

A Prochlorperazine-Induced Decrease in Autonomous Muscle Activity during Hindlimb Unloading Is Accompanied by Preserved Slow Myosin mRNA Expression
 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
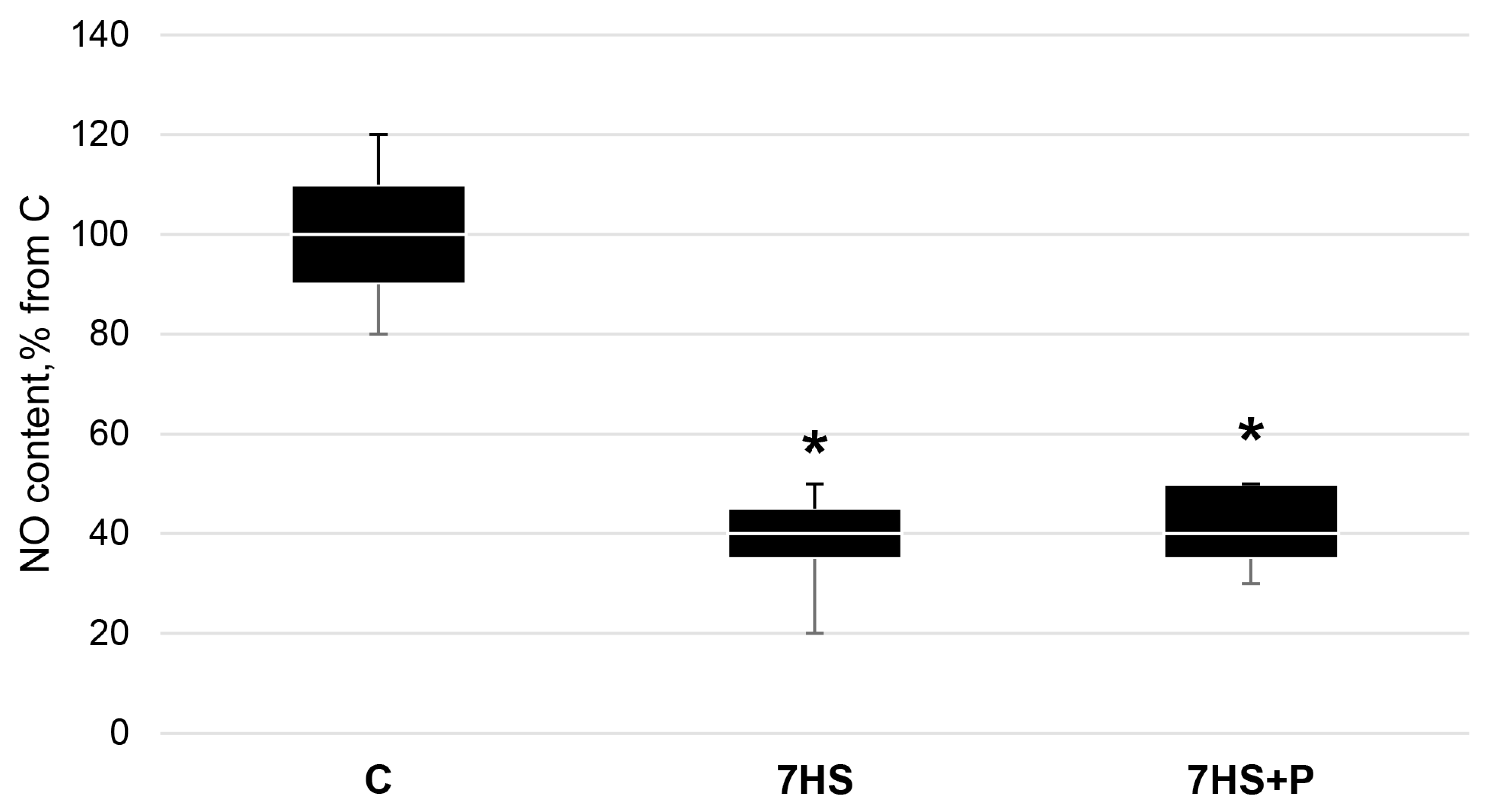
2.2. NO Content Detection
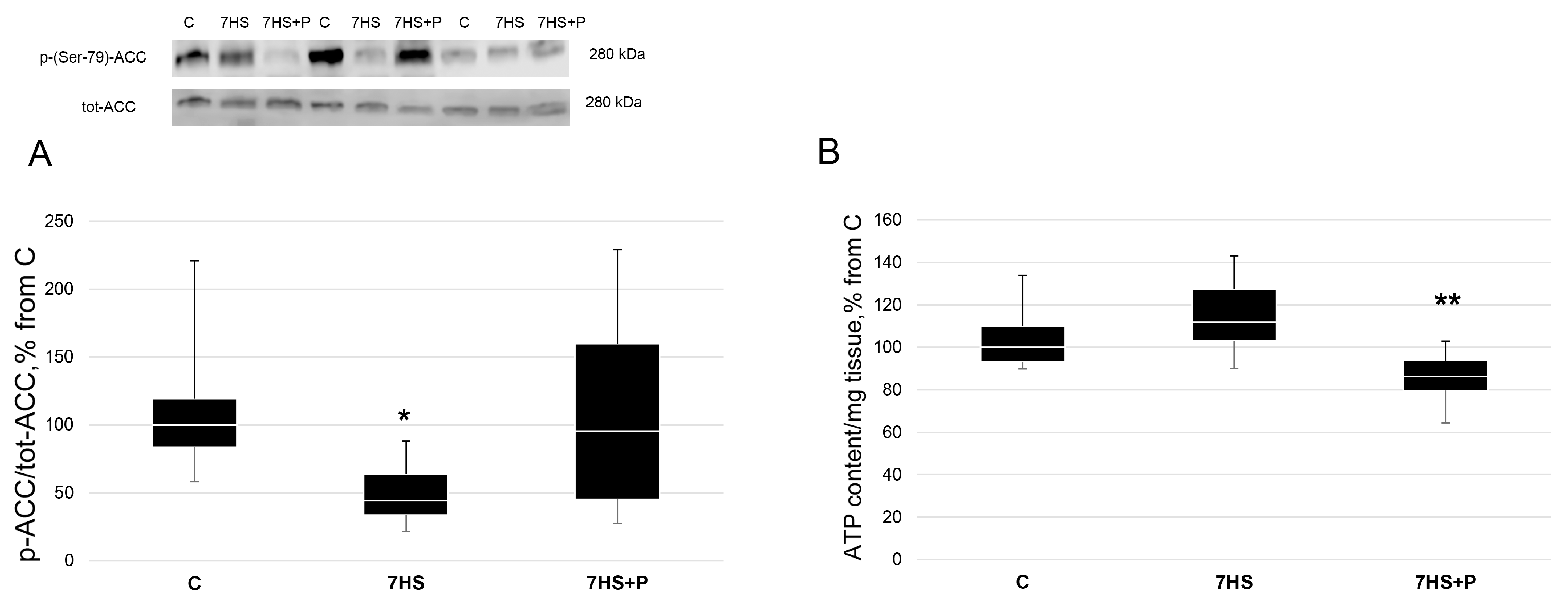
2.3. ATP Content Detection
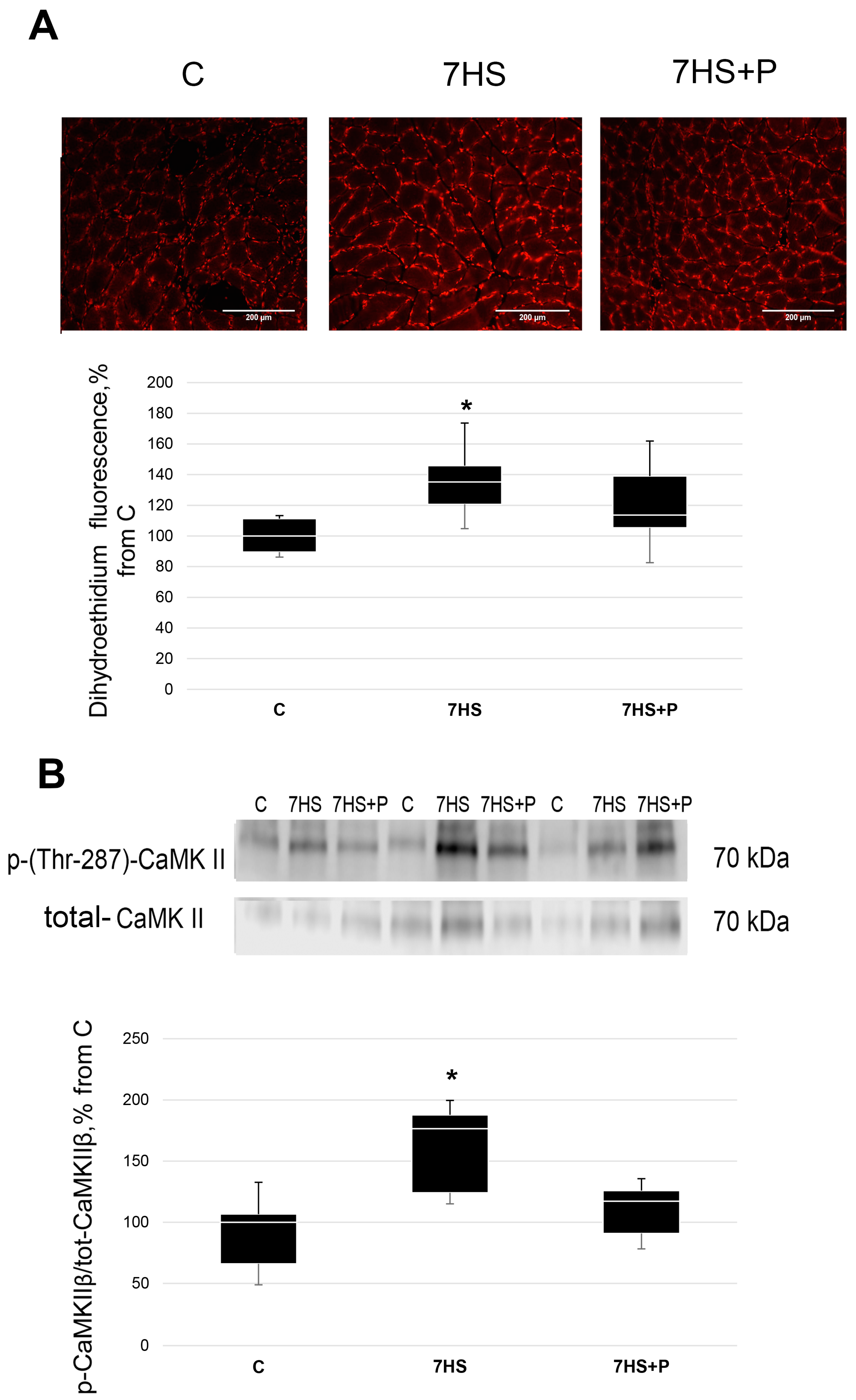
2.4. Detection of Superoxide Anion
2.5. Total Protein Fraction Preparation
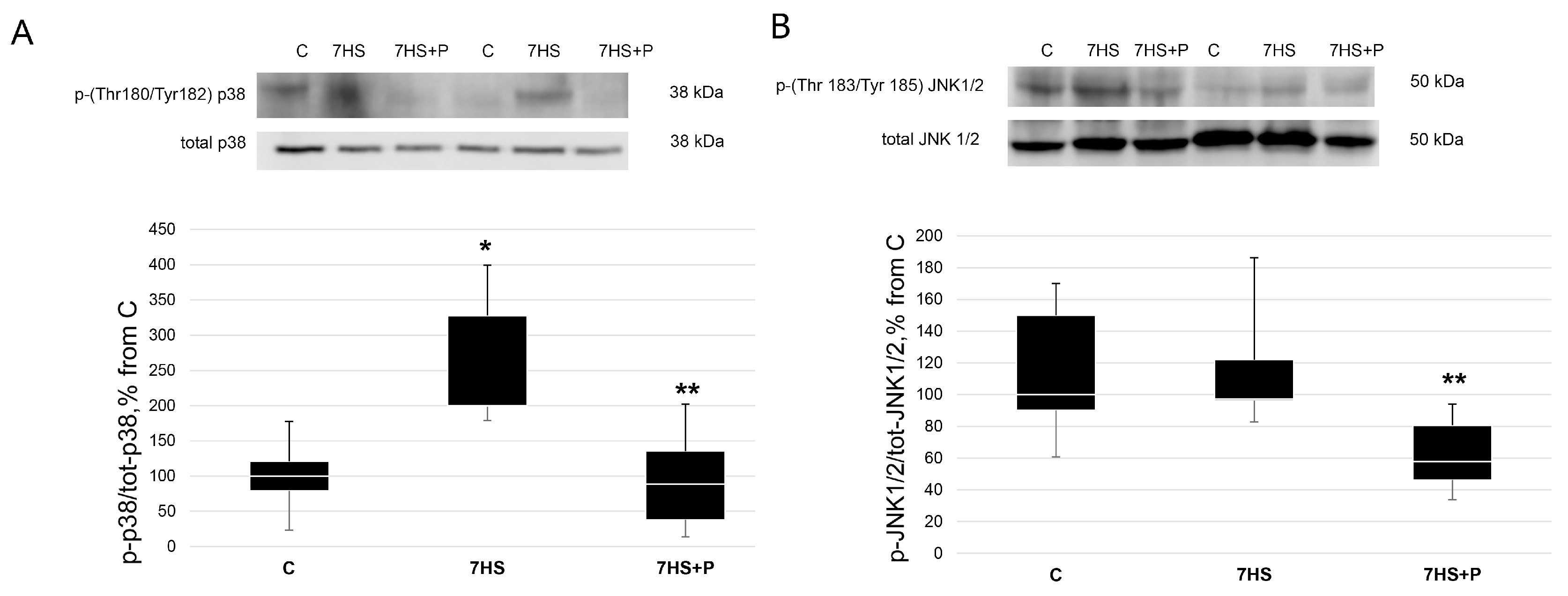
2.6. Western Blot Analysis
2.7. RNA Extraction, Reverse Transcription and PCR
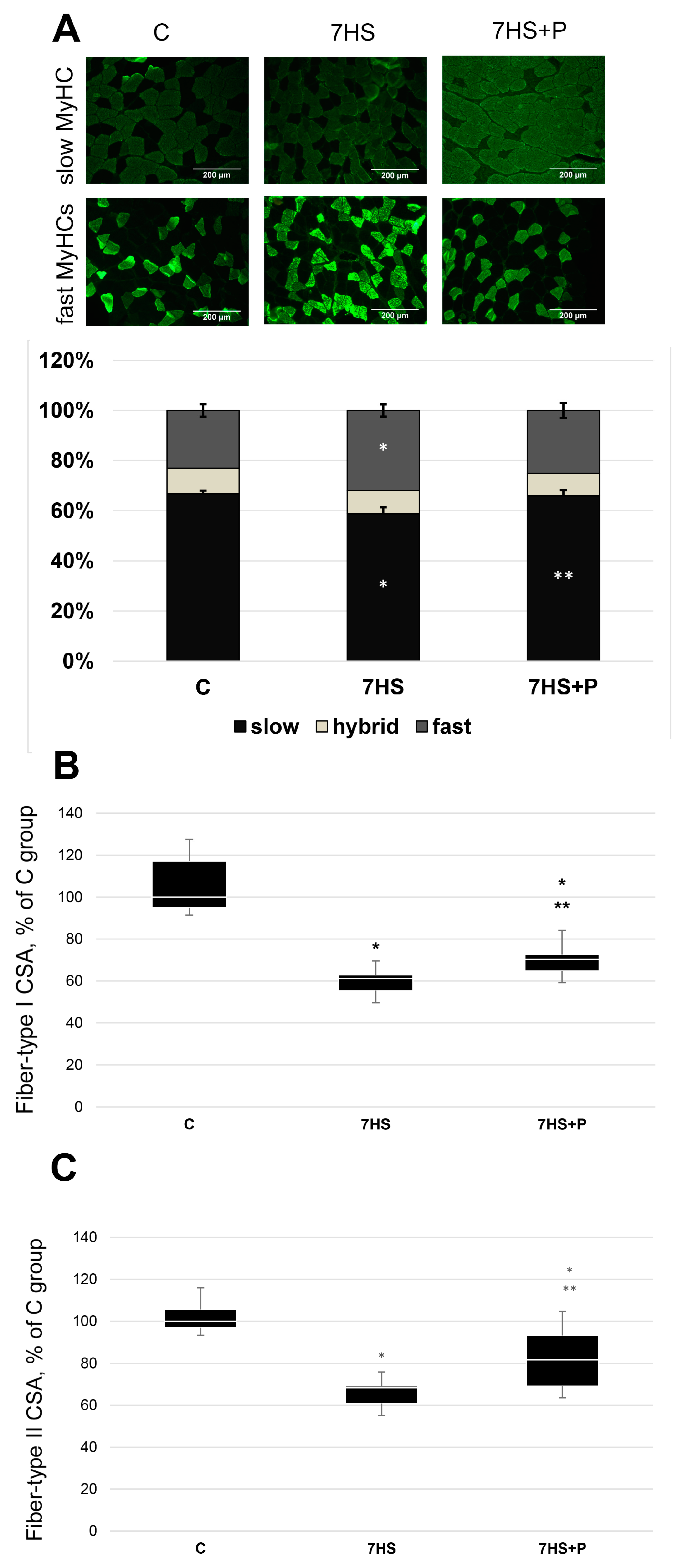
2.8. MyHCs Immunostaining
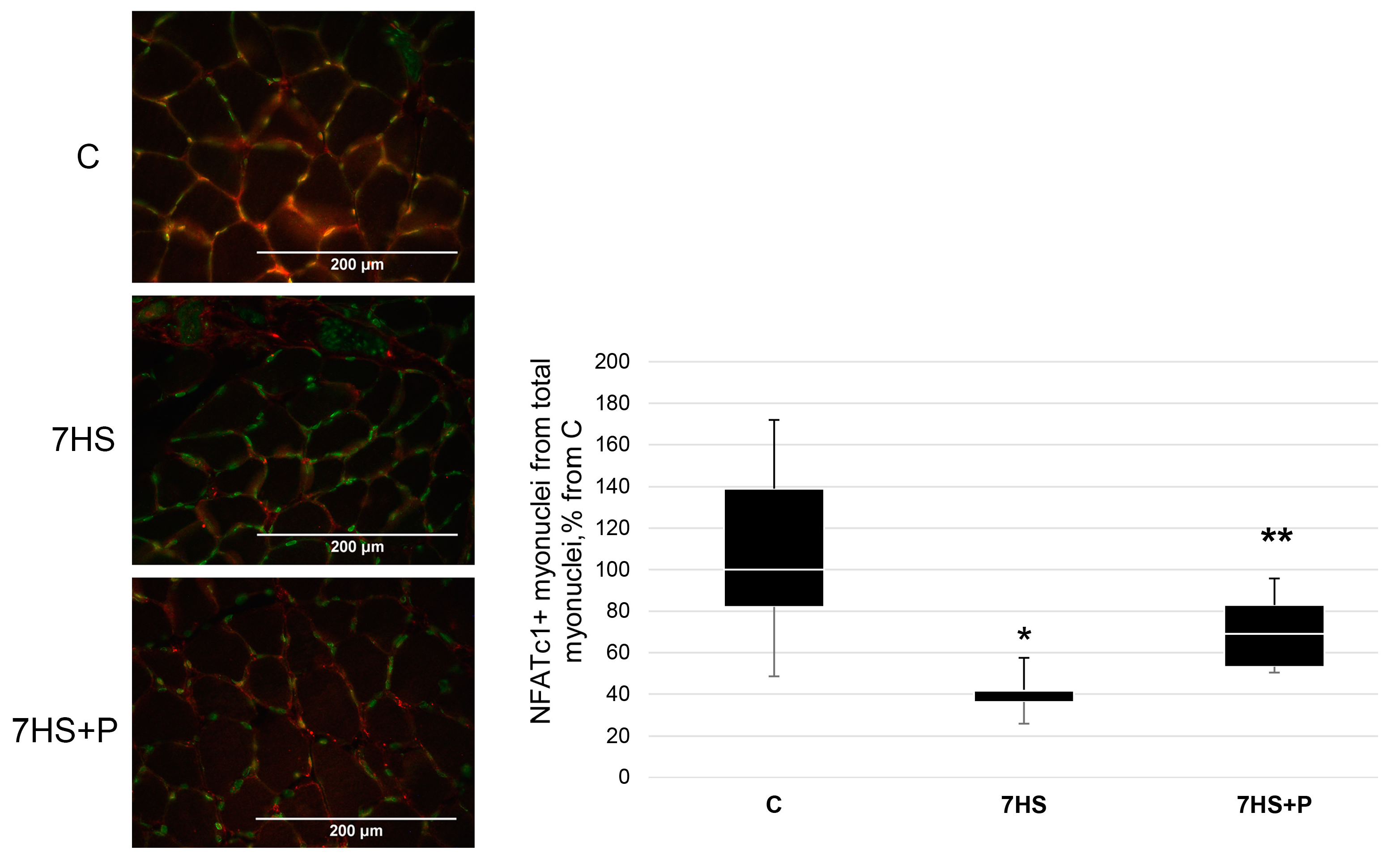
2.9. NFATc1 Detection in Muscle Nuclei
2.10. Statistical Analysis
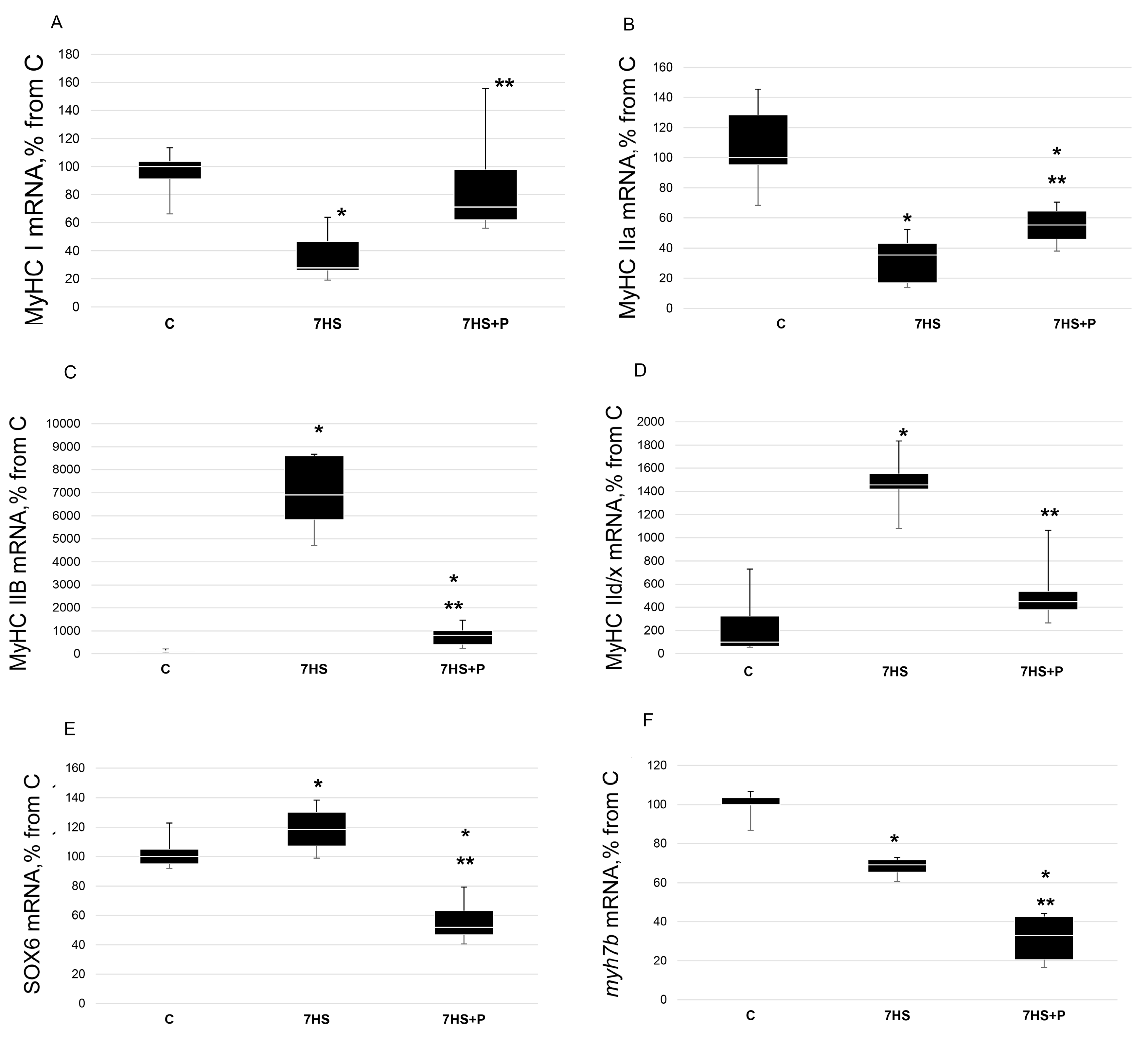
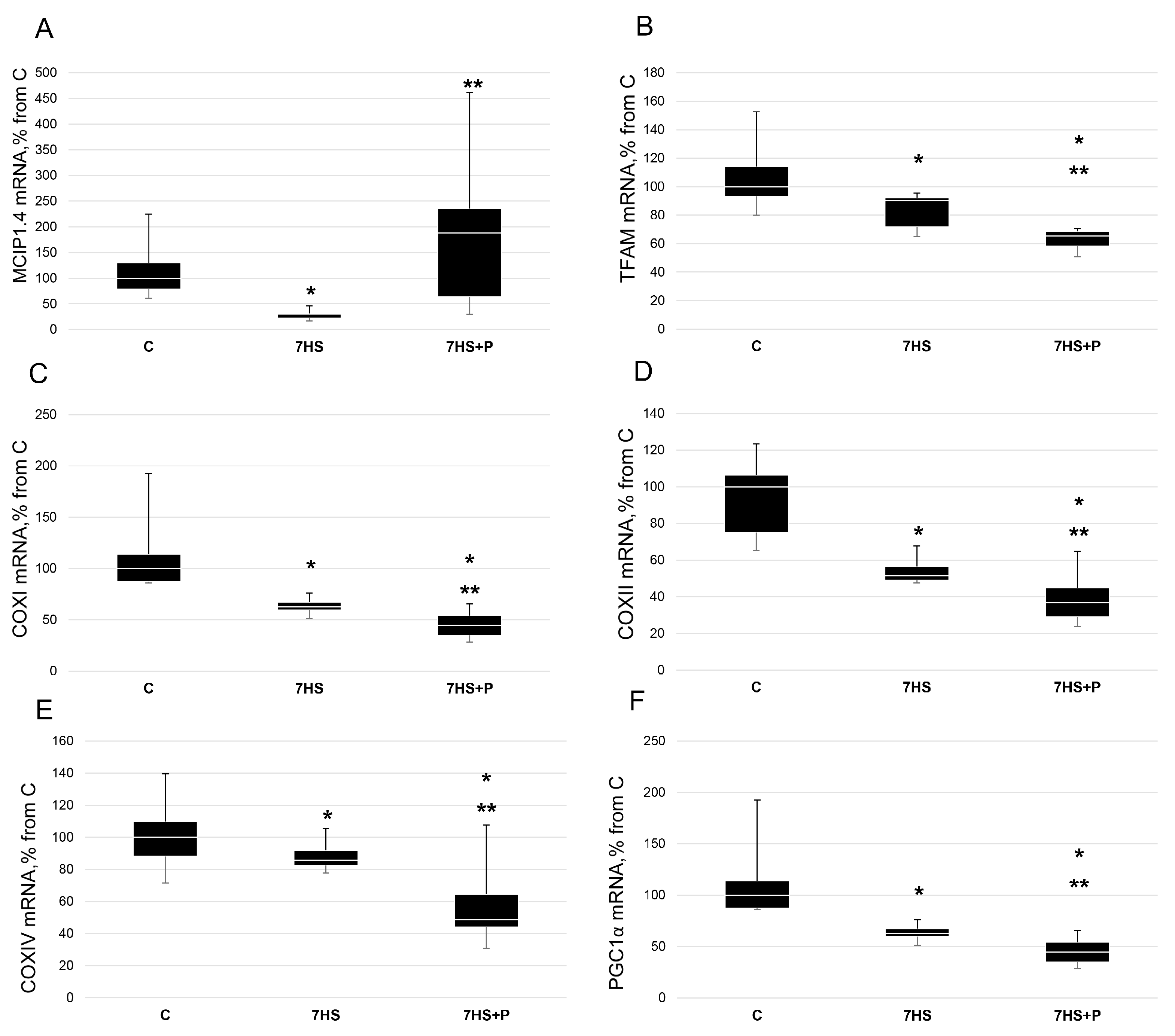
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shenkman, B.S. How Postural Muscle Senses Disuse? Early Signs and Signals. Int. J. Mol. Sci. 2020, 21, 5037. [Google Scholar] [CrossRef] [PubMed]
- Alford, E.K.; Roy, R.R.; Hodgson, J.A.; Edgerton, V.R. Electromyography of rat soleus, medical gastrocnemius, and tibialis anterior during hind limb suspension. Exp. Neurol. 1987, 96, 635–649. [Google Scholar] [CrossRef] [PubMed]
- De-Doncker, L.; Kasri, M.; Picquet, F.; Falempin, M. Physiologically adaptive changes of the L5 afferent neurogram and of the rat soleus EMG activity during 14 days of hindlimb unloading and recovery. J. Exp. Biol. 2005, 208 Pt 24, 4585–4592. [Google Scholar] [CrossRef]
- Kawano, F.; Ishihara, A.; Stevens, J.L.; Wang, X.D.; Ohshima, S.; Horisaka, M.; Maeda, Y.; Nonaka, I.; Ohira, Y. Tension- and afferent input-associated responses of neuromuscular system of rats to hindlimb unloading and/or tenotomy. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R76–R86. [Google Scholar] [CrossRef] [PubMed]
- Boulenguez, P.; Liabeuf, S.; Bos, R.; Bras, H.; Jean-Xavier, C.; Brocard, C.; Stil, A.; Darbon, P.; Cattaert, D.; Delpire, E.; et al. Down-regulation of the potassium-chloride cotransporter KCC2 contributes to spasticity after spinal cord injury. Nat. Med. 2010, 16, 302–307. [Google Scholar] [CrossRef]
- Liabeuf, S.; Stuhl-Gourmand, L.; Gackière, F.; Mancuso, R.; Brualla, I.S.; Marino, P.; Brocard, F.; Vinay, L. Prochlorperazine Increases KCC2 Function and Reduces Spasticity after Spinal Cord Injury. J. Neurotrauma 2017, 34, 3397–3406. [Google Scholar] [CrossRef]
- Kalashnikov, V.E.; Tyganov, S.A.; Turtikova, O.V.; Kalashnikova, E.P.; Glazova, M.V.; Mirzoev, T.M.; Shenkman, B.S. Prochlorperazine Withdraws the Delayed Onset Tonic Activity of Unloaded Rat Soleus Muscle: A Pilot Study. Life 2021, 11, 1161. [Google Scholar] [CrossRef]
- Morey-Holton, E.R.; Globus, R.K. Hindlimb unloading rodent model: Technical aspects. J. Appl. Physiol. 2002, 92, 1367–1377. [Google Scholar] [CrossRef]
- Sharlo, K.A.; Paramonova, I.I.; Lvova, I.D.; Vilchinskaya, N.A.; Bugrova, A.E.; Shevchenko, T.F.; Kalamkarov, G.R.; Shenkman, B.S. NO-Dependent Mechanisms of Myosin Heavy Chain Transcription Regulation in Rat Soleus Muscle after 7-Days Hindlimb Unloading. Front. Physiol. 2020, 11, 814. [Google Scholar] [CrossRef]
- Zaripova, K.A.; Kalashnikova, E.P.; Belova, S.P.; Kostrominova, T.Y.; Shenkman, B.S.; Nemirovskaya, T.L. Role of Pannexin 1 ATP-Permeable Channels in the Regulation of Signaling Pathways during Skeletal Muscle Unloading. Int. J. Mol. Sci. 2021, 22, 10444. [Google Scholar] [CrossRef]
- Sharlo, K.A.; Paramonova, I.I.; Lvova, I.D.; Mochalova, E.P.; Kalashnikov, V.E.; Vilchinskaya, N.A.; Tyganov, S.A.; Konstantinova, T.S.; Shevchenko, T.F.; Kalamkarov, G.R.; et al. Plantar Mechanical Stimulation Maintains Slow Myosin Expression in Disused Rat Soleus Muscle via NO-Dependent Signaling. Int. J. Mol. Sci. 2021, 22, 1372. [Google Scholar] [CrossRef] [PubMed]
- Sharlo, K.; Paramonova, I.; Turtikova, O.; Tyganov, S.; Shenkman, B. Plantar mechanical stimulation prevents calcineurin-NFATc1 inactivation and slow-to-fast fiber type shift in rat soleus muscle under hindlimb unloading. J. Appl. Physiol. 2019, 126, 1769–1781. [Google Scholar] [CrossRef] [PubMed]
- Winje, I.M.; Bengtsen, M.; Eftestøl, E.; Juvkam, I.; Bruusgaard, J.C.; Gundersen, K. Specific labelling of myonuclei by an antibody against pericentriolar material 1 on skeletal muscle tissue sections. Acta Physiol. 2018, 223, e13034. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.B. Role of nitric oxide in skeletal muscle: Synthesis, distribution and functional importance. Acta Physiol. Scand. 1998, 162, 401–409. [Google Scholar] [CrossRef]
- Michaelson, L.P.; Iler, C.; Ward, C.W. ROS and RNS signaling in skeletal muscle: Critical signals and therapeutic targets. Annu. Rev. Nurs. Res. 2013, 31, 367–387. [Google Scholar] [CrossRef]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef]
- McCullagh, K.J.A.; Calabria, E.; Pallafacchina, G.; Ciciliot, S.; Serrano, A.L.; Argentini, C.; Kalhovde, J.M.; Lømo, T.; Schiaffino, S. NFAT is a nerve activity sensor in skeletal muscle and controls activity-dependent myosin switching. Proc. Natl. Acad. Sci. USA 2004, 101, 10590–10595. [Google Scholar] [CrossRef]
- McCarthy, J.J.; Esser, K.A.; Peterson, C.A.; Dupont-Versteegden, E.E. Evidence of MyomiR network regulation of beta-myosin heavy chain gene expression during skeletal muscle atrophy. Physiol. Genom. 2009, 39, 219–226. [Google Scholar] [CrossRef]
- Liu, J.; Liang, X.; Zhou, D.; Lai, L.; Xiao, L.; Liu, L.; Fu, T.; Kong, Y.; Zhou, Q.; Vega, R.B.; et al. Coupling of mitochondrial function and skeletal muscle fiber type by a miR-499/Fnip1/AMPK circuit. EMBO Mol. Med. 2016, 8, 1212–1228. [Google Scholar] [CrossRef]
- Mirzoev, T.M.; Sharlo, K.A.; Shenkman, B.S. The Role of GSK-3β in the Regulation of Protein Turnover, Myosin Phenotype, and Oxidative Capacity in Skeletal Muscle under Disuse Conditions. Int. J. Mol. Sci. 2021, 22, 5081. [Google Scholar] [CrossRef]
- Theeuwes, W.F.; Gosker, H.R.; Schols, A.; Langen, R.C.J.; Remels, A.H.V. Regulation of PGC-1alpha expression by a GSK-3beta-TFEB signaling axis in skeletal muscle. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118610. [Google Scholar] [CrossRef] [PubMed]
- Shenkman, B.S. From Slow to Fast: Hypogravity-Induced Remodeling of Muscle Fiber Myosin Phenotype. Acta Nat. 2016, 8, 47–59. [Google Scholar] [CrossRef]
- Bogdanis, G.C. Effects of Physical Activity and Inactivity on Muscle Fatigue. Front. Physiol. 2012, 3, 142. [Google Scholar] [CrossRef] [PubMed]
- Vilchinskaya, N.A.; Krivoi, I.I.; Shenkman, B.S. AMP-Activated Protein Kinase as a Key Trigger for the Disuse-Induced Skeletal Muscle Remodeling. Int. J. Mol. Sci. 2018, 19, 3558. [Google Scholar] [CrossRef]
- Matoba, H.; Wakatuki, T.; Ohira, Y. β-Guanidinopropionic acid suppresses suspension-induced changes in myosin expression in rat skeletal muscle. Med. Sci. Sports Exerc. 1993, 25, 157. [Google Scholar] [CrossRef]
- Sharlo, K.A.; Mochalova, E.P.; Belova, S.P.; Lvova, I.D.; Nemirovskaya, T.L.; Shenkman, B.S. The role of MAP-kinase p38 in the m. soleus slow myosin mRNA transcription regulation during short-term functional unloading. Arch. Biochem. Biophys. 2020, 695, 108622. [Google Scholar] [CrossRef]
- Paramonova, I.I.; Vilchinskaya, N.A.; Shenkman, B.S. HDAC4 Is Indispensable for Reduced Slow Myosin Expression at the Early Stage of Hindlimb Unloading in Rat Soleus Muscle. Pharmaceuticals 2021, 14, 1167. [Google Scholar] [CrossRef]
- Gupta, R.C.; Misulis, K.E.; Dettbarn, W.-D. Activity dependent characteristics of fast and slow muscle: Biochemical and histochemical considerations. Neurochem. Res. 1989, 14, 647–655. [Google Scholar] [CrossRef]
- Vilchinskaya, N.A.; Mochalova, E.P.; Nemirovskaya, T.L.; Mirzoev, T.M.; Turtikova, O.V.; Shenkman, B.S. Rapid decline in MyHC I(beta) mRNA expression in rat soleus during hindlimb unloading is associated with AMPK dephosphorylation. J. Physiol. 2017, 595, 7123–7134. [Google Scholar] [CrossRef]
- Drenning, J.A.; Lira, V.A.; Simmons, C.G.; Soltow, Q.A.; Sellman, J.E.; Criswell, D.S. Nitric oxide facilitates NFAT-dependent transcription in mouse myotubes. Am. J. Physiol. Cell Physiol. 2008, 294, C1088–C1095. [Google Scholar] [CrossRef]
- Liang, Q.; Bueno, O.F.; Wilkins, B.J.; Kuan, C.Y.; Xia, Y.; Molkentin, J.D. c-Jun N-terminal kinases (JNK) antagonize cardiac growth through cross-talk with calcineurin-NFAT signaling. EMBO J. 2003, 22, 5079–5089. [Google Scholar] [CrossRef] [PubMed]
- Braz, J.C.; Bueno, O.F.; Liang, Q.; Wilkins, B.J.; Dai, Y.-S.; Parsons, S.; Braunwart, J.; Glascock, B.J.; Klevitsky, R.; Kimball, T.F.; et al. Targeted inhibition of p38 MAPK promotes hypertrophic cardiomyopathy through upregulation of calcineurin-NFAT signaling. J. Clin. Investig. 2003, 111, 1475–1486. [Google Scholar] [CrossRef] [PubMed]
- Powers, S.K.; Duarte, J.; Kavazis, A.N.; Talbert, E.E. Reactive oxygen species are signalling molecules for skeletal muscle adaptation. Exp. Physiol. 2010, 95, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Sharma, R.P. Calcium-Mediated Activation of c-Jun NH2-Terminal Kinase (JNK) and Apoptosis in Response to Cadmium in Murine Macrophages. Toxicol. Sci. 2004, 81, 518–527. [Google Scholar] [CrossRef]
- Wright, D.C.; Geiger, P.C.; Han, D.H.; Jones, T.E.; Holloszy, J.O. Calcium induces increases in peroxisome proliferator-activated receptor gamma coactivator-1alpha and mitochondrial biogenesis by a pathway leading to p38 mitogen-activated protein kinase activation. J. Biol. Chem. 2007, 282, 18793–18799. [Google Scholar] [CrossRef]
- Ingalls, C.P.; Warren, G.L.; Armstrong, R.B. Intracellular Ca2+ transients in mouse soleus muscle after hindlimb unloading and reloading. J. Appl. Physiol. 1999, 87, 386–390. [Google Scholar] [CrossRef]
- Kuczmarski, J.M.; Hord, J.M.; Lee, Y.; Guzzoni, V.; Rodriguez, D.; Lawler, M.S.; Garcia-Villatoro, E.L.; Holly, D.; Ryan, P.; Falcon, K.; et al. Effect of Eukarion-134 on Akt-mTOR signalling in the rat soleus during 7 days of mechanical unloading. Exp. Physiol. 2018, 103, 545–558. [Google Scholar] [CrossRef]
- Zhang, X.; Connelly, J.; Levitan, E.S.; Sun, D.; Wang, J.Q. Calcium/Calmodulin–Dependent Protein Kinase II in Cerebrovascular Diseases. Transl. Stroke Res. 2021, 12, 513–529. [Google Scholar] [CrossRef]
- Bouviere, J.; Fortunato, R.S.; Dupuy, C.; Werneck-De-Castro, J.P.; Carvalho, D.P.; Louzada, R.A. Exercise-Stimulated ROS Sensitive Signaling Pathways in Skeletal Muscle. Antioxidants 2021, 10, 537. [Google Scholar] [CrossRef]
- Pandorf, C.E.; Haddad, F.; Owerkowicz, T.; Carroll, L.P.; Baldwin, K.M.; Adams, G.R. Regulation of myosin heavy chain antisense long noncoding RNA in human vastus lateralis in response to exercise training. Am. J. Physiol. Cell Physiol. 2020, 318, C931–C942. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
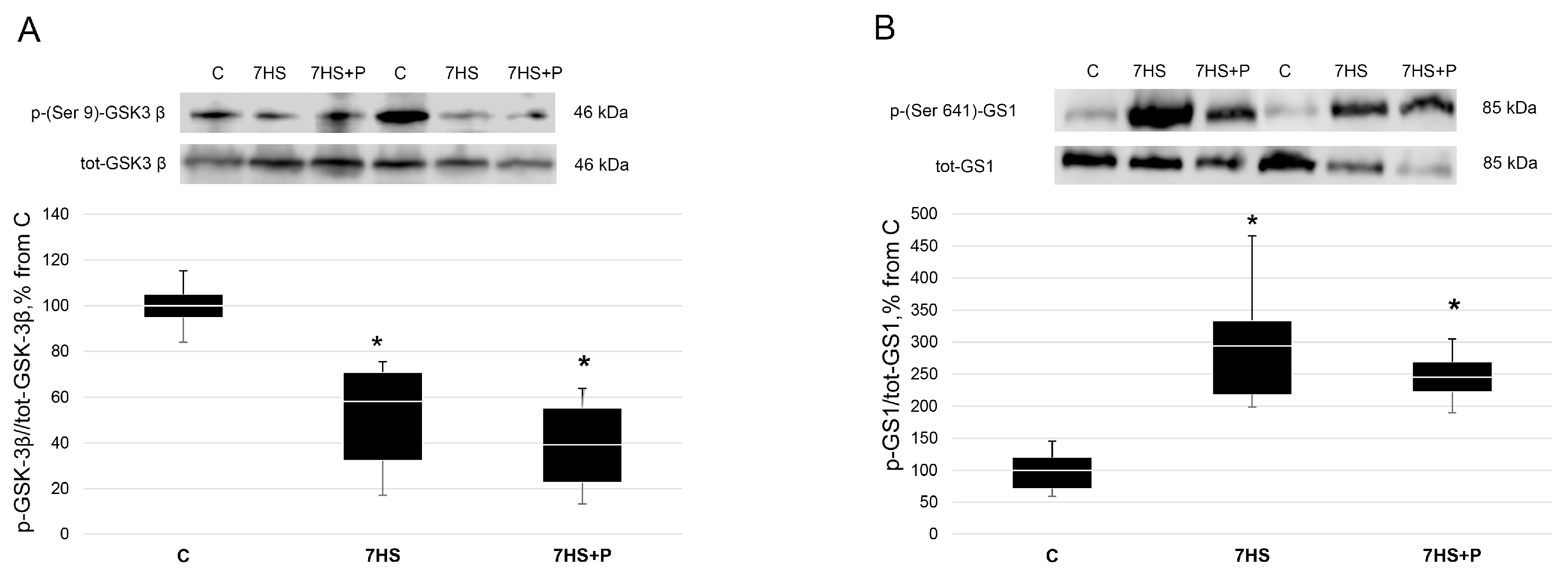{kind=link}
| Group | Body Weight, g | Soleus Weight, mg | Soleus Weight/Body Weight, mg/g |
|---|---|---|---|
| C | 191.4 ± 2.4 | 85.4 ± 3.2 | 44.6 ± 1.5 |
| 7HS | 206.7 ± 6.7 | 72.2 ± 1.9 * | 35.0 ± 0.8 * |
| 7HS + P | 196.0 ± 6.6 | 68.9 ± 3.5 * | 35.6 ± 1.4 * |
| Gene Description | Primer Sequence |
|---|---|
| Myh7 (MyHC I(β)) | 5′-ACAGAGGAAGACAGGAAGAACCTAC-3′ 5′-GGGCTTCACAGGCATCCTTAG-3′ |
| Myh2 (MyHC IIa) | 5′-TATCCTCAGGCTTCAAGATTTG-3′ 5′-TAAATAGAATCACATGGGGACA-3′ |
| Myh4 (MyHC IIb) | 5′-CTGAGGAACAATCCAACGTC-3′ 5′-TTGTGTGATTTCTTCTGTCACCT-3′ |
| Myh1 (MyHC IId/x) | 5′-CGCGAGGTTCACACCAAA-3′ 5′-TCCCAAAGTCGTAAGTACAAAATGG-3′ |
| PGC1alpha | 5′-GTGCAGCCAAGACTCTGTATGG-3′ 5′-GTCCAGGTCATTCACATCAAGTTC-3′ |
| SOX6 | 5′-TCAAAGGCGATTTACCAGTGAC-3′ 5′-TTGTTGTGCATTATGGGGTGC-3′ |
| Rcan1 (MCIP1.4) | 5′-CCGTTGGCTGGAAACAAG-3′ 5′-GGTCACTCTCACACACGTGG-3′ |
| RPL19 | 5′- GTACCCTTCCTCTTCCCTATGC-3′ 5′- CAATGCCAACTCTCGTCAACAG-3′ |
| COXI | 5′-ATTGGAGGCTTCGGGAACTG-3′ 5′-AGATAGAAGACACCCCGGCT-3′ |
| COXII | 5′-ATTGGAGGCTTCGGGAACTG-3′ 5′-AGATAGAAGACACCCCGGCT-3′ |
| COXIV | 5′-TGGGAGTGTTGTGAAGAGTGA-3′ 5′-GCAGTGAAGCCGATGAAGAAC-3′ |
| TFAM | 5′-CGCCTGTCAGCCTTATCTGTA-3′ 5′-TGCATCTGGGTGTTTAGCTTA-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharlo, K.A.; Lvova, I.D.; Tyganov, S.A.; Sergeeva, K.V.; Kalashnikov, V.Y.; Kalashnikova, E.P.; Mirzoev, T.M.; Kalamkarov, G.R.; Shevchenko, T.F.; Shenkman, B.S. A Prochlorperazine-Induced Decrease in Autonomous Muscle Activity during Hindlimb Unloading Is Accompanied by Preserved Slow Myosin mRNA Expression. Curr. Issues Mol. Biol. 2023, 45, 5613-5630. https://doi.org/10.3390/cimb45070354
Sharlo KA, Lvova ID, Tyganov SA, Sergeeva KV, Kalashnikov VY, Kalashnikova EP, Mirzoev TM, Kalamkarov GR, Shevchenko TF, Shenkman BS. A Prochlorperazine-Induced Decrease in Autonomous Muscle Activity during Hindlimb Unloading Is Accompanied by Preserved Slow Myosin mRNA Expression. Current Issues in Molecular Biology. 2023; 45(7):5613-5630. https://doi.org/10.3390/cimb45070354
Chicago/Turabian StyleSharlo, Kristina A., Irina D. Lvova, Sergey A. Tyganov, Ksenia V. Sergeeva, Vitaly Y. Kalashnikov, Ekaterina P. Kalashnikova, Timur M. Mirzoev, Grigoriy R. Kalamkarov, Tatiana F. Shevchenko, and Boris S. Shenkman. 2023. "A Prochlorperazine-Induced Decrease in Autonomous Muscle Activity during Hindlimb Unloading Is Accompanied by Preserved Slow Myosin mRNA Expression" Current Issues in Molecular Biology 45, no. 7: 5613-5630. https://doi.org/10.3390/cimb45070354
APA StyleSharlo, K. A., Lvova, I. D., Tyganov, S. A., Sergeeva, K. V., Kalashnikov, V. Y., Kalashnikova, E. P., Mirzoev, T. M., Kalamkarov, G. R., Shevchenko, T. F., & Shenkman, B. S. (2023). A Prochlorperazine-Induced Decrease in Autonomous Muscle Activity during Hindlimb Unloading Is Accompanied by Preserved Slow Myosin mRNA Expression. Current Issues in Molecular Biology, 45(7), 5613-5630. https://doi.org/10.3390/cimb45070354