.; Christensen, L.P.; Al-Najami, I.; Fretté, X.; Baatrup, G. Dietary polyacetylenes, falcarinol and falcarindiol, isolated from carrots prevents the formation of neoplastic lesions in the colon of azoxymethane-induced rats. Food Funct. 2017, 8, 964–974. [Google Scholar] [CrossRef]
- Deding, U.; Baatrup, G.; Christensen, L.P.; Kobaek-Larsen, M. Carrot intake and risk of colorectal cancer: A prospective cohort study of 57,053 Danes. Nutrients 2020, 12, 332. [Google Scholar] [CrossRef]
- Venkatesan, T.; Choi, Y.-W.; Lee, J.; Kim, Y.-K. Falcarindiol inhibits LPS-induced inflammation via attenuating MAPK and JAK-STAT signaling pathways in murine macrophage RAW 264.7 cells. Mol. Cell. Biochem. 2018, 445, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Metzger, B.T.; Barnes, D.M.; Reed, J.D. Purple carrot (Daucus carota L.) polyacetylenes decrease lipopolysaccharide-induced expression of inflammatory proteins in macrophage and endothelial cells. J. Agric. Food Chem. 2008, 56, 3554–3560. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Ju, J.; Song, J.-L.; Yang, S.-G.; Park, K.-Y. Anti-colitic effect of purple carrot on dextran sulfate sodium (DSS)-induced colitis in C57BL/6J Mice. Prev. Nutr. Food Sci. 2018, 23, 77. [Google Scholar] [CrossRef]
- Kobaek-Larsen, M.; Deding, U.; Al-Najami, I.; Clausen, B.H.; Christensen, L.P. Carrot Juice Intake Affects the Cytokine and Chemokine Response in Human Blood after Ex Vivo Lipopolysaccharide-Induced Inflammation. Nutrients 2023, 15, 5002. [Google Scholar] [CrossRef]
- Stefanson, A. Anti-Inflammatory and Immunomodulatory Effects of Carrot-Derived Polyacetylenes in Acute Intestinal and Systemic Inflammation and an Evaluation of Potential of Agronomic Techniques for Their Enhancement in Carrot. Ph.D. Thesis, University of Guelph, Guelph, ON, Canada, 2018. [Google Scholar]
- Stefanson, A.L.; Bakovic, M. Dietary regulation of Keap1/Nrf2/ARE pathway: Focus on plant-derived compounds and trace minerals. Nutrients 2014, 6, 3777–3801. [Google Scholar] [CrossRef]
- Cavagnaro, P.F.; Dunemann, F.; Selvakumar, R.; Iorizzo, M.; Simon, P.W. Health-enhancing compounds in carrots: Genetics, genomics, and molecular breeding. In Compendium of Crop Genome Designing for Nutraceuticals; Springer: Singapore, 2023; pp. 1–72. [Google Scholar]
- Dolatpanah, M.; Rashtchizadeh, N.; Abbasi, M.M.; Nazari, S.; Mohammadian, J.; Roshangar, L.; Argani, H.; Ghorbanihaghjo, A. Falcarindiol attenuates cisplatin-induced nephrotoxicity through the modulation of NF-kB and Nrf2 signaling pathways in mice. preprint. 2022. [Google Scholar]
- Liu, R.; Choi, H.S.; Kim, S.-L.; Kim, J.-H.; Yun, B.-S.; Lee, D.-S. 6-Methoxymellein isolated from carrot (Daucus carota L.) targets breast cancer stem cells by regulating NF-κB signaling. Molecules 2020, 25, 4374. [Google Scholar] [CrossRef]
- Singh, R.; Tiwari, P.; Sharma, B.; Guerrero-Perilla, C.; Coy-Barrera, E. Analysis of polyacetylenes. In Recent Advances in Natural Products Analysis; Elsevier: Amsterdam, The Netherlands, 2020; pp. 707–722. [Google Scholar]
- Nazar, M.S.; Saeed, M.; Khan, V.; ul Haq, I.; Tariq, M.A.; Tanoli, A.H.; Zakki, S.A. Nutritional and therapeutic importance of carrot pomace: A review article. Int. J. Nat. Med. Health Sci. 2023, 2, 25–33. [Google Scholar]
