Meloside A Protects Dermal Papilla Cells from DHT-Induced Damage via Androgen Receptor Modulation

 , ,
, ,
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. CLE Preparation
2.2. Reagents and Plasmids
2.3. Culture of Human Dermal Papilla Cells (HDPCs)
2.4. pEGFP-C1-AR Subcellular Localization
2.5. Enzyme-Linked Immunosorbent Assay (ELISA)
2.6. Western Blot Analysis
2.7. HPLC–ESI–MS Analysis
2.8. Statistical Analysis
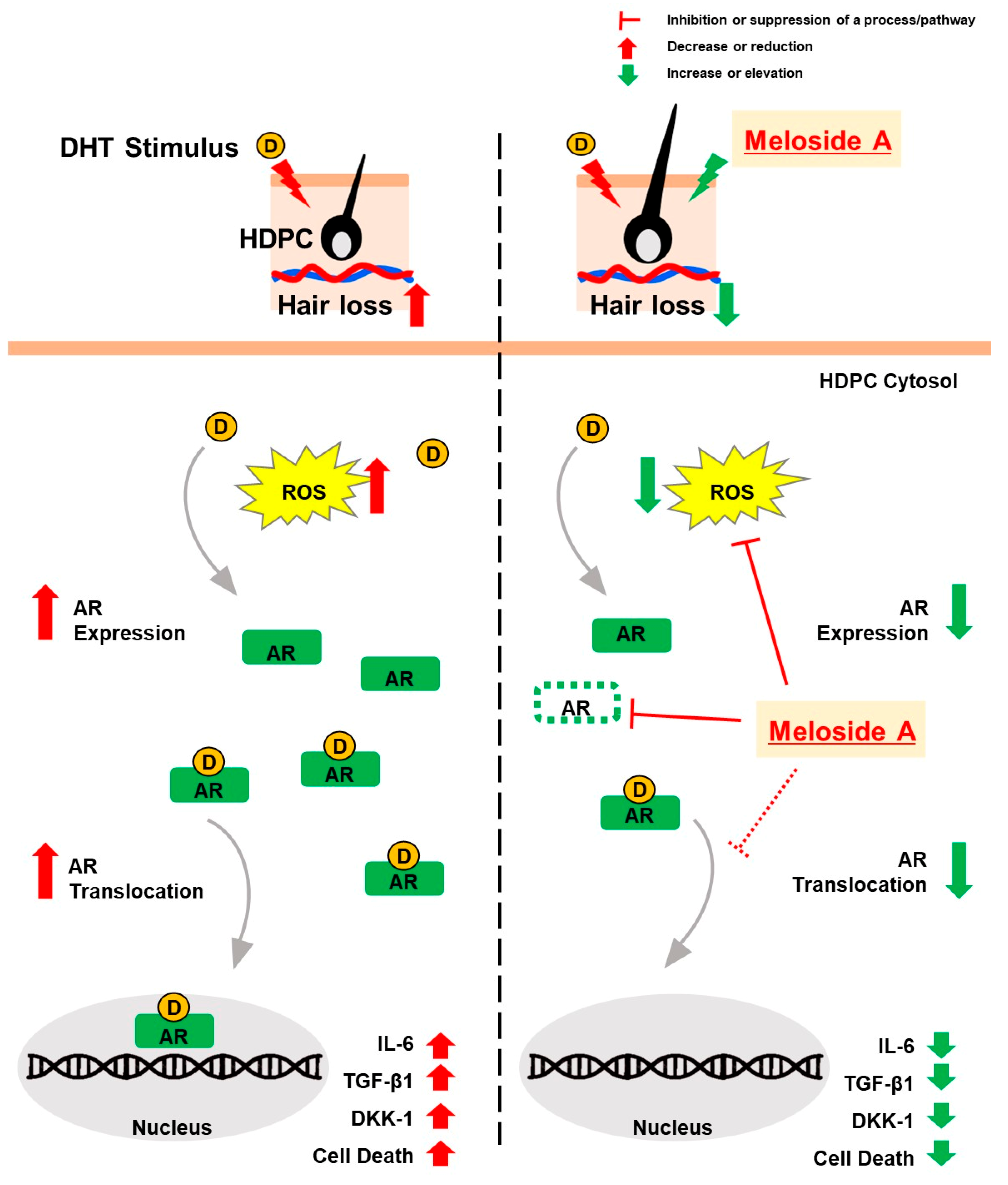
3. Results
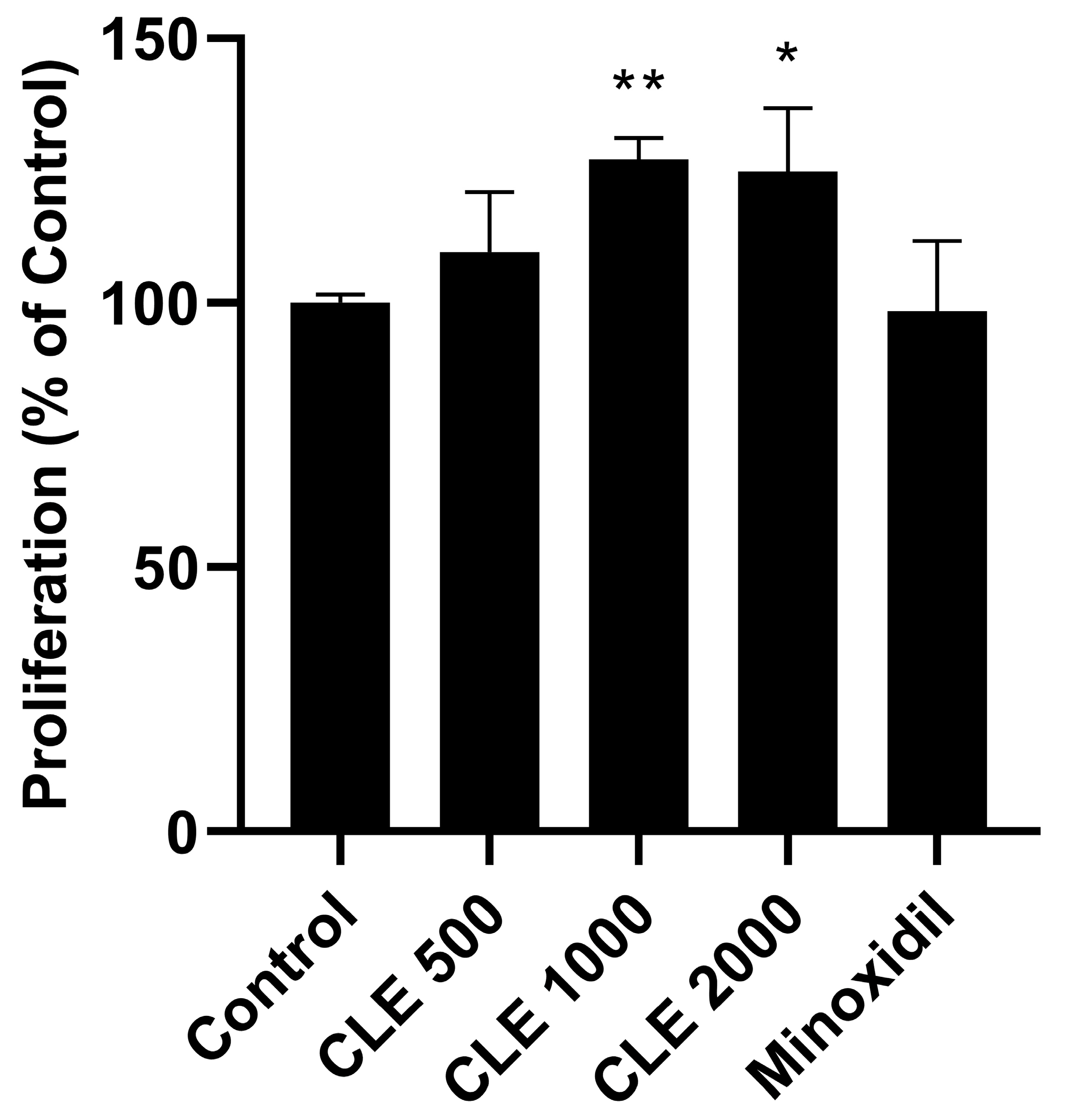
3.1. Evaluation of the Proliferative Capacity in Human Follicle Dermal Papilla Cells (HDPCs)
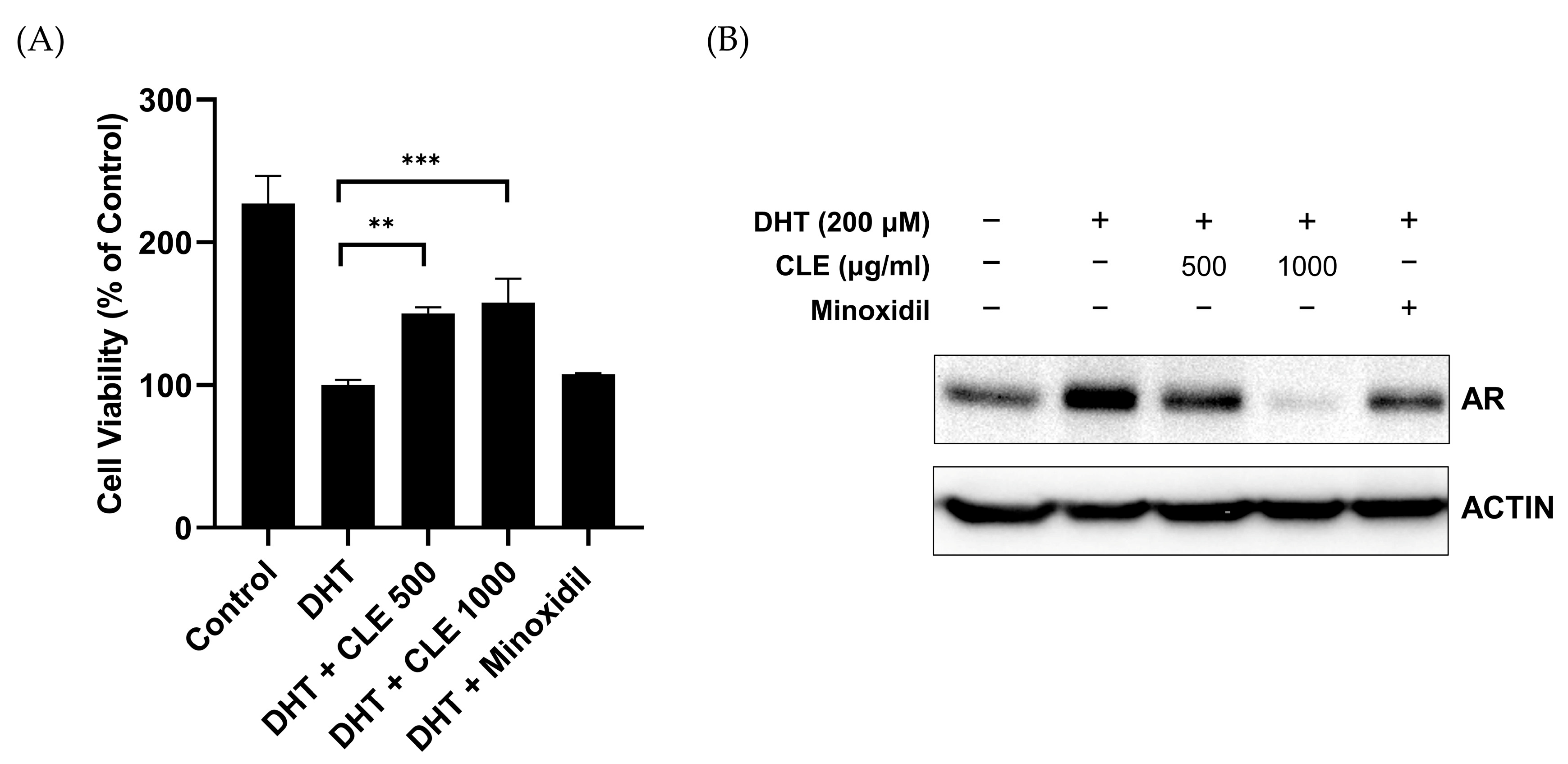
3.2. CLE Inhibits DHT-Induced AR Expression
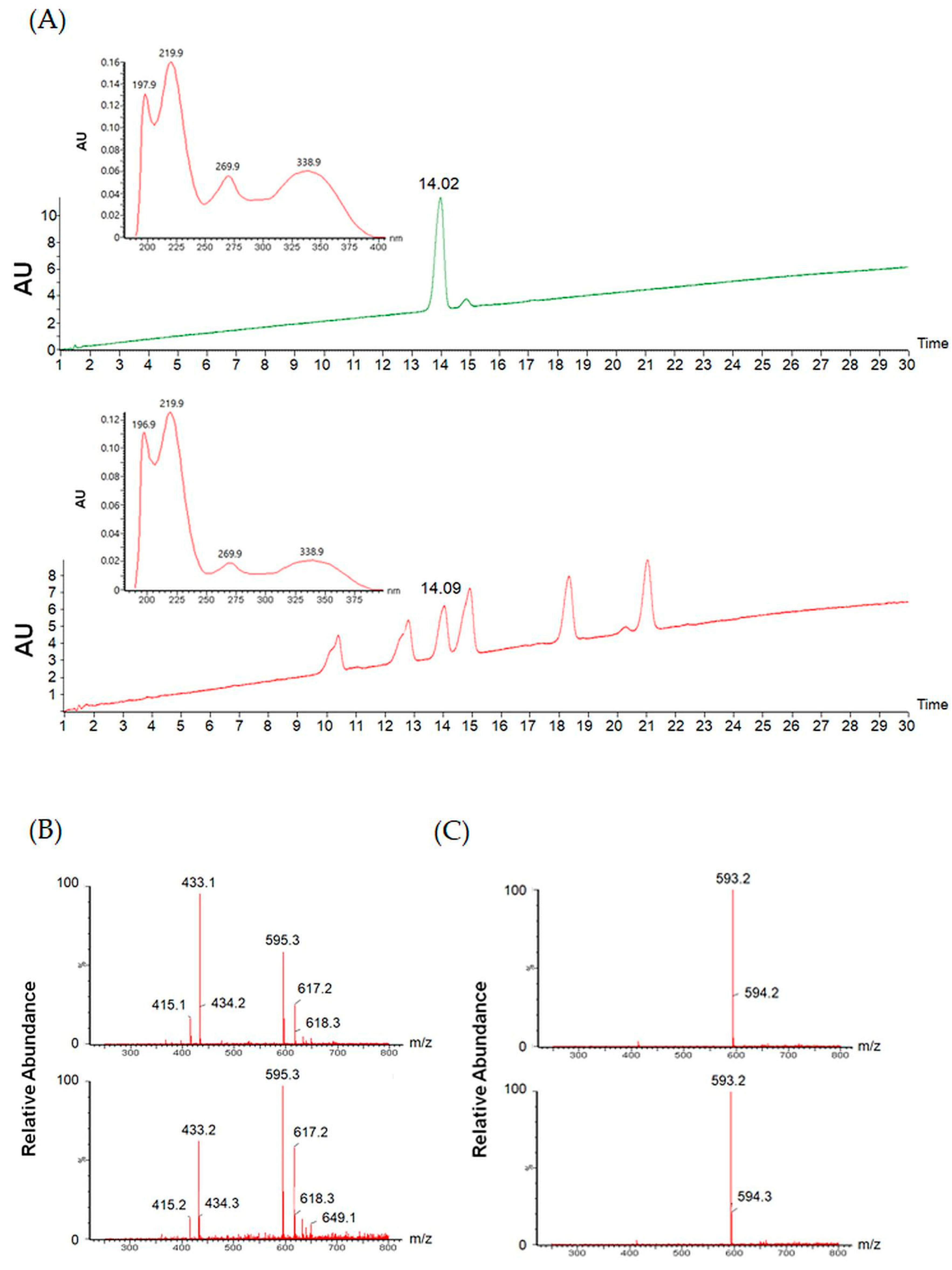
3.3. Meloside A Identification in CLE
3.4. Meloside A Inhibits DHT-Induced AR Nuclear Translocation
3.5. Meloside A Modulates AR Downstream Signaling and Protects Against Oxidative Stress
4. Discussions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hunt, N.; McHale, S. The psychological impact of alopecia. BMJ 2005, 331, 951–953. [Google Scholar] [CrossRef] [PubMed]
- Thom, E. Stress and the hair growth cycle: Cortisol-induced hair growth disruption. J. Drugs Dermatol. 2016, 15, 1001–1004. [Google Scholar] [PubMed]
- Guo, E.L.; Katta, R. Diet and hair loss: Effects of nutrient deficiency and supplement use. Dermatol. Pract. Concept. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Schmidt, J.B.; Lindmaier, A.; Trenz, A.; Schurz, B.; Spona, J. Hormone studies in females with androgenic hair loss. Gynecol. Obstet. Invest. 1991, 31, 235–239. [Google Scholar] [CrossRef]
