

An Elevated IL10 mRNA Combined with Lower TNFA mRNA Level in Active Rheumatoid Arthritis Peripheral Blood
 , ,
, ,  and
and
Abstract

1. Introduction
2. Materials and Methods
2.1. Study Subjects
2.2. Blood Samples
2.3. Reverse Transcription PCR
2.4. Real-Time Quantitative PCR
2.5. Quantification of Serum Cytokines Concentrations
2.6. Data Analysis
3. Results
3.1. Study Subjects
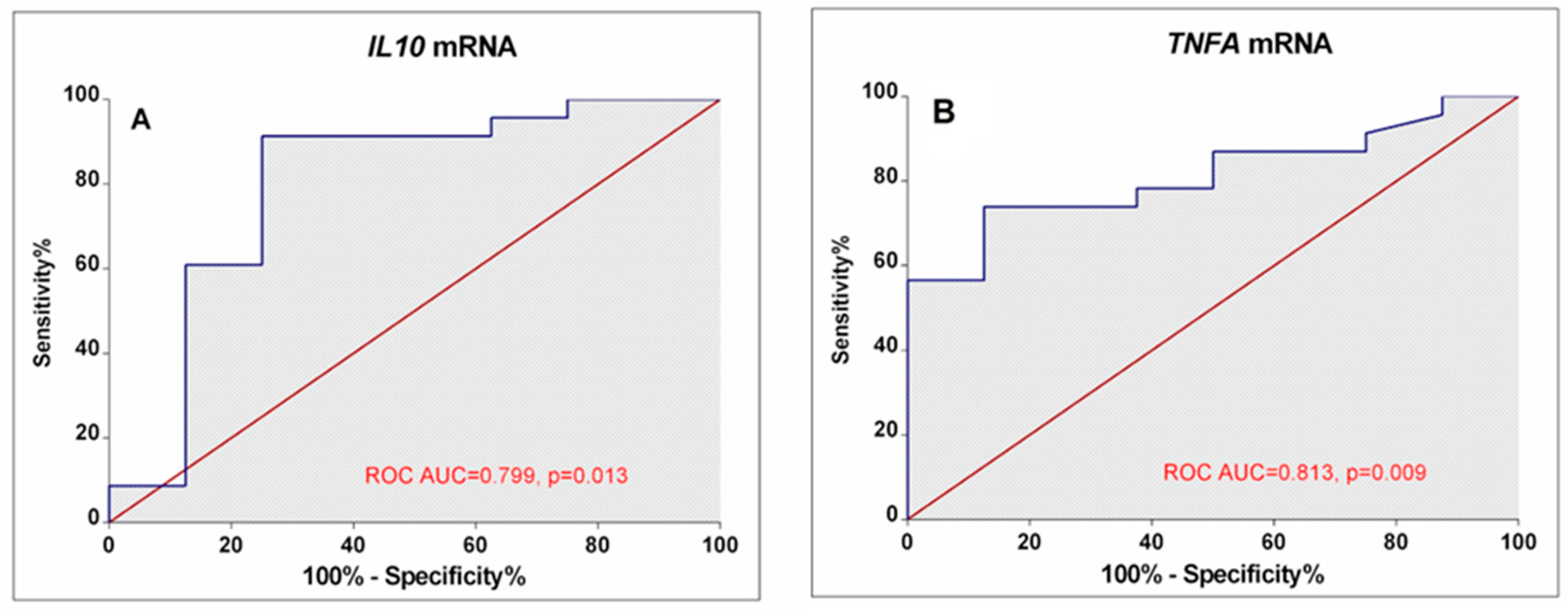
3.2. Gene Expression at mRNA Level in Peripheral Blood Cells
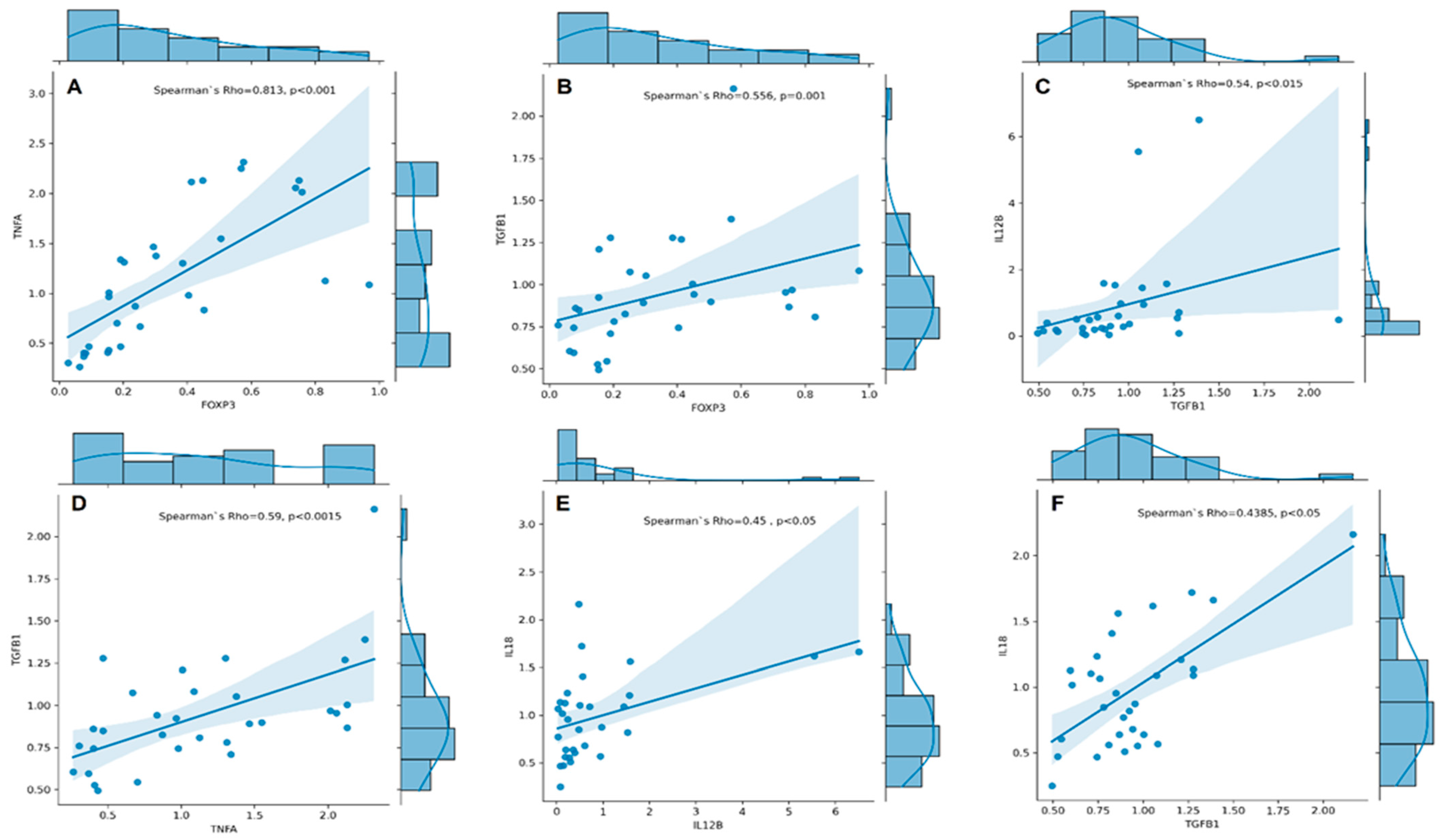
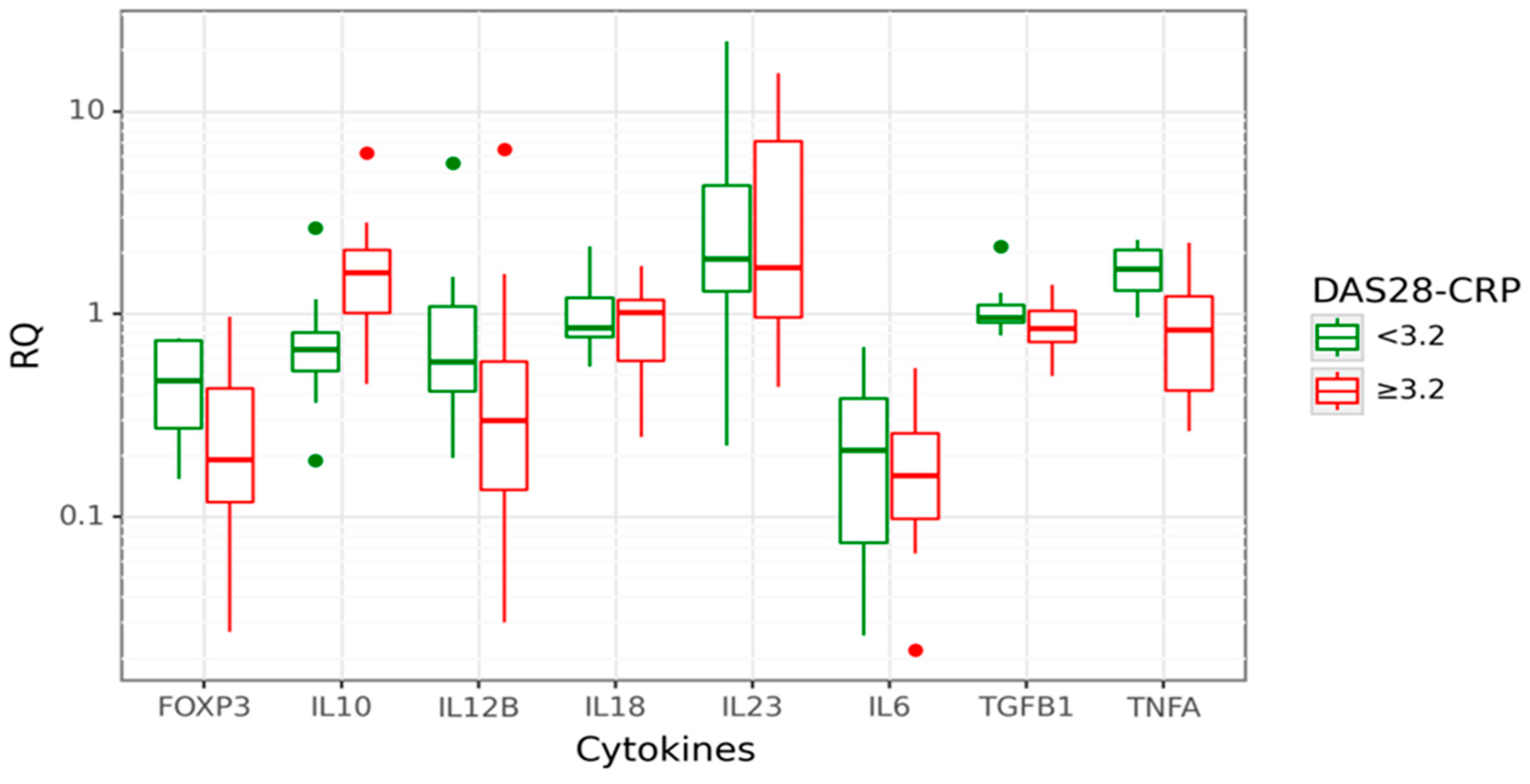
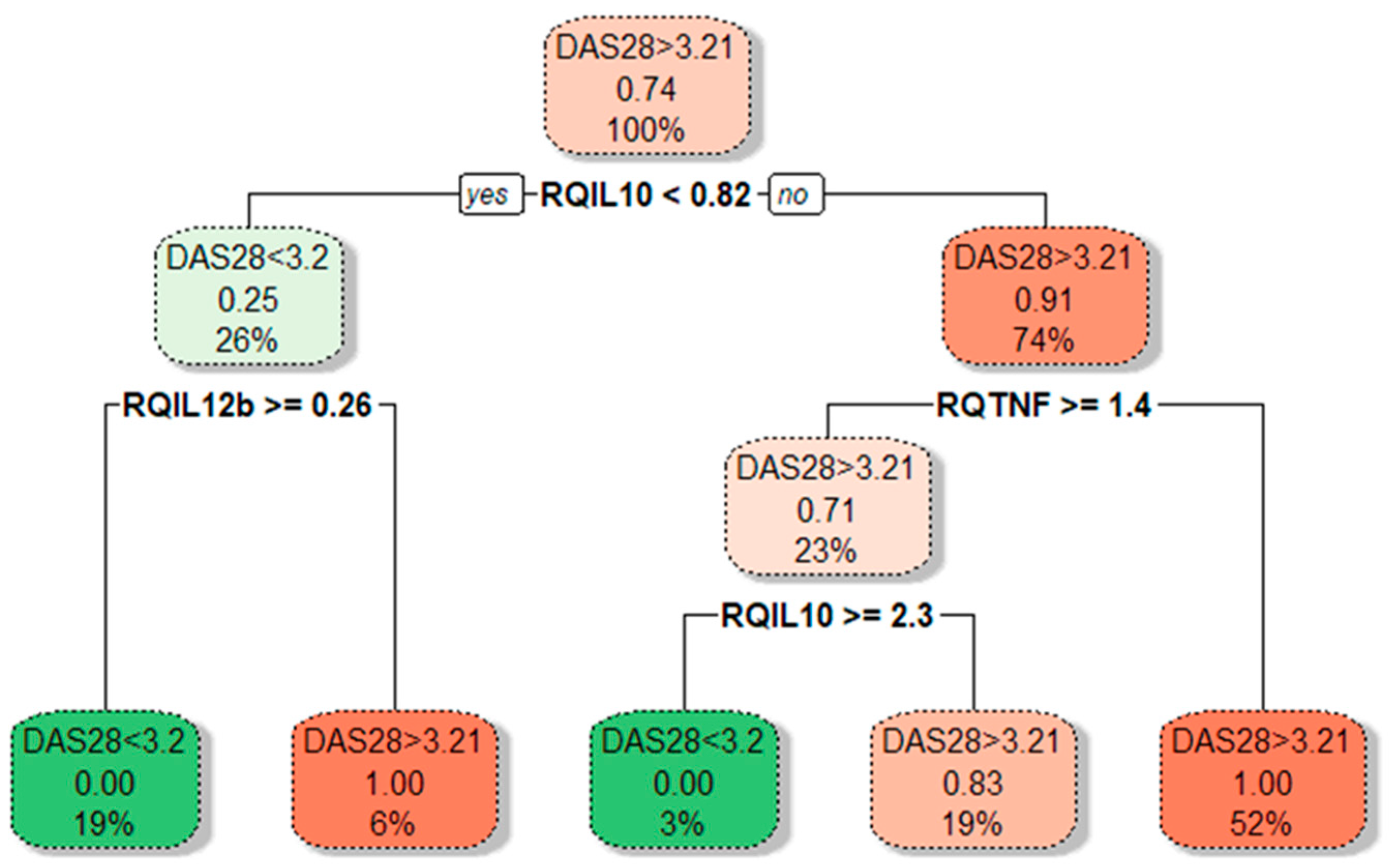