- Jeon, J.E.; Kim, J.-G.; Fischer, C.R.; Mehta, N.; Dufour-Schroif, C.; Wemmer, K.; Mudgett, M.B.; Sattely, E. A pathogen-responsive gene cluster for highly modified fatty acids in tomato. Cell 2020, 180, 176–187.e119. [Google Scholar] [CrossRef] [PubMed]
- Meot-Duros, L.; Cérantola, S.; Talarmin, H.; Le Meur, C.; Le Floch, G.; Magné, C. New antibacterial and cytotoxic activities of falcarindiol isolated in Crithmum maritimum L. leaf extract. Food Chem. Toxicol. 2010, 48, 553–557. [Google Scholar] [CrossRef] [PubMed]
- Arnspang, E.C.; Pipó-Ollé, E.; Notabi, M.K.; Hedegaard, M.A.; Walther, A.; Kobaek-Larsen, M.; Baatrup, G.; El-Houri, R.B.; Christensen, L.P. Effect of falcarindiol and falcarinol purified from carrots on inflammation and cancer. In Proceedings of the Nordic Natural Product Conference (NNPC) 2019, Flúðir, Iceland, 9 June–12 June 2019; p. L26. [Google Scholar]
- Hinds, L.; Kenny, O.; Hossain, M.; Walsh, D.; Sheehy, E.; Evans, P.; Gaffney, M.; Rai, D. Evaluating the antibacterial properties of polyacetylene and glucosinolate compounds with further identification of their presence within various carrot (Daucus carota) and Broccoli (Brassica oleracea) cultivars using high-performance liquid chromatography with a diode array detector and ultra performance liquid chromatography–tandem mass spectrometry analyses. J. Agric. Food Chem. 2017, 65, 7186–7191. [Google Scholar] [PubMed]
- Patra, A.K. An overview of antimicrobial properties of different classes of phytochemicals. Diet. Phytochem. Microbes 2012, 1–32. [Google Scholar] [CrossRef]
- Negri, R. Polyacetylenes from terrestrial plants and fungi: Recent phytochemical and biological advances. Fitoterapia 2015, 106, 92–109. [Google Scholar] [CrossRef]
- Ayoub, N.; Al-Azizi, M.; König, W.; Kubeczka, K.H. Essential oils and a novel polyacetylene from Eryngium yuccifolium Michaux.(Apiaceae). Flavour Fragr. J. 2006, 21, 864–868. [Google Scholar] [CrossRef]
- Garrod, B.; Lea, E.; Lewis, B. Studies on the mechanism of action of the antifungal compound falcarindiol. New Phytol. 1979, 83, 463–471. [Google Scholar] [CrossRef]
- Shiono, Y.; Haga, M.; Koyama, H.; Murayama, T.; Koseki, T. Antifungal activity of a polyacetylene against the fungal pathogen of Japanese oak from the liquid culture of the edible mushroom, Hypsizygus marmoreus. Z. Für Naturforschung B 2013, 68, 293–295. [Google Scholar] [CrossRef]
- Rahman, M.A.A.; Cho, S.-C.; Song, J.; Mun, H.-T.; Moon, S.-S. Dendrazawaynes A and B, antifungal polyacetylenes from Dendranthema zawadskii (Asteraceae). Planta Medica 2007, 73, 1089–1094. [Google Scholar] [CrossRef]
- Zhao, L.; Li, C.; Zhang, M.; Li, H.; Ding, S.; Zhao, X.; Yang, L.; Liu, J.; Li, Q.X.; Na, R. Design, Synthesis, and Antifungal Activity of Polyacetylenic Alcohol Derivatives and Stereoisomers against Phytopathogenic Fungi. J. Agric. Food Chem. 2023, 71, 9753–9761. [Google Scholar] [CrossRef]
- Kobaek-Larsen, M.; Nielsen, D.S.; Kot, W.; Krych, Ł.; Christensen, L.P.; Baatrup, G. Effect of the dietary polyacetylenes falcarinol and falcarindiol on the gut microbiota composition in a rat model of colorectal cancer. BMC Res. Notes 2018, 11, 411. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lee, P.; Son, D.; Kim, H.; Kim, S.Y. Falcarindiol inhibits nitric oxide-mediated neuronal death in lipopolysaccharide-treated organotypic hippocampal cultures. Neuroreport 2003, 14, 1941–1944. [Google Scholar] [CrossRef] [PubMed]