- Rushton, D.H. Nutritional factors and hair loss. Clin. Exp. Dermatol. 2002, 27, 396–404. [Google Scholar] [CrossRef]
- Olsen, E.A. Female pattern hair loss. J. Am. Acad. Dermatol. 2001, 45, S70–S80. [Google Scholar] [CrossRef] [PubMed]
- Trost, L.; Saitz, T.R.; Hellstrom, W.J.G. Side effects of 5-alpha reductase inhibitors: A comprehensive review. Sex. Med. Rev. 2013, 1, 24–41. [Google Scholar] [CrossRef]
- Robert, S.; English, J. A hypothetical pathogenesis model for androgenic alopecia: Clarifying the dihydrotestosterone paradox and rate-limiting recovery factors. Med. Hypotheses 2018, 111, 73–81. [Google Scholar]
- Ceruti, J.M.; Leirós, G.J.; Balañá, M.E. Androgens and androgen receptor action in skin and hair follicles. Mol. Cell Endocrinol. 2018, 456, 122–133. [Google Scholar] [CrossRef]
- Lolli, F.; Pallotti, F.; Rossi, A.; Fortuna, M.C.; Caro, G.; Lenzi, A.; Sansone, A.; Lombardo, F. Androgenetic alopecia: A review. Endocrine 2017, 57, 9–17. [Google Scholar] [CrossRef]
- Foitzik, K.; Lindner, G.; Mueller-Roever, S.; Maurer, M.; Botchkareva, N.; Botchkarev, V.; Handjiski, B.; Metz, M.; Hibino, T.; Soma, T.; et al. Control of murine hair follicle regression (catagen) by TGF-beta1 in vivo. FASEB J. 2000, 14, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Kwack, M.H.; Ahn, J.S.