
Extremophilic Solutions: The Role of Deinoxanthin in Counteracting UV-Induced Skin Harm

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
2.3. Cytotoxicity Assay
2.4. Assessing Deinoxanthin’s Protective Role in HFF-1 Cells Subjected to UVA and UVB Radiation
2.4.1. Assessment of Cell Viability
2.4.2. Apoptosis Assay
2.4.3. Determination of Tyrosinase Activity
2.4.4. Determination of Superoxide Dismutase Activity
2.4.5. Measuring Malondialdehyde Levels
2.4.6. Hydroxyproline Measurement
2.4.7. Intracellular Reactive Oxygen Species Quantification
2.4.8. Real-Time Monitoring and Assessment of Wound Healing Progress
2.5. Statistical Analysis
3. Results
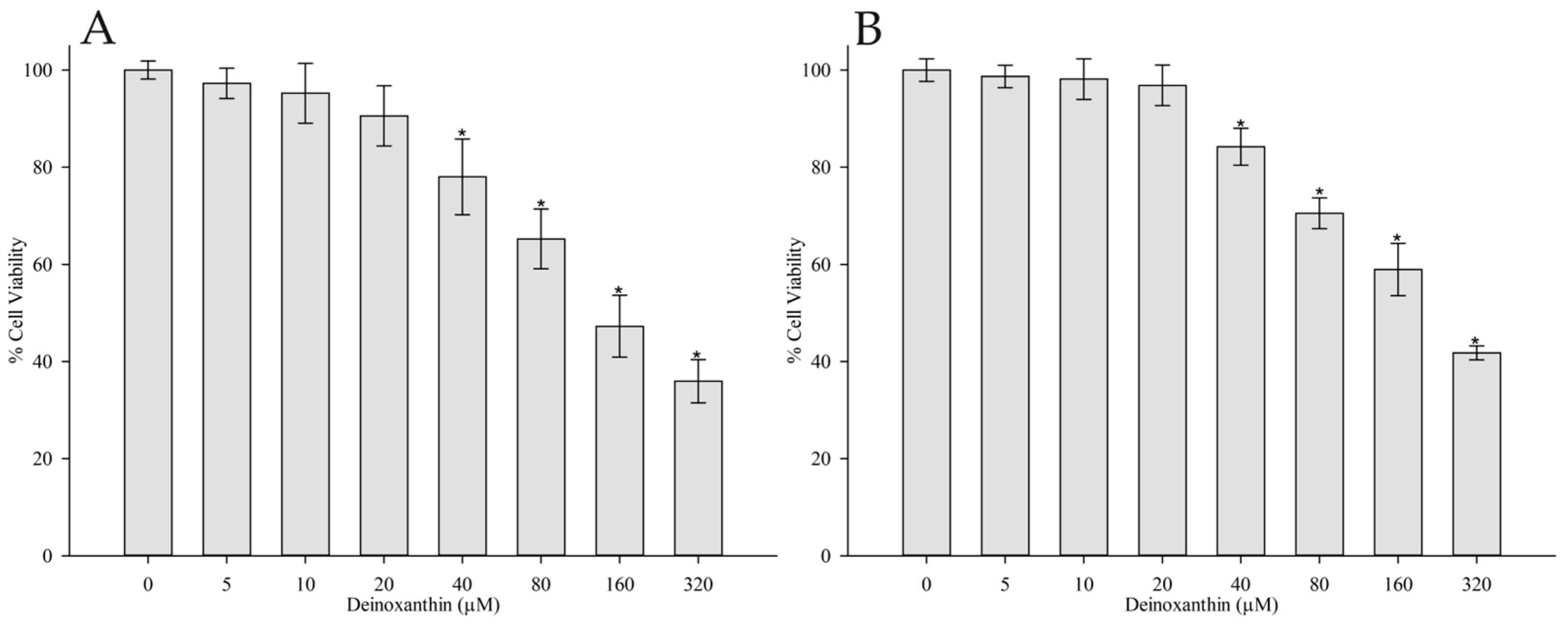
3.1. Cytotoxicity Assay
3.2. Evaluating the Protective Effects of Deinoxanthin on HFF-1 Cells Exposed to UVA and UVB Radiation
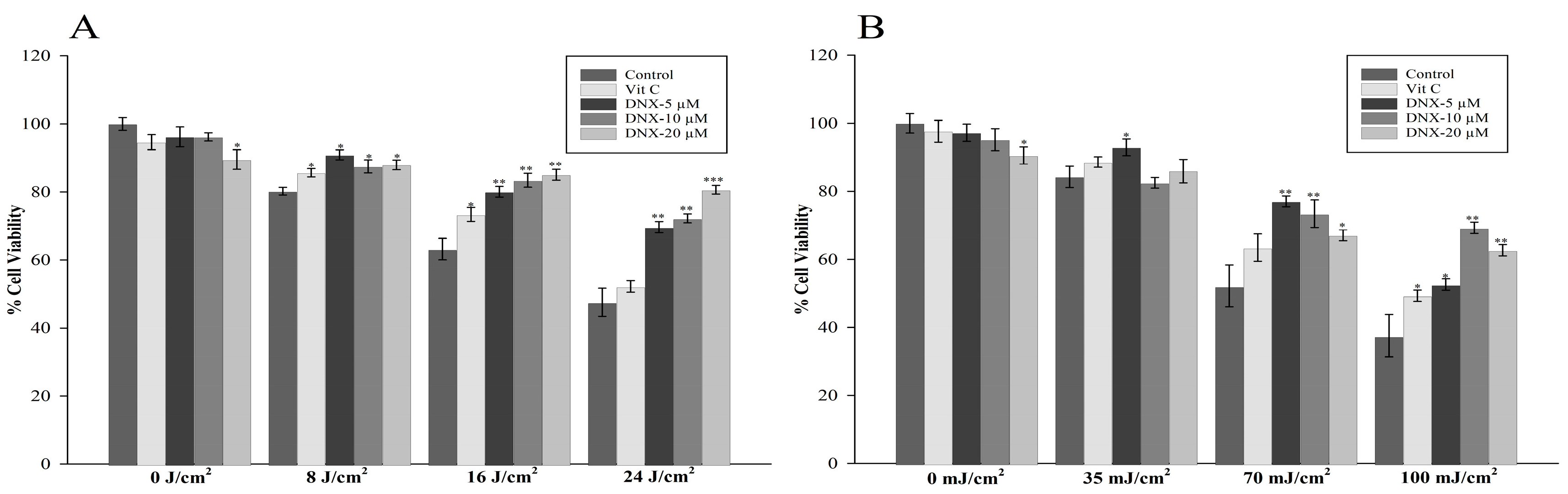
3.2.1. Cell Viability Assay
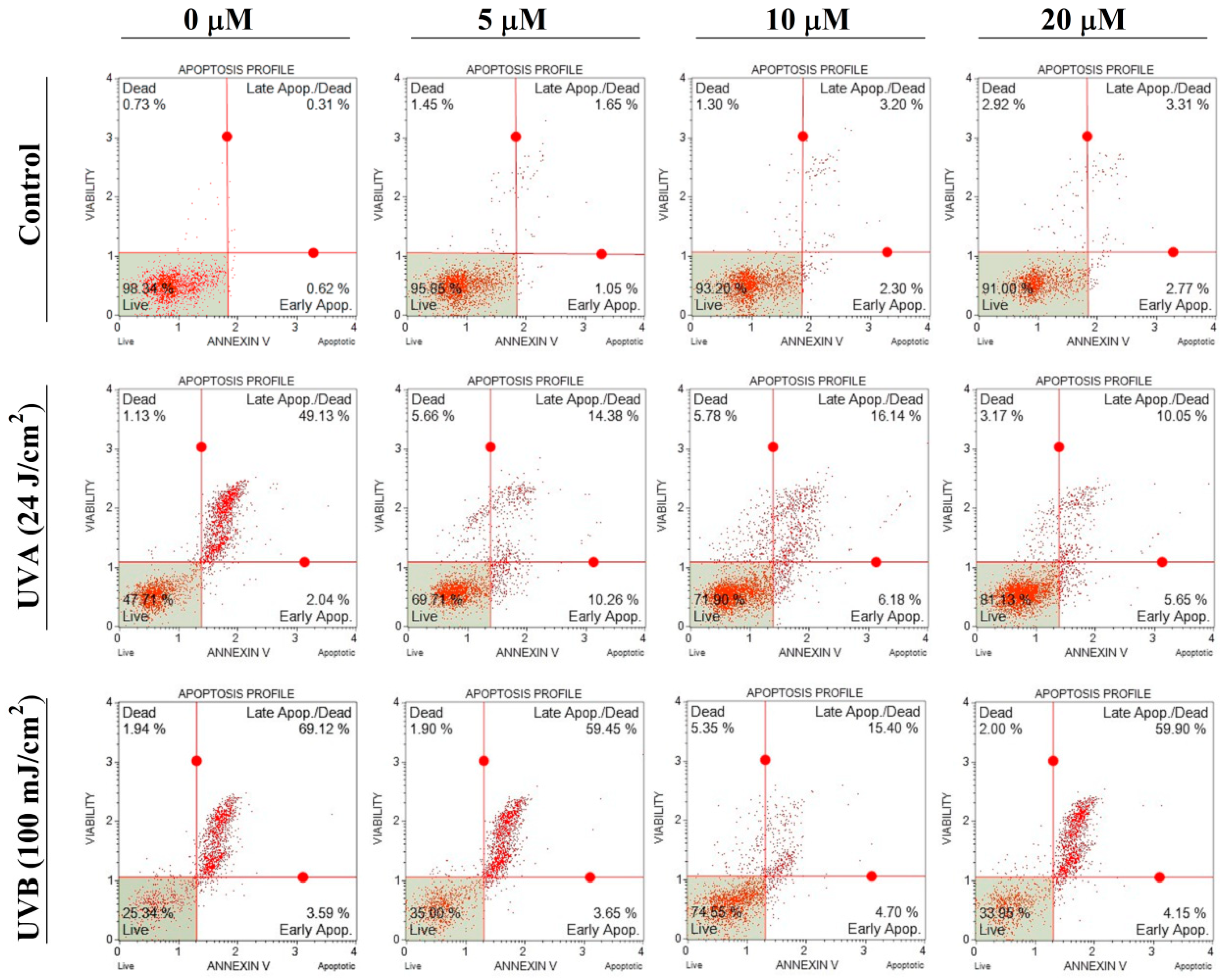
3.2.2. Apoptosis Assay
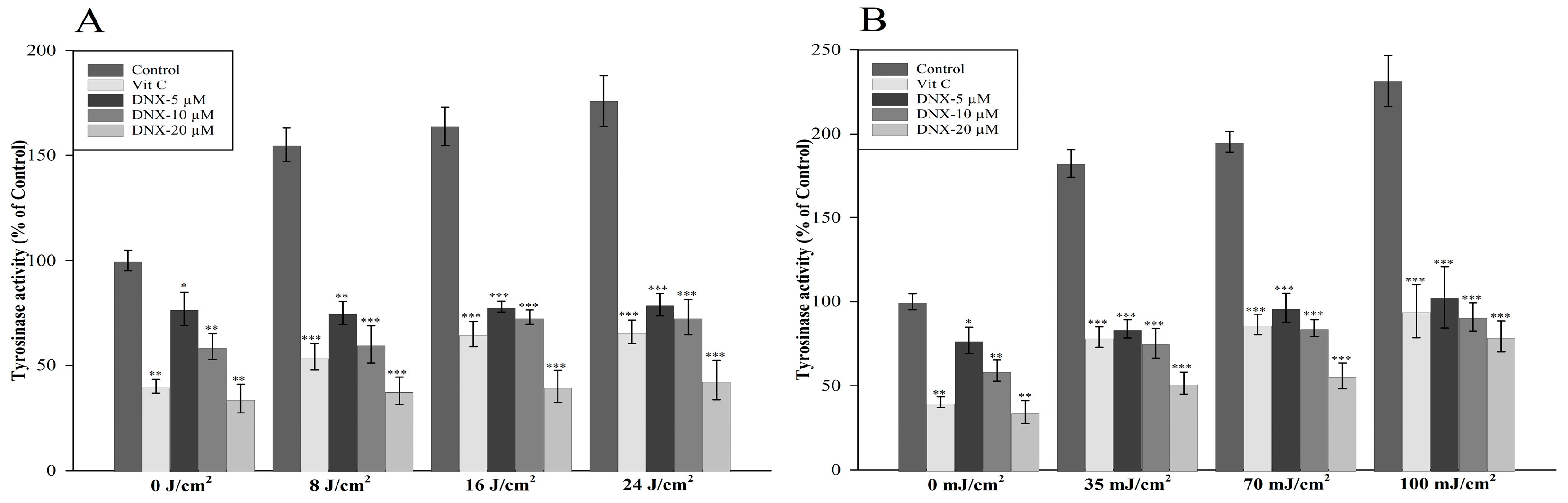
3.2.3. Tyrosinase Activity
3.2.4. Superoxide Dismutase Activity
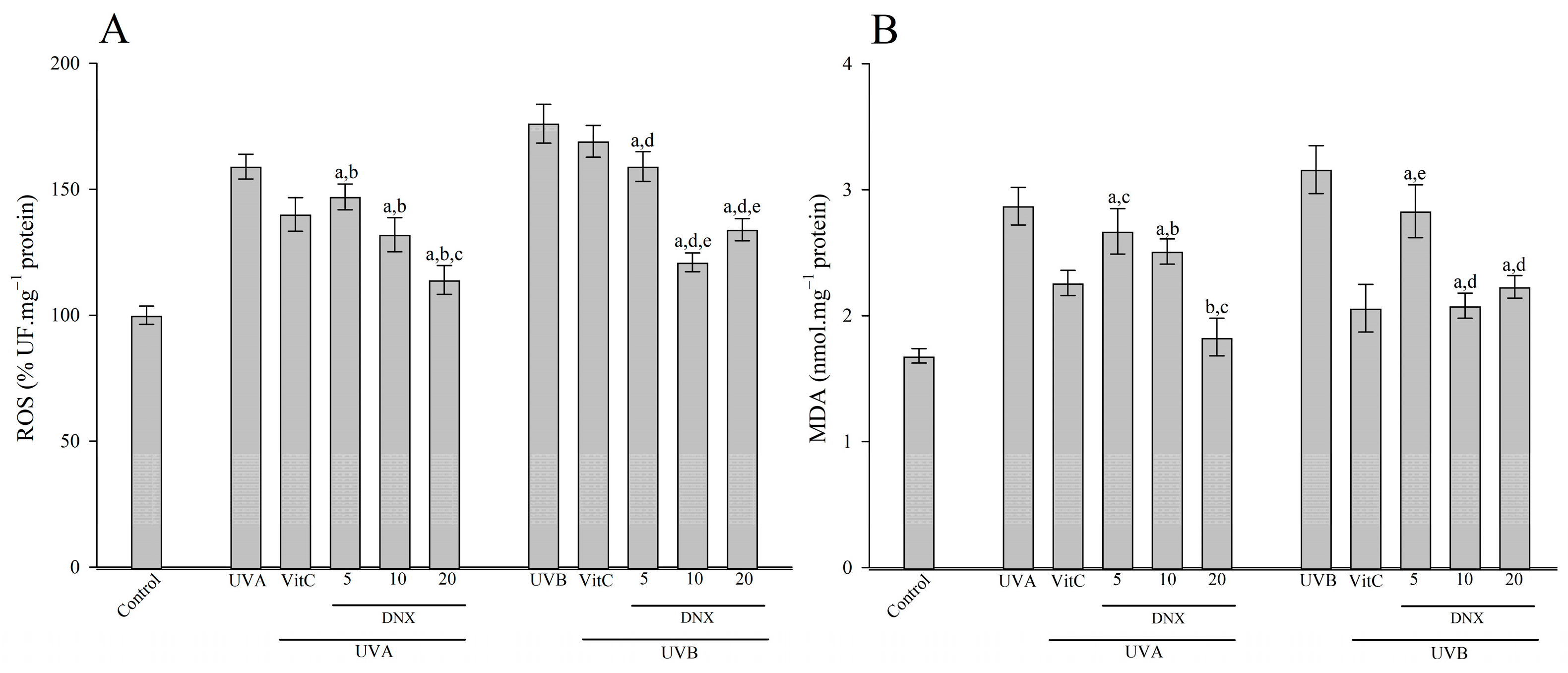
3.2.5. ROS and MDA Content
3.2.6. Hydroxyproline Measurement
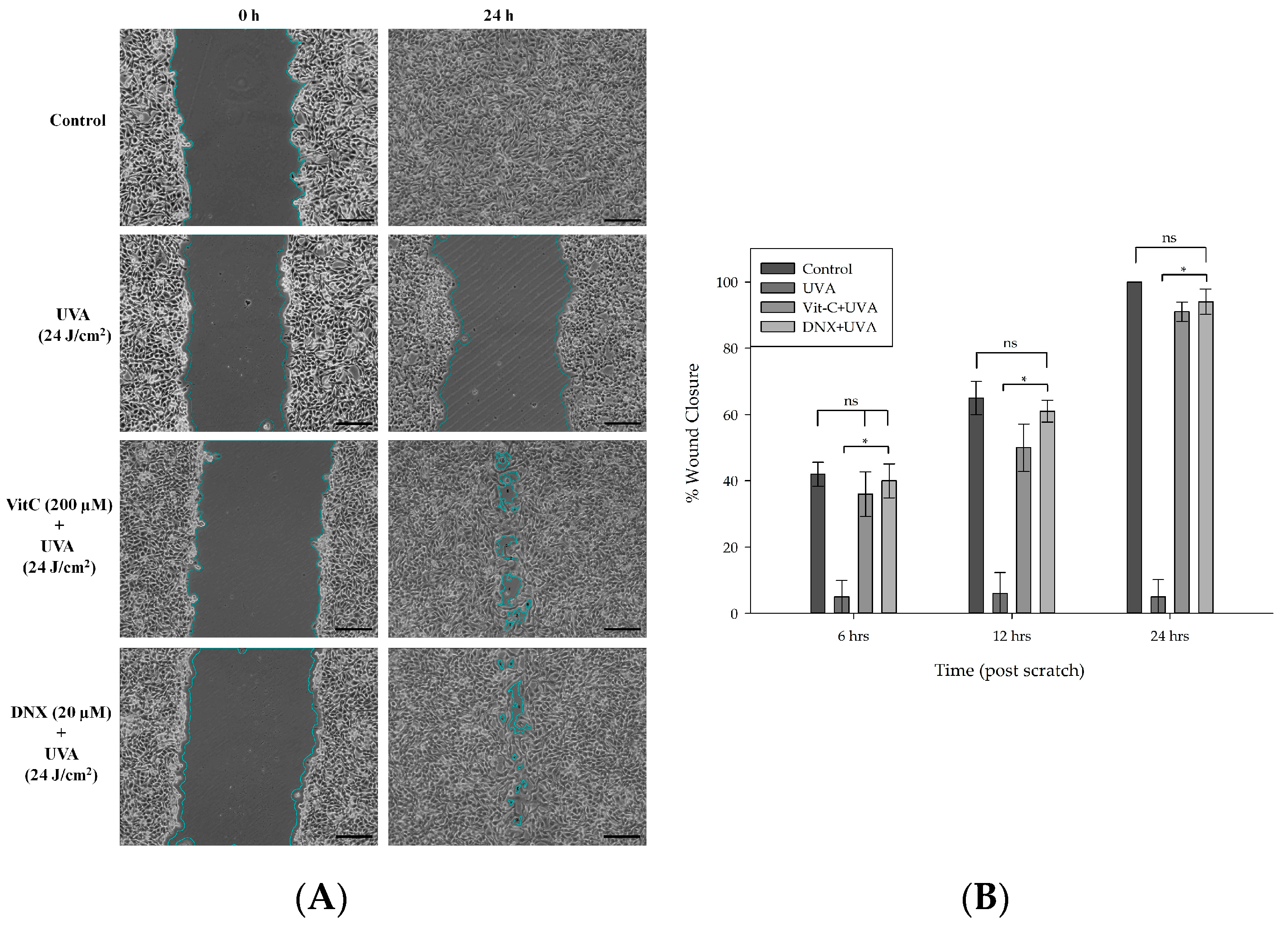
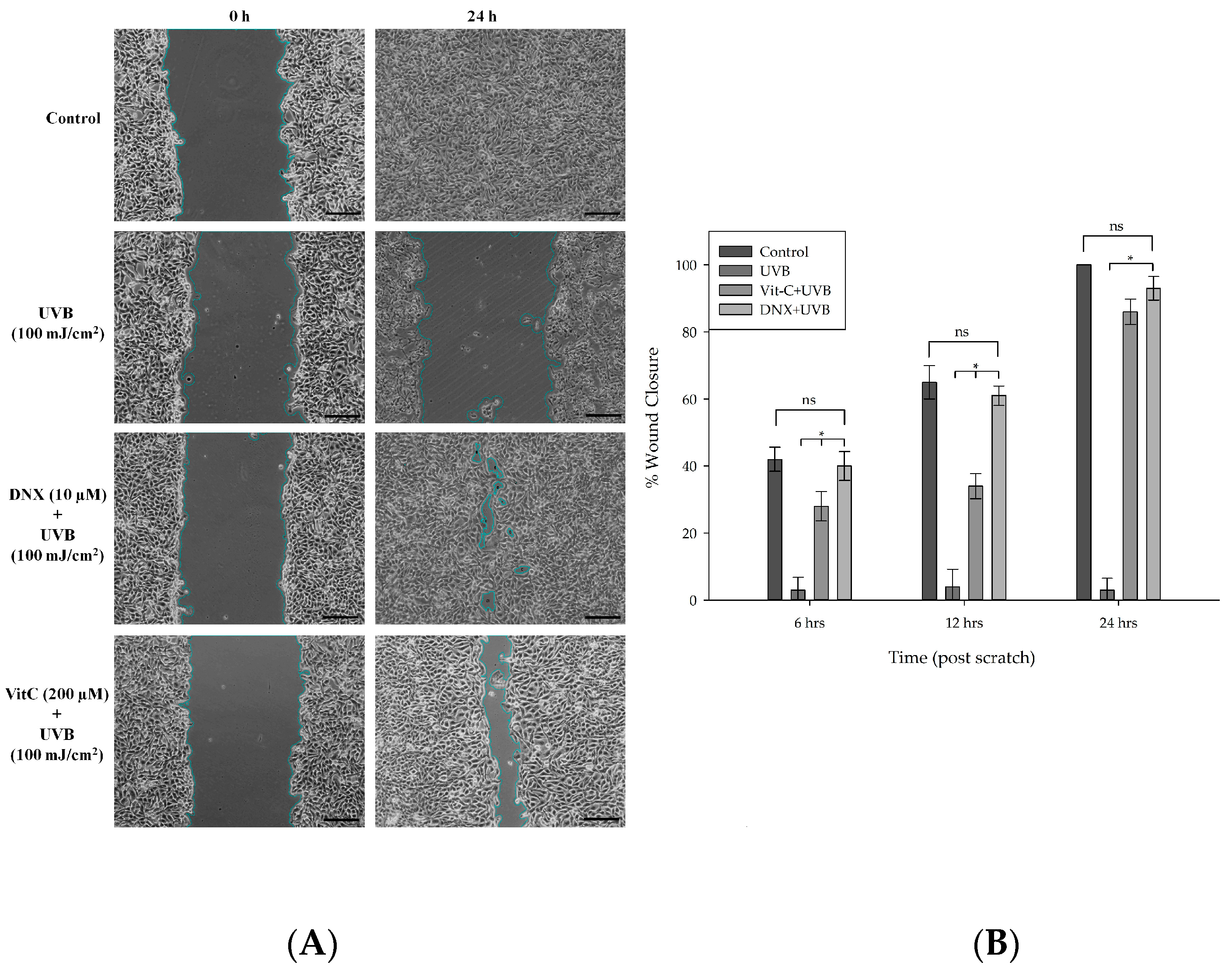
3.2.7. Wound Healing Assay
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Collaboration, P.; Ade, P.A.R.; Aghanim, N.; Alves, M.I.R.; Armitage-Caplan, C.; Arnaud, M.; Ashdown, M.; Atrio-Barandela, F.; Aumont, J.; Aussel, H.; et al. Planck 2013 results. I. Overview of products and scientific results. Astron. Astrophys. 2014, 571, A1. [Google Scholar] [CrossRef]
- Kochhar, N.; I․K, K.; Shrivastava, S.; Ghosh, A.; Rawat, V.S.; Sodhi, K.K.; Kumar, M. Perspectives on the microorganism of extreme environments and their applications. Curr. Res. Microb. Sci. 2022, 3, 100134. [Google Scholar] [CrossRef] [PubMed]
- Koklesova, L.; Liskova, A.; Samec, M.; Buhrmann, C.; Samuel, S.M.; Varghese, E.; Ashrafizadeh, M.; Najafi, M.; Shakibaei, M.; Büsselberg, D.; et al. Carotenoids in Cancer Apoptosis—The Road from Bench to Bedside and Back. Cancers 2020, 12, 2425. [Google Scholar] [CrossRef] [PubMed]
- Darvin, M.E.; Lademann, J.; von Hagen, J.; Lohan, S.B.; Kolmar, H.; Meinke, M.C.; Jung, S. Carotenoids in Human Skin In Vivo: Antioxidant and Photo-Protectant Role against External and Internal Stressors. Antioxidants 2022, 11, 1451. [Google Scholar] [CrossRef]
- McArdle, F.; Rhodes, L.E.; Parslew, R.A.; Close, G.L.; Jack, C.I.; Friedmann, P.S.; Jackson, M.J. Effects of oral vitamin E and β-carotene supplementation on ultraviolet radiation–induced oxidative stress in human skin. Am. J. Clin. Nutr. 2004, 80, 1270–1275. [Google Scholar] [CrossRef]
- Reis-Mansur, M.C.P.; Cardoso-Rurr, J.S.; Silva, J.V.M.A.; de Souza, G.R.; Cardoso, V.d.S.; Mansoldo, F.R.P.; Pinheiro, Y.; Schultz, J.; Lopez Balottin, L.B.; da Silva, A.J.R. Carotenoids from UV-resistant Antarctic Microbacterium sp. LEMMJ01. Sci. Rep. 2019, 9, 9554. [Google Scholar] [CrossRef]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef]