- Kim, H. Neuroprotective herbs for stroke therapy in traditional eastern medicine. Neurol. Res. 2005, 27, 287–301. [Google Scholar] [CrossRef]
- Herrmann, F.; Hamoud, R.; Sporer, F.; Tahrani, A.; Wink, M. Carlina oxide–a natural polyacetylene from Carlina acaulis (Asteraceae) with potent antitrypanosomal and antimicrobial properties. Planta Medica 2011, 77, 1905–1911. [Google Scholar] [CrossRef]
- Takagi, M.; Kimura, K.; Nakashima, K.-I.; Inoue, M. Ameliorative effect of panaxynol on the reduction in high-molecular-weight adiponectin secretion from 3T3-L1 adipocytes treated with palmitic acids. Eur. J. Pharmacol. 2018, 820, 138–145. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, T.; Zhao, Y.; Cai, E.; Zhu, H.; Liu, S. Panaxynol attenuates CUMS-induced anxiety and depressive-like behaviors via regulating neurotransmitters, synapses and the HPA axis in mice. Food Funct. 2020, 11, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kadowaki, T. Physiological and pathophysiological roles of adiponectin and adiponectin receptors in the integrated regulation of metabolic and cardiovascular diseases. Int. J. Obes. 2008, 32, S13–S18. [Google Scholar] [CrossRef]
- Gonçalves, E.C.; Baldasso, G.M.; Bicca, M.A.; Paes, R.S.; Capasso, R.; Dutra, R.C. Terpenoids, cannabimimetic ligands, beyond the cannabis plant. Molecules 2020, 25, 1567. [Google Scholar] [CrossRef]
- McPartland, J.M.; Guy, G.W.; Di Marzo, V. Care and feeding of the endocannabinoid system: A systematic review of potential clinical interventions that upregulate the endocannabinoid system. PLoS ONE 2014, 9, e89566. [Google Scholar] [CrossRef]
- Cao, L.; Huang, X.; Zhu, J.; Xiao, J.; Xie, L. Falcarindiol improves functional recovery and alleviates neuroinflammation after spinal cord injury by inhibiting STAT/MAPK signaling pathways. Biochem. Biophys. Res. Commun. 2024, 736, 150860. [Google Scholar] [CrossRef]
- Salimonti, A.; Carbone, F.; Romano, E.; Pellegrino, M.; Benincasa, C.; Micali, S.; Tondelli, A.; Conforti, F.L.; Perri, E.; Ienco, A. Association study of the 5’ UTR Intron of the FAD2-2 gene with oleic and linoleic acid content in Olea europaea L. Front. Plant Sci. 2020, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Launhardt, L.; Uhlenberg, J.; Stellmach, H.; Schomburg, M.; Hause, B.; Heilmann, I.; Heilmann, M. Association of the Arabidopsis oleoyl Δ12-desaturase FAD2 with pre-cis-Golgi stacks at endoplasmic reticulum-Golgi-exit sites. Plant J. 2024, 117, 242–263. [Google Scholar] [CrossRef] [PubMed]
- Dunemann, F.; He, W.; Böttcher, C.; Reichardt, S.; Nothnagel, T.; Heuvelmans, P.; Hermans, F. The genetic control of polyacetylenes involved in bitterness of carrots (Daucus carota L.): Identification of QTLs and candidate genes from the plant fatty acid metabolism. BMC Plant Biol. 2022, 22, 92. [Google Scholar] [CrossRef]
- Kjellenberg, L.; Johansson, E.; Gustavsson, K.E.; Olsson, M.E. Polyacetylenes in fresh and stored carrots (Daucus carota): Relations to root morphology and sugar content. J. Sci. Food Agric. 2012, 92, 1748–1754. [Google Scholar] [CrossRef]
- He, W.; Dunemann, F. Endogenous falcarinol-type polyacetylenes in carrots and their putative influence on post-harvest fungal pathogens Mycocentrospora acerina and Botrytis cinerea. bioRxiv 2025. [Google Scholar] [CrossRef]