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Dihydrotestosterone-inducible IL-6 inhibits elongation of human hair shafts by suppressing matrix cell proliferation and promotes regression of hair follicles in mice. J. Investig. Dermatol. 2012, 132, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Dai, J.; Ai, H.; Du, W.; Ji, H. β-Nicotinamide mononucleotide promotes cell proliferation and hair growth by reducing oxidative stress. Molecules 2024, 29, 798. [Google Scholar] [CrossRef] [PubMed]
- Hibberts, N.A.; Howell, A.E.; Randall, V.A. Balding hair follicle dermal papilla cells contain higher levels of androgen receptors than those from non-balding scalp. J. Endocrinol. 1998, 156, 59–65. [Google Scholar] [CrossRef]
- Randall, V.A.; Thornton, M.J.; Messenger, A.G. Cultured dermal papilla cells from androgen-dependent human hair follicles (e.g., beard) contain more androgen receptors than those from non-balding areas of scalp. J. Endocrinol. 1992, 133, 141–147. [Google Scholar] [CrossRef]
- Gupta, A.K.; Talukder, M.; Bamimore, M.A. Natural products for male androgenetic alopecia. Dermatol. Ther. 2022, 35, e15323. [Google Scholar] [CrossRef]
- Jung, Y.H.; Choi, C.W.; Shin, H.C.; Lim, J.R.; Chang, H.S.; Park, J.; Cho, J.H.; Park, M.R.; Lee, H.J.; Han, H.J. Cyanidin 3-O-arabinoside suppresses DHT-induced dermal papilla cell senescence by modulating p38-dependent ER-mitochondria contacts. J. Biomed. Sci. 2022, 29, 17. [Google Scholar] [CrossRef]