- Lim, S.; Jung, J.-H.; Blanchard, L.; de Groot, A. Conservation and diversity of radiation and oxidative stress resistance mechanisms in Deinococcus species. FEMS Microbiol. Rev. 2019, 43, 19–52. [Google Scholar] [CrossRef]
- Farci, D.; Slavov, C.; Tramontano, E.; Piano, D. The S-layer protein DR_2577 binds deinoxanthin and under desiccation conditions protects against UV-radiation in Deinococcus radiodurans. Front. Microbiol. 2016, 7, 155. [Google Scholar] [CrossRef]
- Reichrath, J.; Gordon-Thomson, C.; Tongkao-on, W.; Song, E.J.; Carter, S.E.; Dixon, K.M.; Mason, R.S. Protection from ultraviolet damage and photocarcinogenesis by vitamin D compounds. Sunlight Vitam. D Ski. Cancer 2014, 810, 303–328. [Google Scholar] [CrossRef]
- Chen, F.; Ji, H.J.; Choi, J.-I.; Han, S.H.; Lim, S.; Seo, H.S.; Ahn, K.B. Anti-allergic function of the cell wall (DeinoWall) from Deinococcus radiodurans. Mol. Immunol. 2022, 151, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Moens, E.; Veldhoen, M. Epithelial Barrier Biology: Good Fences Make Good Neighbours. Immunology 2011, 135, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Song, G.; Song, Z. Intrinsic Atopic Dermatitis and Extrinsic Atopic Dermatitis: Similarities and Differences. Clin. Cosmet. Investig. Dermatol. 2022, 15, 2621–2628. [Google Scholar] [CrossRef]
- Feng, S.; Song, G.; Liu, L.; Liu, W.; Liang, G.; Song, Z. Allergen-specific Immunotherapy Induces monocyte-derived Dendritic Cells but Attenuates Their Maturation and Cytokine Production in the Lesional Skin of an Atopic Dermatitis Mouse Model. J. Dermatol. 2022, 49, 1310–1319. [Google Scholar] [CrossRef]
- Mullenders, L.H. Solar UV damage to cellular DNA: From mechanisms to biological effects. Photochem. Photobiol. Sci. 2018, 17, 1842–1852. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Photoaging: UV radiation-induced inflammation and immunosuppression accelerate the aging process in the skin. Inflamm. Res. 2022, 71, 817–831. [Google Scholar] [CrossRef]
- Marabini, L.; Melzi, G.; Lolli, F.; Dell’Agli, M.; Piazza, S.; Sangiovanni, E.; Marinovich, M. Effects of Vitis vinifera L. leaves extract on UV radiation damage in human keratinocytes (HaCaT). J. Photochem. Photobiol. B Biol. 2020, 204, 111810. [Google Scholar] [CrossRef]
- Nichols, J.A.; Katiyar, S.K. Skin photoprotection by natural polyphenols: Anti-inflammatory, antioxidant and DNA repair mechanisms. Arch. Dermatol. Res. 2010, 302, 71–83. [Google Scholar] [CrossRef]
- Anbualakan, K.; Tajul Urus, N.Q.; Makpol, S.; Jamil, A.; Mohd Ramli, E.S.; Md Pauzi, S.H.; Muhammad, N. A Scoping Review on the Effects of Carotenoids and Flavonoids on Skin Damage Due to Ultraviolet Radiation. Nutrients 2022, 15, 92. [Google Scholar] [CrossRef]
- Gromkowska-Kępka, K.J.; Puścion-Jakubik, A.; Markiewicz-Żukowska, R.; Socha, K. The impact of ultraviolet radiation on skin photoaging—Review of in vitro studies. J. Cosmet. Dermatol. 2021, 20, 3427–3431. [Google Scholar] [CrossRef]
