
Gender Differences in Insulin Resistance: New Knowledge and Perspectives


Abstract
:1. Background
2. Methods
3. Prevalence
4. Body Fat Distribution
5. Hormonal Factors
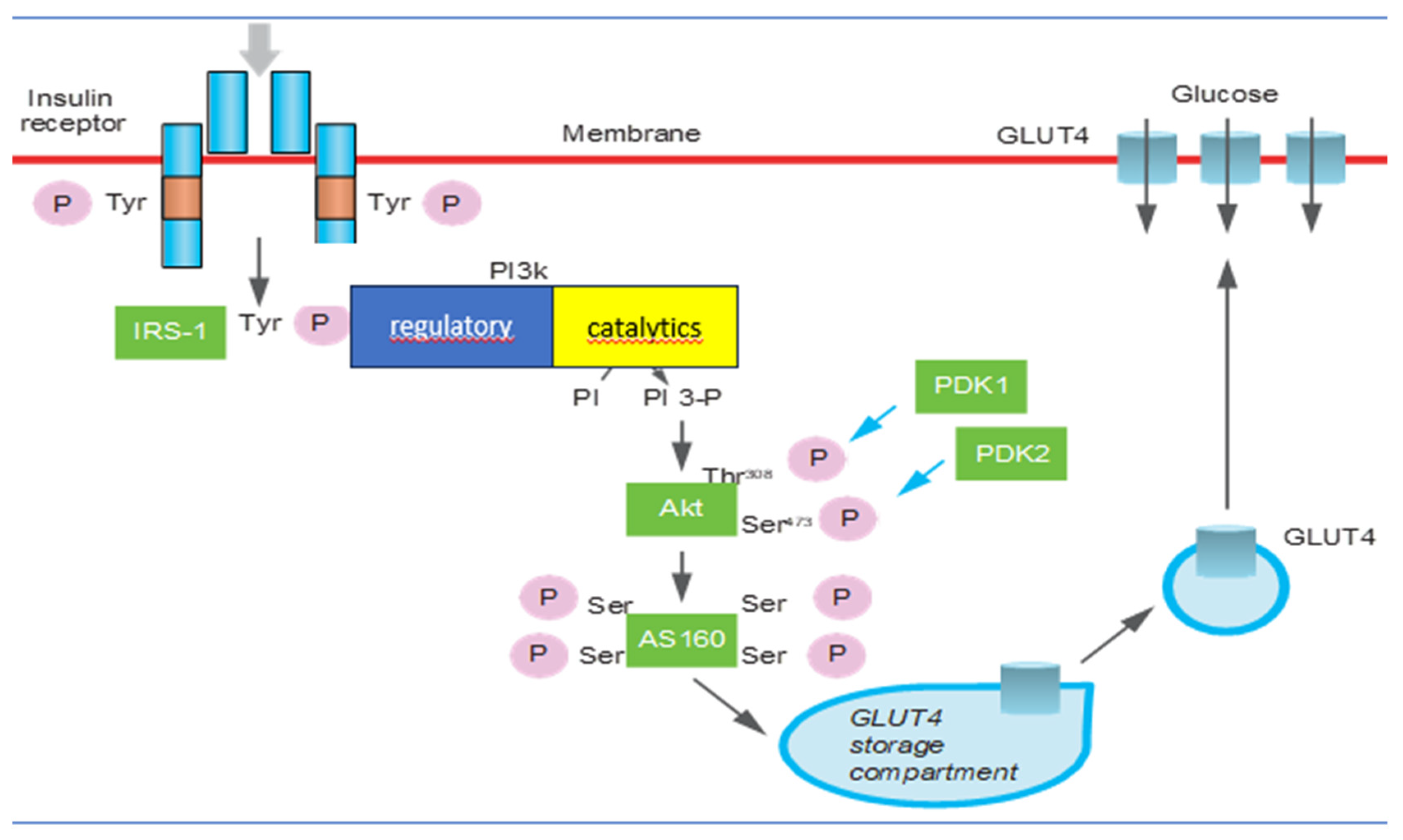
6. Insulin Signal Transduction Perturbations in Insulin Resistance
- Promotion of glucose entry into metabolism target cells by activating glucose transporter 4 (GLUT4) on the cell membrane in the liver, muscle, and adipose tissue, stimulating hepatic glycogen synthesis and inhibiting gluconeogenesis and glycogenolysis;
- Inhibition of lipolysis and stimulation of the synthesis of triglycerides, favoring their deposition in adipocytes (lipogenesis);
- Stimulation of protein synthesis by transcription and translation of mRNA in various cells;
- The phosphokinase pathway mediates the release of nitric oxide from endothelial cells (eNO), promoting vasodilation;
- Promotion of proliferation, contraction, and proinflammatory activity of smooth muscle cells.
7. Associated Health Conditions
7.1. Metabolic Syndrome
7.2. Atherogenesis and Endothelial Damage
7.3. Insulin Resistance and Cognitive and Neurological Diseases
7.4. Insulin Resistance and Cancer
7.5. Insulin Resistance and Hyperandrogenism
8. Lifestyle Factors
9. Conclusions
Take Home Messages
- IR is a condition in which target cells become less responsive to the hormone, leading to hyperglycemia. IR can affect both men and women, but there are some gender differences in its prevalence, risk factors, and associated health conditions.
- IR is generally more prevalent in men compared to premenopausal women. However, after menopause, the incidence of IR in women increases and becomes more comparable to that of men.
- Differences in body fat distribution between men and women contribute to gender variations in IR. Men tend to accumulate more visceral fat, which is fat stored around the abdominal organs, while women typically have a higher proportion of subcutaneous fat, which is located just beneath the skin. Visceral fat is strongly associated with IR and metabolic disturbances.
- Sex hormones play a role in insulin sensitivity. Estrogen, the primary female sex hormone, appears to have protective effects on insulin sensitivity. Women tend to have better insulin sensitivity during their reproductive years, partly due to the presence of estrogen. However, after menopause, when estrogen levels decline, women may experience a decrease in insulin sensitivity.
- IR is a key underlying factor in the development of T2DM. Men with insulin resistance are more likely to develop T2DM at an earlier age compared to women. Additionally, men with insulin resistance have a higher risk of developing nonalcoholic fatty liver disease (NAFLD) and cardiovascular disease compared to women with insulin resistance.
