

Oxidative Stress in Assisted Reproductive Techniques, with a Focus on an Underestimated Risk Factor

 ,
, 

{kind=link}
{kind=link}
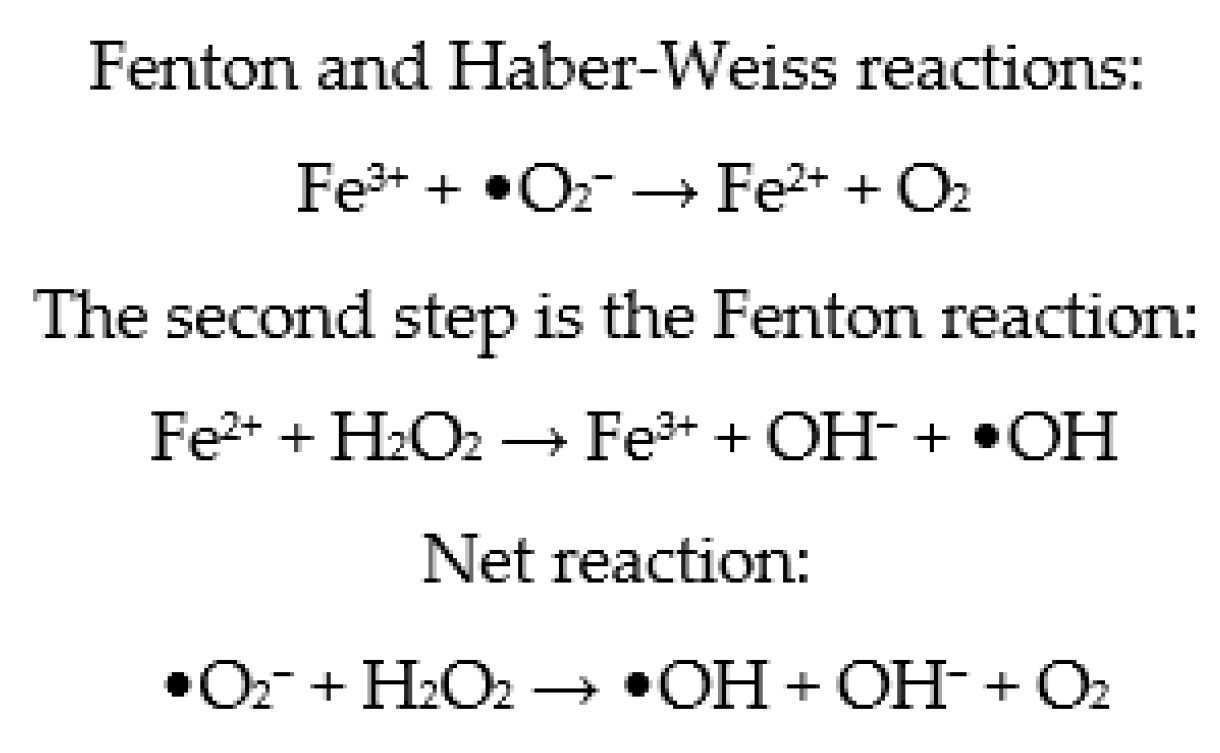{kind=link}
Abstract
1. Introduction
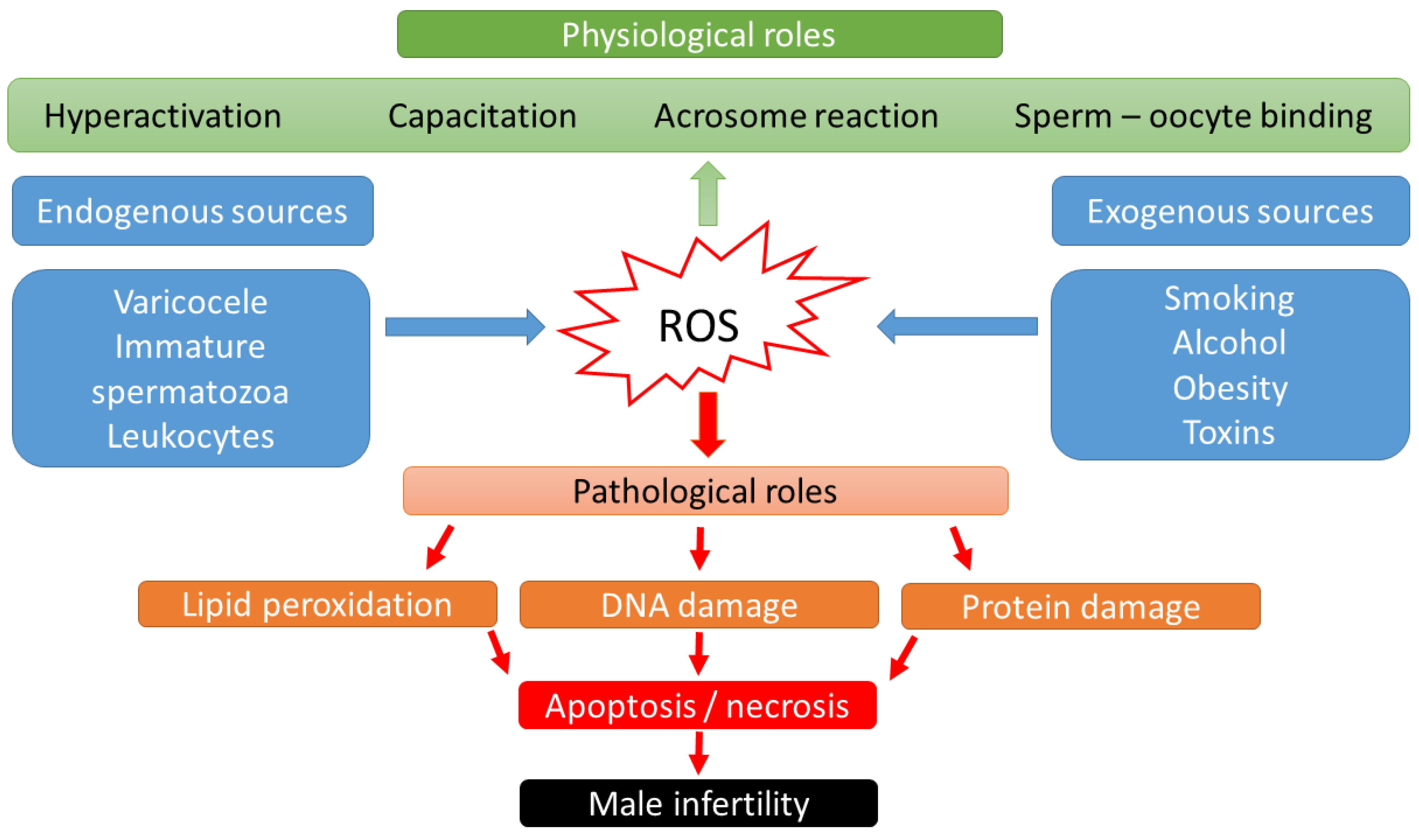
2. Effect of Oxidative Stress on the Reproductive Tract of Males
2.1. Sources of ROS in Sperm
2.2. Physiological Role of ROS in Sperm
2.3. Pathological Role of ROS in Sperm
2.4. Effects of OS on Sperm Functions
2.5. Methods Used to Counteract OS Effects
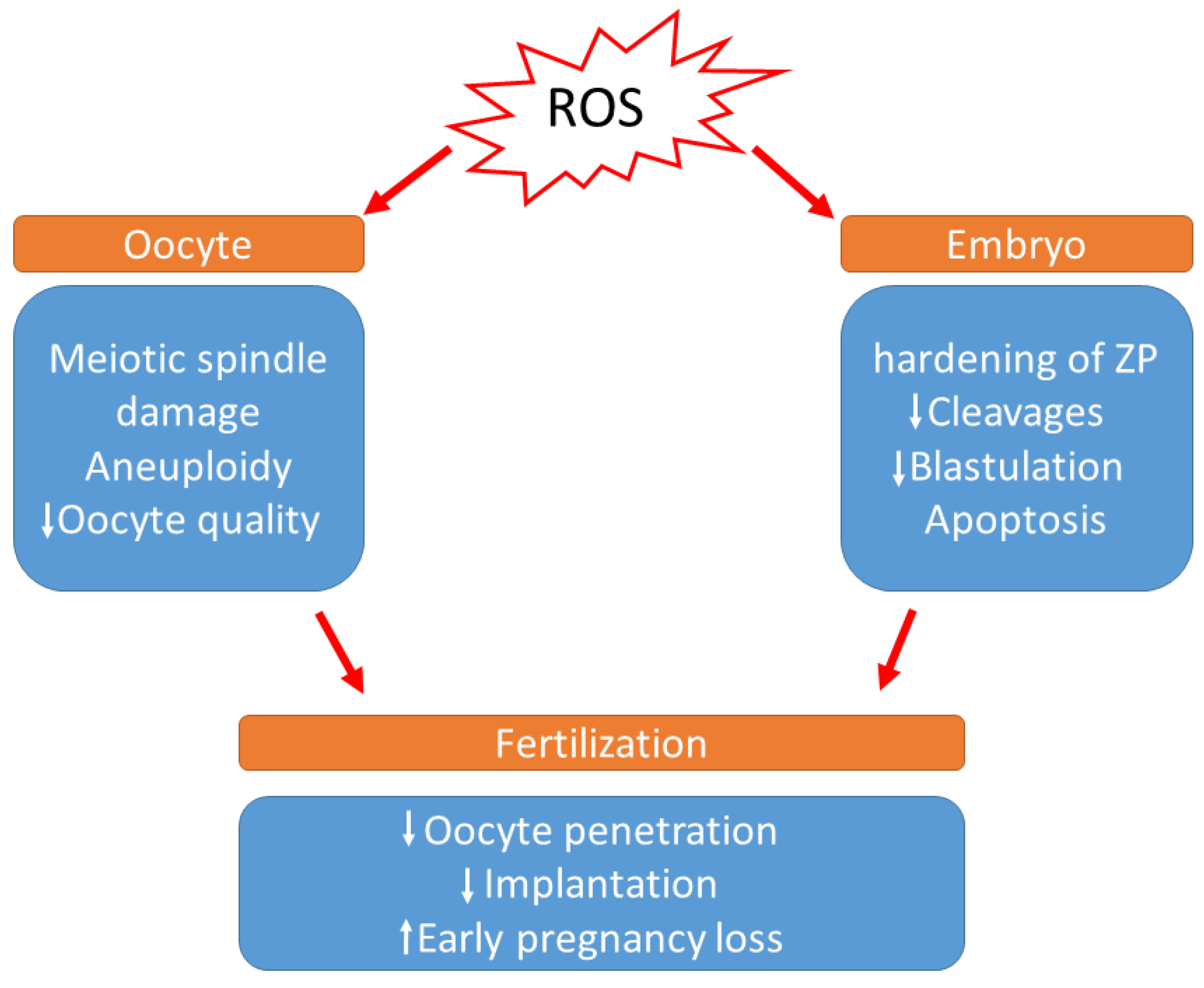
3. Effect of Oxidative Stress on the Reproductive Tract of Females
3.1. Physiological Roles of OS in Females Reproductive Tract
3.2. Pathological Roles of OS in Female Reproductive Tract
4. Oxidative Stress in Pre-Embryos and Their Surroundings
4.1. Sources of ROS during ART
4.2. The Effect of Light Exposure of Gametes and Embryos
4.3. OS in the Embryo Culture Medium
4.4. Methods Used to Counteract OS Effects
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Zarbakhsh, S. Effect of antioxidants on preimplantation embryo development in vitro: A review. Zygote 2021, 29, 179–193. [Google Scholar] [CrossRef]
- Bell, E.L.; Emerling, B.M.; Chandel, N.S. Mitochondrial regulation of oxygen sensing. Mitochondrion 2005, 5, 322–332. [Google Scholar] [CrossRef]
- Bell, E.L.; Chandel, N.S. Mitochondrial oxygen sensing: Regulation of hypoxia-inducible factor by mitochondrial generated reactive oxygen species. Essays Biochem. 2007, 43, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Van Blerkom, J. Mitochondria as regulatory forces in oocytes, preimplantation embryos and stem cells. Reprod. Biomed. Online 2008, 16, 553–569. [Google Scholar] [CrossRef]
- De Giusti, V.C.; Caldiz, C.I.; Ennis, I.L.; Pérez, N.G.; Cingolani, H.E.; Aiello, E.A. Mitochondrial reactive oxygen species (ROS) as signaling molecules of intracellular pathways triggered by the cardiac renin-angiotensin II-aldosterone system (RAAS). Front. Physiol. 2013, 4, 126. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Xu, Y.; Li, Y.; Jin, Q.; Sun, J.; Zhiqiang, E.; Gao, Q. Supplementation of kaempferol to in vitro maturation medium regulates oxidative stress and enhances subsequent embryonic development in vitro. Zygote 2020, 28, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Scialò, F.; Fernández-Ayala, D.J.; Sanz, A. Role of Mitochondrial Reverse Electron Transport in ROS Signaling: Potential Roles in Health and Disease. Front. Physiol. 2017, 8, 428. [Google Scholar] [CrossRef]
- Attaran, M.; Pasqualotto, E.; Falcone, T.; Goldberg, J.M.; Miller, K.F.; Agarwal, A.; Sharma, R. The effect of follicular fluid reactive oxygen species on the outcome of in vitro fertilization. Int. J. Fertil. Women’s Med. 2000, 45, 314–320. [Google Scholar]
- Szczepańska, M.; Koźlik, J.; Skrzypczak, J.; Mikołajczyk, M. Oxidative stress may be a piece in the endometriosis puzzle. Fertil. Steril. 2003, 79, 1288–1293. [Google Scholar] [CrossRef]
- Van Langendonckt, A.; Casanas-Roux, F.; Donnez, J. Oxidative stress and peritoneal endometriosis. Fertil. Steril. 2002, 77, 861–870. [Google Scholar] [CrossRef] [PubMed]
- Pierce, J.D.; Cackler, A.B.; Arnett, M.G. Why should you care about free radicals? RN 2004, 67, 38–42. [Google Scholar]
- Iwayama, K.; Kusakabe, A.; Ohtsu, K.; Nawano, T.; Tatsunami, R.; Ohtaki, K.-I.; Tampo, Y.; Hayase, N. Long-term treatment of clarithromycin at a low concentration improves hydrogen peroxide-induced oxidant/antioxidant imbalance in human small airway epithelial cells by increasing Nrf2 mRNA expression. BMC Pharmacol. Toxicol. 2017, 18, 15. [Google Scholar] [CrossRef] [PubMed]
- Du Plessis, S.S.; Agarwal, A.; Mohanty, G.; van der Linde, M. Oxidative phosphorylation versus glycolysis: What fuel do spermatozoa use? Asian J. Androl. 2015, 17, 230–235. [Google Scholar] [CrossRef]