- Chung, H.-H.; Ting, H.-M.; Wang, W.-H.; Chao, Y.-T.; Hsieh, C.-H.; Apaya, M.K.; Sung, Y.-C.; Lin, S.-S.; Hwu, F.-Y.; Shyur, L.-F. Elucidation of enzymes involved in the biosynthetic pathway of bioactive polyacetylenes in Bidens pilosa using integrated omics approaches. J. Exp. Bot. 2021, 72, 525–541. [Google Scholar] [CrossRef]
- Iorizzo, M.; Ellison, S.; Senalik, D.; Zeng, P.; Satapoomin, P.; Huang, J.; Bowman, M.; Iovene, M.; Sanseverino, W.; Cavagnaro, P. A high-quality carrot genome assembly provides new insights into carotenoid accumulation and asterid genome evolution. Nat. Genet. 2016, 48, 657–666. [Google Scholar] [CrossRef]
- Cahoon, E.B.; Schnurr, J.A.; Huffman, E.A.; Minto, R.E. Fungal responsive fatty acid acetylenases occur widely in evolutionarily distant plant families. Plant J. 2003, 34, 671–683. [Google Scholar] [CrossRef]
- Kjellenberg, L.; Johansson, E.; Gustavsson, K.-E.; Granstedt, A.; Olsson, M.E. Correlations between polyacetylene concentrations in carrot (Daucus carota L.) and various soil parameters. Foods 2016, 5, 60. [Google Scholar] [CrossRef]
- Chen, W.; He, P.; Zhang, H.; Lü, F. Effects of volatile fatty acids on soil properties, microbial communities, and volatile metabolites in wheat rhizosphere of loess. J. Clean. Prod. 2024, 476, 143798. [Google Scholar] [CrossRef]
- Knispel, N.; Ostrozhenkova, E.; Schramek, N.; Huber, C.; Peña-Rodríguez, L.M.; Bonfill, M.; Palazón, J.; Wischmann, G.; Cusidó, R.M.; Eisenreich, W. Biosynthesis of panaxynol and panaxydol in Panax ginseng. Molecules 2013, 18, 7686–7698. [Google Scholar] [CrossRef]
- Rawson, A.; Brunton, N.P.; Rai, D.K.; McLoughlin, P.; Tiwari, B.K.; Tuohy, M.G. Stability of falcarinol type polyacetylenes during processing of Apiaceae vegetables. Trends Food Sci. Technol. 2013, 30, 133–141. [Google Scholar] [CrossRef]
- Farag, M.A.; Sheashea, M.; Zhao, C.; Maamoun, A.A. UV fingerprinting approaches for quality control analyses of food and functional food coupled to chemometrics: A comprehensive analysis of novel trends and applications. Foods 2022, 11, 2867. [Google Scholar] [CrossRef] [PubMed]
- Atanassova, M.; Zahnit, W.; Messaoudi, M.; Rebiai, A.; Benchikha, N.; Sawicka, B. Analytical Techniques for Phytochemical Analysis and Functional Investigation. In Biotechnology and Phytochemical Prospects in Drug Discovery; Springer: Berlin/Heidelberg, Germany, 2025; pp. 151–187. [Google Scholar]
- Cavagnaro, P.F. Genetics and genomics of carrot sugars and polyacetylenes. In The Carrot Genome; Springer: Cham, Switzerland, 2019; pp. 295–315. [Google Scholar]
- Klimek-Chodacka, M.; Lukasiewicz, A.; Szymonik, K.; Baranski, R. Carrot Genetic Engineering. In Genetic Engineering of Vegetable Crops; CABI GB: Oxfordshire, UK, 2024; pp. 91–119. [Google Scholar]
- Lee, M.W.; Padilla, C.S.; Gupta, C.; Galla, A.; Pereira, A.; Li, J.; Goggin, F.L. The FATTY ACID DESATURASE2 family in tomato contributes to primary metabolism and stress responses. Plant Physiol. 2020, 182, 1083–1099. [Google Scholar] [CrossRef]
- Manohar, S.; Kumar, S.; Krishnan, V.; Maheshwari, C.; Dhaka, A.S.; Narwal, S.; Nair, L.S.; Vinutha, T.; Bansal, N.; Sureja, A. Insights into the Genetic Improvement of Carrot (Daucus carota L.). In Genetic Engineering of Crop Plants for Food and Health Security: Volume 2; Springer: Berlin/Heidelberg, Germany, 2024; pp. 185–203. [Google Scholar]