- Lu, W.; Kong, C.; Cheng, S.; Xu, X.; Zhang, J. Succinoglycan riclin relieves UVB-induced skin injury with anti-oxidant and anti-inflammatory properties. Int. J. Biol. Macromol. 2023, 235, 123717. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.-Z.; Li, D.-D.; Luo, H.; Li, W.-J.; Huang, Y.-M.; Li, J.-C.; Hu, Z.; Huang, N.; Guo, M.-H.; Chen, Y. Anti-photoaging effects of chitosan oligosaccharide in ultraviolet-irradiated hairless mouse skin. Exp. Gerontol. 2018, 103, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Jung, S.-H. Recent development of signaling pathways inhibitors of melanogenesis. Cell. Signal. 2017, 40, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Guan, L.L.; Lim, H.W.; Mohammad, T.F. Sunscreens and photoaging: A review of current literature. Am. J. Clin. Dermatol. 2021, 22, 819–828. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Li, A.; Li, S.; Tang, J.; Li, L.; Xiong, L. Natural components in sunscreens: Topical formulations with sun protection factor (SPF). Biomed. Pharmacother. 2020, 134, 111161. [Google Scholar] [CrossRef]
- Narla, S.; Lim, H.W. Sunscreen: FDA regulation, and environmental and health impact. Photochem. Photobiol. Sci. 2020, 19, 66–70. [Google Scholar] [CrossRef]
- Schneider, S.L.; Lim, H.W. Review of environmental effects of oxybenzone and other sunscreen active ingredients. J. Am. Acad. Dermatol. 2019, 80, 266–271. [Google Scholar] [CrossRef]
- Badmus, U.O.; Ač, A.; Klem, K.; Urban, O.; Jansen, M.A. A meta-analysis of the effects of UV radiation on the plant carotenoid pool. Plant Physiol. Biochem. 2022, 183, 36–45. [Google Scholar] [CrossRef]
- Kılınç, S. Purification and Optimization of Deinoxanthin Isolated from Deinococcus Radiodurans R1. Master’s Thesis, Marmara University Institute of Science and Technology, Istanbul, Türkiye, 2019. [Google Scholar]
- Saqib, S.; Nazeer, A.; Ali, M.; Zaman, W.; Younas, M.; Shahzad, A.; Sunera; Nisar, M. Catalytic Potential of Endophytes Facilitates Synthesis of Biometallic Zinc Oxide Nanoparticles for Agricultural Application. BioMetals 2022, 35, 967–985. [Google Scholar] [CrossRef]
- Kawashima, S.; Funakoshi, T.; Sato, Y.; Saito, N.; Ohsawa, H.; Kurita, K.; Nagata, K.; Yoshida, M.; Ishigami, A. Protective effect of pre-and post-vitamin C treatments on UVB-irradiation-induced skin damage. Sci. Rep. 2018, 8, 16199. [Google Scholar] [CrossRef]
- Savini, I.; D’Angelo, I.; Ranalli, M.; Melino, G.; Avigliano, L. Ascorbic acid maintenance in HaCaT revents radical formation and apoptosis by UV-B. Free Radic. Biol. Med. 1999, 26, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Noh, D.; Choi, J.G.; Huh, E.; Oh, M.S. Tectorigenin, a flavonoid-based compound of leopard lily rhizome, attenuates UV-B-induced apoptosis and collagen degradation by inhibiting oxidative stress in human keratinocytes. Nutrients 2018, 10, 1998. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Kim, H.N.; Kim, J.E.; Mun, G.H.; Kim, Y.S.; Cho, D.; Shin, D.H.; Hwang, Y.-I.; Lee, W.J. Regulation of UVB-induced IL-8 and MCP-1 production in skin keratinocytes by increasing vitamin C uptake via the redistribution of SVCT-1 from the cytosol to the membrane. J. Investig. Dermatol. 