
Hydrogen Attenuates Chronic Intermittent Hypoxia-Induced Cardiac Hypertrophy by Regulating Iron Metabolism

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Echocardiography
2.3. Histopathological Examinations
2.4. Mitochondrial Membrane Potential Measurement
2.5. Immunohistochemistry
2.6. Transmission Electron Microscope
2.7. Perls’ Staining
2.8. Determination of Total Iron
2.9. Q-PCR
2.10. Western Blot
2.11. Cell Culture and Associated Assay
2.11.1. Preparation of Hydrogen-Rich Media
2.11.2. Cell Culture
2.11.3. Cell Viability Assay
2.11.4. FerroOrange Staining
2.11.5. ROS Levels
2.11.6. Immunofluorescence Double Staining
2.12. Statistical Analysis
3. Results
3.1. Results
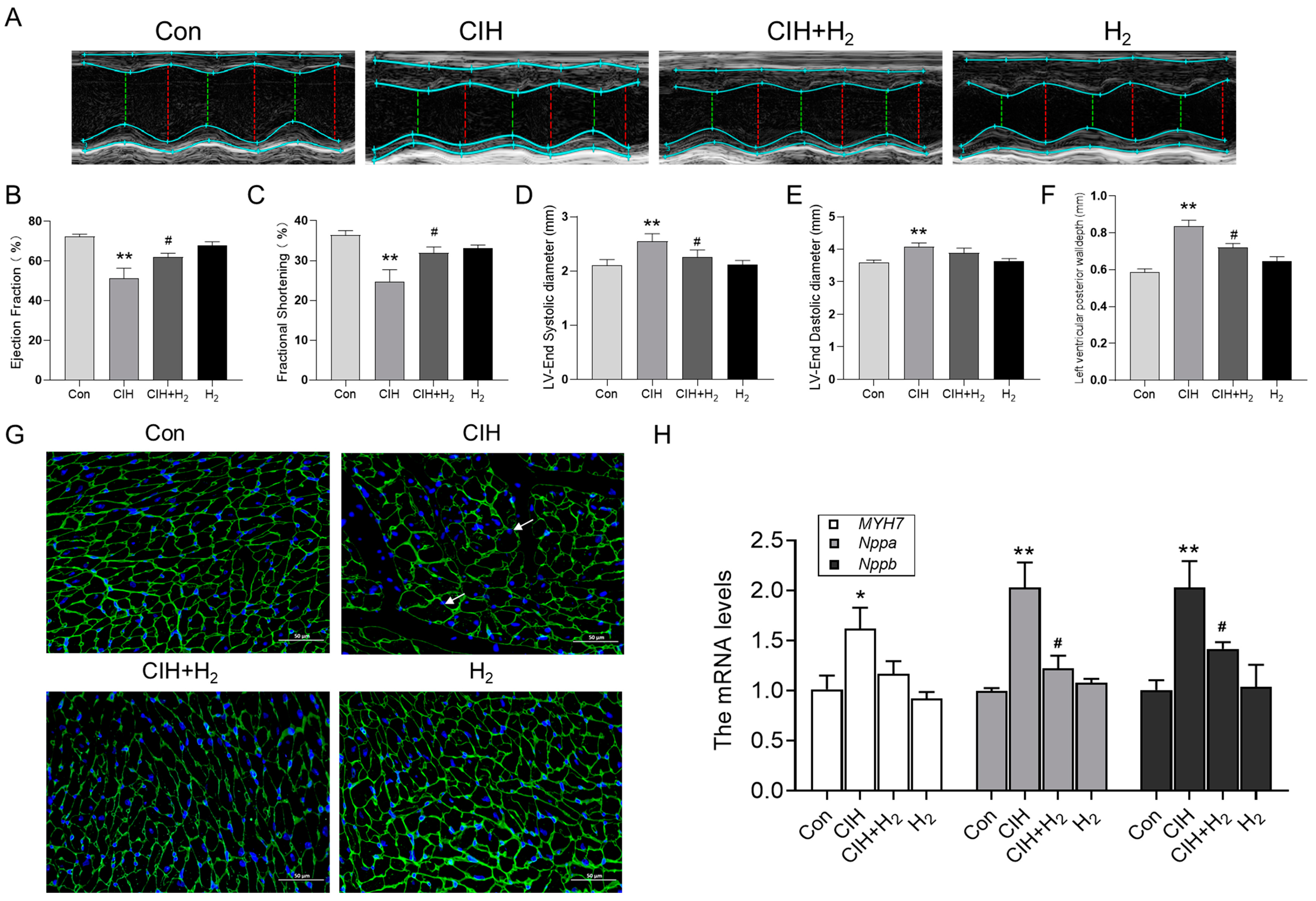
3.1.1. Hydrogen Improved Cardiac Hypertrophy and Histological Changes Induced by CIH
3.1.2. Hydrogen Mitigated the Mitochondrial Dysfunction Induced by CIH
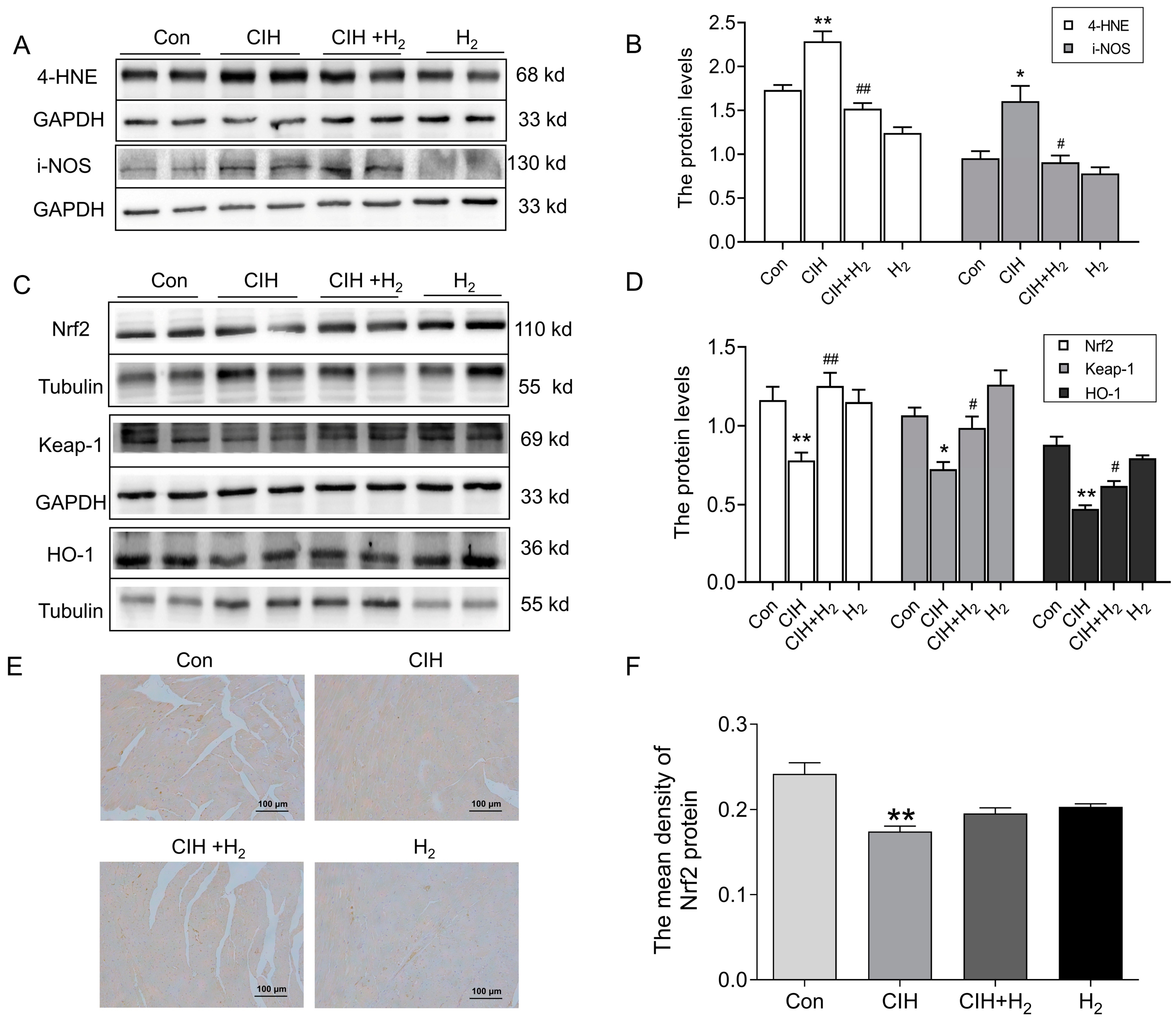
3.1.3. Hydrogen Efficiently Inhibited Oxidative Stress in Cardiac Tissue Induced by CIH
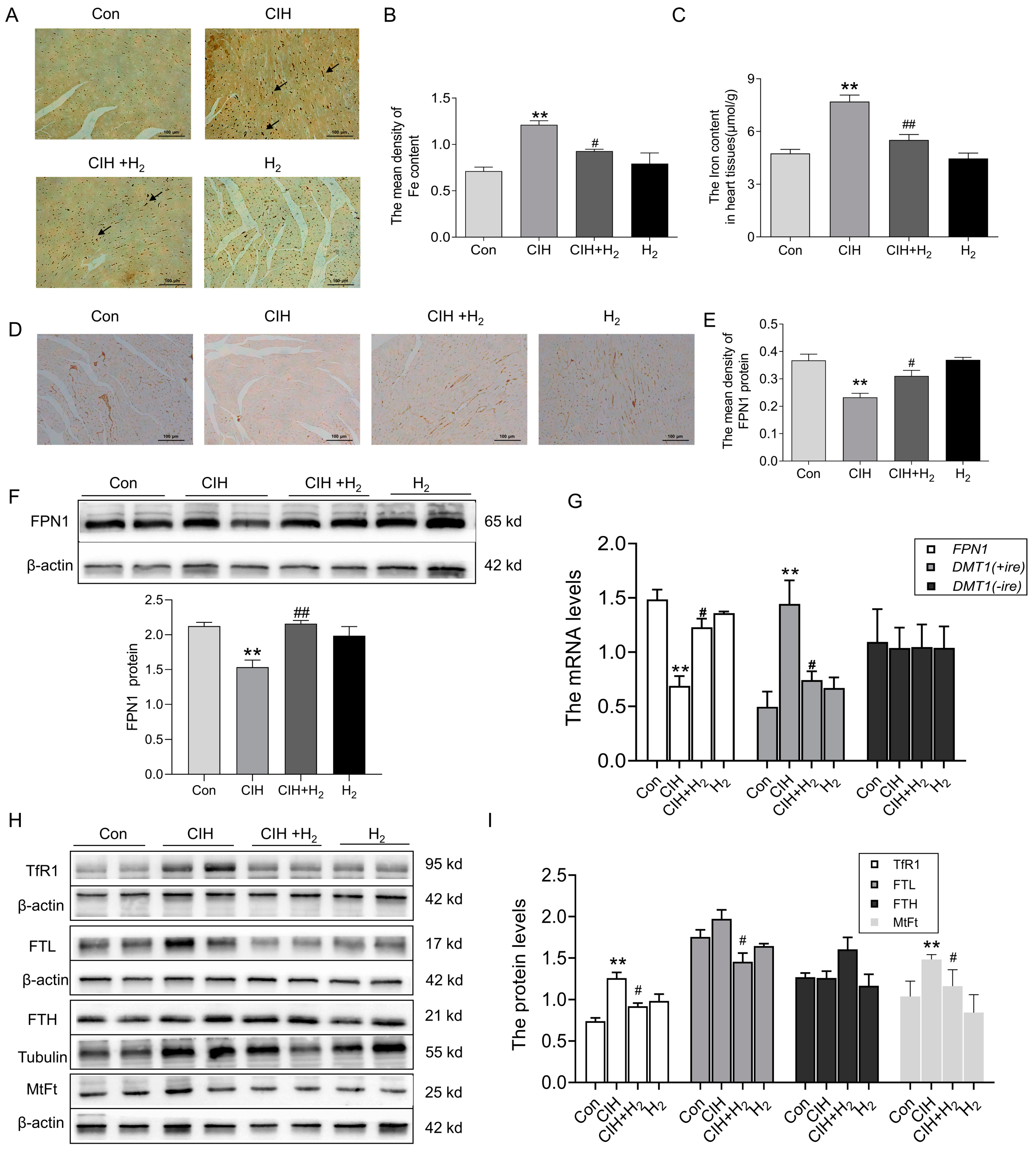
3.1.4. Hydrogen Decreased Iron Deposits in the Cardiac Tissue of CIH Mice by Upregulating FPN1
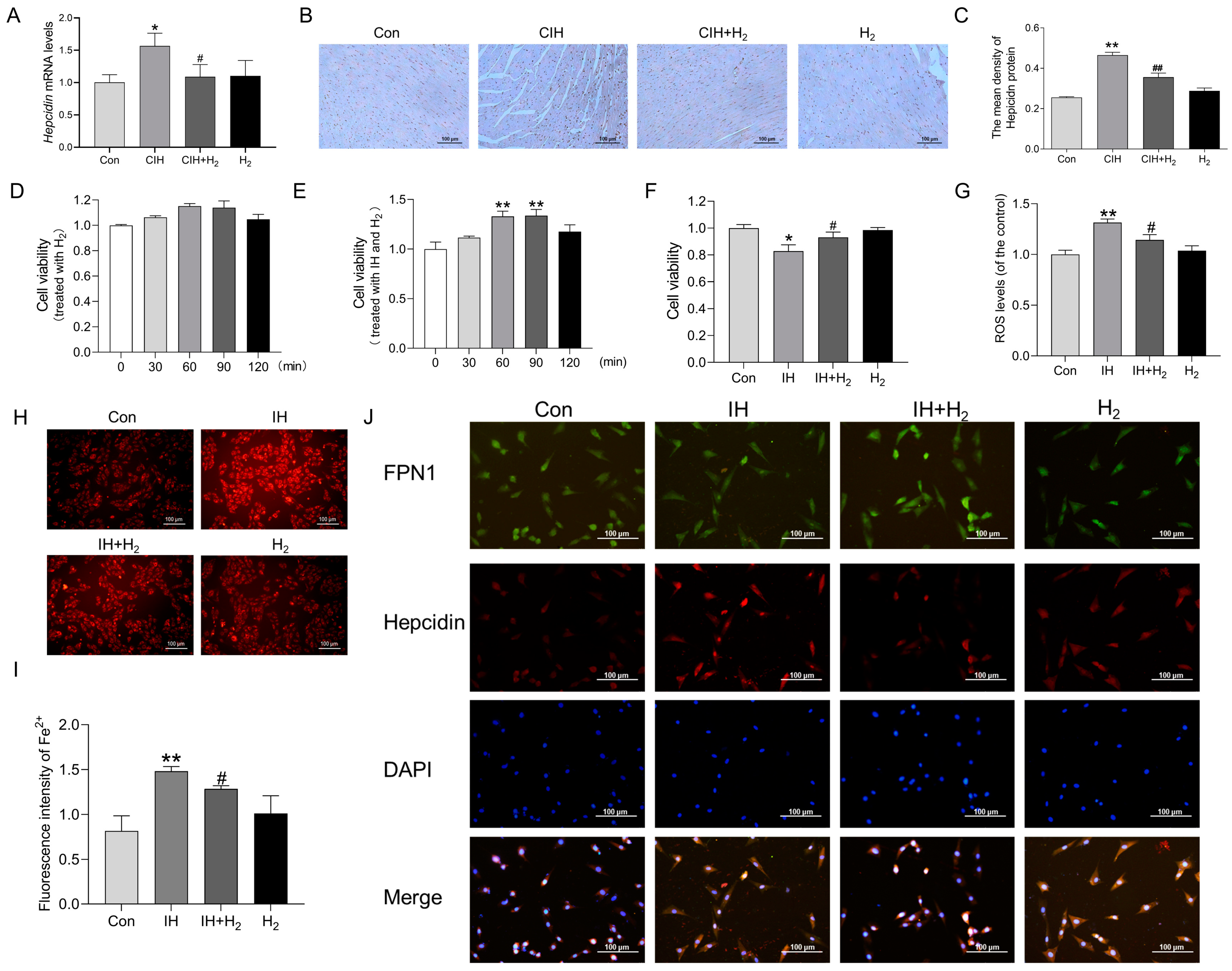
3.1.5. Hydrogen Inhibited Iron Deposition by Regulating Hepcidin
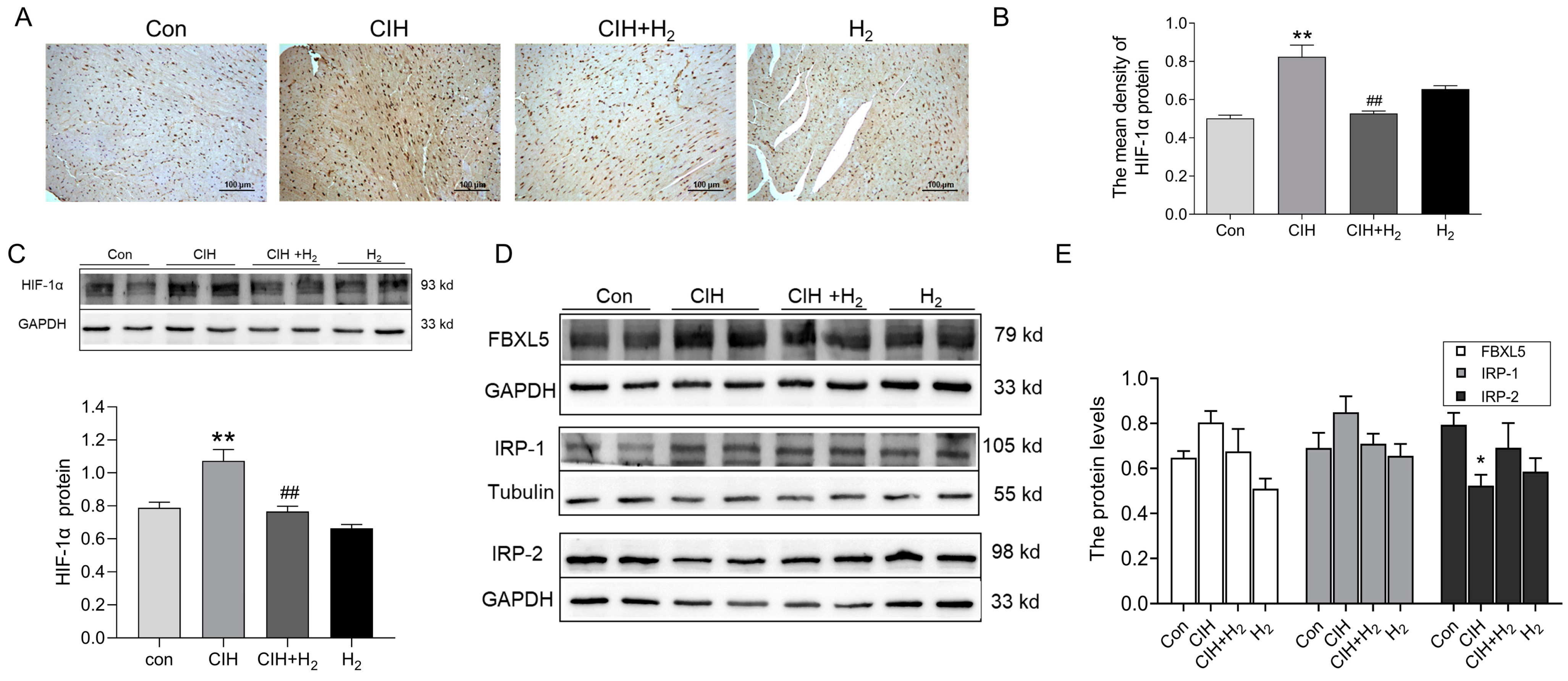
3.1.6. Hydrogen Upregulated HIF-1α Expression
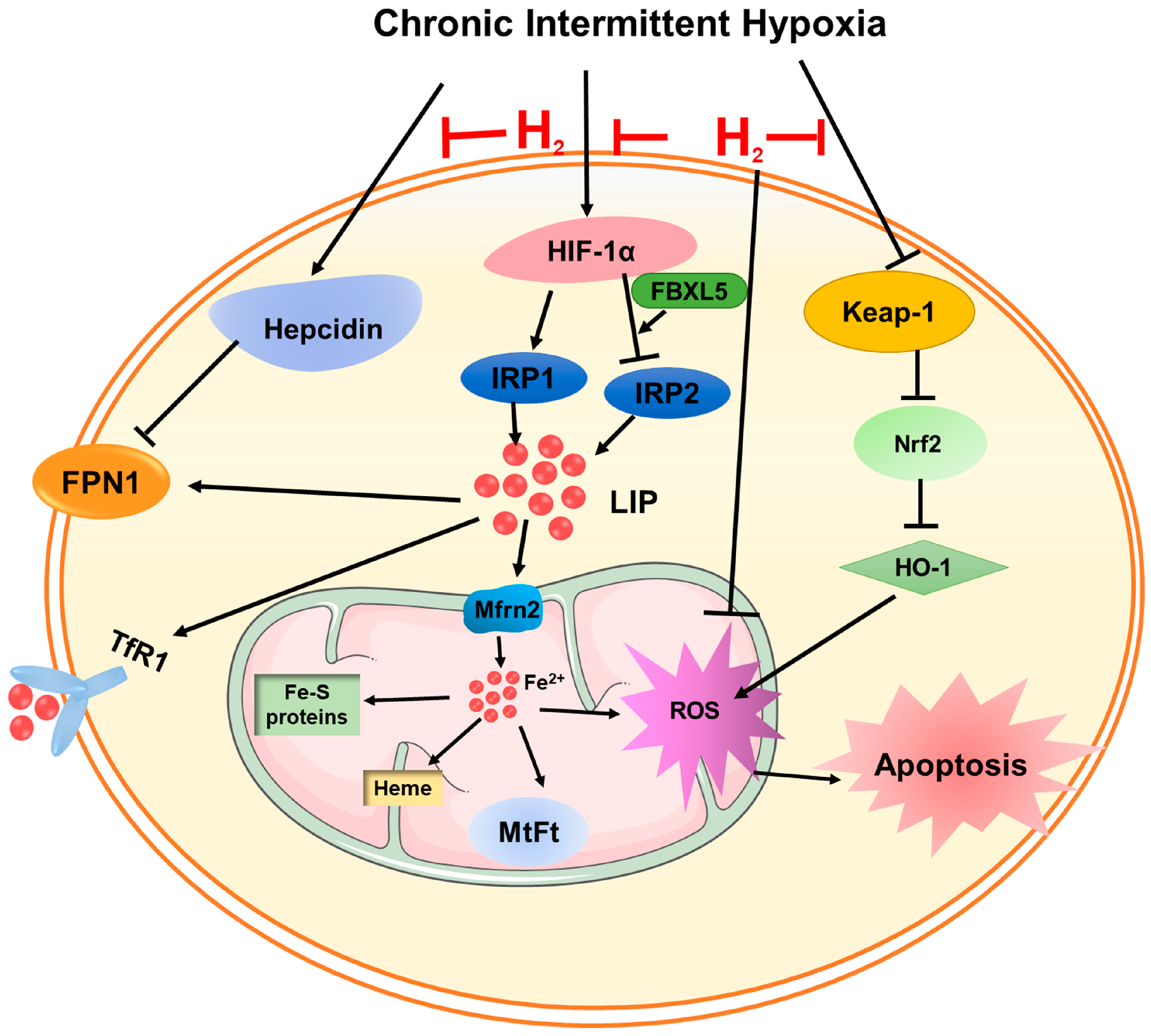
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roche, J.; Rae, D.E.; Redman, K.N.; Knutson, K.L.; von Schantz, M.; Gomez-Olive, F.X.; Scheuermaier, K. Impact of obstructive sleep apnea on cardiometabolic health in a random sample of older adults in rural South Africa: Building the case for the treatment of sleep disorders in underresourced settings. J. Clin. Sleep Med. 2021, 17, 1423–1434. [Google Scholar] [CrossRef] [PubMed]
- Song, J.X.; Zhao, Y.S.; Zhen, Y.Q.; Yang, X.Y.; Chen, Q.; An, J.R.; Ji, E.S. Banxia-Houpu decoction diminishes iron toxicity damage in heart induced by chronic intermittent hypoxia. Pharm. Biol. 2022, 60, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Kura, B.; Szeiffova Bacova, B.; Kalocayova, B.; Sykora, M.; Slezak, J. Oxidative Stress-Responsive MicroRNAs in Heart Injury. Int. J. Mol. Sci. 2020, 21, 358. [Google Scholar] [CrossRef] [PubMed]