- Moon, I.J.; Yoon, H.K.; Kim, D.; Choi, M.E.; Han, S.H.; Park, J.H.; Hong, S.W.; Cho, H.; Lee, D.K.; Won, C.H. Efficacy of asymmetric siRNA targeting androgen receptors for the treatment of androgenetic alopecia. Mol. Pharm. 2023, 20, 128–135. [Google Scholar] [CrossRef]
- Park, I.; Kim, J.; Lee, J.; Kim, S.; Cho, O.; Yang, K.; Ahn, J.; Nahm, S.; Kim, H. Development of SSR markers by next-generation sequencing of Korean landraces of chamae (Cucumis melo var. makuwa). Mol. Biol. Rep. 2013, 40, 6855–6862. [Google Scholar] [CrossRef]
- Hosoya, T.; Masuda, Y.; Ohba, S.; Kumazawa, S. Component analysis of Cucumis melo L. leaves and their antioxidant activity. Nat. Prod. Res. 2024. ahead of print. [Google Scholar] [CrossRef]
- Pi, L.Q.; Lee, W.S.; Min, S.H. Hot water extract of oriental melon leaf promotes hair growth and prolongs anagen hair cycle: In vivo and in vitro evaluation. Food Sci. Biotechnol. 2016, 25, 575–580. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Liu, J.S.; Lin, A.C.; Yang, C.H.; Chung, W.H.; Wu, W.G. Minoxidil may suppress androgen receptor-related functions. Oncotarget 2014, 5, 2187–2197. [Google Scholar] [CrossRef]
- Luengo Escobar, A.; Magnum de Oliveira Silva, F.; Acevedo, P.; Nunes-Nesi, A.; Alberdi, M.; Reyes-Díaz, M. Different levels of UV-B resistance in Vaccinium corymbosum cultivars reveal distinct backgrounds of phenylpropanoid metabolites. Plant Physiol. Biochem. 2017, 118, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Premanand, A.; Rajkumari, B.R. Androgen modulation of Wnt/beta-catenin signaling in androgenetic alopecia. Arch. Dermatol. Res. 2018, 310, 391–399. [Google Scholar] [CrossRef]