2007, 127, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Bian, Y.; Cheng, Y.; Sun, R.; Li, G. Effect of lemon peel flavonoids on UVB-induced skin damage in mice. RSC Adv. 2020, 10, 31470–31478. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Pabón, W.; Schlievert, P.M. Aortic valve damage for the study of left-sided, native valve infective endocarditis in rabbits. Superantigens Methods Protoc. 2016, 1396, 73–80. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovich, I. Superoxide dismutase: An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Uchiyama, M.; Mihara, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef]
- Zhang, C.; Lv, J.; Qin, X.; Peng, Z.; Lin, H. Novel Antioxidant Peptides from Crassostrea Hongkongensis Improve Photo-Oxidation in UV-Induced HaCaT Cells. Mar. Drugs 2022, 20, 100. [Google Scholar] [CrossRef]
- Günay, N.; Kuzucu, M. Agonistic Effects of Deinoxanthin on Tamoxifen Antiproliferative Activity on HER2 Positive Breast Cancer: An In Vitro Study on MDA-MB-453. Erzincan Univ. J. Sci. Technol. 2023, 16, 138–154. [Google Scholar] [CrossRef]
- Tian, B.; Li, J.; Pang, R.; Dai, S.; Li, T.; Weng, Y.; Jin, Y.; Hua, Y. Gold nanoparticles biosynthesized and functionalized using a hydroxylated tetraterpenoid trigger gene expression changes and apoptosis in cancer cells. ACS Appl. Mater. Interfaces 2018, 10, 37353–37363. [Google Scholar] [CrossRef]
- Farci, D.; Haniewicz, P.; de Sanctis, D.; Iesu, L.; Kereïche, S.; Winterhalter, M.; Piano, D. The cryo-EM structure of the S-layer deinoxanthin-binding complex of Deinococcus radiodurans informs properties of its environmental interactions. J. Biol. Chem. 2022, 298, 102031. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.-W.; Kim, J.-H.; Kim, J.-W.; Kim, C.Y.; Kim, S.Y.; Choi, Y.J. Metabolic engineering of extremophilic bacterium Deinococcus radiodurans for the production of the novel carotenoid deinoxanthin. Microorganisms 2020, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Asaka, R.; Ohshima, K.; Kawasaki, S.; Maoka, T.; Tode, C.; Wang-Otomo, Z.-Y.; Takaichi, S. Major Carotenoids of Meiothermus ruber Are Deinoxanthin Glucoside Esters, Not Meiothermoxanthin Glucoside Esters. J. Nat. Prod. 2022, 85, 2266–2273. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Kim, S.; Kim, J.; Kim, J.-W.; Kim, S.Y.; Yeom, B.; Kim, H.; Choi, W.I.; Sung, D. Highly Water-Dispersed and Stable Deinoxanthin Nanocapsule for Effective Antioxidant and Anti-Inflammatory Activity. Int. J. Nanomed. 2023, 18, 4555–4565. [Google Scholar] [CrossRef] [PubMed]
- Han, J.M.; Song, H.-Y.; Jung, J.-H.; Lim, S.; Seo, H.S.; Kim, W.S.; Lim, S.-T.; Byun, E.-B. Deinococcus radiodurans-derived membrane vesicles protect HaCaT cells against H2O2-induced oxidative stress via modulation of MAPK and Nrf2/ARE pathways. Biol. Proced. Online 2023, 25, 17. [Google Scholar] [CrossRef]
- Struwe, M.; Greulich, K.-O.; Junker, U.; Jean, C.; Zimmer, D.; Sutera, W.; Plappert-Helbig, U. Detection of photogenotoxicity in skin and eye in rat with the photo comet assay. Photochem. Photobiol. Sci. 2008, 7, 240–249. [Google Scholar] [CrossRef]