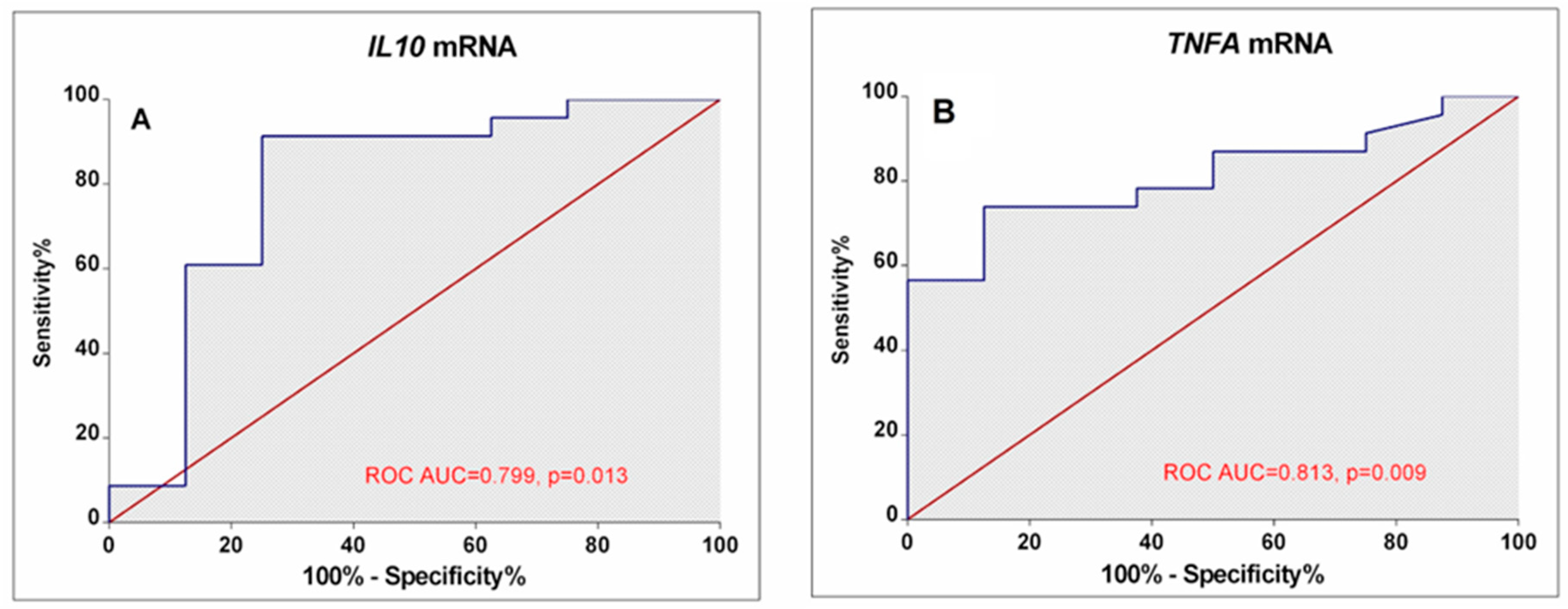
3.3. Associations between Cytokine Gene Expression and RA Activity
3.4. Associations between Cytokine Gene Expression and RA Disease Characteristics
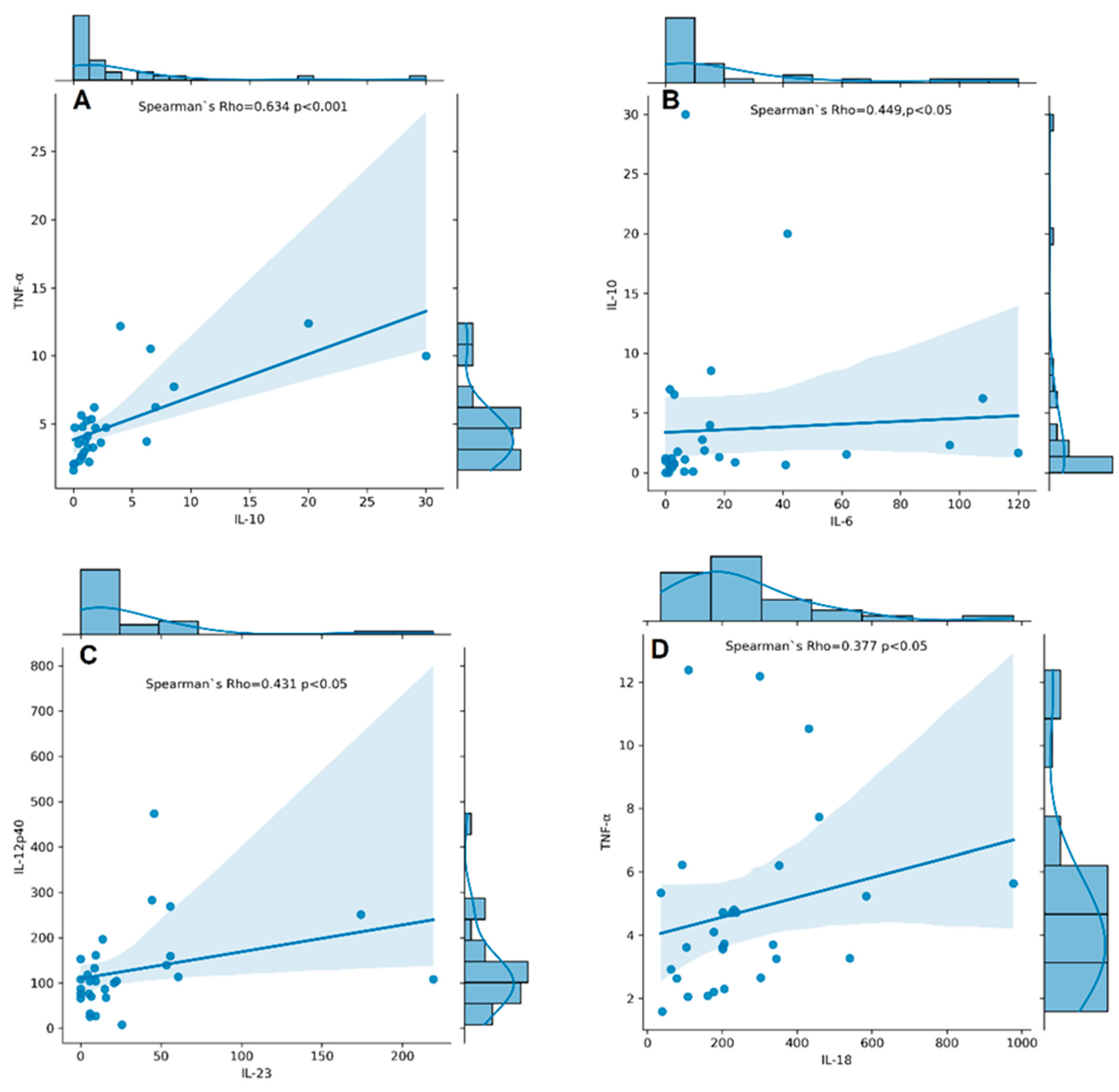
3.5. Associations between Cytokine Serum Levels and RA Disease Characteristics
4. Discussion
Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Silman, A.J.; Pearson, J.E. Epidemiology and Genetics of Rheumatoid Arthritis—Arthritis Research & Therapy. Available online: https://arthritis-research.biomedcentral.com/articles/10.1186/ar578 (accessed on 7 February 2024).
- Niemantsverdriet, E.; van den Akker, E.B.; Boeters, D.M.; van den Eeden, S.J.; Geluk, A.; van der Helm-van Mil, A.H. Gene Expression Identifies Patients Who Develop Inflammatory Arthritis in a Clinically Suspect Arthralgia Cohort. Arthritis Res. Ther. 2020, 22, 266. [Google Scholar] [CrossRef]
- Wu, F.; Gao, J.; Kang, J.; Wang, X.; Niu, Q.; Liu, J.; Zhang, L. B Cells in Rheumatoid Arthritis: Pathogenic Mechanisms and Treatment Prospects. Front. Immunol. 2021, 12, 750753. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Bello, J.; Oregón-Romero, E.; Vázquez-Villamar, M.; García-Arellano, S.; Valle, Y.; Padilla-Gutiérrez, J.R.; Román-Fernández, I.V.; Palafox-Sánchez, C.A.; Martínez-Bonilla, G.E.; Muñoz-Valle, J.F. Aberrant Expression of Interleukin-10 in Rheumatoid Arthritis: Relationship with IL10 Haplotypes and Autoantibodies. Cytokine 2017, 95, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Markusse, I.M.; Dirven, L.; Gerards, A.H.; van Groenendael, J.H.; Ronday, H.K.; Kerstens, P.J.; Lems, W.F.; Huizinga, T.W.; Allaart, C.F. Disease Flares in Rheumatoid Arthritis Are Associated with Joint Damage Progression and Disability: 10-Year Results from the Best Study. Arthritis Res. Ther. 2015, 17, 232. [Google Scholar] [CrossRef] [PubMed]
- Figus, F.A.; Piga, M.; Azzolin, I.; McConnell, R.; Iagnocco, A. Rheumatoid Arthritis: Extra-Articular Manifestations and Comorbidities. Autoimmun. Rev. 2021, 20, 102776. [Google Scholar] [CrossRef] [PubMed]
- McInnes, I.B.; Buckley, C.D.; Isaacs, J.D. Cytokines in Rheumatoid Arthritis—Shaping the Immunological Landscape. Nat. Rev. Rheumatol. 2015, 12, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, G.; Ivanova, M.; Stanilov, I.; Miteva, L.; Stanilova, S.; Manolova, I. Influence of IL10 and TGFB1 Promoter Polymorphisms on Serum Cytokine Levels in Development and Severity of Ra. Int. J. Mol. Sci. 2022, 23, 11955. [Google Scholar] [CrossRef] [PubMed]
- Manolova, I.; Ivanova, M.; Vasilev, G.; Stoilov, R.; Miteva, L.; Stanilova, S. Impact of IL12B Polymorphisms on Genetic Susceptibility and Il-12p40 and IL-23 Serum Levels in Rheumatoid Arthritis. Immunol. Investig. 2019, 49, 1–14. [Google Scholar] [CrossRef]
- Manolova, I.; Ivanova, M.; Stoilov, R.; Rashkov, R.; Stanilova, S. Association of Single Nucleotide Polymorphism at Position −308 of the Tumor Necrosis Factor-Alpha Gene with Ankylosing Spondylitis and Rheumatoid Arthritis. Biotechnol. Biotechnol. Equip. 2014, 28, 1108–1114. [Google Scholar] [CrossRef] [PubMed]
- Nemtsova, M.V.; Zaletaev, D.V.; Bure, I.V.; Mikhaylenko, D.S.; Kuznetsova, E.B.; Alekseeva, E.A.; Beloukhova, M.I.; Deviatkin, A.A.; Lukashev, A.N.; Zamyatnin, A.A. Epigenetic Changes in the Pathogenesis of Rheumatoid Arthritis. Front. Genet. 2019, 10, 570. [Google Scholar] [CrossRef]
- Paradowska-Gorycka, A.; Wajda, A.; Romanowska-Próchnicka, K.; Walczuk, E.; Kuca-Warnawin, E.; Kmiolek, T.; Stypinska, B.; Rzeszotarska, E.; Majewski, D.; Jagodzinski, P.P.; et al. Th17/Treg-Related Transcriptional Factor Expression and Cytokine Profile in Patients with Rheumatoid Arthritis. Front. Immunol. 2020, 11, 572858. [Google Scholar] [CrossRef] [PubMed]
- Vasilev, G.; Manolova, I.; Ivanova, M.; Stanilov, I.; Miteva, L.; Stanilova, S. The Role of IL-18 in Addition to Th17 Cytokines in Rheumatoid Arthritis Development and Treatment in Women. Sci. Rep. 2021, 11, 15391. [Google Scholar] [CrossRef] [PubMed]
- Iyer, S.S.; Cheng, G. Role of Interleukin 10 Transcriptional Regulation in Inflammation and Autoimmune Disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; O’Garra, A. Il-10 Family Cytokines IL-10 and IL-22: From Basic Science to Clinical Translation. Immunity 2019, 50, 871–891. [Google Scholar] [CrossRef] [PubMed]
- van Helvoort, E.M.; van der Heijden, E.; van Roon, J.A.G.; Eijkelkamp, N.; Mastbergen, S.C. The Role of Interleukin-4 and Interleukin-10 in Osteoarthritic Joint Disease: A Systematic Narrative Review. Cartilage 2022, 13, 194760352210981. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Bozec, A.; Ramming, A.; Schett, G. Anti-Inflammatory and Immune-Regulatory Cytokines in Rheumatoid Arthritis. Nat. Rev. Rheumatol. 2018, 15, 9–17. [Google Scholar] [CrossRef] [PubMed]
- Kondo, N.; Kuroda, T.; Kobayashi, D. Cytokine Networks in the Pathogenesis of Rheumatoid Arthritis. Int. J. Mol. Sci. 2021, 22, 10922. [Google Scholar] [CrossRef]
- Oswald, M.; Curran, M.E.; Lamberth, S.L.; Townsend, R.M.; Hamilton, J.D.; Chernoff, D.N.; Carulli, J.; Townsend, M.J.; Weinblatt, M.E.; Kern, M.; et al. Modular Analysis of Peripheral Blood Gene Expression in Rheumatoid Arthritis Captures Reproducible Gene Expression Changes in Tumor Necrosis Factor Responders. Arthritis Rheumatol. 2015, 67, 344–351. [Google Scholar] [CrossRef]
- Tchetina, E.V.; Markova, G.A.; Satybaldyev, A.M.; Lila, A.M. Downregulation of Tumour Necrosis Factor α Gene Expression in Peripheral Blood Mononuclear Cells Cultured in the Presence of Tofacitinib Prior to Therapy Is Associated with Clinical Remission in Patients with Rheumatoid Arthritis. Curr. Issues Mol. Biol. 2022, 44, 1941–1949. [Google Scholar] [CrossRef]
- Hedman, Å.K.; Winter, E.; Yoosuf, N.; Benita, Y.; Berg, L.; Brynedal, B.; Folkersen, L.; Klareskog, L.; Maciejewski, M.; Sirota-Madi, A.; et al. Peripheral Blood Cellular Dynamics of Rheumatoid Arthritis Treatment Informs about Efficacy of Response to Disease Modifying Drugs. Sci. Rep. 2023, 13, 10058. [Google Scholar] [CrossRef]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O.; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. 2010 Rheumatoid Arthritis Classification Criteria: An American College of Rheumatology/European League against Rheumatism Collaborative Initiative. Arthritis Rheum. 2010, 62, 2569–2581. [Google Scholar] [CrossRef]
- Prevoo, M.L.; Van’T Hof, M.A.; Kuper, H.H.; Van Leeuwen, M.A.; Van De Putte, L.B.; Van Riel, P.L. Modified Disease Activity Scores That Include Twenty-Eight-Joint Counts Development and Validation in a Prospective Longitudinal Study of Patients with Rheumatoid Arthritis. Arthritis Rheum. 1995, 38, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Miteva, L.D.; Manolova, I.M.; Ivanova, M.G.; Stoilov, R.M.; Stanilova, S.A. High Interleukin-18 and Low FOXP3 Mrnas in Peripheral Blood of Women with Severe Systemic Lupus Erythematosus: A Cross-Sectional Study. Rheumatol. Int. 2020, 40, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Paradowska-Gorycka, A.; Grzybowska-Kowalczyk, A.; Wojtecka-Lukasik, E.; Maslinski, S. IL-23 in the Pathogenesis of Rheumatoid Arthritis. Scand. J. Immunol. 2010, 71, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Dungan, L.S.; Lalor, S.J.; Sutton, C.E.; Basdeo, S.A.; Fletcher, J.M.; Mills, K.H.G. CS05-6. Caspase-1–Processed Cytokines Il-1β and IL-18 Promote IL-17 Production by and CD4 T Cells That Mediate Autoimmunity. Cytokine 2011, 56, 13. [Google Scholar] [CrossRef]
- Bystrom, J.; Clanchy, F.I.; Taher, T.E.; Mangat, P.; Jawad, A.S.; Williams, R.O.; Mageed, R.A. TNFA in the Regulation of Treg and Th17 Cells in Rheumatoid Arthritis and Other Autoimmune Inflammatory Diseases. Cytokine 2018, 101, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Kmiołek, T.; Rzeszotarska, E.; Wajda, A.; Walczuk, E.; Kuca-Warnawin, E.; Romanowska-Próchnicka, K.; Stypinska, B.; Majewski, D.; Jagodzinski, P.P.; Pawlik, A.; et al. The Interplay between Transcriptional Factors and Micrornas as an Important Factor for Th17/Treg Balance in RA Patients. Int. J. Mol. Sci. 2020, 21, 7169. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, M.; Xie, H.; Hong, F.; Yang, S. Role of Mirnas in Rheumatoid Arthritis Therapy. Cells 2023, 12, 1749. [Google Scholar] [CrossRef]
- Farrugia, M.; Baron, B. The Role of TNF-α in Rheumatoid Arthritis: A Focus on Regulatory T Cells. J. Clin. Transl. Res. 2016, 2, 84–90. [Google Scholar] [CrossRef]
- Chen, W.; Wang, Q.; Ke, Y.; Lin, J. Neutrophil Function in an Inflammatory Milieu of Rheumatoid Arthritis. J. Immunol. Res. 2018, 2018, 8549329. [Google Scholar] [CrossRef] [PubMed]
- Sempere-Ortells, J.M.; Pérez-García, V.; Marín-Alberca, G.; Peris-Pertusa, A.; Benito, J.M.; Marco, F.M.; Zubcoff, J.J.; Navarro-Blasco, F.J. Quantification and Phenotype of Regulatory T Cells in Rheumatoid Arthritis According to Disease Activity Score-28. Autoimmunity 2009, 42, 636–645. [Google Scholar] [CrossRef]
- Galeazzi, M.; Bazzichi, L.; Sebastiani, G.D.; Neri, D.; Garcia, E.; Ravenni, N.; Giovannoni, L.; Wilton, J.; Bardelli, M.; Baldi, C.; et al. A phase IB clinical trial with Dekavil (F8-IL10), an immunoregulatory ‘armed antibody’ for the treatment of rheumatoid arthritis, used in combination with methotrexate. Isr. Med. Assoc. J. 2014, 16, 666. [Google Scholar]
- Saraiva, M.; Vieira, P.; O’Garra, A. Biology and Therapeutic Potential of Interleukin-10. J. Exp. Med. 2019, 217, e20190418. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Ma, C.; Cong, B.; Li, S.; Chen, H.; Zhang, J. Hypomethylation of Proximal CPG Motif of Interleukin-10 Promoter Regulates Its Expression in Human Rheumatoid Arthritis. Acta Pharmacol. Sin. 2011, 32, 1373–1380. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Chen, H.; Wang, F.; Wang, Z.; Zhao, Q.; Liu, S.; Huang, B.; Lou, S.; Zuo, J. Down-Regulation of Microrna-98 Promoted Apoptosis of TNF-α Stimulated Human Fibroblast-like Synoviocytes via up-Regulating IL-10. Gene 2019, 706, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Ai, R.; Boyle, D.L.; Wang, W.; Firestein, G.S. Distinct DNA Methylation Patterns of Rheumatoid Arthritis Peripheral Blood and Synovial Tissue T Cells. ACR Open Rheumatol. 2021, 3, 127–132. [Google Scholar] [CrossRef]
- Adams, C.; Nair, N.; Plant, D.; Verstappen, S.M.; Quach, H.L.; Quach, D.L.; Carvidi, A.; Nititham, J.; Nakamura, M.; Graf, J.; et al. Identification of Cell-Specific Differential DNA Methylation Associated with Methotrexate Treatment Response in Rheumatoid Arthritis. Arthritis Rheumatol. 2023, 75, 1088–1097. [Google Scholar] [CrossRef]
- Hobl, E.-L.; Mader, R.M.; Erlacher, L.; Duhm, B.; Mustak, M.; Bröll, H.; Högger, P.; Kalipciyan, M.; Jilma, B. The Influence of Methotrexate on the Gene Expression of the Pro-Inflammatory Cytokine IL-12A in the Therapy of Rheumatoid Arthritis. Available online: https://www.clinexprheumatol.org/abstract.asp?a=4832 (accessed on 15 January 2024).
- Migita, K.; Iwanaga, N.; Izumi, Y.; Kawahara, C.; Kumagai, K.; Nakamura, T.; Koga, T.; Kawakami, A. TNF-α-Induced Mir-155 Regulates IL-6 Signaling in Rheumatoid Synovial Fibroblasts. BMC Res. Notes 2017, 10, 403. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RA | Controls | p-Value | |
|---|---|---|---|
| n | 31 | 21 | |
| Sex, n (%) | 1.0 | ||
| Male | 5 (16.1) | 3 (14.3) | |
| Female | 26 (83.9) | 18 (85.7) | |
| Age (mean ± SD; years) | 43.1 ± 14.2 | 39.3 ± 12.2 | 0.324 |
| Disease duration (mean ± SD; years) | 9.6 ± 8.5 | ||