- Lifestyle choices, such as diet and physical activity, can influence IR. Men and women may have different patterns of dietary preferences and physical activity levels, which can contribute to variations in insulin sensitivity.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gluvic, Z.; Zaric, B.; Resanovic, I.; Obradovic, M.; Mitrovic, A.; Radak, D.; Isenovic, E.R. Link between Metabolic Syndrome and Insulin Resistance. Curr. Vasc. Pharmacol. 2017, 15, 30–39. [Google Scholar] [CrossRef]
- Tahapary, D.L.; Pratisthita, L.B.; Fitri, N.A.; Marcella, C.; Wafa, S.; Kurniawan, F.; Rizka, A.; Tarigan, T.J.E.; Harbuwono, D.S.; Purnamasari, D.; et al. Challenges in the diagnosis of insulin resistance: Focusing on the role of HOMA-IR and Tryglyceride/glucose index. Diabetes Metab. Syndr. 2022, 16, 102581. [Google Scholar] [CrossRef]
- Kaur, J. A comprehensive review on metabolic syndrome. Cardiol. Res. Pract. 2014, 2014, 943162. [Google Scholar] [CrossRef]
- Nolan, P.B.; Carrick-Ranson, G.; Stinear, J.W.; Reading, S.A.; Dalleck, L.C. Prevalence of metabolic syndrome and metabolic syndrome components in young adults: A pooled analysis. Prev. Med. Rep. 2017, 7, 211–215. [Google Scholar] [CrossRef]
- Aldhoon-Hainerová, I.; Zamrazilová, H.; Dušátková, L.; Sedláčková, B.; Hlavatý, P.; Hill, M.; Hampl, R.; Kunešová, M.; Hainer, V. Glucose homeostasis and insulin resistance: Prevalence, gender differences and predictors in adolescents. Diabetol. Metab. Syndr. 2014, 6, 100. [Google Scholar] [CrossRef]
- White, U.A.; Tchoukalova, Y.D. Sex dimorphism and depot differences in adipose tissue function. Biochim. Biophys. Acta 2014, 1842, 377–392. [Google Scholar] [CrossRef]
- Tramunt, B.; Smati, S.; Grandgeorge, N.; Lenfant, F.; Arnal, J.F.; Montagner, A.; Gourdy, P. Sex differences in metabolic regulation and diabetes susceptibility. Diabetologia 2020, 63, 453–461. [Google Scholar] [CrossRef]
- Chait, A.; Den Hartigh, L.J. Adipose tissue distribution, inflammation and its metabolic consequences, including diabetes and cardiovascular disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef]
- De Paoli, M.; Zakharia, A.; Werstuck, G.H. The Role of Estrogen in Insulin Resistance: A Review of Clinical and Preclinical Data. Am. J. Pathol. 2021, 191, 1490–1498. [Google Scholar] [CrossRef]
- Kautzky-Willer, A.; Harreiter, J.; Pacini, G. Sex and Gender Differences in Risk, Pathophysiology and Complications of Type 2 Diabetes Mellitus. Endocr. Rev. 2016, 37, 278–316. [Google Scholar] [CrossRef]
- Camilleri, G.; Kiani, A.K.; Herbst, K.L.; Kaftalli, J.; Bernini, A.; Dhuli, K.; Manara, E.; Bonetti, G.; Stuppia, L.; Paolacci, S.; et al. Genetics of fat deposition. Eur. Rev. Med. Pharmacol. Sci. 2021, 25 (Suppl. S1), 14–22. [Google Scholar] [CrossRef]
- Chen, X.; McClusky, R.; Itoh, Y.; Reue, K.; Arnold, A.P. X and Y chromosome complement influence adiposity and metabolism in mice. Endocrinology 2013, 154, 1092–1104. [Google Scholar] [CrossRef]
- Christakis, M.K.; Hasan, H.; De Souza, L.R.; Shirreff, L. The effect of menopause on metabolic syndrome: Cross-sectional results from the Canadian Longitudinal Study on Aging. Menopause 2020, 27, 999–1009. [Google Scholar] [CrossRef]
- Li, X.; Liu, J.; Zhou, B.; Li, Y.; Wu, Z.; Meng, H.; Wang, G. Sex Differences in the Effect of Testosterone on Adipose Tissue Insulin Resistance from Overweight to Obese Adults. J. Clin. Endocrinol. Metab. 2021, 106, 2252–2263. [Google Scholar] [CrossRef]
- Khalid, M.; Alkaabi, J.; Khan, M.A.B.; Adem, A. Insulin Signal Transduction Perturbations in Insulin Resistance. Int. J. Mol. Sci. 2021, 22, 8590. [Google Scholar] [CrossRef]
- Meex, R.C.R.; Blaak, E.E.; van Loon, L.J.C. Lipotoxicity plays a key role in the development of both insulin resistance and muscle atrophy in patients with type 2 diabetes. Obes. Rev. Off. J. Int. Assoc. Study Obes. 2019, 20, 1205–1217. [Google Scholar] [CrossRef]
- Tangvarasittichai, S. Oxidative stress, insulin resistance, dyslipidemia, and type 2 diabetes mellitus. World J. Diabetes 2015, 6, 456–480. [Google Scholar] [CrossRef]
- Boucher, J.; Kleinridders, A.; Kahn, C.R. Insulin receptor signaling in normal and insulin-resistant states. Cold Spring Harb. Perspect. Biol. 2014, 6, a009191. [Google Scholar] [CrossRef]
- Humphrey, S.J.; James, D.E.; Mann, M. Protein Phosphorylation: A Major Switch Mechanism for Metabolic Regulation. Trends Endocrinol. Metab. 2015, 26, 676–687. [Google Scholar] [CrossRef]
- Matheny, R.W., Jr.; Geddis, A.V.; Abdalla, M.N.; Leandry, L.A.; Ford, M.; McClung, H.L.; Pasiakos, S.M. AKT 2 Is the Predominant AKT Isoform Expressed in Human Skeletal Muscle. Physiol. Rep. 2018, 6, e13652. [Google Scholar] [CrossRef]
- Mezza, T.; Cinti, F.; Cefalo, C.M.A.; Pontecorvi, A.; Kulkarni, R.N.; Giaccari, A. β-Cell Fate in Human Insulin Resistance and Type 2 Diabetes: A Perspective on Islet Plasticity. Diabetes 2019, 68, 1121–1129. [Google Scholar] [CrossRef]
- Freeman, A.M.; Pennings, N. Insulin Resistance. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK507839 (accessed on 13 September 2023).