- Imano, H.; Kato, R.; Tanikawa, S.; Yoshimura, F.; Nomura, A.; Ijiri, Y.; Yamaguchi, T.; Izumi, Y.; Yoshiyama, M.; Hayashi, T. Factor Xa inhibition by rivaroxaban attenuates cardiac remodeling due to intermittent hypoxia. J. Pharmacol. Sci. 2018, 137, 274–282. [Google Scholar] [CrossRef]
- Carpenter, C.E.; Mahoney, A.W. Contributions of heme and nonheme iron to human nutrition. Crit. Rev. Food Sci. Nutr. 1992, 31, 333–367. [Google Scholar] [CrossRef] [PubMed]
- Koppenol, W.H.; Hider, R.H. Iron and redox cycling. Do’s and don’ts. Free Radic. Biol. Med. 2019, 133, 3–10. [Google Scholar] [CrossRef]
- Hentze, M.W.; Muckenthaler, M.U.; Galy, B.; Camaschellay, C. Two to tango: Regulation of Mammalian iron metabolism. Cell 2010, 142, 24–38. [Google Scholar] [CrossRef]
- Fang, X.; Cai, Z.; Wang, H.; Han, D.; Cheng, Q.; Zhang, P.; Gao, F.; Yu, Y.; Song, Z.; Wu, Q.; et al. Loss of Cardiac Ferritin H Facilitates Cardiomyopathy via Slc7a11-Mediated Ferroptosis. Circ. Res. 2020, 127, 486–501. [Google Scholar] [CrossRef]
- Yang, T.; Brittenham, G.M.; Dong, W.Q.; Levy, M.N.; Obejero-Paz, C.A.; Kuryshev, Y.A.; Brown, A.M. Deferoxamine prevents cardiac hypertrophy and failure in the gerbil model of iron-induced cardiomyopathy. J. Lab. Clin. Med. 2003, 142, 332–340. [Google Scholar] [CrossRef]
- Gao, X.; Qian, M.; Campian, J.L.; Marshall, J.; Zhou, Z.; Roberts, A.M.; Kang, Y.J.; Prabhu, S.D.; Sun, X.F.; Eaton, J.W. Mitochondrial dysfunction may explain the cardiomyopathy of chronic iron overload. Free Radic. Biol. Med. 2010, 49, 401–407. [Google Scholar] [CrossRef]
- Walker, E.M., Jr.; Epling, C.P.; Parris, C.; Cansino, S.; Ghosh, P.; Desai, D.H.; Morrison, R.G.; Wright, G.L.; Wehner, P.; Mangiarua, E.I.; et al. Acetaminophen protects against iron-induced cardiac damage in gerbils. Ann. Clin. Lab. Sci. 2007, 37, 22–33. [Google Scholar] [PubMed]
- Guan, P.; Sun, Z.M.; Luo, L.F.; Zhao, Y.S.; Yang, S.C.; Yu, F.Y.; Wang, N.; Ji, E.S. Hydrogen Gas Alleviates Chronic Intermittent Hypoxia-Induced Renal Injury through Reducing Iron Overload. Molecules 2019, 24, 1184. [Google Scholar] [CrossRef] [PubMed]
- Dorofeyeva, N.; Drachuk, K.; Rajkumar, R.; Sabnis, O.; Sagach, V. H2S donor improves heart function and vascular relaxation in diabetes. Eur. J. Clin. Investig. 2021, 51, e13354. [Google Scholar] [CrossRef] [PubMed]
- Ohta, S. Recent progress toward hydrogen medicine: Potential of molecular hydrogen for preventive and therapeutic applications. Curr. Pharm. Des. 2011, 17, 2241–2252. [Google Scholar] [CrossRef] [PubMed]
- Cole, A.R.; Sperotto, F.; DiNardo, J.A.; Carlisle, S.; Rivkin, M.J.; Sleeper, L.A.; Kheir, J.N. Safety of Prolonged Inhalation of Hydrogen Gas in Air in Healthy Adults. Crit. Care Explor. 2021, 3, e543. [Google Scholar] [CrossRef] [PubMed]
- Nakao, A.; Kaczorowski, D.J.; Wang, Y.; Cardinal, J.S.; Buchholz, B.M.; Sugimoto, R.; Tobita, K.; Lee, S.; Toyoda, Y.; Billiar, T.R.; et al. Amelioration of rat cardiac cold ischemia/reperfusion injury with inhaled hydrogen or carbon monoxide, or both. J. Heart Lung Transplant. 2010, 29, 544–553. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.S.; Zheng, H. Chronic hydrogen-rich saline treatment reduces oxidative stress and attenuates left ventricular hypertrophy in spontaneous hypertensive rats. Mol. Cell Biochem. 2012, 365, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Lin, X.-M.; Yang, S.-C.; Guo, Y.-J.; Li, W.-Y.; Zhao, Y.-S.; Yu, F.-Y.; Sun, Z.-M.; An, J.-R.; Ji, E.-S. Hydrogen gas reduces chronic intermittent hypoxia-induced hypertension by inhibiting sympathetic nerve activity and increasing vasodilator responses via the antioxidation. J. Cell. Biochem. 2019, 120, 3998–4008. [Google Scholar] [CrossRef]
- Goes, C.M.; Falcochio, P.; Drager, L.F. Strategies to manage obstructive sleep apnea to decrease the burden of atrial fibrillation. Expert Rev. Cardiovasc. Ther. 2018, 16, 707–713. [Google Scholar] [CrossRef]
- Lee, S.D.; Kuo, W.W.; Wu, C.H.; Lin, Y.M.; Lin, J.A.; Lu, M.C.; Yang, A.L.; Liu, J.Y.; Wang, S.G.; Liu, C.J.; et al. Effects of short- and long-term hypobaric hypoxia on Bcl2 family in rat heart. Int. J. Cardiol. 2006, 108, 376–384. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Nishikimi, T.; Kuwahara, K. Atrial and brain natriuretic peptides: Hormones secreted from the heart. Peptides 2019, 111, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Rose, J.; Kraft, T.; Brenner, B.; Montag, J. Hypertrophic cardiomyopathy MYH7 mutation R723G alters mRNA secondary structure. Physiol. Genom. 2020, 52, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Jia, L.; Chen, B.; Zhang, L.; Liu, J.; Long, J.; Li, Y. Hydrogen Inhalation is Superior to Mild Hypothermia in Improving Cardiac Function and Neurological Outcome in an Asphyxial Cardiac Arrest Model of Rats. Shock 2016, 46, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.B.; Tang, Y.C.; Xu, X.J.; Guo, S.X.; Wang, H.Z. Hydrogen gas inhalation protects against liver ischemia/reperfusion injury by activating the NF-kappaB signaling pathway. Exp. Ther. Med. 2015, 9, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, T.; Huang, C.S.; Tochigi, N.; Lee, S.; Shigemura, N.; Billiar, T.R.; Okumura, M.; Nakao, A.; Toyoda, Y. Inhaled hydrogen gas therapy for prevention of lung transplant-induced ischemia/reperfusion injury in rats. Transplantation 2010, 90, 1344–1351. [Google Scholar] [CrossRef] [PubMed]
- Kato, R.; Nomura, A.; Sakamoto, A.; Yasuda, Y.; Amatani, K.; Nagai, S.; Sen, Y.; Ijiri, Y.; Okada, Y.; Yamaguchi, T.; et al. Hydrogen gas attenuates embryonic gene expression and prevents left ventricular remodeling induced by intermittent hypoxia in cardiomyopathic hamsters. Am. J. Physiol. Heart Circ. Physiol. 2014, 307, H1626–H1633. [Google Scholar] [CrossRef] [PubMed]
- Ohsawa, I.; Nishimaki, K.; Yamagata, K.; Ishikawa, M.; Ohta, S. Consumption of hydrogen water prevents atherosclerosis in apolipoprotein E knockout mice. Biochem. Biophys. Res. Commun. 2008, 377, 1195–1198. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yang, H.; Fan, Y.; Li, L.; Fang, J.; Yang, W. Hydrogen-Rich Saline Attenuates Cardiac and Hepatic Injury in Doxorubicin Rat Model by Inhibiting Inflammation and Apoptosis. Mediat. Inflamm. 2016, 2016, 1320365. [Google Scholar] [CrossRef]