- Sheng, J.; Ding, S.; Liao, H.; Yao, Y.; Zhai, Y.; Zhan, J.; Wang, X. Polyacrylonitrile/UV329/titanium oxide composite nanofibrous membranes with enhanced UV protection and filtration performance. RSC Adv. 2023, 13, 17622–17627. [Google Scholar] [CrossRef]
- Agar, N.S.; Halliday, G.M.; Barnetson, R.S.; Ananthaswamy, H.N.; Wheeler, M.; Jones, A.M. The basal layer in human squamous tumors harbors more UVA than UVB fingerprint mutations: A role for UVA in human skin carcinogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 4954–4959. [Google Scholar] [CrossRef]
- Barrette, K.; Zutterman, N.; Van Kelst, S.; Proby, C.; Garmyn, M. Pattern of sensitivity of progressive cutaneous squamous cell carcinoma cells to UVB and oxidative stress-induced cell death. Photochem. Photobiol. Sci. 2013, 12, 104–110. [Google Scholar] [CrossRef]
- Dazard, J.-E.; Gal, H.; Amariglio, N.; Rechavi, G.; Domany, E.; Givol, D. Genome-wide comparison of human keratinocyte and squamous cell carcinoma responses to UVB irradiation: Implications for skin and epithelial cancer. Oncogene 2003, 22, 2993–3006. [Google Scholar] [CrossRef]
- Kidd, P. Astaxanthin, cell membrane nutrient with diverse clinical benefits and anti-aging potential. Altern Med. Rev. 2011, 16, 355–364. [Google Scholar] [PubMed]
- O’Connor, I.; O’Brien, N. Modulation of UVA light-induced oxidative stress by β-carotene, lutein and astaxanthin in cultured fibroblasts. J. Dermatol. Sci. 1998, 16, 226–230. [Google Scholar] [CrossRef] [PubMed]
- Santocono, M.; Zurria, M.; Berrettini, M.; Fedeli, D.; Falcioni, G. Influence of astaxanthin, zeaxanthin and lutein on DNA damage and repair in UVA-irradiated cells. J. Photochem. Photobiol. B Biol. 2006, 85, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Radrezza, S.; Carini, M.; Baron, G.; Aldini, G.; Negre-Salvayre, A.; D’Amato, A. Study of Carnosine’s effect on nude mice skin to prevent UV-A damage. Free Radic. Biol. Med. 2021, 173, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Giampieri, F.; Alvarez-Suarez, J.M.; Tulipani, S.; Gonzàles-Paramàs, A.M.; Santos-Buelga, C.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Battino, M. Photoprotective potential of strawberry (Fragaria × ananassa) extract against UV-A irradiation damage on human fibroblasts. J. Agric. Food Chem. 2012, 60, 2322–2327. [Google Scholar] [CrossRef]
- Chung, Y.H.; Jeong, S.A.; Choi, H.S.; Ro, S.; Lee, J.S.; Park, J.K. Protective effects of ginsenoside Rg2 and astaxanthin mixture against UVB-induced DNA damage. Anim. Cells Syst. 2018, 22, 400–406. [Google Scholar] [CrossRef]
- Tomasello, B.; Malfa, G.A.; Acquaviva, R.; La Mantia, A.; Di Giacomo, C. Phytocomplex of a Standardized Extract from Red Orange (Citrus sinensis L. Osbeck) against Photoaging. Cells 2022, 11, 1447. [Google Scholar] [CrossRef]
- Kulms, D.; Schwarz, T. Molecular mechanisms of UV-induced apoptosis. Photodermatol. Photoimmunol. Photomed. Rev. Artic. 2000, 16, 195–201. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, J.E.; Choi, Y.J.; Gong, J.E.; Park, S.H.; Douangdeuane, B.; Souliya, O.; Park, J.M.; Lee, H.S.; Kim, B.-H. Therapeutic effects of Dipterocarpus tuberculatus with high antioxidative activity against UV-induced photoaging of NHDF cells and nude mice. Antioxidants 2021, 10, 791. [Google Scholar] [CrossRef]
- Lee, S.J.; Kim, J.E.; Choi, Y.J.; Jin, Y.J.; Roh, Y.J.; Seol, A.Y.; Song, H.J.; Park, S.H.; Uddin, M.S.; Lee, S.W. Antioxidative role of Hygrophila erecta (Brum. F.) Hochr. on UV-induced photoaging of dermal fibroblasts and melanoma cells. Antioxidants 2022, 11, 1317. [Google Scholar] [CrossRef]
- Guo, W.; An, Y.; Jiang, L.; Geng, C.; Zhong, L. The protective effects of hydroxytyrosol against UVB-induced DNA damage in HaCaT cells. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2010, 24, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.H.; Hwang, S.T.; Um, J.-Y.; Ahn, K.S. Cycloastragenol exerts protective effects against UVB irradiation in human dermal fibroblasts and HaCaT keratinocytes. J. Dermatol. Sci. 2023, 111, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Wang, F.; Chen, Y. Protective effect and mechanism research of Phyllanthus emblica Linn. fruit extract on UV-induced photodamage in keratinocytes. Photochem. Photobiol. Sci. 2023, 22, 1945–1959. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-J.; Hur, J.-M.; Lim, S.; Jo, M.; Kim, D.H.; Choi, J.-I. Induction of apoptosis by deinoxanthin in human cancer cells. Anticancer Res. 2014, 34, 1829–1835. [Google Scholar] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Jung, H.; Kim, K.; Lim, K.M.; Kim, J.y.; Jho, E.h.; Oh, E.S. D-tyrosine negatively regulates melanin synthesis by competitively inhibiting tyrosinase activity. Pigment. Cell Melanoma Res. 2018, 31, 374–383. [Google Scholar] [CrossRef]
- Slominski, A.; Zmijewski, M.A.; Pawelek, J. L-tyrosine and L-dihydroxyphenylalanine as hormone-like regulators of melanocyte functions. Pigment. Cell Melanoma Res. 2012, 25, 14–27. [Google Scholar] [CrossRef]
- Young, A.J.; Lowe, G.L. Carotenoids—Antioxidant properties. Antioxidants 2018, 7, 28. [Google Scholar] [CrossRef]
- Mordi, R.C.; Ademosun, O.T.; Ajanaku, C.O.; Olanrewaju, I.O.; Walton, J.C. Free radical mediated oxidative degradation of carotenes and xanthophylls. Molecules 2020, 25, 1038. [Google Scholar] [CrossRef]
- Zaripheh, S.; Erdman, J.W., Jr. Factors that influence the bioavailablity of xanthophylls. J. Nutr. 2002, 132, 531S–534S. [Google Scholar] [CrossRef]
- Nakajima, H.; Fukazawa, K.; Wakabayashi, Y.; Wakamatsu, K.; Senda, K.; Imokawa, G. Abrogating effect of a xanthophyll carotenoid astaxanthin on the stem cell factor-induced stimulation of human epidermal pigmentation. Arch. Dermatol. Res. 2012, 304, 803–816. [Google Scholar] [CrossRef]
- Kim, H.M.; An, H.S.; Bae, J.-S.; Kim, J.Y.; Choi, C.H.; Kim, J.Y.; Lim, J.H.; Choi, J.-h.; Song, H.; Moon, S.H. Effects of palmitoyl-KVK-L-ascorbic acid on skin wrinkles and pigmentation. Arch. Dermatol. Res. 2017, 309, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Miao, F.; Su, M.-Y.; Jiang, S.; Luo, L.-F.; Shi, Y.; Lei, T.-C. Intramelanocytic acidification plays a role in the antimelanogenic and antioxidative properties of vitamin C and its derivatives. Oxidative Med. Cell. Longev. 2019, 2019, 2084805. [Google Scholar] [CrossRef] [PubMed]
- Shimada, Y.; Tai, H.; Tanaka, A.; Ikezawa-Suzuki, I.; Takagi, K.; Yoshida, Y.; Yoshie, H. Effects of ascorbic acid on gingival melanin pigmentation in vitro and in vivo. J. Periodontol. 2009, 80, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Panich, U.; Tangsupa-A-Nan, V.; Onkoksoong, T.; Kongtaphan, K.; Kasetsinsombat, K.; Akarasereenont, P.; Wongkajornsilp, A. Inhibition of UVA-mediated melanogenesis by ascorbic acid through modulation of antioxidant defense and nitric oxide system. Arch. Pharmacal Res. 2011, 34, 811–820. [Google Scholar] [CrossRef]
- Deng, H.; Wan, M.; Li, H.; Chen, Q.; Li, R.; Liang, B.; Zhu, H. Curcumin protection against ultraviolet-induced photo-damage in Hacat cells by regulating nuclear factor erythroid 2-related factor 2. Bioengineered 2021, 12, 9993–10006. [Google Scholar] [CrossRef]
- Kobayashi, T.; Matsumoto, M.; Iizuka, H.; Suzuki, K.; Taniguchi, N. Superoxide dismutase in psoriasis, squamous cell carcinoma and basal cell epithelioma: An immunohistochemical study. Br. J. Dermatol. 1991, 124, 555–559. [Google Scholar] [CrossRef]
- Kobayashi, T.; Saito, N.; Takemori, N.; Iizuka, S.; Suzuki, K.; Taniguchi, N.; Iizuka, H. Ultrastructural localization of superoxide dismutase in human skin. Acta Derm. Venereol. 1993, 73, 41–45. [Google Scholar] [CrossRef]
- Sasaki, H.; Akamatsu, H.; Horio, T. Protective role of copper, zinc superoxide dismutase against UVB-induced injury of the human keratinocyte cell line HaCaT. J. Investig. Dermatol. 2000, 114, 502–507. [Google Scholar] [CrossRef]
- Lyons, N.M.; O’Brien, N.M. Modulatory effects of an algal extract containing astaxanthin on UVA-irradiated cells in culture. J. Dermatol. Sci. 2002, 30, 73–84. [Google Scholar] [CrossRef]
- Ansary, T.M.; Hossain, M.R.; Kamiya, K.; Komine, M.; Ohtsuki, M. Inflammatory molecules associated with ultraviolet radiation-mediated skin aging. Int. J. Mol. Sci. 2021, 22, 3974. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.D.; Bermudez, Y.; Park, S.L.; Stratton, S.P.; Uchida, K.; Hurst, C.A.; Wondrak, G.T. Malondialdehyde-derived epitopes in human skin result from acute exposure to solar UV and occur in nonmelanoma skin cancer tissue. J. Photochem. Photobiol. B Biol. 2014, 132, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Çavuşoğlu, K.; Kalefetoğlu Macar, T.; Macar, O.; Çavuşoğlu, D.; Yalçın, E. Comparative investigation of toxicity induced by UV-A and UV-C radiation using allium test. Environ. Sci. Pollut. Res. 2022, 29, 33988–33998. [Google Scholar] [CrossRef]
- Yakaew, S.; Itsarasook, K.; Ngoenkam, J.; Jessadayannamaetha, A.; Viyoch, J.; Ungsurungsie, M. Ethanol extract of Terminalia chebula fruit protects against UVB-induced skin damage. Pharm. Biol. 2016, 54, 2701–2707. [Google Scholar] [CrossRef] [PubMed]
- Faikrua, A.; Jeenapongsa, R.; Sila-Asna, M.; Viyoch, J. Properties of β-glycerol phosphate/collagen/chitosan blend scaffolds for application in skin tissue engineering. ScienceAsia 2009, 35, 247–254. [Google Scholar] [CrossRef]
- Inpanya, P.; Faikrua, A.; Ounaroon, A.; Sittichokechaiwut, A.; Viyoch, J. Effects of the blended fibroin/aloe gel film on wound healing in streptozotocin-induced diabetic rats. Biomed. Mater. 2012, 7, 035008. [Google Scholar] [CrossRef]
- Zhang, L.; Liang, S.; Zhang, Z.; Wang, K.; Cao, J.; Yao, M.; Qin, L.; Qu, C.; Miao, J. Protective Effects of ζ-Carotene-like Compounds against Acute UVB-Induced Skin Damage. Int. J. Mol. Sci. 2023, 24, 13970. [Google Scholar] [CrossRef]
- Martin, P. Wound healing--aiming for perfect skin regeneration. Science 1997, 276, 75–81. [Google Scholar] [CrossRef]
- Shaw, T.J.; Martin, P. Wound repair at a glance. J. Cell Sci. 2009, 122, 3209–3213. [Google Scholar] [CrossRef]
- Singer, A.J.; Clark, R.A. Cutaneous wound healing. N. Engl. J. Med. 1999, 341, 738–746. [Google Scholar] [CrossRef]
- Willenborg, S.; Knipper, J.; Ranjan, R.; Krieg, T.; Eming, S. Chronic wounds and inflammation. Adv. Wound Care 2010, 1, 259. [Google Scholar]
- Schultz, G.S.; Davidson, J.M.; Kirsner, R.S.; Bornstein, P.; Herman, I.M. Dynamic reciprocity in the wound microenvironment. Wound Repair Regen. 2011, 19, 134–148. [Google Scholar] [CrossRef]
- Sepp, K.J.; Auld, V.J. RhoA and Rac1 GTPases mediate the dynamic rearrangement of actin in peripheral glia. Development 2003, 130, 1825–1835. [Google Scholar] [CrossRef]
- Enciso, J.M.; Konecny, C.M.; Karpen, H.E.; Hirschi, K.K. Endothelial cell migration during murine yolk sac vascular remodeling occurs by means of a Rac1 and FAK activation pathway in vivo. Dev. Dyn. 2010, 239, 2570–2583. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, J.; Jiang, W.; Zhang, S. Astaxanthin induces angiogenesis through Wnt/β-catenin signaling pathway. Phytomedicine 2015, 22, 744–751. [Google Scholar] [CrossRef]
- Hodge, R.G.; Ridley, A.J. Regulating Rho GTPases and their regulators. Nat. Rev. Mol. Cell Biol. 2016, 17, 496–510. [Google Scholar] [CrossRef]
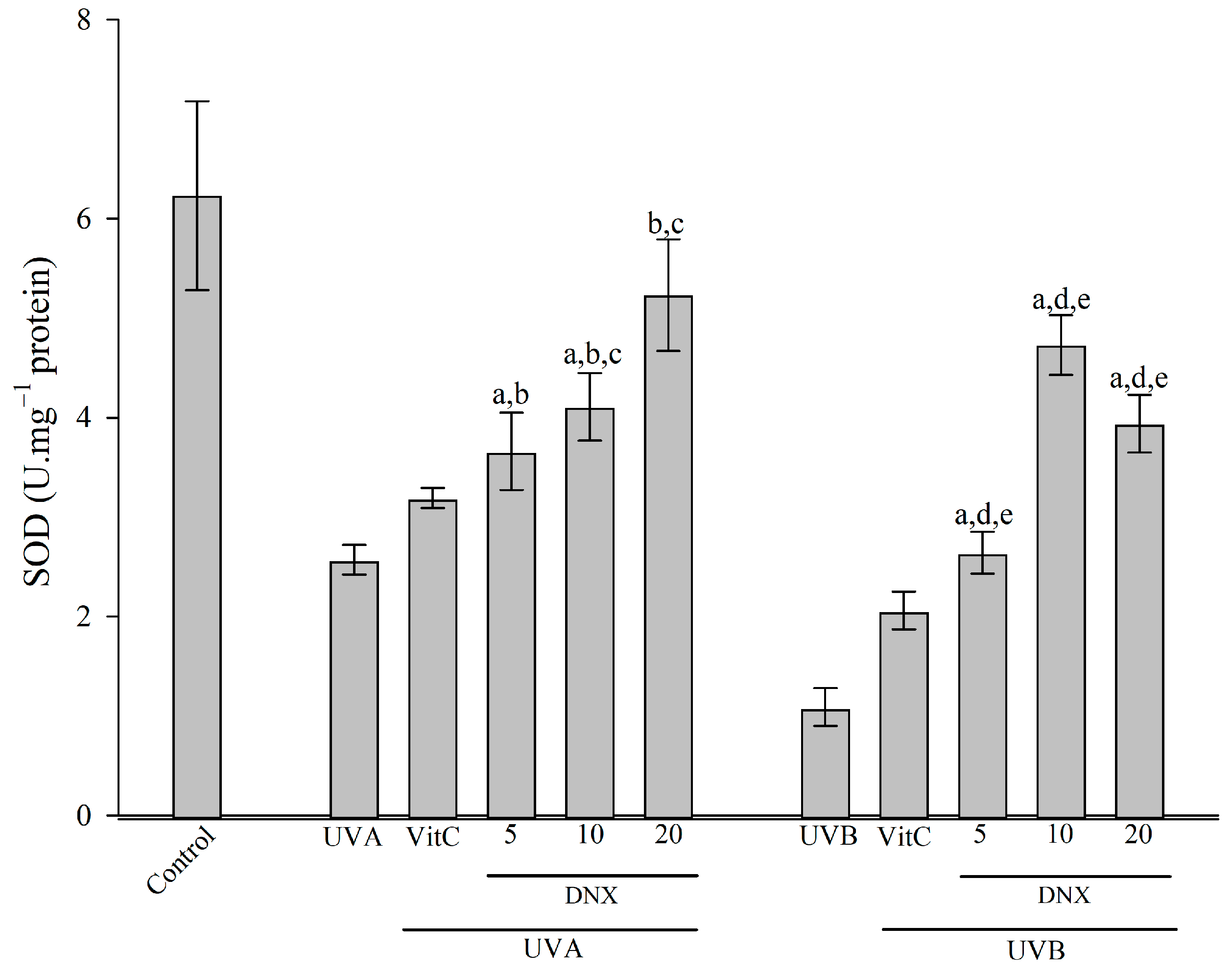

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuzucu, M. Extremophilic Solutions: The Role of Deinoxanthin in Counteracting UV-Induced Skin Harm. Curr. Issues Mol. Biol. 2023, 45, 8372-8394. https://doi.org/10.3390/cimb45100528
Kuzucu M. Extremophilic Solutions: The Role of Deinoxanthin in Counteracting UV-Induced Skin Harm. Current Issues in Molecular Biology. 2023; 45(10):8372-8394. https://doi.org/10.3390/cimb45100528
Chicago/Turabian StyleKuzucu, Mehmet. 2023. "Extremophilic Solutions: The Role of Deinoxanthin in Counteracting UV-Induced Skin Harm" Current Issues in Molecular Biology 45, no. 10: 8372-8394. https://doi.org/10.3390/cimb45100528
APA StyleKuzucu, M. (2023). Extremophilic Solutions: The Role of Deinoxanthin in Counteracting UV-Induced Skin Harm. Current Issues in Molecular Biology, 45(10), 8372-8394. https://doi.org/10.3390/cimb45100528