- Cerf, M.E. Beta Cell Dysfunction and Insulin Resistance. Front. Endocrinol. 2013, 4, 37. [Google Scholar] [CrossRef]
- Bergman, R.N.; Finegood, D.T.; Kahn, S.E. The Evolution of Β-cell Dysfunction and Insulin Resistance in Type 2 Diabetes. Eur. J. Clin. Investig. 2002, 32, 35–45. [Google Scholar] [CrossRef]
- Itoh, Y.; Kawamata, Y.; Harada, M.; Kobayashi, M.; Fujii, R.; Fukusumi, S.; Ogi, K.; Hosoya, M.; Tanaka, Y.; Uejima, H. Free Fatty Acids Regulate Insulin Secretion from Pancreatic β Cells through GPR40. Nature 2003, 422, 173–176. [Google Scholar] [CrossRef]
- Abedini, A.; Cao, P.; Plesner, A.; Zhang, J.; He, M.; Derk, J.; Patil, S.A.; Rosario, R.; Lonier, J.; Song, F. RAGE Binds Preamyloid IAPP Intermediates and Mediates Pancreatic β Cell Proteotoxicity. J. Clin. Investig. 2018, 128, 682–698. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms Linking Obesity to Insulin Resistance and Type 2 Diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef]
- Rabiee, A.; Krüger, M.; Ardenkjær-Larsen, J.; Kahn, C.R.; Emanuelli, B. Distinct Signalling Properties of Insulin Receptor Substrate (IRS)-1 and IRS-2 in Mediating Insulin/IGF-1 Action. Cell. Signal. 2018, 47, 1–15. [Google Scholar] [CrossRef]
- Sharma, M.; Aggarwal, S.; Nayar, U.; Vikram, N.K.; Misra, A.; Luthra, K. Differential Expression of Insulin Receptor Substrate-1 (IRS-1) in Visceral and Subcutaneous Adipose Depots of Morbidly Obese Subjects Undergoing Bariatric Surgery in a Tertiary Care Center in North India; SNP Analysis and Correlation with Metabolic Profile. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 981–986. [Google Scholar]
- Wisessaowapak, C.; Watcharasit, P.; Satayavivad, J. Arsenic Disrupts Neuronal Insulin Signaling through Increasing Free PI3K-P85 and Decreasing PI3K Activity. Toxicol. Lett. 2021, 349, 40–50. [Google Scholar] [CrossRef]
- Carnagarin, R.; Dharmarajan, A.M.; Dass, C.R. Molecular Aspects of Glucose Homeostasis in Skeletal Muscle–A Focus on the Molecular Mechanisms of Insulin Resistance. Mol. Cell. Endocrinol. 2015, 417, 52–62. [Google Scholar] [CrossRef]
- Goncalves, M.D.; Hopkins, B.D.; Cantley, L.C. Phosphatidylinositol 3-Kinase, Growth Disorders, and Cancer. N. Engl. J. Med. 2018, 379, 2052–2062. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Moten, A.; Lin, H.-K. Akt: A New Activation Mechanism. Cell Res. 2014, 24, 785. [Google Scholar] [CrossRef] [PubMed]
- Bizzarri, M.; Dinicola, S.; Cucina, A. Modulation of Both Insulin Resistance and Cancer Growth by Inositol. Curr. Pharm. Des. 2017, 23, 5200–5210. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Liu, G.; Guo, J.; Su, Z. The PI3K/AKT Pathway in Obesity and Type 2 Diabetes. Int. J. Biol. Sci. 2018, 14, 1483. [Google Scholar] [CrossRef]
- Saha, S. Association between the Membrane Transporter Proteins and Type 2 Diabetes Mellitus. Expert Rev. Clin. Pharmacol. 2020, 13, 287–297. [Google Scholar] [CrossRef]
- Klip, A.; McGraw, T.E.; James, D.E. Thirty Sweet Years of GLUT4. J. Biol. Chem. 2019, 294, 11369–11381. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. MTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Yoon, M.-S. MTOR as a Key Regulator in Maintaining Skeletal Muscle Mass. Front. Physiol. 2017, 8, 788. [Google Scholar] [CrossRef]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of Cellular Energy Sensing and Restoration of Metabolic Balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuñiga, F.A. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef]
- Nyman, K.; Granér, M.; Pentikäinen, M.O.; Lundbom, J.; Hakkarainen, A.; Sirén, R.; Nieminen, M.S.; Taskinen, M.-R.; Lundbom, N.; Lauerma, K. Cardiac steatosis and left ventricular function in men with metabolic syndrome. J. Cardiovasc. Magn. Reson. Off. J. Soc. Cardiovasc. Magn. Reson. 2013, 15, 103. [Google Scholar] [CrossRef] [PubMed]
- Kitade, H.; Chen, G.; Ni, Y.; Ota, T. Nonalcoholic Fatty Liver Disease and Insulin Resistance: New Insights and Potential New Treatments. Nutrients 2017, 9, 387. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Feng, J.; Zhang, G.; Yuan, X.; Li, F.; Yang, T.; Hao, S.; Huang, D.; Hsue, C.; Lou, Q. Visceral adipose tissue is more strongly associated with insulin resistance than subcutaneous adipose tissue in Chinese subjects with pre-diabetes. Curr. Med. Res. Opin. 2018, 34, 123–129. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, K.; Mercer, A.; Mawhinney, H.; Pulinilkunnil, T.; Udenigwe, C.C.; Kienesberger, P.C. Whey Peptides Stimulate Differentiation and Lipid Metabolism in Adipocytes and Ameliorate Lipotoxicity-Induced Insulin Resistance in Muscle Cells. Nutrients 2020, 12, 425. [Google Scholar] [CrossRef]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef]
- Xu, G.-X.; Wei, S.; Yu, C.; Zhao, S.-Q.; Yang, W.-J.; Feng, Y.-H.; Pan, C.; Yang, K.-X.; Ma, Y. Activation of Kupffer cells in NAFLD and NASH: Mechanisms and therapeutic interventions. Front. Cell Dev. Biol. 2023, 11, 1199519. [Google Scholar] [CrossRef]
- Mikolasevic, I.; Milic, S.; Turk Wensveen, T.; Grgic, I.; Jakopcic, I.; Stimac, D.; Wensveen, F.; Orlic, L. Nonalcoholic fatty liver disease - A multisystem disease? World J. Gastroenterol. 2016, 22, 9488–9505. [Google Scholar] [CrossRef]