- You, J.; Yue, Z.; Chen, S.; Chen, Y.; Lu, X.; Zhang, X.; Shen, P.; Li, J.; Han, Q.; Li, Z.; et al. Receptor-interacting Protein 140 represses Sirtuin 3 to facilitate hypertrophy, mitochondrial dysfunction and energy metabolic dysfunction in cardiomyocytes. Acta Physiol. 2017, 220, 58–71. [Google Scholar] [CrossRef]
- Rababa’h, A.M.; Guillory, A.N.; Mustafa, R.; Hijjawi, T. Oxidative Stress and Cardiac Remodeling: An Updated Edge. Curr. Cardiol. Rev. 2018, 14, 53–59. [Google Scholar] [CrossRef]
- Hasan, P.; Saotome, M.; Ikoma, T.; Iguchi, K.; Kawasaki, H.; Iwashita, T.; Hayashi, H.; Maekawa, Y. Mitochondrial fission protein, dynamin-related protein 1, contributes to the promotion of hypertensive cardiac hypertrophy and fibrosis in Dahl-salt sensitive rats. J. Mol. Cell Cardiol. 2018, 121, 103–106. [Google Scholar] [CrossRef]
- Liu, F.; Su, H.; Liu, B.; Mei, Y.; Ke, Q.; Sun, X.; Tan, W. STVNa Attenuates Isoproterenol-Induced Cardiac Hypertrophy Response through the HDAC4 and Prdx2/ROS/Trx1 Pathways. Int. J. Mol. Sci. 2020, 21, 682. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J. Redox signaling: An evolution from free radicals to aging. Free Radic. Biol. Med. 2016, 97, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Ying, W. NAD+/NADH and NADP+/NADPH in cellular functions and cell death: Regulation and biological consequences. Antioxid. Redox Signal. 2008, 10, 179–206. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Zhong, H.; Xia, L.; Tao, Y.; Yin, H. Pathophysiology of mitochondrial lipid oxidation: Role of 4-hydroxynonenal (4-HNE) and other bioactive lipids in mitochondria. Free Radic. Biol. Med. 2017, 111, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, H.; Kinugawa, S.; Matsushima, S. Oxidative stress and heart failure. Am. J. Physiol. Heart Circ. Physiol. 2011, 301, H2181–H2190. [Google Scholar] [CrossRef] [PubMed]
- Rajapakse, N.W.; Giam, B.; Kuruppu, S.; Head, G.A.; Kaye, D.M. Impaired l-arginine-nitric oxide pathway contributes to the pathogenesis of resistant hypertension. Clin. Sci. 2019, 133, 2061–2067. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Hu, L. Nrf2 activation through the inhibition of Keap1-Nrf2 protein-protein interaction. Med. Chem. Res. 2020, 29, 846–867. [Google Scholar] [CrossRef]
- Lee, J.M.; Calkins, M.J.; Chan, K.; Kan, Y.W.; Johnson, J.A. Identification of the NF-E2-related factor-2-dependent genes conferring protection against oxidative stress in primary cortical astrocytes using oligonucleotide microarray analysis. J. Biol. Chem. 2003, 278, 12029–12038. [Google Scholar] [CrossRef]
- Lakhal-Littleton, S. Mechanisms of cardiac iron homeostasis and their importance to heart function. Free Radic. Biol. Med. 2019, 133, 234–237. [Google Scholar] [CrossRef]
- Kell, D.B. Iron behaving badly: Inappropriate iron chelation as a major contributor to the aetiology of vascular and other progressive inflammatory and degenerative diseases. BMC Med. Genom. 2009, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Oudit, G.Y.; Trivieri, M.G.; Khaper, N.; Liu, P.P.; Backx, P.H. Role of L-type Ca2+ channels in iron transport and iron-overload cardiomyopathy. J. Mol. Med. 2006, 84, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Paul, B.T.; Manz, D.H.; Torti, F.M.; Torti, S.V. Mitochondria and Iron: Current questions. Expert Rev. Hematol. 2017, 10, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Xu, A.; Li, Y.; Zhao, S.; Zhou, D.; Wu, L.; Zhang, B.; Zhao, X.; Wang, Y.; Wang, X.; et al. A novel SLC40A1 p.Y333H mutation with gain of function of ferroportin: A recurrent cause of haemochromatosis in China. Liver Int. 2019, 39, 1120–1127. [Google Scholar] [CrossRef] [PubMed]
- Okonko, D.O.; Mandal, A.K.; Missouris, C.G.; Poole-Wilson, P.A. Disordered iron homeostasis in chronic heart failure: Prevalence, predictors, and relation to anemia, exercise capacity, and survival. J. Am. Coll. Cardiol. 2011, 58, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Ganz, T. Hepcidin-Ferroportin Interaction Controls Systemic Iron Homeostasis. Int. J. Mol. Sci. 2021, 22, 6493. [Google Scholar] [CrossRef]
- Zhao, Y.S.; Tan, M.; Song, J.X.; An, J.R.; Yang, X.Y.; Li, W.Y.; Guo, Y.J.; Ji, E.S. Involvement of Hepcidin in Cognitive Damage Induced by Chronic Intermittent Hypoxia in Mice. Oxid. Med. Cell. Longev. 2021, 2021, 8520967. [Google Scholar] [CrossRef]
- Lakhal-Littleton, S.; Wolna, M.; Carr, C.A.; Miller, J.J.; Christian, H.C.; Ball, V.; Santos, A.; Diaz, R.; Biggs, D.; Stillion, R.; et al. Cardiac ferroportin regulates cellular iron homeostasis and is important for cardiac function. Proc. Natl. Acad. Sci. USA 2015, 112, 3164–3169. [Google Scholar] [CrossRef]
- Charlebois, E.; Fillebeen, C.; Katsarou, A.; Rabinovich, A.; Wisniewski, K.; Venkataramani, V.; Michalke, B.; Velentza, A.; Pantopoulos, K. A crosstalk between hepcidin and IRE/IRP pathways controls ferroportin expression and determines serum iron levels in mice. eLife 2022, 11, e81332. [Google Scholar] [CrossRef]
- Meyron-Holtz, E.G.; Ghosh, M.C.; Iwai, K.; LaVaute, T.; Brazzolotto, X.; Berger, U.V.; Land, W.; Ollivierre-Wilson, H.; Grinberg, A.; Love, P.; et al. Genetic ablations of iron regulatory proteins 1 and 2 reveal why iron regulatory protein 2 dominates iron homeostasis. EMBO J. 2004, 23, 386–395. [Google Scholar] [CrossRef]
- Jiang, H.; Luan, Z.; Wang, J.; Xie, J. Neuroprotective effects of iron chelator Desferal on dopaminergic neurons in the substantia nigra of rats with iron-overload. Neurochem. Int. 2006, 49, 605–609. [Google Scholar] [CrossRef] [PubMed]
- Yanatori, I.; Kishi, F. DMT1 and iron transport. Free Radic. Biol. Med. 2019, 133, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.A.; Horbinski, C.; Feng, L.; Dolan, K.G.; Higgins, D.; Garrick, M.D. Differential localization of divalent metal transporter 1 with and without iron response element in rat PC12 and sympathetic neuronal cells. J. Neurosci. 2000, 20, 7595–7601. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | Length |