| RF-positive patients; n (%) (RF positivity > 14.0 U/mL) | 25 (81) | ||
| Anti-CCP-positive patients; n (%) (anti-CCP positivity > 5 RU/mL) | 22 (71) | ||
| CRP (mean ± SD; mg/L) (CRP reference range 0–6 mg/L) | 19.2 ± 39.64 | ||
| DAS28-CRP (mean ± SD) | 4.1 ± 1.05 | ||
| <3.2; n (%) | 8 (26) | ||
| ≥3.2; n (%) | 23 (74) | ||
| Therapy, n (%) | |||
| Symptomatic | 9 (29) | ||
| csDMARDs | 22 (71) |
| mRNA (RQ) | IL6 | IL10 | IL12B | IL18 | IL23 | TNFA | TGFB1 | FOXP3 |
|---|---|---|---|---|---|---|---|---|
| Disease onset | 0.25 ± 0.24 | 1.33 ± 0.71 | 0.92 ± 1.56 | 0.91 ± 0.37 | 4.74 ± 6.0 | 1.28 ± 0.67 | 0.90 ± 0.23 | 0.40 ± 0.28 |
| <40 years (n = 16) | 0.21 ± 0.17 | 1.71 ± 1.45 | 0.82 ± 1.39 | 1.05 ± 0.50 | 4.62 ± 5.5 | 0.98 ± 0.65 | 0.98 ± 0.41 | 0.28 ± 0.23 |
| >40 years (n = 15) | p = 0.833 | p = 0.833 | p = 0.698 | p = 0.495 | p = 0.711 | p = 0.299 | p = 0.892 | p = 0.247 |
| Disease duration | 0.28 ± 0.21 | 1.01 ± 0.26 | 0.39 ± 0.50 | 0.76 ± 0.38 | 4.74 ± 6.0 | 0.74 ± 0.43 | 0.81 ± 0.19 | 0.27 ± 0.19 |
| <2 years (n = 7) | 0.22 ± 0.22 | 1.66 ± 1.24 | 1.00 ± 1.62 | 1.04 ± 0.45 | 4.62 ± 5.5 | 1.25 ± 0.68 | 0.98 ± 0.35 | 0.37 ± 0.27 |
| >2 years (n = 24) | p = 0.692 | p = 0.167 | p = 0.182 | p = 0.495 | p = 0.139 | p = 0.085 | p = 0.216 | p = 0.473 |
| RF | 0.14 ± 0.10 | 1.25 ± 0.95 | 1.54 ± 2.44 | 0.91 ± 0.37 | 2.79 ± 2.52 | 1.53 ± 0.80 | 1.15 ± 0.55 | 0.43 ± 0.25 |
| negative (n = 6) | 0.25 ± 0.22 | 1.58 ± 1.17 | 0.70 ± 1.13 | 1.05 ± 0.50 | 5.14 ± 6.18 | 1.04 ± 0.61 | 0.89 ± 0.23 | 0.32 ± 0.26 |
| positive (n = 25) | p = 0.513 | p = 0.417 | p = 0.698 | p = 0.295 | p = 0.764 | p = 0.153 | p = 0.563 | p = 0.104 |
| Anti-CCP | 0.28 ± 0.28 | 1.95 ± 1.71 | 1.13 ± 2.06 | 1.16 ± 0.57 | 4.45 ± 5.78 | 1.30 ± 0.81 | 1.01 ± 0.49 | 0.39 ± 0.27 |
| negative (n = 9) | 0.22 ± 0.19 | 1.34 ± 0.77 | 0.76 ± 1.17 | 0.90 ± 0.37 | 4.78 ± 5.79 | 1.02 ± 0.58 | 0.91 ± 0.23 | 0.33 ± 0.26 |
| positive (n = 22) | p = 0.604 | p = 0.428 | p = 0.881 | p = 0.273 | p = 0.881 | p = 0.292 | p = 0.949 | p = 0.507 |
| Therapy | 0.23 ± 0.18 | 2.08 ± 1.65 | 1.35 ± 2.0 | 0.92 ± 0.43 | 6.76 ± 6.02 | 1.22 ± 0.75 | 0.99 ± 0.25 | 0.42 ± 0.30 |
| Symptomatic (n = 9) | 0.24 ± 0.26 | 1.3 ± 0.78 | 0.69 ± 1.19 | 1.03 ± 0.46 | 3.77 ± 5.6 | 1.1 ± 0.66 | 0.92 ± 0.36 | 0.29 ± 0.22 |
| csDMARDs (n = 22) | p = 0.903 | p = 0.334 | p = 0.294 | p = 0.468 | p = 0.360 | p = 0.884 | p = 0.381 | p = 0.152 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasilev, G.; Vasileva, V.; Ivanova, M.; Stanilova, S.; Manolova, I.; Miteva, L. An Elevated IL10 mRNA Combined with Lower TNFA mRNA Level in Active Rheumatoid Arthritis Peripheral Blood. Curr. Issues Mol. Biol. 2024, 46, 2644-2657. https://doi.org/10.3390/cimb46030167
Vasilev G, Vasileva V, Ivanova M, Stanilova S, Manolova I, Miteva L. An Elevated IL10 mRNA Combined with Lower TNFA mRNA Level in Active Rheumatoid Arthritis Peripheral Blood. Current Issues in Molecular Biology. 2024; 46(3):2644-2657. https://doi.org/10.3390/cimb46030167
Chicago/Turabian StyleVasilev, Georgi, Viktoria Vasileva, Mariana Ivanova, Spaska Stanilova, Irena Manolova, and Lyuba Miteva. 2024. "An Elevated IL10 mRNA Combined with Lower TNFA mRNA Level in Active Rheumatoid Arthritis Peripheral Blood" Current Issues in Molecular Biology 46, no. 3: 2644-2657. https://doi.org/10.3390/cimb46030167
APA StyleVasilev, G., Vasileva, V., Ivanova, M., Stanilova, S., Manolova, I., & Miteva, L. (2024). An Elevated IL10 mRNA Combined with Lower TNFA mRNA Level in Active Rheumatoid Arthritis Peripheral Blood. Current Issues in Molecular Biology, 46(3), 2644-2657. https://doi.org/10.3390/cimb46030167