- Vishnu, K.; Aiswarya, K.; Sumesh, K. Recent advances in sample preparation in chromatographic techniques. Adv. Sep. Sci. 2025, 87–105. [Google Scholar]
- Killeen, D.P.; Sansom, C.E.; Lill, R.E.; Eason, J.R.; Gordon, K.C.; Perry, N.B. Quantitative Raman spectroscopy for the analysis of carrot bioactives. J. Agric. Food Chem. 2013, 61, 2701–2708. [Google Scholar] [CrossRef]
- Wang, N.; Huo, Y.-X. Using genome and transcriptome analysis to elucidate biosynthetic pathways. Curr. Opin. Biotechnol. 2022, 75, 102708. [Google Scholar] [CrossRef]
- Petersen, B.L.; Möller, S.R.; Mravec, J.; Jørgensen, B.; Christensen, M.; Liu, Y.; Wandall, H.H.; Bennett, E.P.; Yang, Z. Improved CRISPR/Cas9 gene editing by fluorescence activated cell sorting of green fluorescence protein tagged protoplasts. BMC Biotechnol. 2019, 19, 36. [Google Scholar] [CrossRef]
- Reyna-Llorens, I.; Ferro-Costa, M.; Burgess, S.J. Plant protoplasts in the age of synthetic biology. J. Exp. Bot. 2023, 74, 3821–3832. [Google Scholar] [CrossRef]
- Wolfender, J.-L.; Marti, G.; Ferreira Queiroz, E. Advances in techniques for profiling crude extracts and for the rapid identificationof natural products: Dereplication, quality control and metabolomics. Curr. Org. Chem. 2010, 14, 1808–1832. [Google Scholar] [CrossRef]
- Mukherjee, D.; Mukherjee, A. Understanding Plant Secondary Metabolism Using Bioinformatics Tools: Recent Advances and Prospects. In Bioinformatics for Plant Research and Crop Breeding; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2024; pp. 459–488. [Google Scholar]
- Kim, G.B.; Kim, W.J.; Kim, H.U.; Lee, S.Y. Machine learning applications in systems metabolic engineering. Curr. Opin. Biotechnol. 2020, 64, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Durand, M.; Besseau, S.; Papon, N.; Courdavault, V. Unlocking plant bioactive pathways: Omics data harnessing and machine learning assisting. Curr. Opin. Biotechnol. 2024, 87, 103135. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.S.; Lee, B.T.; Lee, S.Y.; Kim, H.U. Modeling regulatory networks using machine learning for systems metabolic engineering. Curr. Opin. Biotechnol. 2020, 65, 163–170. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Umar, A.W.; Hussain, H.; Ahmad, N. The Known Unknowns: An Enigmatic Pathway of C17-Polyacetylenic Oxylipins in Carrot (Daucus carota L.). Curr. Issues Mol. Biol. 2025, 47, 471. https://doi.org/10.3390/cimb47060471
Umar AW, Hussain H, Ahmad N. The Known Unknowns: An Enigmatic Pathway of C17-Polyacetylenic Oxylipins in Carrot (Daucus carota L.). Current Issues in Molecular Biology. 2025; 47(6):471. https://doi.org/10.3390/cimb47060471
Chicago/Turabian StyleUmar, Abdul Wakeel, Hamad Hussain, and Naveed Ahmad. 2025. "The Known Unknowns: An Enigmatic Pathway of C17-Polyacetylenic Oxylipins in Carrot (Daucus carota L.)" Current Issues in Molecular Biology 47, no. 6: 471. https://doi.org/10.3390/cimb47060471
APA StyleUmar, A. W., Hussain, H., & Ahmad, N. (2025). The Known Unknowns: An Enigmatic Pathway of C17-Polyacetylenic Oxylipins in Carrot (Daucus carota L.). Current Issues in Molecular Biology, 47(6), 471. https://doi.org/10.3390/cimb47060471