- Villanueva-Ortega, E.; Garcés-Hernández, M.J.; Herrera-Rosas, A.; López-Alvarenga, J.C.; Laresgoiti-Servitje, E.; Escobedo, G.; Queipo, G.; Cuevas-Covarrubias, S.; Garibay-Nieto, G.N. Gender-specific differences in clinical and metabolic variables associated with NAFLD in a Mexican pediatric population. Ann. Hepatol. 2019, 18, 693–700. [Google Scholar] [CrossRef]
- Benedict, M.; Zhang, X. Non-alcoholic fatty liver disease: An expanded review. World J. Hepatol. 2017, 9, 715. [Google Scholar] [CrossRef]
- Ballestri, S.; Nascimbeni, F.; Baldelli, E.; Marrazzo, A.; Romagnoli, D.; Lonardo, A. NAFLD as a sexual dimorphic disease: Role of gender and reproductive status in the development and progression of nonalcoholic fatty liver disease and inherent cardiovascular risk. Adv. Ther. 2017, 34, 1291–1326. [Google Scholar] [CrossRef]
- Lonardo, A.; Trande, P. Are there any sex differences in fatty liver? A study of glucose metabolism and body fat distribution. J. Gastroenterol. Hepatol. 2000, 15, 775–782. [Google Scholar] [CrossRef] [PubMed]
- Lonardo, A.; Suzuki, A. Sexual Dimorphism of NAFLD in Adults. Focus on Clinical Aspects and Implications for Practice and Translational Research. J. Clin. Med. 2020, 9, 1278. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, A.; Ciobanu, O.S.; Ismaiel, M.; Leucuta, D.C.; Popa, S.L.; David, L.; Ensar, D.; Al Srouji, N.; Dumitrascu, D.L. Atherogenic Index of Plasma in Non-Alcoholic Fatty Liver Disease: Systematic Review and Meta-Analysis. Biomedicines 2022, 10, 2101. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-H.; Kim, S.H.; Kim, S.-N.; Kwon, H.-J.; Kim, J.-D.; Oh, J.Y.; Jung, Y.-S. Sex-specific metabolic interactions between liver and adipose tissue in MCD diet-induced non-alcoholic fatty liver disease. Oncotarget 2016, 7, 46959–46971. [Google Scholar] [CrossRef] [PubMed]
- Abu-Freha, N.; Cohen, B.; Weissmann, S.; Hizkiya, R.; Abu-Hammad, R.; Taha, G.; Gordon, M. Comorbidities and Outcomes among Females with Non-Alcoholic Fatty Liver Disease Compared to Males. Biomedicines 2022, 10, 2908. [Google Scholar] [CrossRef]
- Dolce, A.; Della Torre, S. Sex, Nutrition, and NAFLD: Relevance of Environmental Pollution. Nutrients 2023, 15, 2335. [Google Scholar] [CrossRef]
- Al-Nimer, M.S. Concomitant dysregulation of androgen secretion and dysfunction of adipose tissue induced insulin resistance. World J. Diabetes 2022, 13, 417–419. [Google Scholar] [CrossRef]
- Dumesic, D.A.; Akopians, A.L.; Madrigal, V.K.; Ramirez, E.; Margolis, D.J.; Sarma, M.K.; Thomas, A.M.; Grogan, T.R.; Haykal, R.; Schooler, T.A.; et al. Hyperandrogenism Accompanies Increased Intra-Abdominal Fat Storage in Normal Weight Polycystic Ovary Syndrome Women. J. Clin. Endocrinol. Metab. 2016, 101, 4178–4188. [Google Scholar] [CrossRef]
- Sanchez-Garrido, M.A.; Tena-Sempere, M. Metabolic dysfunction in polycystic ovary syndrome: Pathogenic role of androgen excess and potential therapeutic strategies. Mol. Metab. 2020, 35, 100937. [Google Scholar] [CrossRef]
- Pramfalk, C.; Pavlides, M.; Banerjee, R.; McNeil, C.A.; Neubauer, S.; Karpe, F.; Hodson, L. Sex-Specific Differences in Hepatic Fat Oxidation and Synthesis May Explain the Higher Propensity for NAFLD in Men. J. Clin. Endocrinol. Metab. 2015, 100, 4425–4433. [Google Scholar] [CrossRef]
- Nagral, A.; Bangar, M.; Menezes, S.; Bhatia, S.; Butt, N.; Ghosh, J.; Manchanayake, J.H.; Mahtab, M.A.; Singh, S.P. Gender Differences in Nonalcoholic Fatty Liver Disease. Euroasian J. Hepato-Gastroenterol. 2022; 12, (Suppl. S1), S19–S25. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Y.; Wang, L.; Li, Z.; Zhang, H.; Wu, J.; Rahman, N.; Guo, Y.; Li, D.; Li, N.; et al. Differential effects of estrogen/androgen on the prevention of nonalcoholic fatty liver disease in the male rat. J. Lipid Res. 2013, 54, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Song, M.J.; Choi, J.Y. Androgen dysfunction in non-alcoholic fatty liver disease: Role of sex hormone binding globulin. Front. Endocrinol. 2022, 13, 1053709. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, M.; Srinivasan, V.; Mohan, M.B.; Varghese, J.; Venkataraman, J. A study of gender-wise risk association between fatty liver and metabolic syndrome components (Asia-Pacific criteria) in a South Indian urban cohort. Indian J. Gastroenterol. 2015, 34, 38–42. [Google Scholar] [CrossRef]
- Wang, Q.; Zheng, D.; Liu, J.; Fang, L.; Li, Q. Atherogenic index of plasma is a novel predictor of non-alcoholic fatty liver disease in obese participants: A cross-sectional study. Lipids Health Dis. 2018, 17, 284. [Google Scholar] [CrossRef] [PubMed]
- Ciarambino, T.; Crispino, P.; Leto, G.; Mastrolorenzo, E.; Para, O.; Giordano, M. Influence of Gender in Diabetes Mellitus and Its Complication. Int. J. Mol. Sci. 2022, 23, 8850. [Google Scholar] [CrossRef]
- Unwin, N.; Shaw, J.; Zimmet, P.; Alberti, K.G. Impaired glucose tolerance and impaired fasting glycaemia: The current status definition and intervention. Diabet. Med. 2002, 19, 708–723. [Google Scholar]
- Ali, J.; Haider, S.M.S.; Ali, S.M.; Haider, T.; Anwar, A.; Hashmi, A.A. Overall Clinical Features of Type 2 Diabetes Mellitus with Respect to Gender. Cureus 2023, 15, e35771. [Google Scholar] [CrossRef]