- Takeshima, T.; Usui, K.; Mori, K.; Asai, T.; Yasuda, K.; Kuroda, S.; Yumura, Y. Oxidative stress and male infertility. Reprod. Med. Biol. 2020, 20, 41–52. [Google Scholar] [CrossRef]
- Carrell, D.T.; Liu, L.; Peterson, C.M.; Jones, K.P.; Hatasaka, H.H.; Erickson, L.; Campbell, B. Sperm DNA fragmentation is increased in couples with unexplained recurrent pregnancy loss. Arch. Androl. 2003, 49, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.E.M.; Aitken, R.J. DNA damage to spermatozoa has impacts on fertilization and pregnancy. Cell Tissue Res. 2005, 322, 33–41. [Google Scholar] [CrossRef]
- Khan, A.U.; Wilson, T. Reactive oxygen species as cellular messengers. Chem. Biol. 1995, 2, 437–445. [Google Scholar] [CrossRef]
- Thompson, A.; Agarwal, A.; du Plessis, S.S. Physiological Role of Reactive Oxygen Species in Sperm Function: A Review. In Antioxidants in Male Infertility: A Guide for Clinicians and Researchers; Parekatil, S.J., Agarwal, A., Eds.; Springer Science+Business Media: New York, NY, USA, 2013; pp. 69–89. [Google Scholar] [CrossRef]
- Du Plessis, S.S.; Agarwal, A.; Halabi, J.; Tvrda, E. Contemporary evidence on the physiological role of reactive oxygen species in human sperm function. J. Assist. Reprod. Genet. 2015, 32, 509–520. [Google Scholar] [CrossRef]
- Griveau, J.F.; Le Lannou, D. Reactive oxygen species and human spermatozoa: Physiology and pathology. Int. J. Androl. 1997, 20, 61–69. [Google Scholar] [CrossRef]
- Dutta, S.; Henkel, R.; Sengupta, P.; Agarwal, A. Physiological role of ROS in sperm function. In Male Infertility: Contemporary Clinical Approaches, Andrology, ART and Antioxidants; Springer: New York, NY, USA, 2020; pp. 337–345. [Google Scholar]
- Evans, E.P.P.; Scholten, J.T.M.; Mzyk, A.; Reyes-San-Martin, C.; Llumbet, A.E.; Hamoh, T.; Arts, E.G.J.M.; Schirhagl, R.; Cantineau, A.E. Male subfertility and oxidative stress. Redox Biol. 2021, 46, 102071. [Google Scholar] [CrossRef] [PubMed]
- Durairajanayagam, D. Physiological Role of Reactive Oxygen Species in Male Reproduction. In Oxidants, Antioxidants and Impact of the Oxidative Status in Male Reproduction; Henkel, R., Samanta, L., Agarwal, A., Eds.; Academic Press: Cambridge, MA, USA, 2019; Chapter 1.8; pp. 65–78. [Google Scholar]
- de Lamirande, E. Reactive oxygen species and sperm physiology. Rev. Reprod. 1997, 2, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Makker, K.; Agarwal, A.; Sharma, R. Oxidative stress & male infertility. Indian J. Med. Res. 2009, 129, 357–367. [Google Scholar] [PubMed]
- Kumar, N.; Singh, A.K. Reactive oxygen species in seminal plasma as a cause of male infertility. J. Gynecol. Obstet. Hum. Reprod. 2018, 47, 565–572. [Google Scholar] [CrossRef]
- Cooper, T.G. The epididymis, cytoplasmic droplets and male fertility. Asian J. Androl. 2010, 13, 130–138. [Google Scholar] [CrossRef]
- Aziz, N.; Saleh, R.A.; Sharma, R.K.; Lewis-Jones, I.; Esfandiari, N.; Thomas, A.J., Jr.; Agarwal, A. Novel association between sperm reactive oxygen species production, sperm morphological defects, and the sperm deformity index. Fertil. Steril. 2004, 81, 349–354. [Google Scholar] [CrossRef]
- Plante, M.; de Lamirande, E.; Gagnon, C. Reactive oxygen species released by activated neutrophils, but not by deficient spermatozoa, are sufficient to affect normal sperm motility. Fertil. Steril. 1994, 62, 387–393. [Google Scholar] [CrossRef]
- Agarwal, A.; Rosas, I.M.; Anagnostopoulou, C.; Cannarella, R.; Boitrelle, F.; Munoz, L.V.; Finelli, R.; Durairajanayagam, D.; Henkel, R.; Saleh, R. Oxidative Stress and Assisted Reproduction: A Comprehensive Review of Its Pathophysiological Role and Strategies for Optimizing Embryo Culture Environment. Antioxidants 2022, 11, 477. [Google Scholar] [CrossRef]
- Aitken, R.J.; Finnie, J.M.; Muscio, L.; Whiting, S.; Connaughton, H.S.; Kuczera, L.; Rothkirch, T.B.; De Iuliis, G.N. Potential importance of transition metals in the induction of DNA damage by sperm preparation medium. Hum. Reprod. 2014, 29, 2136–2147. [Google Scholar] [CrossRef]
- Muratori, M.; Tarozzi, N.; Carpentiero, F.; Danti, S.; Perrone, F.M.; Cambi, M.; Casini, A.; Azzari, C.; Boni, L.; Maggi, M.; et al. Sperm selection with density gradient centrifugation and swim up: Effect on DNA fragmentation in viable spermatozoa. Sci. Rep. 2019, 9, 7492. [Google Scholar] [CrossRef]
- Barbonetti, A.; Castellini, C.; Di Giammarco, N.; Santilli, G.; Francavilla, S.; Francavilla, F. In vitro exposure of human spermatozoa to bisphenol A induces pro-oxidative/apoptotic mitochondrial dysfunction. Reprod. Toxicol. 2016, 66, 61–67. [Google Scholar] [CrossRef]
- Kotwicka, M.; Skibinska, I.; Jendraszak, M.; Jedrzejczak, P. 17beta-estradiol modifies human spermatozoa mitochondrial function in vitro. Reprod. Biol. Endocrinol. 2016, 14, 50. [Google Scholar]
- Gualtieri, R.; Kalthur, G.; Barbato, V.; Longobardi, S.; Di Rella, F.; Adiga, S.K.; Talevi, R. Sperm Oxidative Stress during In Vitro Manipulation and Its Effects on Sperm Function and Embryo Development. Antioxidants 2021, 10, 1025. [Google Scholar] [CrossRef]
- Alvarez, J.G.; Storey, B.T. Differential incorporation of fatty acids into and peroxidative loss of fatty acids from phospholipids of human spermatozoa. Mol. Reprod. Dev. 1995, 42, 334–346. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Saleh, R.A.; Bedaiwy, M.A. Role of reactive oxygen species in the pathophysiology of human reproduction. Fertil. Steril. 2003, 79, 829–843. [Google Scholar] [CrossRef] [PubMed]
- De Lamirande, E.; Gagnon, C. Reactive oxygen species and human spermatozoa. I. Effects on the motility of intact spermatozoa and on sperm axonemes. J. Androl. 1992, 13, 368–378. [Google Scholar]
- Twigg, J.P.; Irvine, D.S.; Aitken, R.J. Oxidative damage to DNA in human spermatozoa does not preclude pronucleus formation at intracytoplasmic sperm injection. Hum. Reprod. 1998, 13, 1864–1871. [Google Scholar] [CrossRef]
- Kemal Duru, N.; Morshedi, M.; Oehninger, S. Effects of hydrogen peroxide on DNA and plasma membrane integrity of human spermatozoa. Fertil. Steril. 2000, 74, 1200–1207. [Google Scholar] [CrossRef]
- Aitken, R.; Krausz, C. Oxidative stress, DNA damage and the Y chromosome. Reproduction 2001, 122, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Mahfouz, R.; Sharma, R.; Thiyagarajan, A.; Kale, V.; Gupta, S.; Sabanegh, E.; Agarwal, A. Semen characteristics and sperm DNA fragmentation in infertile men with low and high levels of seminal reactive oxygen species. Fertil. Steril. 2010, 94, 2141–2146. [Google Scholar] [CrossRef]
- Bauer, N.C.; Corbett, A.H.; Doetsch, P.W. The current state of eukaryotic DNA base damage and repair. Nucleic Acids Res. 2015, 43, 10083–10101. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.B.; Dun, M.D.; Smith, N.D.; Curry, B.J.; Connaughton, H.S.; Aitken, R.J. The presence of a truncated base excision repair pathway in human spermatozoa, Mediated by OGG1. J. Cell Sci. 2013, 126, 1488–1497. [Google Scholar] [CrossRef]
- Martin-Hidalgo, D.; Bragado, M.J.; Batista, A.R.; Oliveira, P.F.; Alves, M.G. Antioxidants and Male Fertility: From Molecular Studies to Clinical Evidence. Antioxidants 2019, 8, 89. [Google Scholar] [CrossRef]
- Humaidan, P.; Haahr, T.; Povlsen, B.B.; Kofod, L.; Laursen, R.J.; Alsbjerg, B.; Elbaek, H.O.; Esteves, S.C. The combined effect of lifestyle intervention and antioxidant therapy on sperm DNA fragmentation and seminal oxidative stress in IVF patients: A pilot study. Int. Braz. J. Urol. 2022, 48, 131–156. [Google Scholar] [CrossRef] [PubMed]
- Panner Selvam, M.K.; Agarwal, A.; Henkel, R.; Finelli, R.; Robert, K.A.; Iovine, C.; Baskaran, S. The effect of oxidative and reductive stress on semen parameters and functions of physiologically normal human spermatozoa. Free Radic. Biol. Med. 2020, 152, 375–385. [Google Scholar] [CrossRef]
- Wang, L.; Tang, J.; Wang, L.; Tan, F.; Song, H.; Zhou, J.; Li, F. Oxidative stress in oocyte aging and female reproduction. J. Cell. Physiol. 2021, 236, 7966–7983. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Sharma, R.K.; Falcone, T.; Goldberg, J.; Agarwal, A. Importance portance of reactive oxygen species in the peritoneal fluid of women with endometriosis or idiopathic infertility. Fertil. Steril. 1997, 68, 826–830. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.; Sharma, R. Oxidative stress and its implications in female infertility–A clinician’s perspective. Reprod. Biomed. Online 2005, 11, 641–650. [Google Scholar] [CrossRef]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrinol. 2005, 3, 28–48. [Google Scholar] [CrossRef]
- Wall, P.D.; Pressman, E.K.; Woods, J.R. Preterm premature rupture of the membranes and antioxidants: The free radical connection. J. Périnat. Med. 2002, 30, 447–457. [Google Scholar] [CrossRef]
- Myatt, L.; Cui, X. Oxidative stress in the placenta. Histochem. Cell Biol. 2004, 122, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Murphy, A.A.; Arlene, N.S.; Morales, J.; Parthasarathy, S. Lysophosphatidyl Choline, a Chemotactic Factor for Monocytes/T-Lymphocytes Is Elevated in Endometriosis. J. Clin. Endocrinol. Metab. 1998, 83, 2110–2113. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Sugino, N.; Fukaya, T.; Sugiyama, S.; Uda, T.; Takaya, R.; Yajima, A.; Sasano, H. Superoxide dismutase in normal cycling human ovaries: Immunohistochemical localization and characterization. Fertil. Steril. 1999, 72, 720–726. [Google Scholar] [CrossRef] [PubMed]
- Vega, M.; Carrasca, I.; Castillo, T.; Troncoso, J.L.; Videla, I.A.; Devoto, L. Functional luteolysis in response to hydrogen peroxide in human luteal cells. J. Endocrinol. 1995, 147, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.L.; Skepper, J.N.; Jauniaux, E.; Burton, G.J. Changes in concentration, localization and activity of catalase within the human placenta during early gestation. Placenta 1998, 19, 27–34. [Google Scholar] [CrossRef]
- Watson, A.l.; Palmer, M.E.; Jauniaux, E.; Burton, G.J. Variations in expression of copper/zinc superoxide dismutase in villous trophoblast of the human placenta with gestational age. Placenta 1997, 18, 295–299. [Google Scholar] [CrossRef]
- Kaya, C.; Erkan, A.F.; Cengiz, S.D.; Dünder, I.; Demirel, E.; Bilgihan, A. Advanced oxidation protein products are increased in women with polycystic ovary syndrome: Relationship with traditional and nontraditional cardiovascular risk factors in patients with polycystic ovary syndrome. Fertil. Steril. 2009, 92, 1372–1377. [Google Scholar] [CrossRef]
- Amato, G.; Conte, M.; Mazziotti, G.; Lalli, E.; Vitolo, G.; Tucker, A.T.; Bellastella, A.; Carella, C.; Izzo, A. Serum and follicular fluid cytokines in polycystic ovary syndrome during stimulated cycles. Obstet. Gynecol. 2003, 101, 1177–1182. [Google Scholar]
- Rashidi, B.; Haghollahi, F.; Shariat, M.; Zayerii, F. The Effects of Calcium-Vitamin D and Metformin on Polycystic Ovary Syndrome: A Pilot Study. Taiwan. J. Obstet. Gynecol. 2009, 48, 142–147. [Google Scholar] [CrossRef]
- Cuffe, J.S.M.; Holland, O.; Salomon, C.; Rice, G.E.; Perkins, A.V. Review: Placental derived biomarkers of pregnancy disorders. Placenta 2017, 54, 104–110. [Google Scholar] [CrossRef]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, Ö.; Durak, I. Role of Oxidative Stress in Intrauterine Growth Restriction. Gynecol. Obstet. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Wiener-Megnazi, Z.; Vardi, L.; Lissak, A.; Shnizer, S.; Reznick, A.Z.; Ishai, D.; Lahav-Baratz, S.; Shiloh, H.; Koifman, M.; Dirnfeld, M. Oxidative stress indices in follicular fluid as measured by the thermochemiluminescence assay correlate with outcome parameters in in vitro fertilization. Fertil. Steril. 2004, 82, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Shkolnik, K.; Tadmor, A.; Ben-Dor, S.; Nevo, N.; Galiani, D.; Dekel, N. Reactive oxygen species are indispensable in ovulation. Proc. Natl. Acad. Sci. USA 2011, 108, 1462–1467. [Google Scholar] [CrossRef]
- Babuška, V.; Cedíková, M.; Rajdl, D.; Racek, J.; Zech, N.H.; Trefil, L.; Mocková, A.; Ulčová-Gallová, Z.; Novotný, Z.; Králíčková, M. Srovnání vybraných parametrů oxidačního stresu ve folikulární tekutině u žen s poruchou plodnosti a u zdravých fertilních dárkyň oocytů. [Comparison of selective oxidative stress parameters in the follicular fluid of infertile women and healthy fertile oocyte donors]. Ceská Gynekol. Ceská Lékarská Spolecnost. J. Ev. Purkyne 2012, 77, 543–548. [Google Scholar]
- Sasaki, H.; Hamatani, T.; Kamijo, S.; Iwai, M.; Kobanawa, M.; Ogawa, S.; Miyado, K.; Tanaka, M. Impact of Oxidative Stress on Age-Associated Decline in Oocyte Developmental Competence. Front. Endocrinol. 2019, 10, 811. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, K.; Sato, E.F.; Kasahara, E.; Jikumaru, M.; Hiramoto, K.; Tabata, H.; Katsuragi, M.; Odo, S.; Utsumi, K.; Inoue, M. Effect of oxidative stress during repeated ovulation on the structure and functions of the ovary, oocytes, and their mitochondria. Free. Radic. Biol. Med. 2010, 49, 674–681. [Google Scholar] [CrossRef]
- Agarwal, A.; Allamaneni, S.S. Role of free radicals in female reproductive diseases and assisted reproduction. Reprod. Biomed. Online 2004, 9, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Tarin, J.J. Potential effects of age-associated oxidative stress on mammalian oocytes/embryos. Mol. Hum. Reprod. 1996, 2, 717–724. [Google Scholar] [CrossRef]
- Tatone, C.; Carbone, M.; Falone, S.; Aimola, P.; Giardinelli, A.; Caserta, D.; Marci, R.; Pandolfi, A.; Ragnelli, A.; Amicarelli, F. Age-dependent changes in the expression of superoxide dismutases and catalase are associated with ultrastructural modifications in human granulosa cells. Mol. Hum. Reprod. 2006, 12, 655–660. [Google Scholar] [CrossRef]
- Myllynen, P.; Pasanen, M.; Pelkonen, O. Human placenta: A human organ for developmental toxicology research and biomonitoring. Placenta 2005, 26, 361–371. [Google Scholar] [CrossRef]
- Strakovsky, R.S.; Pan, Y.-X. In Utero Oxidative Stress Epigenetically Programs Antioxidant Defense Capacity and Adulthood Diseases. Antioxid. Redox Signal. 2012, 17, 237–253. [Google Scholar] [CrossRef] [PubMed]
- Global Supplement Report. Available online: https://store.newhope.com/products/2019-global-supplement-report2019. (accessed on 28 January 2023).
- Showell, M.G.; Mackenzie-Proctor, R.; Jordan, V.; Hart, R.J. Antioxidants for female sub-fertility. Cochrane Database Syst. Rev. 2020, 8, CD007807. [Google Scholar] [PubMed]
- Elnashar, A. Antioxidants for female infertility: Review of systematic reviews. Eur. J. Obstet. Gynecol. Reprod. Biol. 2019, 234, e84. [Google Scholar] [CrossRef]
- Fang, F.; Ni, K.; Cai, Y.; Shang, J.; Zhang, X.; Xiong, C. Effect of vitamin D supplementa-tion on polycystic ovary syndrome: A systematic review and meta-analysis of randomized controlled trials. Complement. Clin. Pract. 2017, 26, 53–60. [Google Scholar] [CrossRef]
- Arhin, S.K.; Zhao, Y.; Lu, X.; Chetry, M.; Lu, J. Effect of micronutrient supplementation on IVF outcomes: A systematic review of the literature. Reprod. Biomed. Online 2017, 35, 715–722. [Google Scholar] [CrossRef]
- Guérin, P.; El Mouatassim, S.; Ménézo, Y. Oxidative stress and protection against reactive oxygen species in the pre-implantation embryo and its surroundings. Hum. Reprod. Update 2001, 7, 175–189. [Google Scholar] [CrossRef]
- Harvey, A.J.; Kind, K.L.; Thompson, J.G. REDOX regulation of early embryo development. Reproduction 2002, 123, 479–486. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M.; Bedaiwy, M.A.; Banerjee, J.; Alvarez, J.G. Oxidative stress in an assisted reproductive techniques setting. Fertil. Steril. 2006, 86, 503–512. [Google Scholar] [CrossRef]
- Bedaiwy, M.A.; Falcone, T.; Mohamed, M.S.; Aleem, A.A.N.; Sharma, R.K.; Worley, S.E.; Thornton, J.; Agarwal, A. Differential growth of human embryos in vitro: Role of reactive oxygen species. Fertil. Steril. 2004, 82, 593–600. [Google Scholar] [CrossRef]
- Mohammadi, M. Oxidative stress and polycystic ovary syndrome: A brief review. Int. J. Prev. Med. 2019, 10, 86. [Google Scholar] [CrossRef]
- Scutiero, G.; Iannone, P.; Bernardi, G.; Bonaccorsi, G.; Spadaro, S.; Volta, C.A.; Greco, P.; Nappi, L. Oxidative Stress and Endometriosis: A Systematic Review of the Literature. Oxidative Med. Cell. Longev. 2017, 2017, 7265238. [Google Scholar] [CrossRef]
- von Mengden, L.; Klamt, F.; Smitz, J. Redox Biology of Human Cumulus Cells: Basic Concepts, Impact on Oocyte Quality, and Potential Clinical Use. Antioxid. Redox Signal. 2020, 32, 522–535. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Irvine, D.S.; Wu, F.C. Prospective analysis of sperm-oocyte fusion and reactive oxygen species generation as criteria for the diagnosis of infertility. Am. J. Obstet. Gynecol. 1991, 164, 542–551. [Google Scholar] [CrossRef]
- Gianaroli, L.; Magli, M.; Ferraretti, A.P.; Fiorentino, A.; Tosti, E.; Panzella, S.; Dale, B. Reducing the time of sperm-oocyte interaction in human in-vitro fertilization improves the implantation rate. Hum. Reprod. 1996, 11, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Quinn, P.; Lydic, M.L.; Ho, M.; Bastuba, M.; Hendee, F.; Brody, S.A. Confirmation of the Beneficial Effects of Brief Coincubation of Gametes in Human In Vitro Fertilization. Fertil. Steril. 1998, 69, 399–402. [Google Scholar] [CrossRef]
- Boone, W.R.; Johnson, J.E. Clinical Assisted Reproduction: Extending the Coincubation Time of Gametes Improves In Vitro Fertilization. J. Assist. Reprod. Genet. 2001, 18, 18–20. [Google Scholar] [CrossRef]
- Fujii, J.; Iuchi, Y.; Okada, F. Fundamental roles of reactive oxygen species and protective mechanisms in the female reproductive system. Reprod. Biol. Endocrinol. 2005, 3, 43. [Google Scholar] [CrossRef]
- Lee, T.-H.; Lee, M.-S.; Liu, C.-H.; Tsao, H.-M.; Huang, C.-C.; Yang, Y.-S. The Association Between Microenvironmental Reactive Oxygen Species and Embryo Development in Assisted Reproduction Technology Cycles. Reprod. Sci. 2012, 19, 725–732. [Google Scholar] [CrossRef]
- Yang, H.W.; Hwang, K.J.; Kwon, H.C.; Kim, H.S.; Choi, K.W.; Oh, K.S. Detection of reactive oxygen species (ROS) and apoptosis in human fragmented embryos. Hum. Reprod. 1998, 13, 998–1002. [Google Scholar] [CrossRef]
- Várnagy, A.; Kőszegi, T.; Györgyi, E.; Szegedi, S.; Sulyok, E.; Prémusz, V.; Bódis, J. Levels of total antioxidant capacity and 8-hydroxy-2′-deoxyguanosine of serum and follicular fluid in women undergoing in vitro fertilization: Focusing on endometriosis. Hum. Fertil. 2018, 23, 200–208. [Google Scholar] [CrossRef]
- Du Plessis, S.S.; Makker, K.; Desai, N.R.; Agarwal, A. Impact of oxidative stress on IVF. Expert Rev. Obstet. Gynecol. 2008, 3, 539–554. [Google Scholar] [CrossRef]
- Zhang, X.D.; Liu, J.X.; Liu, W.W.; Gao, Y.; Han, W.; Xiong, S.; Wu, L.H.; Huang, G.N. Time of insemination culture and outcomes of in vitro fertilization: A systematic review and meta-analysis. Hum. Reprod. Updat. 2013, 19, 685–695. [Google Scholar] [CrossRef]
- Martin, K.L. Nutritional and metabolic requirements of early cleavage stage embryos and blastocysts. Hum. Fertil. 2000, 3, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.G.; Sherman, A.N.M.; Allen, N.W.; McGowan, L.T.; Tervit, H.R. Total protein content and protein synthesis within pre-elongation stage bovine embryos. Mol. Reprod. Dev. 1998, 50, 139–145. [Google Scholar] [CrossRef]
- Sunderam, S.; Kissin, D.M.; Crawford, S.B.; Folger, S.G.; Jamieson, D.J.; Barfield, W.D. Centers for Disease C and Prevention (2014). Assisted reproductive technology surveillance–United States. MMWR Surveill Summ. 2011, 63, 1–28. [Google Scholar]
- Pomeroy, K.O.; Reed, M. The effect of light on embryos and embryo culture. J. Reprod. Biotechnol. Fertil. 2013, 5, 104–116. [Google Scholar] [CrossRef]
- Ottosen, L.D.; Hindkjaer, J.; Ingerslev, J. Light exposure of the ovum and preimplantation embryo during ART procedures. J. Assist. Reprod. Genet. 2007, 24, 99–103. [Google Scholar] [CrossRef]
- Bódis, J.; Gödöny, K.; Várnagy, Á.; Kovács, K.; Koppán, M.; Nagy, B.; Erostyák, J.; Herczeg, R.; Szekeres-Barthó, J.; Gyenesei, A.; et al. How to Reduce the Potential Harmful Effects of Light on Blastocyst Development during IVF. Med. Princ. Pract. 2020, 29, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Hockberger, P.E.; Skimina, T.A.; Centonze, V.E.; Lavin, C.; Chu, S.; Dadras, S.; Reddy, J.K.; White, J.G. Activation of flavin-containing oxidases underlies light-induced production of H2O2 in mammalian cells. Proc. Natl. Acad. Sci. USA 1999, 96, 6255–6260. [Google Scholar] [CrossRef]
- Hirao, Y.; Yanagimachi, R. Detrimental effect of visible light on meiosis of mammalian eggs in vitro. J. Exp. Zool. 1978, 206, 365–369. [Google Scholar] [CrossRef]
- Bognar, Z.; Csabai, T.J.; Pallinger, E.; Balassa, T.; Farkas, N.; Schmidt, J.; Görgey, E.; Berta, G.; Szekeres-Bartho, J.; Bodis, J. The effect of light exposure on the cleavage rate and implantation capacity of preimplantation murine embryos. J. Reprod. Immunol. 2019, 132, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Lavi, R.; Ankri, R.; Sinyakov, M.; Eichler, M.; Friedmann, H.; Shainberg, A.; Breitbart, H.; Lubart, R. The plasma membrane is involved in the visible light-tissue interaction. Photomed. Laser Surg. 2012, 30, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Abdelrazik, H.; Sharma, R.; Mahfouz, R.; Agarwal, A. l-Carnitine decreases DNA damage and improves the in vitro blastocyst development rate in mouse embryos. Fertil. Steril. 2009, 91, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Martin-Romero, F.J.; Miguel-Lasobras, E.M.; Dominguez-Arroyo, J.A.; Gonzalez-Carrera, E.; Alvarez, I.S. Contribution of culture media to oxidative stress and its effect on human oocytes. Reprod. Biomed. Online 2008, 17, 652–661. [Google Scholar] [CrossRef]
- Menezo, Y.; Dale, B.; Cohen, M. DNA damage and repair in human oocytes and embryos: A review. Zygote 2010, 18, 357–365. [Google Scholar] [CrossRef]
- Alvarez, J.G.; Storey, B.T. Taurine, hypotaurine, epinephrine and albumin inhibit lipid peroxidation in rabbit spermatozoa and protect against loss of motility. Biol. Reprod. 1983, 29, 548–555. [Google Scholar] [CrossRef]
- Shannon, P. Factors affecting semen preservation and conception rates in cattle. J. Reprod. Fertil. 1978, 54, 519–527. [Google Scholar] [CrossRef]
- Labied, S.; Jouan, C.; Wenders, F.; Ravet, S.; Gaspard, O.; Thonon, F.; Gridelet, V.; Henry, L.; Perrier d’Hauterive, S.; Nisolle, M. Comparison between paraffin and mineral oil covering on early human embryo culture: A prospective randomized study. Syst. Biol. Reprod. Med. 2019, 65, 81–86. [Google Scholar] [CrossRef]
- Lan, K.C.; Lin, Y.C.; Chang, Y.C.; Lin, H.J.; Tsai, Y.R.; Kang, H.Y. Limited relationships between reactive oxygen species levels in culture media and zygote and embryo development. J. Assist. Reprod. Genet. 2019, 36, 325–334. [Google Scholar] [CrossRef]
- Pool, T.B.; Martin, J.E. High continuing pregnancy rates after in vitro fertilization-embryo transfer using medium supplemented with a plasma protein fraction containing alpha- and beta-globulins. Fertil. Steril. 1994, 61, 714–719. [Google Scholar] [CrossRef]
- Aitken, R.J. Impact of oxidative stress on male and female germ cells: Implications for fertility. Reproduction 2020, 159, R189–R201. [Google Scholar] [CrossRef] [PubMed]
- Shih, Y.F.; Lee, T.H.; Liu, C.H.; Tsao, H.M.; Huang, C.C.; Lee, M.S. Effects of reactive oxygen species levels in prepared culture media on embryo development: A comparison of two media. Taiwan J. Obstet. Gynecol. 2014, 53, 504–508. [Google Scholar] [CrossRef]
- Sills, E.S.; Palermo, G.D. Human blastocyst culture in IVF: Current laboratory applications in reproductive medicine practice. Rom. J. Morphol. Embryol. 2010, 51, 441–445. [Google Scholar]
- Yamanaka, K.I.; Khatun, H.; Egashira, J.; Balboula, A.Z.; Tatemoto, H.; Sakatani, M.; Takenouchi, N.; Wada, Y.; Takahashi, M. Heat-shock-induced cathepsin B activity during IVF and culture compromises the developmental competence of bovine embryos. Theriogenology 2018, 114, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Hyun, C.S.; Cha, J.H.; Son, W.Y.; Yoon, S.H.; Kim, K.A.; Lim, J.H. Optimal ICSI timing after the first polar body extrusion in in vitro matured human oocytes. Hum. Reprod. 2007, 22, 1991–1995. [Google Scholar] [CrossRef] [PubMed]
- Kharche, S.D.; Pathak, J.; Agarwal, S.; Kushwah, B.; Sikarwar, A. Effect of Ca Ionophore on Blastocyst Production Following Intracytoplasmic Sperm Injection in Caprine Oocytes. Reprod. Domest. Anim. 2016, 51, 611–617. [Google Scholar] [CrossRef]
- Samiec, M.; Skrzyszowska, M. Molecular conditions of the cell nucleus remodelling/reprogramming process and nuclear transferred embryo development in the intraooplasmic karyoplast injection technique: A review. Czech J. Anim. Sci. 2005, 50, 185–195. [Google Scholar] [CrossRef]
- Qu, P.; Shen, C.; Du, Y.; Qin, H.; Luo, S.; Fu, S.; Dong, Y.; Guo, S.; Hu, F.; Xue, Y.; et al. Melatonin Protects Rabbit Somatic Cell Nuclear Transfer (SCNT) Embryos from Electrofusion Damage. Sci. Rep. 2020, 10, 2186. [Google Scholar] [CrossRef]
- Wiater, J.; Samiec, M.; Wartalski, K.; Smorąg, Z.; Jura, J.; Słomski, R.; Skrzyszowska, M.; Romek, M. Characterization of Mono- and Bi-Transgenic Pig-Derived Epidermal Keratinocytes Expressing Human FUT2 and GLA Genes–In Vitro Studies. Int. J. Mol. Sci. 2021, 22, 9683. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mauchart, P.; Vass, R.A.; Nagy, B.; Sulyok, E.; Bódis, J.; Kovács, K. Oxidative Stress in Assisted Reproductive Techniques, with a Focus on an Underestimated Risk Factor. Curr. Issues Mol. Biol. 2023, 45, 1272-1286. https://doi.org/10.3390/cimb45020083
Mauchart P, Vass RA, Nagy B, Sulyok E, Bódis J, Kovács K. Oxidative Stress in Assisted Reproductive Techniques, with a Focus on an Underestimated Risk Factor. Current Issues in Molecular Biology. 2023; 45(2):1272-1286. https://doi.org/10.3390/cimb45020083
Chicago/Turabian StyleMauchart, Péter, Réka Anna Vass, Bernadett Nagy, Endre Sulyok, József Bódis, and Kálmán Kovács. 2023. "Oxidative Stress in Assisted Reproductive Techniques, with a Focus on an Underestimated Risk Factor" Current Issues in Molecular Biology 45, no. 2: 1272-1286. https://doi.org/10.3390/cimb45020083
APA StyleMauchart, P., Vass, R. A., Nagy, B., Sulyok, E., Bódis, J., & Kovács, K. (2023). Oxidative Stress in Assisted Reproductive Techniques, with a Focus on an Underestimated Risk Factor. Current Issues in Molecular Biology, 45(2), 1272-1286. https://doi.org/10.3390/cimb45020083