Inflammation-Related Molecules at the Maternal–Fetal Interface during Pregnancy and in Pathologically Altered Endometrium

 , , , ,
, , , ,
Abstract
1. Introduction
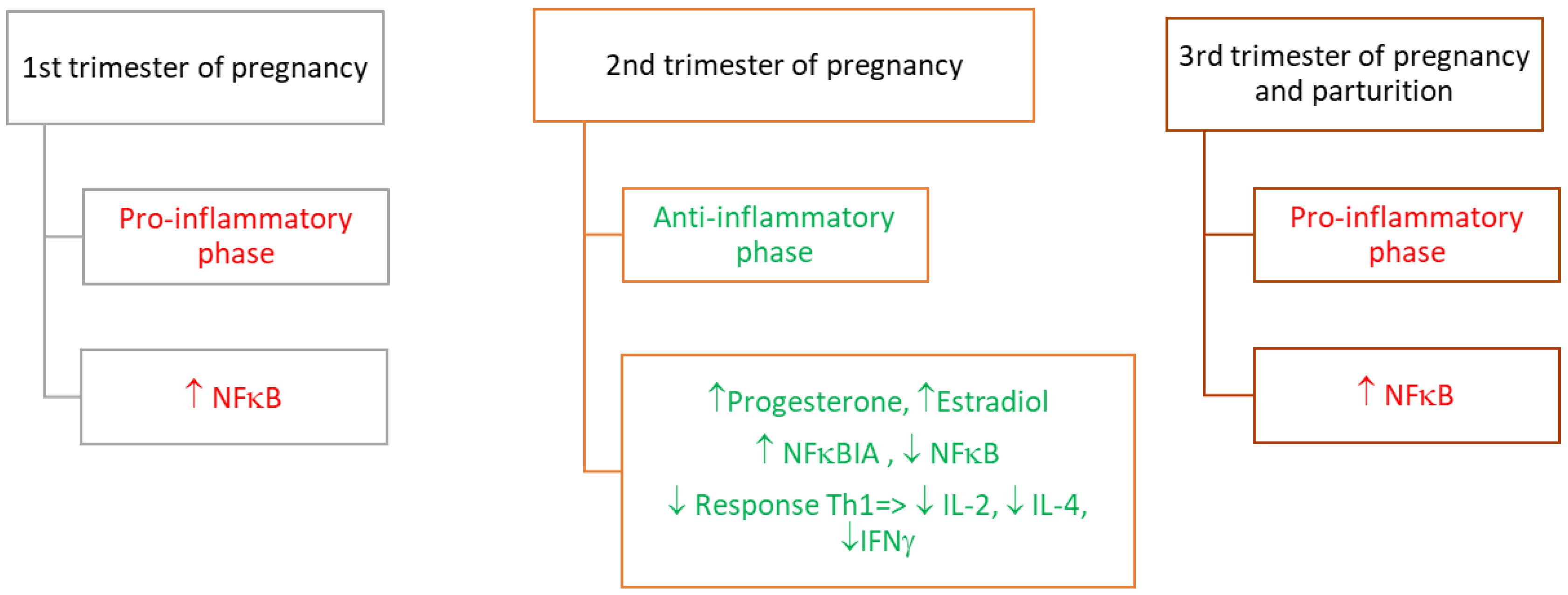
2. Inflammation-Related Molecules at the Maternal–Fetal Interface
2.1. Involvement of the Transcription Factor—NFκB in the Inflammatory Response
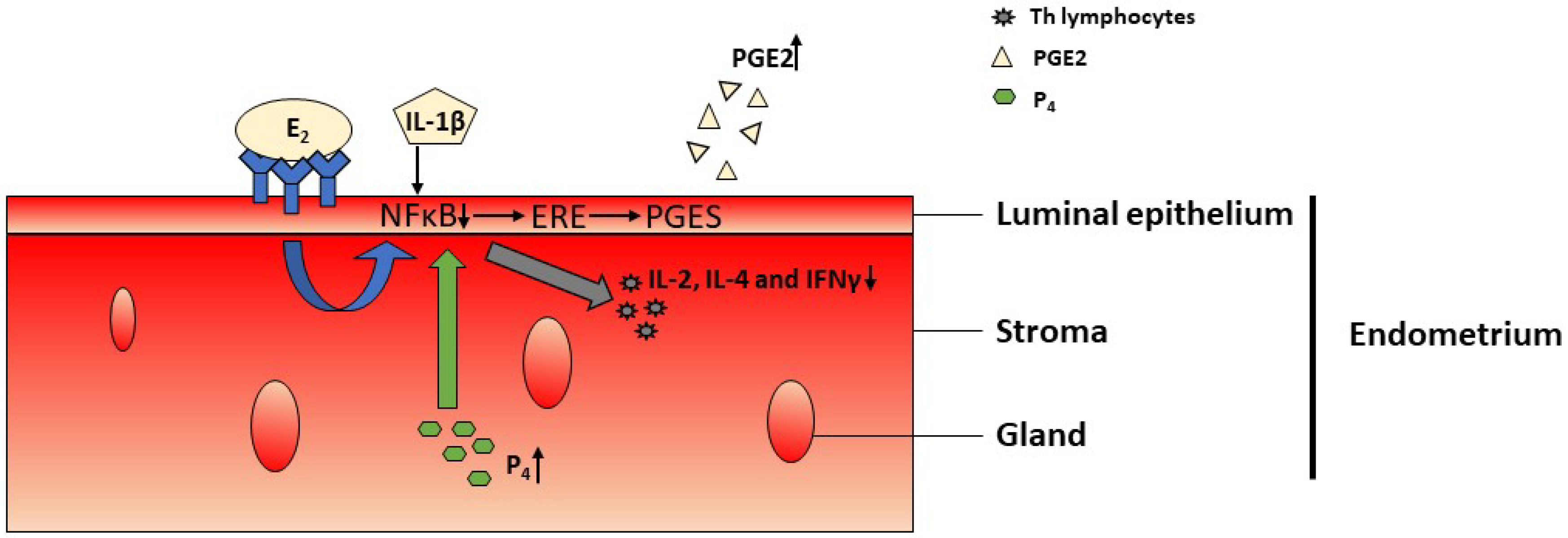
2.2. Involvement of PGs in the Inflammatory Response
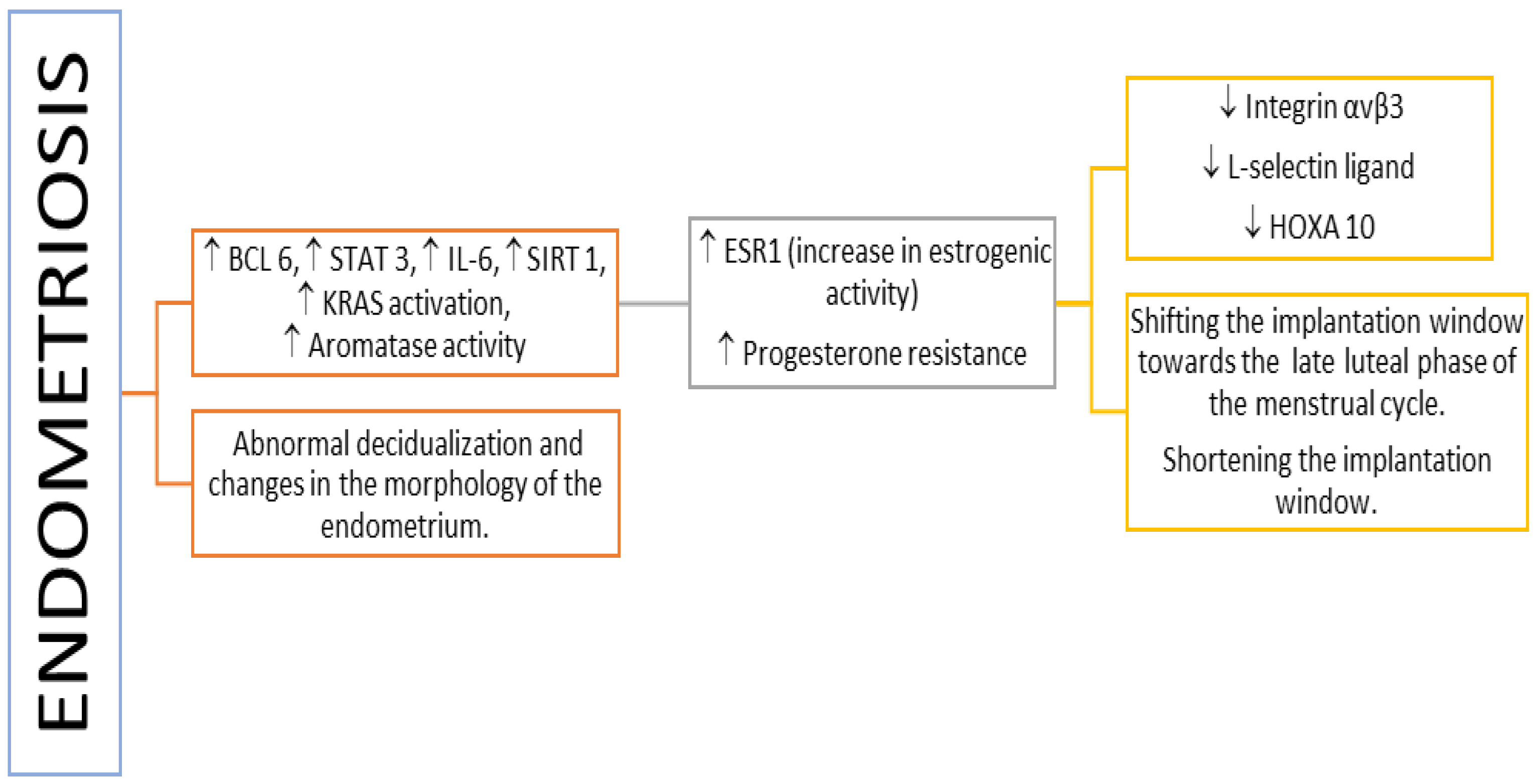
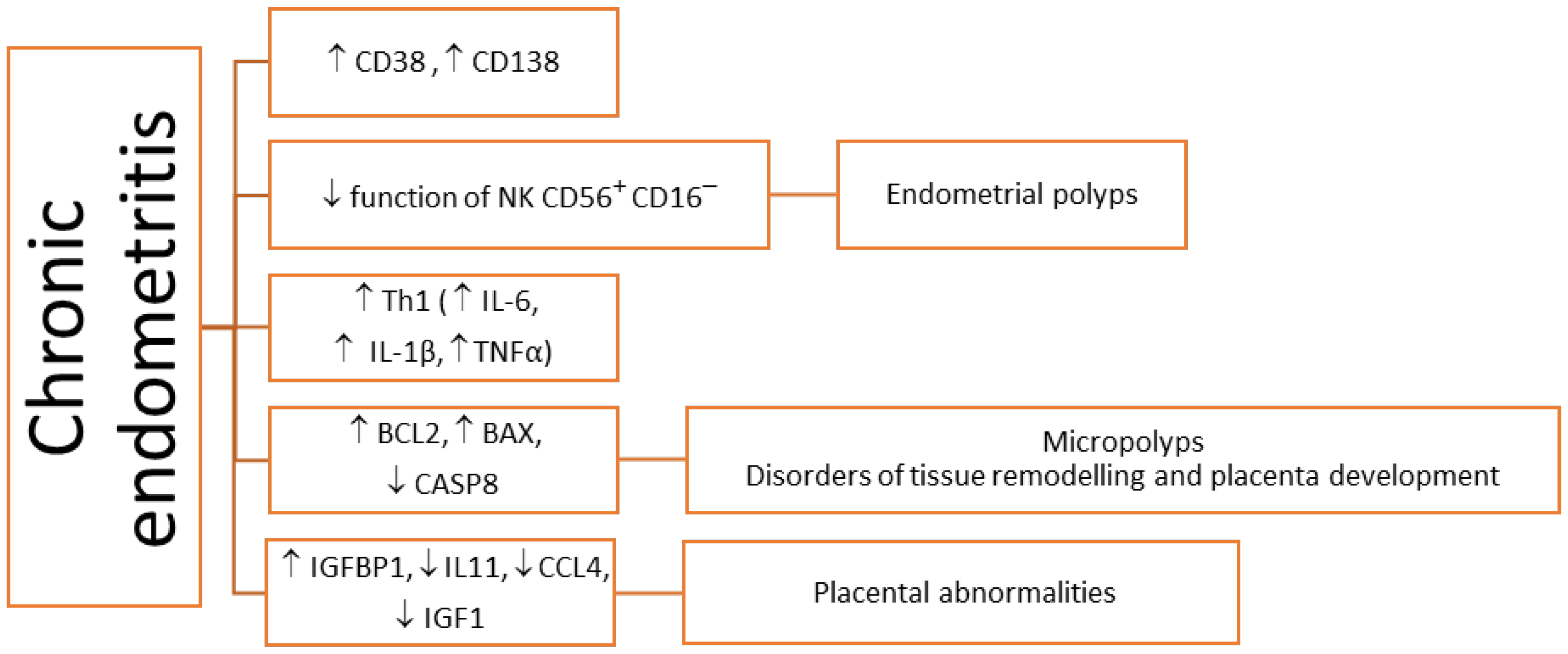
3. Inflammation-Related Molecules in Pathologically Altered Endometrium
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mor, G.; Cardenas, I.; Abrahams, V.; Guller, S. Inflammation and Pregnancy: The Role of the Immune System at the Implantation Site. Ann. N. Y. Acad. Sci. 2011, 1221, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Benner, M.; Ferwerda, G.; Joosten, I.; van der Molen, R.G. How Uterine Microbiota Might Be Responsible for a Receptive, Fertile Endometrium. Hum. Reprod. Update 2018, 24, 393–415. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Chávez, F.; Correa, D.; Navarrete-Meneses, P.; Cancino-Diaz, J.C.; Cancino-Diaz, M.E.; Rodríguez-Martínez, S. NF-κB and Its Regulators during Pregnancy. Front. Immunol. 2021, 12, 679106. [Google Scholar] [CrossRef] [PubMed]
- Pathare, A.; Hinduja, I.; Zaveri, K. Immunological Approach of Personalized Treatment for Recurrent Implantation Failure Patients Undergoing IVF. Glob. J. Reprod. Med. 2018, 5, 555667. [Google Scholar] [CrossRef]
- Salama, K.M.; Alloush, M.K. Are the Cytokines TNF Alpha and IL 1Beta Early Predictors of Embryo Implantation? Cross Sectional Study. J. Reprod. Immunol. 2020, 137, 102618. [Google Scholar] [CrossRef]
- Achache, H.; Revel, A. Endometrial Receptivity Markers, the Journey to Successful Embryo Implantation. Hum. Reprod. Update 2006, 12, 731–746. [Google Scholar] [CrossRef]
- Chimote, N.; Chimote, M.; Mehta, B.; Nath, N. Cytokines and Growth Factors in Implantation. J. Reprod. Stem Cell Biotechnol. 2010, 1, 219–243. [Google Scholar] [CrossRef]
- Massimiani, M.; Lacconi, V.; la Civita, F.; Ticconi, C.; Rago, R.; Campagnolo, L. Molecular Signaling Regulating Endometrium–Blastocyst Crosstalk. Int. J. Mol. Sci. 2020, 21, 23. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, T.; Guo, X.; Wong, C.K.; Chen, X.; Chan, Y.L.; Wang, C.C.; Laird, S.; Li, T.C. Successful Implantation Is Associated with a Transient Increase in Serum Pro-Inflammatory Cytokine Profile Followed by a Switch to Anti-Inflammatory Cytokine Profile Prior to Confirmation of Pregnancy. Fertil. Steril. 2021, 115, 1044–1053. [Google Scholar] [CrossRef]
- Maria, J.; Alecsandru, D. Circulating Cytokines during the Blastocyst Peri-Implantation Period. Fertil. Steril. 2021, 115, 905–906. [Google Scholar]
- Fülöp, V.; Vermes, G.; Demeter, J. The Relationship between Inflammatory and Immunological Processes during Pregnancy. Practical Aspects. Orv. Hetil. 2019, 160, 1247–1259. [Google Scholar] [CrossRef] [PubMed]
- Rehman, R.; Ashraf, M.; Jasmine, A.; Lal, K.; Alam, F. Cytokines and Endometrial Receptivity after Intracytoplasmic Sperm Injection—A Cohort Study at Islamabad. J. Pak. Med. Assoc. 2018, 68, 862–866. [Google Scholar] [PubMed]
- Dekel, N.; Gnainsky, Y.; Granot, I.; Mor, G. Inflammation and Implantation. Am. J. Reprod. Immunol. 2010, 63, 17–21. [Google Scholar] [CrossRef] [PubMed]
- Chavan, A.R.; Griffith, O.W.; Wagner, G.P. The Inflammation Paradox in the Evolution of Mammalian Pregnancy: Turning a Foe into a Friend. Curr. Opin. Genet. Dev. 2017, 47, 24–32. [Google Scholar] [CrossRef]
- Sharma, S.; Godbole, G.; Modi, D. Decidual Control of Trophoblast Invasion. Am. J. Reprod. Immunol. 2016, 75, 341–350. [Google Scholar] [CrossRef]
- Simón, C.; Mercader, A.; Gimeno, M.J.; Pellicer, A. The Interleukin-1 System and Human Implantation. Am. J. Reprod. Immunol. 1997, 37, 64–72. [Google Scholar] [CrossRef]
- Fazleabas, A.T.; Kim, J.J.; Strakova, Z. Implantation: Embryonic Signals and the Modulation of the Uterine Environment—A Review. Placenta 2004, 25, S26–S31. [Google Scholar] [CrossRef]
- Prutsch, N.; Fock, V.; Haslinger, P.; Haider, S.; Fiala, C.; Pollheimer, J.; Knöfler, M. The Role of Interleukin-1β in Human Trophoblast Motility. Placenta 2012, 33, 696–703. [Google Scholar] [CrossRef]
- Nakamura, H.; Jasper, M.J.; Hull, M.L.; Aplin, J.D.; Robertson, S.A. Macrophages Regulate Expression of A1, 2-Fucosyltransferase Genes in Human Endometrial Epithelial Cells. Mol. Hum. Reprod. 2012, 18, 204–215. [Google Scholar] [CrossRef]
- Robertson, S.A.; Moldenhauer, L.M. Immunological Determinants of Implantation Success. Int. J. Dev. Biol. 2014, 58, 205–217. [Google Scholar] [CrossRef]
- Minas, V.; Loutradis, D.; Makrigiannakis, A. Factors Controlling Blastocyst Implantation. Reprod. Biomed. Online 2005, 10, 205–216. [Google Scholar] [CrossRef]
- De Sousa, F.L.P.; Chaiwangyen, W.; Morales-Prieto, D.M.; Ospina-Prieto, S.; Weber, M.; Photini, S.M.; Sass, N.; Daher, S.; Schleussner, E.; Markert, U.R. Involvement of STAT1 in Proliferation and Invasiveness of Trophoblastic Cells. Reprod. Biol. 2017, 17, 218–224. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Yang, C.; Cheng, Y.; Wang, J.; Zhang, S.; Yan, S.; He, F.; Yin, T.; Yang, J. Trophoblast-Derived IL-6 Serves as an Important Factor for Normal Pregnancy by Activating Stat3-Mediated M2 Macrophages Polarization. Int. Immunopharmacol. 2021, 90, 106788. [Google Scholar] [CrossRef]
- Banerjee, P.; Malik, A.; Malhotra, S.S.; Gupta, S.K. Role of STAT Signaling and Autocrine Action of Chemokines during H2O2 Induced HTR-8/SVneo Trophoblastic Cells Invasion. J. Cell Physiol. 2019, 234, 1380–1397. [Google Scholar] [CrossRef] [PubMed]
- Ryu, B.J.; Han, J.W.; Kim, R.H.; Yun, S.; Kim, T.H.; Hur, S.E.; Kim, C.J.; Lee, S.K. Activation of NOD-1/JNK/IL-8 Signal Axis in Decidual Stromal Cells Facilitates Trophoblast Invasion. Am. J. Reprod. Immunol. 2017, 78, e12672. [Google Scholar] [CrossRef]
- Tanaka, K.; Watanabe, M.; Matsushima, M.; Matsuzawa, Y.; Izawa, T.; Nagashima, T.; Kobayashi, Y.; Iwashita, M. Synergistic Effects of Tumor Necrosis Factor-α and Insulin-like Growth Factor-I on Survival of Human Trophoblast-Derived BeWo Cell Line. Growth Horm. IGF Res. 2018, 41, 34–41. [Google Scholar] [CrossRef]
- Basu, J.; Agamasu, E.; Bendek, B.; Salafia, C.M.; Mishra, A.; Benfield, N.; Prasad, P.; Mikhail, M. Placental Tumor Necrosis Factor-α Protein Expression during Normal Human Gestation. J. Matern.-Fetal Neonatal Med. 2016, 29, 3934–3938. [Google Scholar] [CrossRef]
- Bauer, S.; Pollheimer, J.; Hartmann, J.; Husslein, P.; Aplin, J.D.; Knöfler, M. Tumor Necrosis Factor-α Inhibits Trophoblast Migration through Elevation of Plasminogen Activator Inhibitor-1 in First-Trimester Villous Explant Cultures. J. Clin. Endocrinol. Metab. 2004, 89, 812–822. [Google Scholar] [CrossRef]
- Otun, H.A.; Lash, G.E.; Innes, B.A.; Bulmer, J.N.; Naruse, K.; Hannon, T.; Searle, R.F.; Robson, S.C. Effect of Tumour Necrosis Factor-α in Combination with Interferon-γ on First Trimester Extravillous Trophoblast Invasion. J. Reprod. Immunol. 2011, 88, 1–11. [Google Scholar] [CrossRef]
- Lash, G.E.; Otun, H.A.; Innes, B.A.; Kirkley, M.; de Oliveira, L.; Searle, R.F.; Robson, S.C.; Bulmer, J.N. Interferon-γ Inhibits Extravillous Trophoblast Cell Invasion by a Mechanism That Involves Both Changes in Apoptosis and Protease Levels. FASEB J. 2006, 20, 2512–2518. [Google Scholar] [CrossRef]
- Verma, S.; Pal, R.; Gupta, S.K. Decrease in Invasion of HTR-8/SVneo Trophoblastic Cells by Interferon Gamma Involves Cross-Communication of STAT1 and BATF2 That Regulates the Expression of JUN. Cell Adhes. Migr. 2018, 12, 432–446. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Mann, D.A. Signal Transduction via the NF-κB Pathway: A Targeted Treatment Modality for Infection, Inflammation and Repair. Cell Biochem. Funct. Cell. Biochem. Its Modul. Act. Agents Dis. 2004, 22, 67–79. [Google Scholar] [CrossRef] [PubMed]
- Geisert, R.; Fazleabas, A.; Lucy, M.; Mathew, D. Interaction of the Conceptus and Endometrium to Establish Pregnancy in Mammals: Role of Interleukin 1β. Cell Tissue Res. 2012, 349, 825–838. [Google Scholar] [CrossRef]
- Novembri, R.; de Clemente, C.; Funghi, L.; Torricelli, M.; Voltolini, C.; Challis, J.R.; Petraglia, F. Corticotropin Releasing Hormone and Urocortin 2 Activate Inflammatory Pathways in Cultured Trophoblast Cell Lines. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 195, 200–205. [Google Scholar] [CrossRef] [PubMed]
- McCracken, S.A.; Hadfield, K.A.; Yenson, V.M.; Ariyakumar, K.J.; McKelvey, K.; Woodland, N.; ASHTON, A.W.; MORRIS, J.M. NF-KB Regulation in T-Cells in Pregnancy Is Mediated via Fas/FasL Interactions: The Signal for Which Is Derived from Exosomes Present in Maternal Plasma. Reprod. Immunol. 2016, 1, 8–18. [Google Scholar] [CrossRef][Green Version]
- Lindstrom, T.M.; Bennett, P.R. The Role of Nuclear Factor Kappa B in Human Labour. Reproduction 2005, 130, 569–581. [Google Scholar] [CrossRef]
- Biswas, D.K.; Singh, S.; Shi, Q.; Pardee, A.B.; Iglehart, J.D. Crossroads of Estrogen Receptor and NF-κB Signaling. Sci. STKE 2005, 2005, pe27. [Google Scholar] [CrossRef]
- Harnish, D.C. Estrogen Receptor Ligands in the Control of Pathogenic Inflammation. Curr. Opin. Investig. Drugs 2006, 7, 997–1001. [Google Scholar]
- Lou, Y.; Hu, M.; Wang, Q.; Yuan, M.; Wang, N.; Le, F.; Li, L.; Huang, S.; Wang, L.; Xu, X. Estradiol Suppresses TLR4-Triggered Apoptosis of Decidual Stromal Cells and Drives an Anti-Inflammatory TH2 Shift by Activating SGK1. Int. J. Biol. Sci. 2017, 13, 434. [Google Scholar] [CrossRef]
- King, A.E.; Critchley, H.O.D.; Kelly, R.W. The NF-ΚB Pathway in Human Endometrium and First Trimester Decidua. Mol. Hum. Reprod. 2001, 7, 175–183. [Google Scholar] [CrossRef]
- Ross, J.W.; Ashworth, M.D.; Mathew, D.; Reagan, P.; Ritchey, J.W.; Hayashi, K.; Spencer, T.E.; Lucy, M.; Geisert, R.D. Activation of the Transcription Factor, Nuclear Factor Kappa-B, during the Estrous Cycle and Early Pregnancy in the Pig. Reprod. Biol. Endocrinol. 2010, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Geisert, R.D.; Pratt, T.N.; Bazer, F.W.; Mayes, J.S.; Watson, G.H. Immunocytochemical Localization and Changes in Endometrial Progestin Receptor Protein during the Porcine Oestrous Cycle and Early Pregnancy. Reprod. Fertil. Dev. 1994, 6, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Cui, L.; Li, A.; Zhang, J.; Liu, Y.; Zhao, J.; Xu, X.; He, B.; Wang, J.; Chu, L. Endometrial Breakdown with Sustained Progesterone Release Involves NF-κB-mediated Functional Progesterone Withdrawal in a Mouse Implant Model. Mol. Reprod. Dev. 2016, 83, 780–791. [Google Scholar] [CrossRef]
- Kalkhoven, E.; Wissink, S.; van der Saag, P.T.; van der Burg, B. Negative Interaction between the RelA (P65) Subunit of NF-κB and the Progesterone Receptor. J. Biol. Chem. 1996, 271, 6217–6224. [Google Scholar] [CrossRef] [PubMed]
- Davies, S.; Dai, D.; Feldman, I.; Pickett, G.; Leslie, K.K. Identification of a Novel Mechanism of NF-ΚB Inactivation by Progesterone through Progesterone Receptors in Hec50co Poorly Differentiated Endometrial Cancer Cells: Induction of A20 and ABIN-2. Gynecol. Oncol. 2004, 94, 463–470. [Google Scholar] [CrossRef]
- Hardy, D.B.; Janowski, B.A.; Chen, C.-C.; Mendelson, C.R. Progesterone Receptor Inhibits Aromatase and Inflammatory Response Pathways in Breast Cancer Cells via Ligand-Dependent and Ligand-Independent Mechanisms. Mol. Endocrinol. 2008, 22, 1812–1824. [Google Scholar] [CrossRef] [PubMed]
- Bettelli, E.; Dastrange, M.; Oukka, M. Foxp3 Interacts with Nuclear Factor of Activated T Cells and NF-ΚB to Repress Cytokine Gene Expression and Effector Functions of T Helper Cells. Proc. Natl. Acad. Sci. USA 2005, 102, 5138–5143. [Google Scholar] [CrossRef]
- Kwiatek, M.; Gęca, T.; Kwaśniewska, A. Pro-and Anti-Inflammatory Cytokines in the First Trimester—Comparison of Missed Miscarriage and Normal Pregnancy. Int. J. Environ. Res. Public Health 2021, 18, 8538. [Google Scholar] [CrossRef]
- Wang, W.; Sung, N.; Gilman-Sachs, A.; Kwak-Kim, J. T Helper (Th) Cell Profiles in Pregnancy and Recurrent Pregnancy Losses: Th1/Th2/Th9/Th17/Th22/Tfh Cells. Front. Immunol. 2020, 11, 2025. [Google Scholar] [CrossRef]
- Lappas, M. NOD1 and NOD2 Regulate Proinflammatory and Prolabor Mediators in Human Fetal Membranes and Myometrium via Nuclear Factor-Kappa B. Biol. Reprod. 2013, 89, 11–14. [Google Scholar] [CrossRef]
- Ross, K.M.; Carroll, J.E.; Dunkel Schetter, C.; Hobel, C.; Cole, S.W. Pro-inflammatory Immune Cell Gene Expression during the Third Trimester of Pregnancy Is Associated with Shorter Gestational Length and Lower Birthweight. Am. J. Reprod. Immunol. 2019, 82, e13190. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, J.E.; Walsh, S.W. Activation of NF-ΚB in Placentas of Women with Preeclampsia. Hypertens Pregnancy 2012, 31, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Aban, M.; Cinel, L.; Arslan, M.; Dilek, U.; Kaplanoglu, M.; Arpaci, R.; Dilek, S. Expression of Nuclear Factor-Kappa B and Placental Apoptosis in Pregnancies Complicated with Intrauterine Growth Restriction and Preeclampsia: An Immunohistochemical Study. Tohoku J. Exp. Med. 2004, 204, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Vilella, F.; Ramirez, L.B.; Simón, C. Lipidomics as an Emerging Tool to Predict Endometrial Receptivity. Fertil. Steril. 2013, 99, 1100–1106. [Google Scholar] [CrossRef] [PubMed]
- Salleh, N. Diverse Roles of Prostaglandins in Blastocyst Implantation. Sci. World J. 2014, 2014, 968141. [Google Scholar] [CrossRef] [PubMed]
- Niringiyumukiza, J.D.; Cai, H.; Xiang, W. Prostaglandin E2 Involvement in Mammalian Female Fertility: Ovulation, Fertilization, Embryo Development and Early Implantation. Reprod. Biol. Endocrinol. 2018, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Waclawik, A.; Kaczynski, P.; Jabbour, H.N. Autocrine and Paracrine Mechanisms of Prostaglandin E2 Action on Trophoblast/Conceptus Cells through the Prostaglandin E2 Receptor (PTGER2) during Implantation. Endocrinology 2013, 154, 3864–3876. [Google Scholar] [CrossRef]
- Hallé, C.; Goff, A.K.; Petit, H.V.; Blouin, R.; Palin, M.-F. Effects of Different N-6: N-3 Fatty Acid Ratios and of Enterolactone on Gene Expression and PG Secretion in Bovine Endometrial Cells. Br. J. Nutr. 2015, 113, 56–71. [Google Scholar] [CrossRef][Green Version]
- Achache, H.; Tsafrir, A.; Prus, D.; Reich, R.; Revel, A. Defective Endometrial Prostaglandin Synthesis Identified in Patients with Repeated Implantation Failure Undergoing in Vitro Fertilization. Fertil. Steril. 2010, 94, 1271–1278. [Google Scholar] [CrossRef]
- Miravet-Valenciano, J.A.; Rincon-Bertolin, A.; Vilella, F.; Simon, C. Understanding and Improving Endometrial Receptivity. Curr. Opin. Obstet. Gynecol. 2015, 27, 187–192. [Google Scholar] [CrossRef]
- Huang, X.; Liu, H.; Li, R. Prostaglandin E2 Promotes BeWo Spheroids Implantation in RL95-2 Cell Monolayers. Gynecol. Endocrinol. 2017, 33, 548–552. [Google Scholar] [CrossRef] [PubMed]
- McCracken, J.A.; Custer, E.E.; Lamsa, J.C. Luteolysis: A Neuroendocrine-Mediated Event. Physiol. Rev. 1999, 79, 263–323. [Google Scholar] [CrossRef] [PubMed]
- Milne, S.A.; Jabbour, H.N. Prostaglandin (PG) F2α Receptor Expression and Signaling in Human Endometrium: Role of PGF2α in Epithelial Cell Proliferation. J. Clin. Endocrinol. Metab. 2003, 88, 1825–1832. [Google Scholar] [CrossRef]
- Xiao, L.; Zhang, Q.; Huang, X.; He, A.; Xie, S.; Li, Y. Endometrial Stromal Cell MiR-29c-3p Regulates Uterine Contraction. Reproduction 2019, 158, 493–501. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, J.; Yan, H.; Tian, D.; Li, H.; Zhang, Y.; Guo, T.; Wu, X.; Luo, W.; Zhou, Y. TP and/or EP3 Receptors Mediate the Vasoconstrictor and Pressor Responses of Prostaglandin F2α in Mice and/or Humans. FASEB J. 2019, 33, 2451–2459. [Google Scholar] [CrossRef] [PubMed]
- