
Effects of Melatonin on Liver of D-Galactose-Induced Aged Mouse Model


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Liver Index Determination
2.3. Cell Culture and Treatment
2.4. Histological Analyses
2.5. Immunofluorescence
2.6. Western Blotting
2.7. RNA Isolation and Quantitative PCR
2.8. Detection of IL1-β, AST, and ALT Levels
2.9. Statistical Analysis
3. Results
3.1. Melatonin Protected Mice against D-Gal-Induced Liver Dysfunction
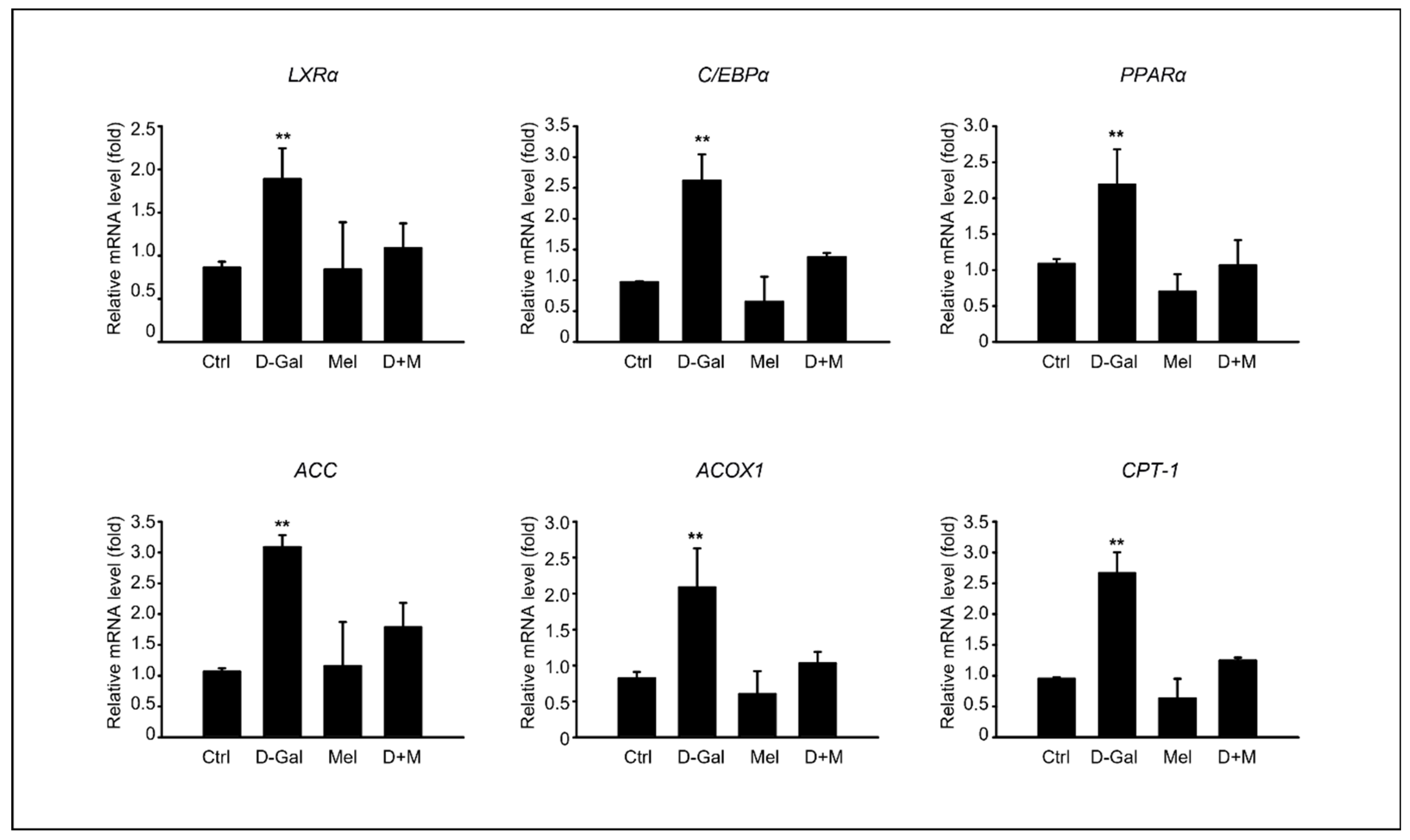
3.2. Melatonin Ameliorated the Hepatic Lipid Profiles in D-Gal-Induced Liver Disease in Aging Mice
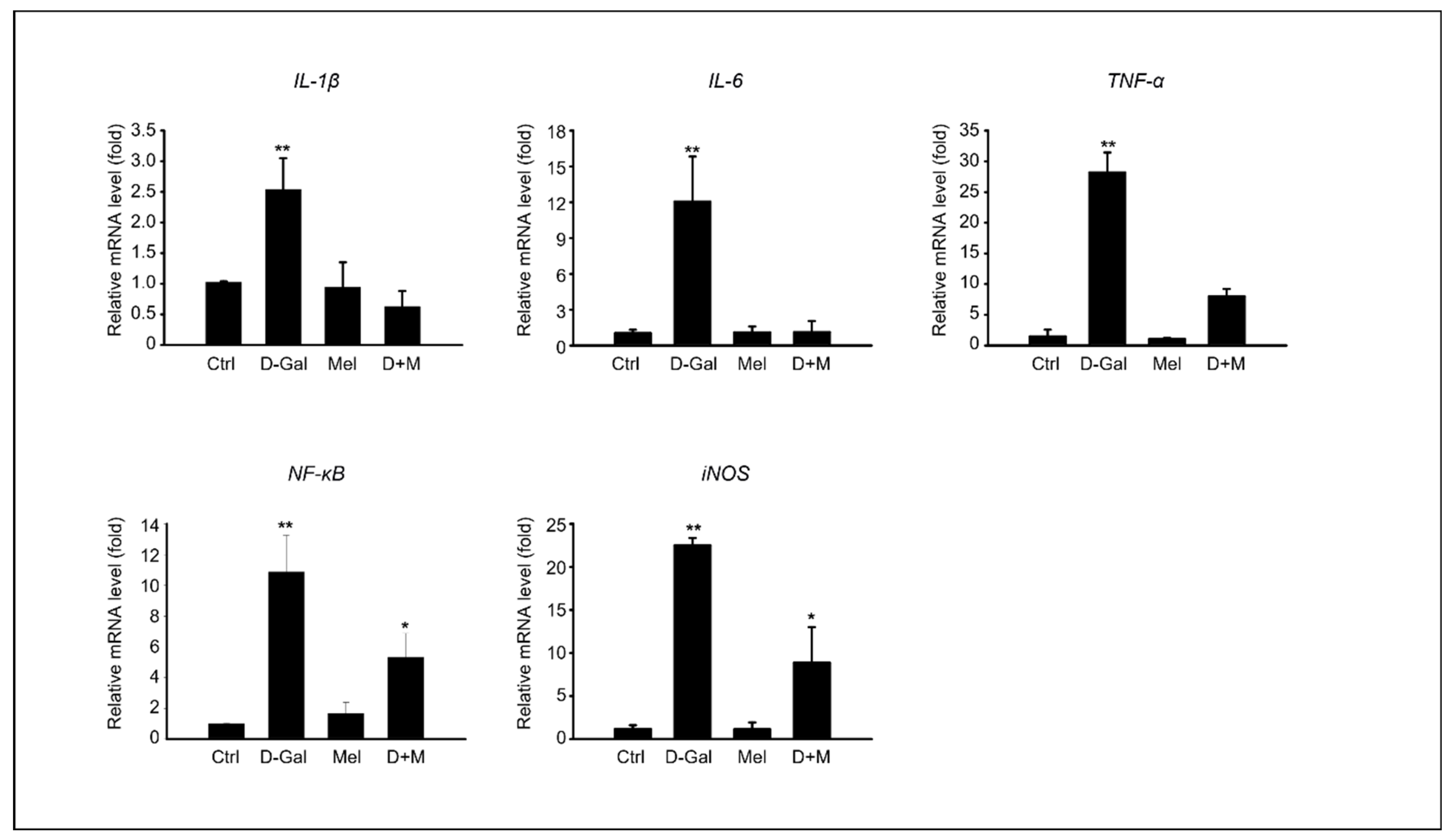
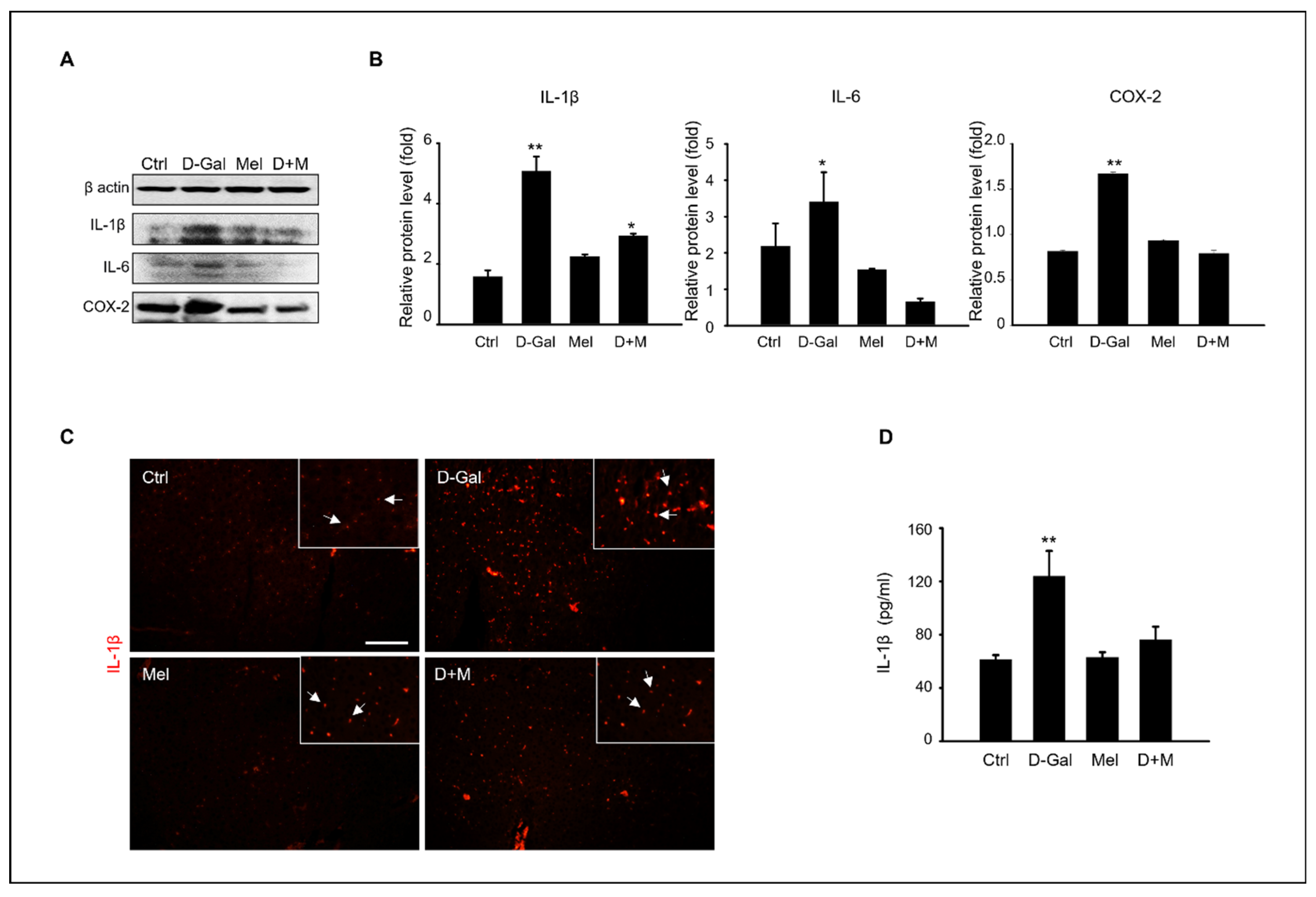
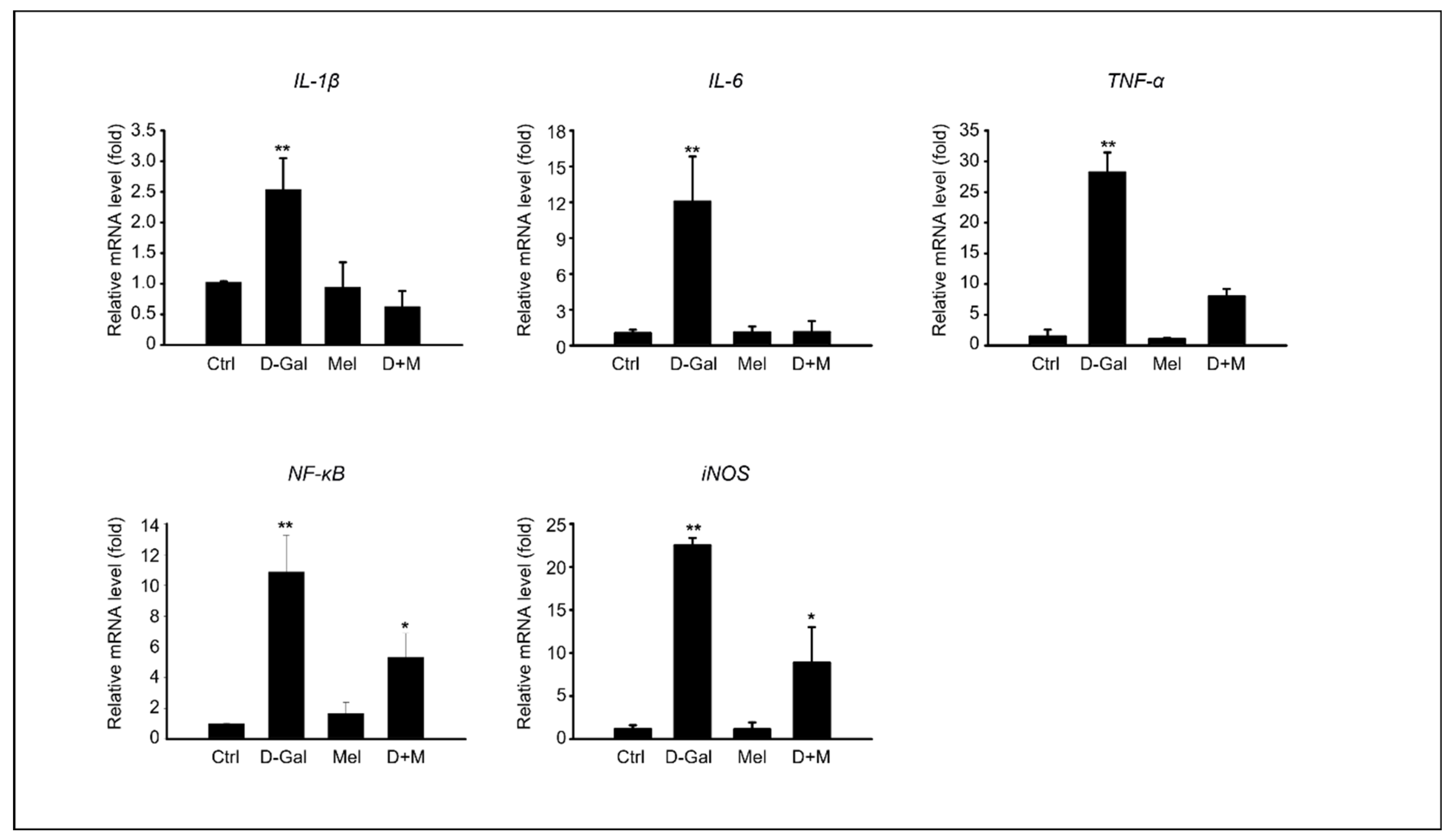
3.3. Melatonin Suppressed Pro-Inflammatory Cytokines in Induced Liver Damage in Aging Mice
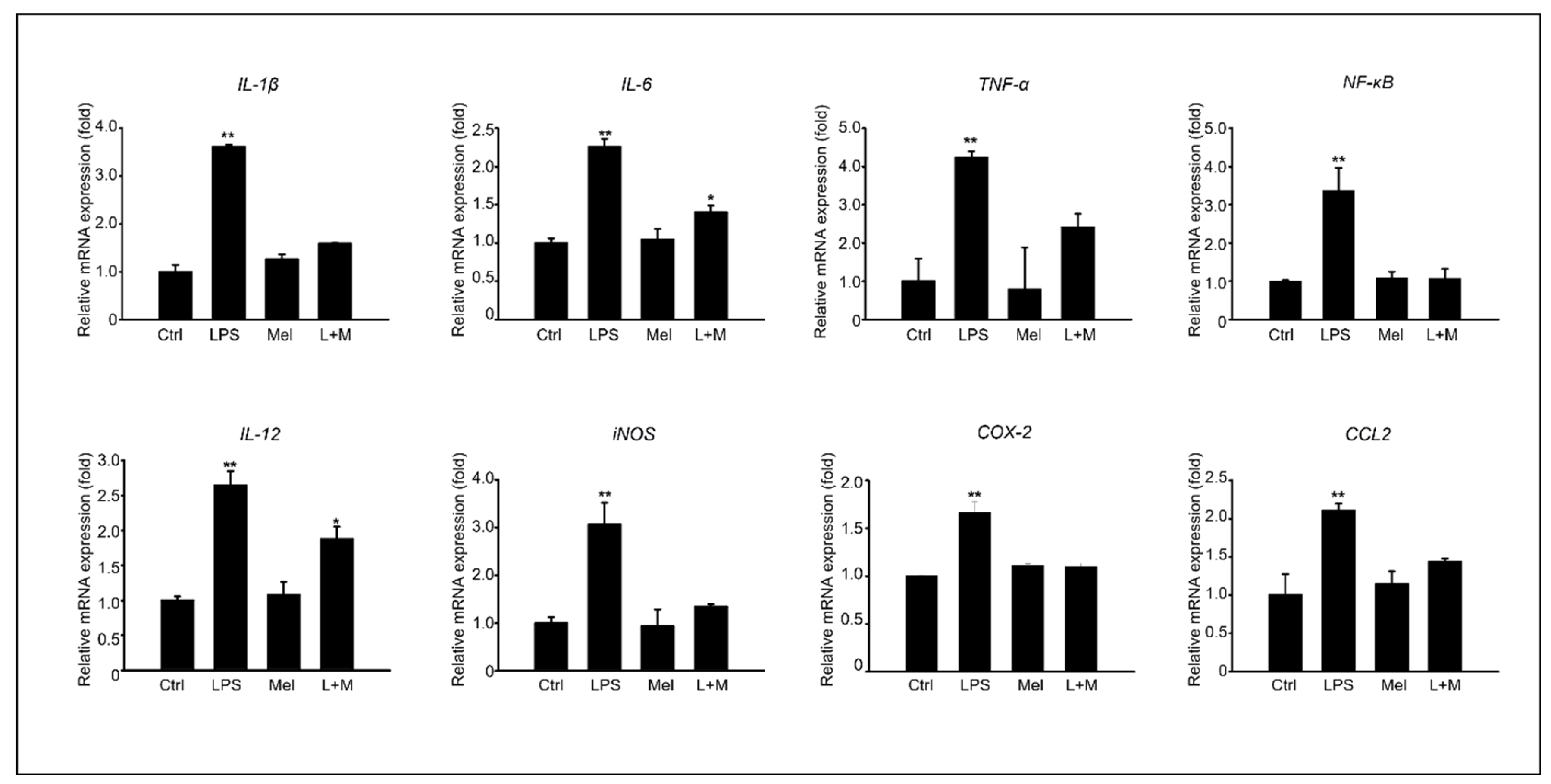
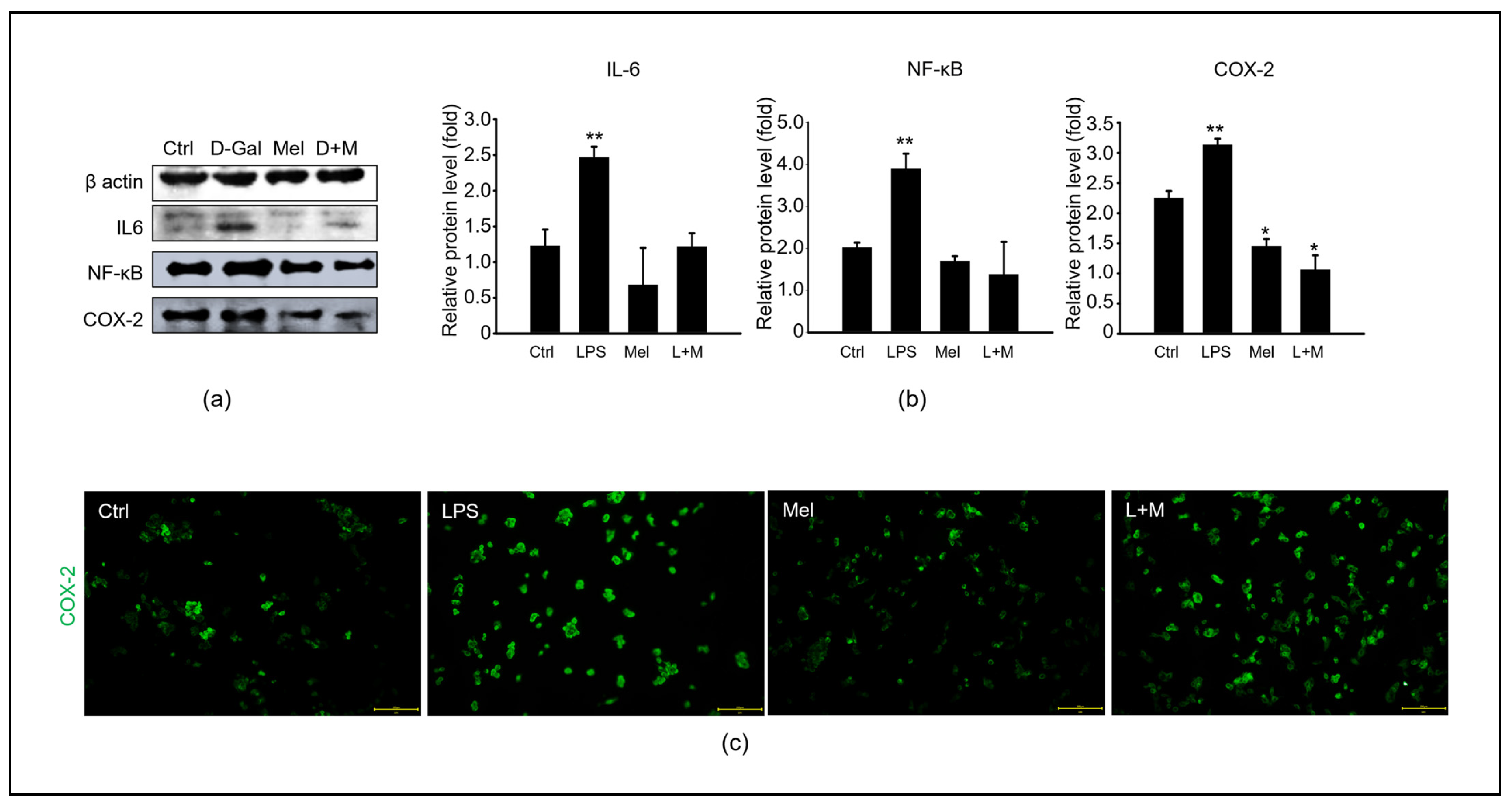
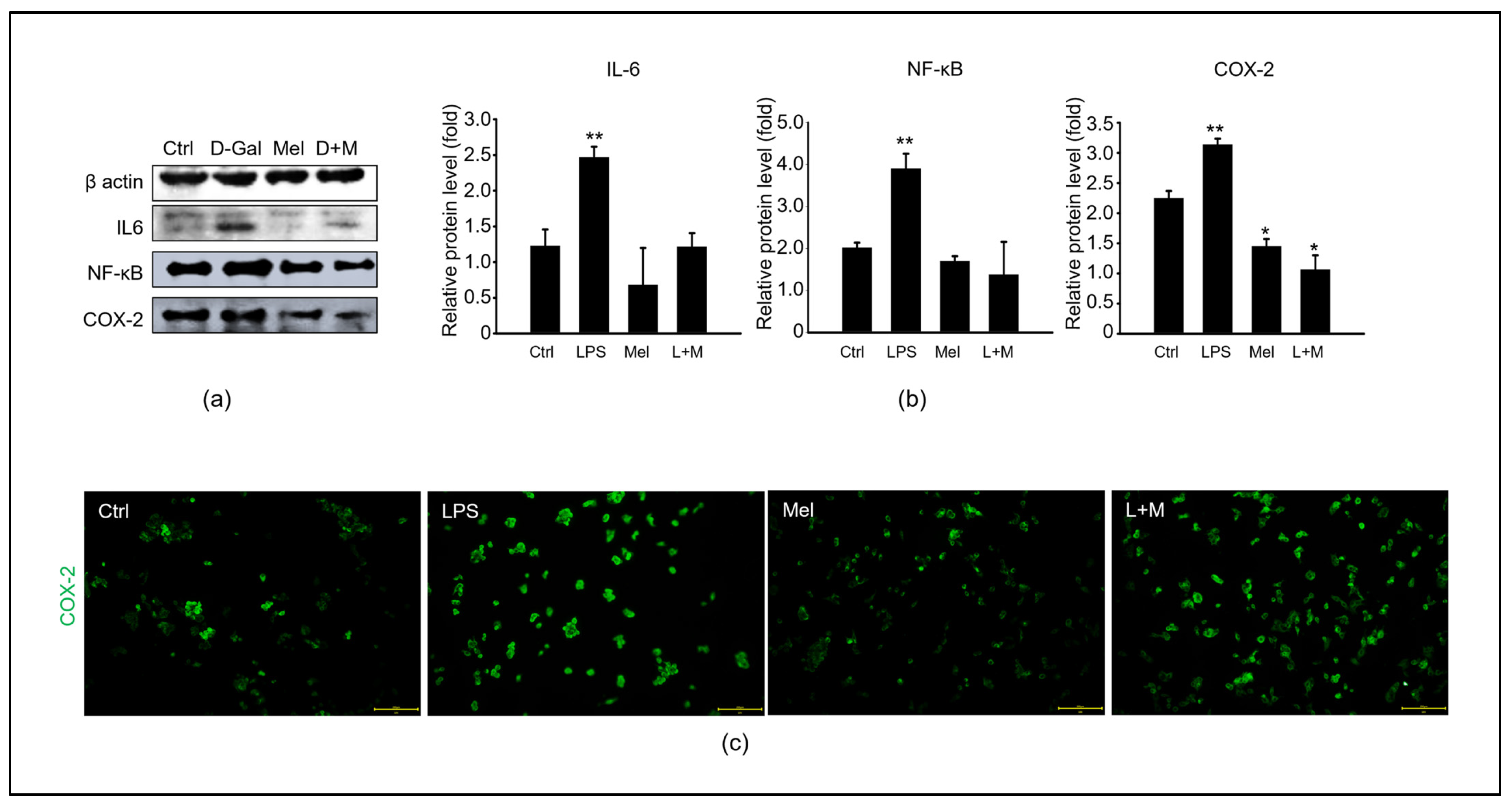
3.4. Melatonin-Enhanced Stabilization of LPS-Induced Inflammatory Mediators in HepG2 Cells
3.5. Melatonin-Regulated Stabilization of LPS-Induced Inflammatory Mediators in HepG2 Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, J.; Huang, X.; Dou, L.; Yan, M.; Shen, T.; Tang, W.; Li, J. Aging and aging-related diseases: From molecular mechanisms to interventions and treatments. Signal Transduct. Target. Ther. 2022, 7, 391. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, H.L. Problems of an Aging Population: Public Health Aspects of Diabetes. Am. J. Public Health Nations Health 1947, 37, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Cortes-Canteli, M.; Iadecola, C. Alzheimer’s Disease and Vascular Aging: JACC Focus Seminar. J. Am. Coll. Cardiol. 2020, 75, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Hoehn, M.M.; Yahr, M.D. Parkinsonism: Onset, progression, and mortality. Neurology 1967, 17, 427–442. [Google Scholar] [CrossRef]
- Kaufman, P.; Poliakoff, H. Studies on the aging heart; the pattern of rheumatic heart disease in old age (a clinical pathological study). Ann. Intern. Med. 1950, 32, 889–904. [Google Scholar]
- Cordero, A.I.H.; Yang, C.X.; Yang, J.; Horvath, S.; Shaipanich, T.; MacIsaac, J.; Lin, D.T.S.; Kobor, M.S.; Guillemi, S.; Harris, M.; et al. Airway Aging and Methylation Disruptions in HIV-associated Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2022, 206, 150–160. [Google Scholar] [CrossRef]
- Cisneros, B.; García-Aguirre, I.; Unzueta, J.; Arrieta-Cruz, I.; González-Morales, O.; Domínguez-Larrieta, J.M.; Tamez-González, A.; Leyva-Gómez, G.; Magaña, J.J. Imune system modulation in aging: Molecular mechanisms and therapeutic targets. Front. Immunol. 2022, 15, 1059173. [Google Scholar] [CrossRef]