- Masoodi, K.Z.; Xu, Y.; Dar, J.A.; Eisermann, K.; Pascal, L.E.; Parrinello, E.; Ai, J.; Johnston, P.A.; Nelson, J.B.; Wipf, P.; et al. Inhibition of androgen receptor nuclear localization and castration-resistant prostate tumor growth by pyrroloimidazole-based small molecules. Mol. Cancer Ther. 2017, 16, 2120–2129. [Google Scholar] [CrossRef]
- Cwynar, A.; Olszewska-Słonina, D.M.; Czajkowski, R. The impact of oxidative stress in androgenic alopecia in women. Postep. Dermatol. Alergol. 2020, 37, 119–120. [Google Scholar] [CrossRef]
- Raja Soh, R.S.S.; Hapidin, H.; Kasiram, M.Z. A scoping review on Cucumis melo and its anti-cancer properties. Malays. J. Med. Sci. 2024, 31, 63–77. [Google Scholar] [CrossRef]
- Paradisi, R.; Porcu, E.; Fabbri, R.; Seracchioli, R.; Battaglia, C.; Venturoli, S. Prospective cohort study on the effects and tolerability of flutamide in patients with female pattern hair loss. Ann. Pharmacother. 2011, 45, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Wysowski, D.K.; Fourcroy, J.L. Flutamide hepatotoxicity. J. Urol. 1996, 155, 209–212. [Google Scholar] [CrossRef]
- Hu, X.; Li, X.; Wu, S.; Jiang, X.; Chen, G.; Hu, Y.; Sun, J.; Bai, W. Cyanidin-3-O-glucoside and its derivative vitisin A alleviate androgenetic alopecia by exerting anti-androgen effect and inhibiting dermal papilla cell apoptosis. Eur. J. Pharmacol. 2024, 963, 176237. [Google Scholar] [CrossRef]
- Jung, E.S.; Lee, S.; Lim, S.H.; Ha, S.H.; Liu, K.H.; Lee, C.H. Metabolite profiling of the short-term responses of rice leaves (Oryza sativa cv. Ilmi) cultivated under different LED lights and its correlations with antioxidant activities. Plant Sci. 2013, 210, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Brauch, D.; Porzel, A.; Schumann, E.; Pillen, K.; Mock, H.P. Changes in isovitexin-O-glycosylation during the development of young barley plants. Phytochemistry 2018, 148, 11–20. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Júnior, R.G.; Reis, S.A.G.B.; de Oliveira, A.P.; Ferraz, C.A.A.; Rolim, L.A.; Lopes, N.P.; Rocha, J.M.; El Aouad, N.; Kritsanida, M.; Almeida, J.R.G.D.S. Photoprotective potential of Passiflora cincinnata Mast. (Passifloraceae) hydro-alcoholic extracts. Chem. Biodivers. 2024, 21, 11. [Google Scholar] [CrossRef]
- Choi, J.Y.; Boo, M.Y.; Boo, Y.C. Can Plant Extracts Help Prevent Hair Loss or Promote Hair Growth? A Review Comparing Their Therapeutic Efficacies, Phytochemical Components, and Modulatory Targets. Molecules 2024, 29, 2288. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Vida, A.; Galazi, M.; Rudman, S.; Chowdhury, S.; Sternberg, C.N. Enzalutamide for the treatment of metastatic castration-resistant prostate cancer. Drug Des. Dev. Ther. 2015, 9, 3325–3339. [Google Scholar] [CrossRef]
- Thanomkitti, K.; Fong-Ngern, K.; Sueksakit, K.; Thuangtong, R.; Thongboonkerd, V. Molecular functional analyses revealed essential roles of HSP90 and lamin A/C in growth, migration, and self-aggregation of dermal papilla cells. Cell Death Discov. 2018, 4, 53. [Google Scholar] [CrossRef]
- Tien, A.H.; Sadar, M.D. Keys to unlock androgen receptor translocation. J. Biol. Chem. 2019, 294, 8711–8712. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.J.; Song, B.G.; Song, J.H.; Park, S.H.; Wang, D.H.; Kwon, H.K.; Lee, J.-E. Meloside A Protects Dermal Papilla Cells from DHT-Induced Damage via Androgen Receptor Modulation. Curr. Issues Mol. Biol. 2025, 47, 436. https://doi.org/10.3390/cimb47060436
Park HJ, Song BG, Song JH, Park SH, Wang DH, Kwon HK, Lee J-E. Meloside A Protects Dermal Papilla Cells from DHT-Induced Damage via Androgen Receptor Modulation. Current Issues in Molecular Biology. 2025; 47(6):436. https://doi.org/10.3390/cimb47060436
Chicago/Turabian StylePark, Hyun Jun, Bong Geun Song, Ji Hoon Song, Seung Hee Park, Da Hye Wang, Ho Kyun Kwon, and Ji-Ean Lee. 2025. "Meloside A Protects Dermal Papilla Cells from DHT-Induced Damage via Androgen Receptor Modulation" Current Issues in Molecular Biology 47, no. 6: 436. https://doi.org/10.3390/cimb47060436
APA StylePark, H. J., Song, B. G., Song, J. H., Park, S. H., Wang, D. H., Kwon, H. K., & Lee, J.-E. (2025). Meloside A Protects Dermal Papilla Cells from DHT-Induced Damage via Androgen Receptor Modulation. Current Issues in Molecular Biology, 47(6), 436. https://doi.org/10.3390/cimb47060436