- Maestro, B.; Campión, J.; Dávila, N.; Calle, C. Stimulation by 1,25-dihydroxyvitamin D3 of insulin receptor expression and insulin responsiveness for glucose transport in U-937 human promonocytic cells. Endocr. J. 2000, 47, 383–391. [Google Scholar] [CrossRef]
- Stadlmayr, A.; Aigner, E.; Huber-Schönauer, U.; Niederseer, D.; Zwerina, J.; Husar-Memmer, E.; Hohla, F.; Schett, G.; Patsch, W.; Datz, C. Relations of vitamin D status, gender and type 2 diabetes in middle-aged Caucasians. Acta Diabetol. 2015, 52, 39–46. [Google Scholar] [CrossRef]
- Ciarambino, T.; Crispino, P.; Minervini, G.; Giordano, M. Vitamin D: Can Gender Medicine Have a Role? Biomedicines 2023, 11, 1762. [Google Scholar] [CrossRef]
- Ciarambino, T.; Crispino, P.; Leto, G.; Minervini, G.; Para, O.; Giordano, M. Microbiota and Glucidic Metabolism: A Link with Multiple Aspects and Perspectives. Int. J. Mol. Sci. 2023, 24, 10409. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Manson, J.E.; Stevenson, J.C.; Fonseca, V.A. Menopausal Hormone Therapy and Type 2 Diabetes Prevention: Evidence, Mechanisms, and Clinical Implications. Endocr. Rev. 2017, 38, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.-L.; Lau, E.S.; Kong, A.P.; Fu, A.W.; Lau, V.; Jia, W.; Sheu, W.H.; Sobrepena, L.; Yoon, K.; Tan, A.T.; et al. Gender-associated cardiometabolic risk profiles and health behaviors in patients with type 2 diabetes: A cross-sectional analysis of the Joint Asia Diabetes Evaluation (JADE) program. The Lancet regional health. West. Pac. 2022, 32, 100663. [Google Scholar] [CrossRef]
- Taghibiglou, C.; Carpentier, A.; Van Iderstine, S.C.; Chen, B.; Rudy, D.; Aiton, A.; Lewis, G.F.; Adeli, K. Mechanisms of hepatic very low-density lipoprotein overproduction in insulin resistance. Evidence for enhanced lipoprotein assembly, reduced intracellular ApoB degradation, and increased microsomal triglyceride transfer protein in a fructose-fed hamster model. J. Biol. Chem. 2000, 275, 8416–8425. [Google Scholar] [CrossRef]
- Dilworth, L.; Facey, A.; Omoruyi, F. Diabetes Mellitus and Its Metabolic Complications: The Role of Adipose Tissues. Int. J. Mol. Sci. 2021, 22, 7644. [Google Scholar] [CrossRef] [PubMed]
- Başaran, Ö.; Doğan, V.; Mert, K.U.; Özlek, B.; Özlek, E.; Çelik, O.; Çil, C.; Özdemir, İ.H.; Rencüzoğulları, İ.; Özpamuk Karadeniz, F.; et al. How Did the Updated 2019 European Society of Cardiology/European Atherosclerosis Society Risk Categorization for Patients with Diabetes Affect the Risk Perception and Lipid Goals? A Simulated Analysis of Real-life Data from EPHESUS Study. Anatol. J. Cardiol. 2023, 27, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Villard, E.F.; Federspiel, M.C.; Cherfils, C.; Fesel-Fouquier, V.; Bruckert, E.; Clement, K.; Bonnefont-Rousselot, D.; Le Goff, W.; Bittar, R.; Couvert, P.; et al. Endogenous CETP activity as a predictor of cardiovascular risk: Determination of the optimal range. Atherosclerosis 2013, 227, 165–171. [Google Scholar] [CrossRef]
- Barzilai, N.; Atzmon, G.; Schechter, C.; Schaefer, E.J.; Cupples, A.L.; Lipton, R.; Cheng, S.; Shuldiner, A.R. Unique lipoprotein phenotype and genotype associated with exceptional longevity. JAMA 2003, 290, 2030–2040. [Google Scholar] [CrossRef]
- Cai, H.; Harrison, D.G. Endothelial dysfunction in cardiovascular diseases: The role of oxidant stress. Circ. Res. 2000, 87, 840–844. [Google Scholar] [CrossRef]
- Kajikawa, M.; Higashi, Y. Obesity and Endothelial Function. Biomedicines 2022, 10, 1745. [Google Scholar] [CrossRef]
- Kotlyarov, S. Immune Function of Endothelial Cells: Evolutionary Aspects, Molecular Biology and Role in Atherogenesis. Int. J. Mol. Sci. 2022, 23, 9770. [Google Scholar] [CrossRef] [PubMed]
- Rüster, C.; Wolf, G. The role of the renin-angiotensin-aldosterone system in obesity-related renal diseases. Semin. Nephrol. 2013, 33, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.S.; Schulman, I.H.; Zeng, Q. Link between the renin-angiotensin system and insulin resistance: Implications for cardiovascular disease. Vasc. Med. 2012, 17, 330–341. [Google Scholar] [CrossRef] [PubMed]
- Lastra-Lastra, G.; Sowers, J.R.; Restrepo-Erazo, K.; Manrique-Acevedo, C.; Lastra-González, G. Role of aldosterone and angiotensin II in insulin resistance: An update. Clin. Endocrinol. 2009, 71, 1–6. [Google Scholar] [CrossRef]
- Padilla, J.; Manrique-Acevedo, C.; Martinez-Lemus, L.A. New insights into mechanisms of endothelial insulin resistance in type 2 diabetes. Am. J. Physiol. Heart Circ. Physiol. 2022, 323, H1231–H1238. [Google Scholar] [CrossRef]
- Nabipoorashrafi, S.A.; Adeli, A.; Seyedi, S.A.; Rabizadeh, S.; Bahri, R.A.; Mohammadi, F.; Yadegar, A.; Nakhjavani, M.; Esteghamati, A. Comparison of insulin resistance indices in predicting albuminuria among patients with type 2 diabetes. Eur. J. Med. Res. 2023, 28, 166. [Google Scholar] [CrossRef]
- Nawaz, S.; Chinnadurai, R.; Al-Chalabi, S.; Evans, P.; Kalra, P.A.; Syed, A.A.; Sinha, S. Obesity and chronic kidney disease: A current review. Obes. Sci. Pract. 2022, 9, 61–74. [Google Scholar] [CrossRef]
- Boekholdt, S.; Sacks, F.; Jukema, J.; Shepherd, J.; Freeman, D.; McMahon, A.; Cambien, F.; Nicaud, V.; de Grooth, G.; Talmud, P.; et al. Cholesteryl ester transfer protein TaqIB variant, high-density lipoprotein cholesterol levels, cardiovascular risk, and efficacy of pravastatin treatment: Individual patient meta-analysis of 13,677 subjects. Circulation 2005, 111, 278–287. [Google Scholar] [CrossRef]
- Brousseau, M.E.; O’connor, J.J.; Ordovas, J.M.; Collins, D.; Otvos, J.D.; Massov, T.; McNamara, J.R.; Rubins, H.B.; Robins, S.J.; Schaefer, E.J. Cholesteryl ester transfer protein TaqI B2B2 genotype is associated with higher HDL cholesterol levels and lower risk of coronary heart disease end points in men with HDL deficiency: Veterans Affairs HDL Cholesterol Intervention Trial. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 1148–1154. [Google Scholar] [CrossRef]
- Ordovas, J.M.; Cupples, L.A.; Corella, D.; Otvos, J.D.; Osgood, D.; Martinez, A.; Lahoz, C.; Coltell, O.; Wilson, P.W.F.; Schaefer, E.J. Association of cholesteryl ester transfer protein-TaqIB polymorphism with variations in lipoprotein subclasses and coronary heart disease risk: The Framingham study. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 1323–1329. [Google Scholar] [CrossRef]
- Duan, Y.; Gong, K.; Xu, S.; Zhang, F.; Meng, X.; Han, J. Regulation of cholesterol homeostasis in health and diseases: From mechanisms to targeted therapeutics. Signal Transduct. Target. Ther. 2022, 7, 265. [Google Scholar] [CrossRef] [PubMed]
- Andrikopoulos, G.K.; Richter, D.J. Genetic factors in exceptional longevity. JAMA 2004, 291, 941–943. [Google Scholar] [CrossRef] [PubMed]
- Russo, G.T.; Horvath, K.V.; Di Benedetto, A.; Giandalia, A.; Cucinotta, D.; Asztalos, B. Influence of menopause and cholesteryl ester transfer protein (CETP) TaqIB polymorphism on lipid profile and HDL subpopulations distribution in women with and without type 2 diabetes. Atherosclerosis 2010, 210, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Ciarambino, T.; Crispino, P.; Giordano, M. Hyperuricemia and Endothelial Function: Is It a Simple Association or Do Gender Differences Play a Role in This Binomial? Biomedicines 2022, 10, 3067. [Google Scholar] [CrossRef]
- Ciarambino, T.; Crispino, P.; Giordano, M. Gender and Renal Insufficiency: Opportunities for Their Therapeutic Management? Cells 2022, 11, 3820. [Google Scholar] [CrossRef]
- Berlanga-Acosta, J.; Guillén-Nieto, G.; Rodríguez-Rodríguez, N.; Bringas-Vega, M.L.; García-Del-Barco-Herrera, D.; Berlanga-Saez, J.O.; García-Ojalvo, A.; Valdés-Sosa, M.J.; Valdés-Sosa, P.A. Insulin Resistance at the Crossroad of Alzheimer Disease Pathology: A Review. Front. Endocrinol. 2020, 11, 560375. [Google Scholar] [CrossRef]
- Kellar, D.; Craft, S. Brain insulin resistance in Alzheimer’s disease and related disorders: Mechanisms and therapeutic approaches. Lancet Neurol. 2020, 19, 758–766. [Google Scholar] [CrossRef]
- de la Monte, S.M. Contributions of brain insulin resistance and deficiency in amyloid-related neurodegeneration in Alzheimer’s disease. Drugs 2012, 72, 49–66. [Google Scholar] [CrossRef]
- Zhang, Y.; Deng, S.; Zhong, H.; Liu, M.; Ding, J.; Geng, R.; Tu, Q. Exploration and Clinical Verification of the Blood Co-Expression Genes of Type 2 Diabetes Mellitus and Mild Cognitive Dysfunction in the Elderly. Biomedicines 2023, 11, 993. [Google Scholar] [CrossRef]
- Paudel, Y.N.; Angelopoulou, E.; Piperi, C.; Shaikh, M.F.; Othman, I. Emerging neuroprotective effect of metformin in Parkinson’s disease: A molecular crosstalk. Pharmacol. Res. 2020, 152, 104593. [Google Scholar] [CrossRef]
- Haaxma, C.A.; Bloem, B.R.; Borm, G.F.; Oyen, W.J.G.; Leenders, K.L.; Eshuis, S.; Booij, J.; Dluzen, D.E.; Horstink, M.W.I.M. Gender differences in Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2007, 78, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Crispino, P.; Gino, M.; Barbagelata, E.; Ciarambino, T.; Politi, C.; Ambrosino, I.; Ragusa, R.; Marranzano, M.; Biondi, A.; Vacante, M. Gender Differences and Quality of Life in Parkinson’s Disease. Int. J. Environ. Res. Public Health 2020, 18, 198. [Google Scholar] [CrossRef]
- Schernhammer, E.; Hansen, J.; Rugbjerg, K.; Wermuth, L.; Ritz, B. Diabetes and the risk of developing Parkinson’s disease in Denmark. Diabetes Care 2011, 34, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Heller, J.; Dogan, I.; Schulz, J.B.; Reetz, K. Evidence for Gender Differences in Cognition, Emotion and Quality of Life in Parkinson’s Disease? Aging Dis. 2013, 5, 63–75. [Google Scholar] [PubMed]
- Amra, B.; Shafiei, A.; Soltaninejad, F.; Asgari, A.; Farajzadegan, Z. The Prevalence of Metabolic Syndrome in First Degree Relatives of Patients with Obstructive Sleep Apnea Syndrome: A Case-Control Study. Int. J. Prev. Med. 2022, 13, 76. [Google Scholar] [PubMed]
- Peled, N.; Kassirer, M.; Shitrit, D.; Kogan, Y.; Shlomi, D.; Berliner, A.S.; Kramer, M.R. The association of OSA with insulin resistance, inflammation and metabolic syndrome. Respir. Med. 2007, 101, 1696–1701. [Google Scholar] [CrossRef]
- Bajpai, J.; Pradhan, A.; Bajaj, D.; Verma, A.K.; Kant, S.; Pandey, A.K.; Sethi, R.; Dubey, A. Prevalence of dyslipidaemia in OSA patients at a tertiary care center. Am. J. Cardiovasc. Dis. 2023, 13, 1–9. [Google Scholar]
- Perrini, S.; Cignarelli, A.; Quaranta, V.N.; Falcone, V.A.; Kounaki, S.; Porro, S.; Ciavarella, A.; Ficarella, R.; Barbaro, M.; Genchi, V.A.; et al. Correction of intermittent hypoxia reduces inflammation in obese subjects with obstructive sleep apnea. JCI Insight 2017, 2, e94379. [Google Scholar] [CrossRef]
- Alterki, A.; Abu-Farha, M.; Al Shawaf, E.; Al-Mulla, F.; Abubaker, J. Investigating the Relationship between Obstructive Sleep Apnoea, Inflammation and Cardio-Metabolic Diseases. Int. J. Mol. Sci. 2023, 24, 6807. [Google Scholar] [CrossRef]
- Neumann, K.F.; Rojo, L.; Navarrete, L.P.; Farías, G.; Reyes, P.; Maccioni, R.B. Insulin resistance and Alzheimer’s disease: Molecular links & clinical implications. Curr. Alzheimer Res. 2008, 5, 438–447. [Google Scholar] [CrossRef]
- Beeri, M.S.; Bendlin, B.B. The link between type 2 diabetes and dementia: From biomarkers to treatment. Lancet Diabetes Endocrinol. 2020, 8, 736–738. [Google Scholar] [CrossRef] [PubMed]
- Bigal, M.E.; Kurth, T.; Santanello, N.; Buse, D.; Golden, W.; Robbins, M.; Lipton, R.B. Migraine and cardiovascular disease: A population-based study. Neurology 2010, 74, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Burch, R.C.; Rist, P.M.; Winter, A.C.; Buring, J.E.; Pradhan, A.D.; Loder, E.W.; Kurth, T. Migraine and risk of incident diabetes in women: A prospective study. Cephalalgia 2012, 32, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Rainero, I.; Limone, P.; Ferrero, M.; Valfrè, W.; Pelissetto, C.; Rubino, E.; Gentile, S.; Lo Giudice, R.; Pinessi, L. Insulin sensitivity is impaired in patients with migraine. Cephalalgia Int. J. Headache 2005, 25, 593–597. [Google Scholar] [CrossRef]
- Fagherazzi, G.; El Fatouhi, D.; Fournier, A.; Gusto, G.; Mancini, F.R.; Balkau, B.; Boutron-Ruault, M.-C.; Kurth, T.; Bonnet, F. Associations Between Migraine and Type 2 Diabetes in Women: Findings From the E3N Cohort Study. JAMA Neurol. 2019, 76, 257–263. [Google Scholar] [CrossRef]
- Varshney, P.; Parveen, R.; Khan, M.A.; Kohli, S.; Agarwal, N.B. Increased serum interleukin-9 and interleukin-1β are associated with depression in type 2 diabetes patients. Arq. Neuropsiquiatr. 2020, 78, 255–261. [Google Scholar] [CrossRef]
- Walker, C.S.; Li, X.; Whiting, L.; Glyn-Jones, S.; Zhang, S.; Hickey, A.J.; Sewell, M.A.; Ruggiero, K.; Phillips, A.R.J.; Kraegen, E.W.; et al. Mice lacking the neuropeptide alpha-calcitonin gene-related peptide are protected against diet-induced obesity. Endocrinology 2010, 151, 4257–4269. [Google Scholar] [CrossRef]
- Marics, B.; Peitl, B.; Varga, A.; Pázmándi, K.; Bácsi, A.; Németh, J.; Szilvássy, Z.; Jancsó, G.; Dux, M. Diet-induced obesity alters dural CGRP release and potentiates TRPA1-mediated trigeminovascular responses. Cephalalgia Int. J. Headache 2017, 37, 581–591. [Google Scholar] [CrossRef]
- Holland, P.R.; Akerman, S.; Goadsby, P.J. Orexin 1 receptor activation attenuates neurogenic dural vasodilation in an animal model of trigeminovascular nociception. J. Pharmacol. Exp. Ther. 2005, 315, 1380–1385. [Google Scholar] [CrossRef]
- Pisanu, C.; Preisig, M.; Castelao, E.; Glaus, J.; Cunningham, J.L.; Del Zompo, M.; Merikangas, K.R.; Schiöth, H.B.; Mwinyi, J. High leptin levels are associated with migraine with aura. Cephalalgia Int. J. Headache 2017, 37, 435–441. [Google Scholar] [CrossRef]
- Peterlin, B.L.; Sacco, S.; Bernecker, C.; Scher, A.I. Adipokines and Migraine: A Systematic Review. Headache 2016, 56, 622–644. [Google Scholar] [CrossRef] [PubMed]
- Kurth, T.; Gaziano, J.M.; Cook, N.R.; Logroscino, G.; Diener, H.C.; Buring, J.E. Migraine and risk of cardiovascular disease in women. JAMA. 2006, 296, 283–291; Correction in JAMA 2006, 296, 654. [Google Scholar] [CrossRef] [PubMed]
- Berge, L.I.; Riise, T.; Fasmer, O.B.; Hundal, Ø.; Oedegaard, K.J.; Midthjell, K.; Lund, A. Does diabetes have a protective effect on migraine? Epidemiology 2013, 24, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Haghighi, F.S.; Rahmanian, M.; Namiranian, N.; Arzaghi, S.M.; Dehghan, F.; Chavoshzade, F.; Sepehri, F. Migraine and type 2 diabetes; is there any association? J. Diabetes Metab. Disord. 2015, 15, 37. [Google Scholar] [CrossRef]
- Split, W.; Szydlowska, M. Headaches in non-insulin-dependent diabetes mellitus. Funct. Neurol. 1997, 12, 327–332. [Google Scholar]
- Rizvi, F.; Sheikh, A.; Ahmed, H.; Fahmi, S.; Ikramuddin, Z.; Asif, M. Miracle medicine for prevention of migraine attack: Metformin. Prof. Med. J. 2020, 27, 812–819. [Google Scholar] [CrossRef]
- Siva, Z.O.; Uluduz, D.; Keskin, F.E.; Erenler, F.; Balcı, H.; Uygunoğlu, U.; Saip, S.; Göksan, B.; Siva, A. Determinants of glucose metabolism and the role of NPY in the progression of insulin resistance in chronic migraine. Cephalalgia Int. J. Headache 2018, 38, 1773–1781. [Google Scholar] [CrossRef]
- Abancens, M.; Bustos, V.; Harvey, H.; McBryan, J.; Harvey, B.J. Sexual Dimorphism in Colon Cancer. Front. Oncol. 2020, 10, 607909. [Google Scholar] [CrossRef]
- Giorgino, F.; Belfiore, A.; Milazzo, G.; Costantino, A.; Maddux, B.; Whittaker, J.; Goldfine, I.D.; Vigneri, R. Overexpression of Insulin Receptors in Fibroblast and Ovary Cells Induces a Ligand-Mediated Transformed Phenotype. Mol. Endocrinol. 1991, 5, 452–459. [Google Scholar] [CrossRef]
- Lee, J.S.; Tocheny, C.E.; Shaw, L.M. The Insulin-like Growth Factor Signaling Pathway in Breast Cancer: An Elusive Therapeutic Target. Life 2022, 12, 1992. [Google Scholar] [CrossRef]
- Vigneri, R.; Goldfine, I.D.; Frittitta, L. Insulin, insulin receptors, and cancer. J. Endocrinol. Investig. 2016, 39, 1365–1376. [Google Scholar] [CrossRef]
- Arcidiacono, B.; Iiritano, S.; Nocera, A.; Possidente, K.; Nevolo, M.T.; Ventura, V.; Foti, D.; Chiefari, E.; Brunetti, A. Insulin Resistance and Cancer Risk: An Overview of the Pathogenetic Mechanisms. Exp. Diabetes Res. 2012, 2012, 789174. [Google Scholar] [CrossRef] [PubMed]
- Lønning, P.E.; Helle, S.-I.; Johannessen, D.C.; Adlercreutz, H.; Lien, E.A.; Tally, M.; Ekse, D.; Fotsis, T.; Anker, G.B.; Hall, K. Relations between sex hormones, sex hormone binding globulin, insulin-like growth factor-I and insulin-like growth factor binding protein-I in post-menopausal breast cancer patients. Clin. Endocrinol. 1995, 42, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Mahamat-Saleh, Y.; Rinaldi, S.; Kaaks, R.; Biessy, C.; Gonzalez-Gil, E.M.; Murphy, N.; Le Cornet, C.; Huerta, J.M.; Sieri, S.; Tjønneland, A.; et al. Metabolically defined body size and body shape phenotypes and risk of postmenopausal breast cancer in the European Prospective Investigation into Cancer and Nutrition. Cancer Med. 2023, 12, 12668–12682. [Google Scholar] [CrossRef] [PubMed]
- Dimou, N.; Kim, A.E.; Flanagan, O.; Murphy, N.; Diez-Obrero, V.; Shcherbina, A.; Aglago, E.K.; Bouras, E.; Campbell, P.T.; Casey, G.; et al. Probing the diabetes and colorectal cancer relationship using gene—Environment interaction analyses. Br. J. Cancer 2023, 129, 511–520. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, L.; Kempegowda, P.; Arlt, W.; O’Reilly, M.W. Mechanisms in endocrinology: The sexually dimorphic role of androgens in human metabolic disease. Eur. J. Endocrinol. 2017, 177, R125–R143. [Google Scholar]
- Ciarambino, T.; Crispino, P.; Para, O.; Giordano, M. Gender Medicine: A New Possible Frontiers of Venous Thromboembolism. Stresses 2023, 3, 167–181. [Google Scholar] [CrossRef]
- Stanhewicz, A.E.; Wenner, M.M.; Stachenfeld, N.S. Sex differences in endothelial function important to vascular health and overall cardiovascular disease risk across the lifespan. Am. J. Physiol.-Heart Circ. Physiol. 2018, 315, H1569–H1588. [Google Scholar] [CrossRef]
- Boese, A.C.; Kim, S.C.; Yin, K.-J.; Lee, J.-P.; Hamblin, M.H.; Yu, Y.; Wei, S.-G.; Weiss, R.M.; Felder, R.B.; Stanhewicz, A.E.; et al. Sex differences in vascular physiology and pathophysiology: Estrogen and androgen signaling in health and disease. Am. J. Physiol.-Heart Circ. Physiol. 2017, 313, H524–H545. [Google Scholar] [CrossRef]
- de Medeiros, S.F.; Rodgers, R.J.; Norman, R.J. Adipocyte and steroidogenic cell cross-talk in polycystic ovary syndrome. Hum. Reprod. Update 2021, 27, 771–796. [Google Scholar] [CrossRef]
- Rosenfield, R.L.; Ehrmann, D.A. The Pathogenesis of Polycystic Ovary Syndrome (PCOS): The Hypothesis of PCOS as Functional Ovarian Hyperandrogenism Revisited. Endocr. Rev. 2016, 37, 467–520. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Female Metabolic Characteristics | Male Metabolic Characteristics |
|---|---|
| Increase in total fat mass | Prevalent skeletal muscle mass |
| Prevalent subcutaneous adiposity | Prevalent visceral adiposity |
| Increase in insulin sensitivity with age | Increase in ectopic fat |
| Prevalent NEFA storage at rest | Prevalent NEFA oxidation at rest |
| Prevalent NEFA oxidation during exercise | Prevalent glucose oxidation during exercise |
| Prevalent glycemia alteration after a meal | Prevalent glycemia alteration at fast |
| Organ/Tissue | Estrogen Activity |
|---|---|
| Adipose tissue |
|
| Heart |
|
| Liver |
|
| Pancreas |
|
| Muscular system |
|
| Vascular endothelium |
|
| Associated Health Conditions of Insulin Resistance |
|---|
| Metabolic syndrome |
| Type-2 diabetes mellitus |
| Non-alcoholic fatty liver disease |
| Syndrome of polycystic ovary disease |
| Atherosclerosis and cardiovascular disease |
| Neurodegenerative disease(Parkinson’s disease and Alzheimer’s disease) |
| Neoplastic diseases(breast, colorectal, pancreas, and liver cancer) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciarambino, T.; Crispino, P.; Guarisco, G.; Giordano, M. Gender Differences in Insulin Resistance: New Knowledge and Perspectives. Curr. Issues Mol. Biol. 2023, 45, 7845-7861. https://doi.org/10.3390/cimb45100496
Ciarambino T, Crispino P, Guarisco G, Giordano M. Gender Differences in Insulin Resistance: New Knowledge and Perspectives. Current Issues in Molecular Biology. 2023; 45(10):7845-7861. https://doi.org/10.3390/cimb45100496
Chicago/Turabian StyleCiarambino, Tiziana, Pietro Crispino, Gloria Guarisco, and Mauro Giordano. 2023. "Gender Differences in Insulin Resistance: New Knowledge and Perspectives" Current Issues in Molecular Biology 45, no. 10: 7845-7861. https://doi.org/10.3390/cimb45100496
APA StyleCiarambino, T., Crispino, P., Guarisco, G., & Giordano, M. (2023). Gender Differences in Insulin Resistance: New Knowledge and Perspectives. Current Issues in Molecular Biology, 45(10), 7845-7861. https://doi.org/10.3390/cimb45100496