- Fan, J.; Yang, X.; Li, J.; Shu, Z.; Dai, J.; Liu, X.; Li, B.; Jia, S.; Kou, X.; Yang, Y.; et al. Spermidine coupled with exercise rescues skeletal muscle atrophy from D-gal-induced aging rats through enhanced autophagy and reduced apoptosis via AMPK-FOXO3a signal pathway. Oncotarget 2017, 14, 17475–17490. [Google Scholar] [CrossRef]
- Tolosano, E.; Fagoonee, S.; Hirsch, E.; Berger, F.G.; Baumann, H.; Silengo, L.; Altruda, F. Enhanced splenomegaly and severe liver inflammation in haptoglobin/hemopexin double-null mice after acute hemolysis. Blood 2002, 100, 4201–4208. [Google Scholar] [CrossRef]
- Morava, E. Galactose supplementation in phosphoglucomutase-1 deficiency; review and outlook for a novel treatable CDG. Mol. Genet. Metab. 2014, 112, 275–279. [Google Scholar] [CrossRef]
- Coelho, A.I.; Berry, G.T.; Rubio-Gozalbo, M.E. Galactose metabolism and health. Curr. Opin. Clin. Nutr. Metab. Care. 2015, 18, 422–427. [Google Scholar] [CrossRef] [PubMed]
- 12 Homolak, J.; Perhoc, A.B.; Virag, D.; Knezovic, A.; Barilar, J.O.; salkovic-Petrisic, M. gactose might protect against ionizing radiation by stimulating oxidative metabolism and modulating redox homeostasis. J. Radiat. Res. 2023, 64, 743–745. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Li, L.; Song, Q.; Ai, H.; Chu, J.; Li, W. Behavioural study of the D-galactose induced aging model in C57BL/6J mice. Behav. Brain Res. 2005, 157, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Azman, K.F.; Safdar, A.; Zakaria, A. D-galactose-induced liver aging model: Its underlying mechanisms and potential therapeutic interventions. Exp. Gerontol. 2021, 150, 111372. [Google Scholar] [CrossRef]
- Azman, K.F.; Zakaria, R. D-Galactose-induced accelerated aging model: An overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef]
- Zhang, X.L.; Jiang, B.; Li, Z.B.; Hao, S.; An, L.J. Catalpol ameliorates cognition deficits and attenuates oxidative damage in the brain of senescent mice induced by D-galactose. Pharmacol. Biochem. Behav. 2007, 88, 64–72. [Google Scholar] [CrossRef]
- Theiss, A.L.; Vijay-Kumar, M.; Obertone, T.S.; Jones, D.P.; Hansen, J.M.; Gewirtz, A.T.; Merlin, D.; Sitaraman, S.V. Prohibitin is a novel regulator of antioxidant response that attenuates colonic inflammation in mice. Gastroenterology 2009, 137, 199–208. [Google Scholar] [CrossRef]
- Malhi, H.; Gores, G.J. Cellular and molecular mechanisms of liver injury. Gastroenterology 2008, 134, 1641–1654. [Google Scholar] [CrossRef]
- Jing, Z.T.; Liu, W.; Xue, C.R.; Wu, S.X.; Chen, W.N.; Lin, X.J.; Lin, X. AKT activator SC79 protects hepatocytes from TNF-α-mediated apoptosis and alleviates d-Gal/LPS-induced liver injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 316, G387–G396. [Google Scholar] [CrossRef]
- Li, C.; Lai, S.; Yi, R.; Zhou, X.; Zhao, X.; Li, Q. Blood Coral Polysaccharide Helps Prevent D-Gal/LPS-Induced Acute Liver Failure in Mice. J. Inflamm. Res. 2022, 15, 4499–4513. [Google Scholar] [CrossRef]
- Sha, J.Y.; Li, J.H.; Zhou, Y.D.; Yang, J.Y.; Liu, W.; Jiang, S.; Wang, Y.P.; Zhang, R.; Di, P.; Li, W. The p53/p21/p16 and PI3K/Akt signaling pathways are involved in the ameliorative effects of maltol on D-galactose-induced liver and kidney aging and injury. Phytother. Res. 2021, 35, 4411–4424. [Google Scholar] [CrossRef]
- Feng, Y.; Yu, Y.H.; Wang, S.T.; Ren, J.; Camer, D.; Hua, Y.Z.; Zhang, Q.; Huang, J.; Xue, D.L.; Zhang, X.F.; et al. Chlorogenic acid protects D-galactose-induced liver and kidney injury via antioxidation and anti-inflammation effects in mice. Pharm. Biol. 2016, 54, 1027–1034. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Esteban-Zubero, E.; Zhou, Z.; Reiter, R.J. Melatonin as a Potent and Inducible Endogenous Antioxidant: Synthesis and Metabolism. Molecules 2015, 20, 18886–18906. [Google Scholar] [CrossRef]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front. Endocrinol. 2019, 17, 249. [Google Scholar] [CrossRef] [PubMed]
- Kennaway, D.J. Are the proposed benefits of melatonin-rich foods too hard to swallow? Crit. Rev. Food Sci. Nutr. 2017, 57, 958–962. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Meng, X.; Li, Y.; Zhou, Y.; Xu, D.P.; Li, S.; Li, H.B. Effects of Melatonin on Liver Injuries and Diseases. Int. J. Mol. Sci. 2017, 23, 673. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, W.Y.; Zhang, M.; Hong, K.H.; Park, C.; Kim, J.H.; Song, H. Evaluation of Resmethrin Toxicity to Neonatal Testes in Organ Culture. Toxicol. Sci. 2020, 173, 53–64. [Google Scholar] [CrossRef]
- Park, H.J.; Zhang, M.; Lee, W.Y.; Hong, K.H.; Do, J.T.; Park, C.; Song, H. Toxic Effects of Nonylphenol on Neonatal Testicular Development in Mouse Organ Culture. Int. J. Mol. Sci. 2020, 15, 3491. [Google Scholar] [CrossRef]
- Lee, W.Y.; Sim, H.W.; Park, H.J. Effects of melatonin on a d-galactose-induced male reproductive aging mouse model. Theriogenology 2023, 206, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Alberts, A.; Klingberg, A.; Hoffmeister, L.; Wessig, A.K.; Brand, K.; Pich, A.; Neumann, K. Binding of Macrophage Receptor MARCO, LDL, and LDLR to Disease-Associated Crystalline Structures. Front. Immunol. 2020, 8, 596103. [Google Scholar] [CrossRef]
- Rinella, M.E. Nonalcoholic fatty liver disease: A systematic review. JAMA 2015, 9, 2263–2273. [Google Scholar] [CrossRef]
- Cariou, B.; Byrne, C.D.; Loomba, R.; Sanyal, A.J. Nonalcoholic fatty liver disease as a metabolic disease in humans: A literature review. Diabetes Obes. Metab. 2021, 23, 1069–1083. [Google Scholar] [CrossRef] [PubMed]
- Meynial-Denis, D. Glutamine metabolism in advanced age. Nutr. Rev. 2016, 74, 225–236. [Google Scholar]
- Liu, Y.X.; Liu, X.; Cen, C.; Li, X.; Liu, J.M.; Ming, Z.Y.; Yu, S.F.; Tang, X.F.; Zhou, L.; Yu, J.; et al. Comparison and development of advanced machine learning tools to predict nonalcoholic fatty liver disease: An extended study. Hepatobiliary Pancreat. Dis. Int. 2021, 20, 409–415. [Google Scholar] [CrossRef]
- Kim, I.H.; Kisseleva, T.; Brenner, D.A. Aging and liver disease. Curr. Opin. Gastroenterol. 2015, 31, 184–191. [Google Scholar] [CrossRef]
- Zhuang, Y.; Ma, Q.; Guo, Y.; Sun, L. Protective effects of rambutan (Nephelium lappaceum) peel phenolics on H2O2-induced oxidative damages in HepG2 cells and d-galactose-induced aging mice. Food Chem. Toxicol. 2017, 108, 554–562. [Google Scholar] [CrossRef]
- Xu, L.Q.; Xie, Y.L.; Gui, S.H.; Zhang, X.; Mo, Z.Z.; Sun, C.Y.; Li, C.L.; Luo, D.D.; Zhang, Z.B.; Su, Z.R.; et al. Polydatin attenuates d-galactose-induced liver and brain damage through its anti-oxidative, anti-inflammatory and anti-apoptotic effects in mice. Food Funct. 2016, 9, 4545–4555. [Google Scholar] [CrossRef] [PubMed]
- Hall, P.; Cash, J. What is the real function of the liver ‘function’ tests. Ulster Med. J. 2012, 81, 30–36. [Google Scholar]
- Chen, H.L.; Wang, C.H.; Kuo, Y.W.; Tsai, C.H. Antioxidative and hepatoprotective effects of fructo-oligosaccharide in d-galactose-treated Balb/cJ mice. Br. J. Nutr. 2011, 105, 805–809. [Google Scholar] [CrossRef]
- Shahroudi, M.J.; Mehri, S.; Hosseinzadeh, H. Anti-Aging Effect of Nigella Sativa Fixed Oil on D-Galactose-Induced Aging in Mice. J. Pharmacopunct. 2017, 20, 29–35. [Google Scholar]
- Yang, J.; He, Y.; Zou, J.; Xu, L.; Fan, F.; Ge, Z. Effect of Polygonum Multiflorum Thunb on liver fatty acid content in aging mice induced by D-galactose. Lipids Health Dis. 2019, 1, 128. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Zhao, Q.; Xiao, Y.; Liu, X.; Li, Y.; Zhang, J.; Pan, J.; Zhang, Z. Trehalose targets Nrf2 signal to alleviate d-galactose induced aging and improve behavioral ability. Biochem. Biophys. Res. Commun. 2020, 512, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Tarocco, A.; Caroccia, N.; Morciano, G.; Wieckowski, M.R.; Ancora, G.; Garani, G.; Pinton, P. Melatonin as a master regulator of cell death and inflammation: Molecular mechanisms and clinical implications for newborn care. Cell Death Dis. 2019, 8, 317. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Rosales-Corral, S.A.; Tan, D.X.; Acuna-Castroviejo, D.; Qin, L.; Yang, S.F.; Xu, K. Melatonin, a Full Service Anti-Cancer Agent: Inhibition of Initiation, Progression and Metastasis. Int. J. Mol. Sci. 2017, 17, 843. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Meng, F.; Francis, H.; Wu, N.; Chen, L.; Kennedy, L.; Zhou, T.; Franchitto, A.; Onori, P.; Gaudio, E.; et al. Melatonin and circadian rhythms in liver diseases: Functional roles and potential therapies. J. Pineal Res. 2020, 68, e12639. [Google Scholar] [CrossRef]
- Ferramosca, A.; Zara, V. Modulation of hepatic steatosis by dietary fatty acids. World J. Gastroenterol. 2014, 21, 1746–1755. [Google Scholar] [CrossRef]
- Ferramosca, A.; Zara, V. Dietary fat and hepatic lipogenesis: Mitochondrial citrate carrier as a sensor of metabolic changes. Adv. Nutr. 2014, 14, 217–225. [Google Scholar] [CrossRef]
- Musso, G.; Gambino, R.; Cassader, M. Recent insights into hepatic lipid metabolism in non-alcoholic fatty liver disease (NAFLD). Prog. Lipid Res. 2009, 48, 1–26. [Google Scholar] [CrossRef]
- Peet, D.J.; Turley, S.D.; Ma, W.; Janowski, B.A.; Lobaccaro, J.M.; Hammer, R.E.; Mangelsdorf, D.J. Cholesterol and bile acid metabolism are impaired in mice lacking the nuclear oxysterol receptor LXR alpha. Cell 1998, 29, 693–704. [Google Scholar] [CrossRef]
- Kotliarov, P.M. Potential for ultrasonic diagnosis in determining the nature of a bulky lesion of the liver. Med. Radiol. 1990, 35, 14–17. [Google Scholar]
- Ou, T.H.; Tung, Y.T.; Yang, T.H.; Chien, Y.W. Melatonin Improves Fatty Liver Syndrome by Inhibiting the Lipogenesis Pathway in Hamsters with High-Fat Diet-Induced Hyperlipidemia. Nutrients 2019, 30, 748. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, Y.; Chen, Q.; Liu, S.; Xu, W.; Shang, W.; Wang, L.; Yu, J. Melatonin Alleviates Glucose and Lipid Metabolism Disorders in Guinea Pigs Caused by Different Artificial Light Rhythms. J. Diabetes Res. 2020, 22, 4927403. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhang, C.; Zhao, M.; Shi, C.E.; Zhu, R.M.; Wang, H.; Zhao, H.; Wei, W.; Li, J.B.; Xu, D.X. Melatonin alleviates lipopolysaccharide-induced hepatic SREBP-1c activation and lipid accumulation in mice. J. Pineal Res. 2011, 51, 416–425. [Google Scholar] [CrossRef]
- Heo, J.I.; Yoon, D.W.; Yu, J.H.; Kim, N.H.; Yoo, H.J.; Seo, J.A.; Kim, S.G.; Choi, K.M.; Baik, S.H.; Choi, D.S.; et al. Melatonin improves insulin resistance and hepatic steatosis through attenuation of alpha-2-HS-glycoprotein. J. Pineal Res. 2018, 65, e12493. [Google Scholar] [CrossRef]
- Mooli, R.G.R.; Ramakrishnan, S.K. Liver Steatosis is a Driving Factor of Inflammation. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1267–1270. [Google Scholar] [CrossRef]
- Hritcu, L.; Bagci, E.; Aydin, E.; Mihasan, M. Antiamnesic and Antioxidants Effects of Ferulago angulata Essential Oil Against Scopolamine-Induced Memory Impairment in Laboratory Rats. Neurochem. Res. 2015, 40, 1799–1809. [Google Scholar] [CrossRef]
- Crespo, E.; Macías, M.; Pozo, D.; Escames, G.; Martín, M.; Vives, F.; Guerrero, J.M.; Acuña-Castroviejo, D. Melatonin inhibits expression of the inducible NO synthase II in liver and lung and prevents endotoxemia in lipopolysaccharide-induced multiple organ dysfunction syndrome in rats. FASEB J. 1999, 13, 1537–1546. [Google Scholar] [CrossRef]
- Rea, I.M.; Gibson, D.S.; McGilligan, V.; McNerlan, S.E.; Alexander, H.D.; Ross, O.A. Age and Age-Related Diseases: Role of Inflammation Triggers and Cytokines. Front. Immunol. 2018, 9, 586. [Google Scholar] [CrossRef] [PubMed]
- Freund, A.; Orjalo, A.V.; Desprez, P.Y.; Campisi, J. Inflammatory networks during cellular senescence: Causes and consequences. Trends Mol. Med. 2010, 16, 238–246. [Google Scholar] [CrossRef]
- Liu, C.; Chu, D.; Kalantar-Zadeh, K.; George, J.; Young, H.A.; Liu, G. Cytokines: From Clinical Significance to Quantification. Adv. Sci. 2021, 8, e2004433. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wei, X.; Wei, X.; Sun, X.; Huang, X.; Liang, Y.; Xu, W.; Zhu, X.; Lin, X.; Lin, J. 4-hydroxybenzo[d]oxazol-2(3H)-one ameliorates LPS/D-GalN-induced acute liver injury by inhibiting TLR4/NF-κB and MAPK signaling pathways in mice. Int. Immunopharmacol. 2020, 83, 106445. [Google Scholar] [CrossRef]
- Seif, M.; Deabes, M.; El-Askary, A.; El-Kott, A.F.; Albadrani, G.M.; Seif, A.; Wang, Z. Ephedra sinica mitigates hepatic oxidative stress and inflammation via suppressing the TLR4/MyD88/NF-κB pathway in fipronil-treated rats. Environ. Sci. Pollut. Res. Int. 2021, 28, 62943–62958. [Google Scholar] [CrossRef] [PubMed]
- Escribano, B.M.; Colín-González, A.L.; Santamaría, A.; Túnez, I. The role of melatonin in multiple sclerosis, Huntington’s disease and cerebral ischemia. CNS Neurol. Disord. Drug Targets 2014, 13, 1096–1119. [Google Scholar] [CrossRef]
- Gusarov, A.A.; Fetisov, V.A.; Kucheryavets, Y.O. Demonstration of the impossibility of simultaneous Dupuytren and Destot fractures in the result of the car-pedestrian collision. Sud. Med. Ekspertiza 2016, 59, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, D.M.S.; Simão, A.L.; Rodrigues, C.M.P.; Castro, R.E. Revisiting the metabolic syndrome and paving the way for microRNAs in non-alcoholic fatty liver disease. FEBS J. 2014, 281, 2503–2524. [Google Scholar] [CrossRef]
- Illingworth, R.S.; BirdMi, A.P. CpG islands—‘a rough guide’. FEBS Lett. 2009, 583, 1713–1720. [Google Scholar] [CrossRef]
- Gibbs, J.R.; van der Brug, M.P.; Hernandez, D.G.; Traynor, B.J.; Nalls, M.A.; Lai, S.L.; Arepalli, S.; Dillman, A.; Rafferty, I.P.; Troncoso, J.; et al. Abundant quantitative trait loci exist for DNA methylation and gene expression in human brain. PLoS Genet. 2010, 13, e1000952. [Google Scholar] [CrossRef] [PubMed]
- Tryndyak, V.p.; Han, T.; Muskhelishvili, L.; Fuscoe, J.C.; Ross, S.A.; Beland, F.A.; Pogribny, I.P. Coupling global methylation and gene expression profiles reveal key pathophysiological events in liver injury induced by a methyl-deficient diet. Mol. Nutr. Food Res. 2011, 55, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Cordero, P.; Campion, J.; Milagro, F.I.; Martinez, J.A. Transcriptomic and epigenetic changes in early liver steatosis associated to obesity: Effect of dietary methyl donor supplementation. Mol. Genet. Metab. 2013, 110, 388–395. [Google Scholar] [CrossRef]
- Cordero, P.; Gomez-Uriz, A.M.; Campion, J.; Milagro, F.I.; Martinez, J.A. Dietary supplementation with methyl donors reduces fatty liver and modifies the fatty acid synthase DNA methylation profile in rats fed an obesogenic diet. Genes Nutr. 2013, 8, 105–113. [Google Scholar] [CrossRef]
- Wolff, G.L.; Kodell, R.L.; Moore, S.R.; Cooney, C.A. Maternal epigenetics and methyl supplements affect agouti gene expression in Avy/a mice. FASEB J. 1998, 12, 949–957. [Google Scholar] [CrossRef]
- Dudley, K.J.; Sloboda, D.M.; Connor, K.L.; Beltrand, J.; Vickers, M.H. Offspring of mothers fed a high fat diet display hepatic cell cycle inhibition and associated changes in gene expression and DNA methylation. PLoS ONE 2011, 6, e21662. [Google Scholar] [CrossRef]
- Chen, H.C.; Chen, Y.Z.; Wang, C.H.; Lin, F.J. The nonalcoholic fatty liver disease-like phenotype and lowered serum VLDL are associated with decreased expression and DNA hypermethylation of hepatic ApoB in male offspring of ApoE deficient mothers fed a with Western diet. J. Nutr. Biochem. 2020, 77, 108319. [Google Scholar] [CrossRef] [PubMed]
- Rezaeejam, H.; Shirazi, A.; Izadi, P.; Bazzaz, J.T.; Ghazi-Khansari, M.; Valizadeh, M.; Tabesh, G.A. Radioprotective effect of melatonin on expression of Cdkn1a and Rad50 genes in rat peripheral blood. J. Cancer Res. Ther. 2018, 14, S1070–S1075. [Google Scholar] [PubMed]
- Sehgal, R.; Perfilyev, A.; Männistö, V.; Ågren, J.; Nilsson, E.; Käkelä, P.; Ling, C.; de Mello, V.D.; Pihlajamäki, J. Liver saturated fat content associates with hepatic DNA methylation in obese individuals. Clin. Epigenet. 2023, 15, 21. [Google Scholar] [CrossRef]
- Mi, Y.; Tan, D.; He, Y.; Zhou, X.; Zhou, Q.; Ji, S. Melatonin Modulates lipid Metabolism in HepG2 Cells Cultured in High Concentrations of Oleic Acid: AMPK Pathway Activation may Play an Important Role. Cell Biochem. Biophys. 2018, 76, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Yu, Z.; Jing, S.; Jiang, W.; Liu, C.; Yu, C.; Sun, J.; Wang, C.; Chen, J.; Li, H. Protective effect of Anwulignan against D-galactose-induced hepatic injury through activating p38 MAPK-Nrf2-HO-1 pathway in mice. Clin. Interv. Aging. 2018, 2, 1859–1869. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Company | Catalog Number | Dilution |
|---|---|---|---|
| NF-κB | Santa Cruz Biotech | sc8008 | 1:2000 |
| IL-6 | Bio-RAD | AHP424 | 1:500 |
| IL-1B | Bio-RAD | MCA1658 | 1:3000 |
| COX-2 | Santa Cruz Biotech | sc376861 | 1:2000 |
| β-actin | Santa Cruz Biotech | sc47778 | 1:2000 |
| Species | Gene | Forward Primer | Reverse Primer |
|---|---|---|---|
| Mouse | IL-1B | 5′-ACCTTCCAGGATGAGGACATGA-3′ | 5′-CTAATGGGAACGTCACACACCA-3′ |
| NF-κB | 5′-TGGAGTTCGTGACCGCCGCCG-3′ | 5′-GCTGGCTCTGCCGGGAAGATG-3′ | |
| IL-6 | 5′-TGATGCTGGTGACAACCACG-3 | 5′-CAGAATTGCCATTGCACAACTC-3′ | |
| TNF-α | 5′-CAGGCGGTGCCTATGTCTC-3′ | 5′-CGATCACCCCGAAGTTCAGTAG-3′ | |
| iNOS | 5′-GTTCTCAGCCCAACAATACAAGA-3′ | 5′-GTGGACGGGTCGATGTCAC-3′ | |
| Gapdh | 5′-GTCGGTGTGAACGGATTTG-3′ | 5′-CTTGCCGTGGGTAGAGTCAT-3′ | |
| Human | IL-1B | 5′- GTCCTGCGTGTTGAAAGATG-3′ | 5′-CTGCTTGAGAGGTGCTGATG-3′ |
| TNF-α | 5′-GGCGTGGAGCTGAGAGATAAC-3′ | 5′-TGATGGCAGAGAGGAGGTTG-3′ | |
| IL-6 | 5′-CCAAGGTCGGCTACTGAAAG-3′ | 5′-GATCCATAGCTGCGTGTCTTC-3′ | |
| iNOS | 5′-AACATCAGGTCGGCCATTAC-3′ | 5′-ACTGGGTGAACTCCAAGGTG-3′ | |
| IL-12 | 5′-CCTCCTCCTTGTGGCTACC-3′ | 5′-GAGTTTGTCTGGCCTTCTGG-3′ | |
| CCL2 | 5′-TCTGTGCCTGCTGCTCATAG-3′ | 5′-GAGTTTGTCTGGCCTTCTGG-3′ | |
| NF-kB | 5′-TCTGTGTTTGTCCAGCTTCG-3′ | 5′-AGCTCCAGCACCACTACCAC-3′ | |
| Gapdh | 5′-GGCTCTCCAGAACATCATCC-3′ | 5′-CCTGCTTCACCACCTTCTTG-3′ | |
| COX-2 | 5′-TGAGCATCTACGGTTTGCTG -3′ | 5′-TGCTTGTCTGGAACAACTGC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, R.; Lee, W.-Y.; Park, H.-J. Effects of Melatonin on Liver of D-Galactose-Induced Aged Mouse Model. Curr. Issues Mol. Biol. 2023, 45, 8412-8426. https://doi.org/10.3390/cimb45100530
Lee R, Lee W-Y, Park H-J. Effects of Melatonin on Liver of D-Galactose-Induced Aged Mouse Model. Current Issues in Molecular Biology. 2023; 45(10):8412-8426. https://doi.org/10.3390/cimb45100530
Chicago/Turabian StyleLee, Ran, Won-Yong Lee, and Hyun-Jung Park. 2023. "Effects of Melatonin on Liver of D-Galactose-Induced Aged Mouse Model" Current Issues in Molecular Biology 45, no. 10: 8412-8426. https://doi.org/10.3390/cimb45100530
APA StyleLee, R., Lee, W.-Y., & Park, H.-J. (2023). Effects of Melatonin on Liver of D-Galactose-Induced Aged Mouse Model. Current Issues in Molecular Biology, 45(10), 8412-8426. https://doi.org/10.3390/cimb45100530