|---|---|---|---|
| β-actin | AGGCCCAGAGCAAGAGAGGTA | TCTCCATGTCGTCCCAGTTG | 81 bp |
| Nppa | GGGTAGGATTGACAGGATTGG | CCTCCTTGGCTGTTATCTTC | 79 bp |
| Nppb | ATCCGTCAGTCGTTTGGG | CAGAGTCAGAAACTGGAGTC | 84 bp |
| MYH7 | TGTTTCCTTACTTGCTACCC | GGATTCTCAAACGTGTCTAGTG | 115 bp |
| Fis-1 | AATATGCCTGGTGCCTGGTT | GCTGTTCCTCTTTGCTCCCT | 102 bp |
| Drp-1 | AGGTTGCCCGTGACAAATGA | TCAGCAAAGTCGGGGTGTTT | 86 bp |
| Opa-1 | GTGACTATAAGTGGATTGTGCCTG | AACTGGCAAGGTCTTCTGAGC | 105 bp |
| FPN1 | TGGATGGGTCCTTACTGTCTGCTA | TGCTAATCTGCTCCTGTTTTCTCC | 139 bp |
| DMT1(+ire) | ACAGCCCAGGAGACCTTAAGAACA | ACCTTTGAACAAGCTCACCTCCGA | 97 bp |
| DMT1(−ire) | CGCCCAGATTTTACACAGTG | TTGGAGTGTCGGTGCTTAAA | 91 bp |
| hepcidin | AGACATTGCGATACCAATGCA | GCAACAGATACCACACTGGGAA | 108 bp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, J.; Chen, Q.; Xu, S.; Gou, Y.; Guo, Y.; Jia, C.; Zhao, C.; Zhang, Z.; Li, B.; Zhao, Y.; et al. Hydrogen Attenuates Chronic Intermittent Hypoxia-Induced Cardiac Hypertrophy by Regulating Iron Metabolism. Curr. Issues Mol. Biol. 2023, 45, 10193-10210. https://doi.org/10.3390/cimb45120636
Song J, Chen Q, Xu S, Gou Y, Guo Y, Jia C, Zhao C, Zhang Z, Li B, Zhao Y, et al. Hydrogen Attenuates Chronic Intermittent Hypoxia-Induced Cardiac Hypertrophy by Regulating Iron Metabolism. Current Issues in Molecular Biology. 2023; 45(12):10193-10210. https://doi.org/10.3390/cimb45120636
Chicago/Turabian StyleSong, Jixian, Qi Chen, Shan Xu, Yujing Gou, Yajing Guo, Cuiling Jia, Chenbing Zhao, Zhi Zhang, Boliang Li, Yashuo Zhao, and et al. 2023. "Hydrogen Attenuates Chronic Intermittent Hypoxia-Induced Cardiac Hypertrophy by Regulating Iron Metabolism" Current Issues in Molecular Biology 45, no. 12: 10193-10210. https://doi.org/10.3390/cimb45120636
APA StyleSong, J., Chen, Q., Xu, S., Gou, Y., Guo, Y., Jia, C., Zhao, C., Zhang, Z., Li, B., Zhao, Y., & Ji, E. (2023). Hydrogen Attenuates Chronic Intermittent Hypoxia-Induced Cardiac Hypertrophy by Regulating Iron Metabolism. Current Issues in Molecular Biology, 45(12), 10193-10210. https://doi.org/10.3390/cimb45120636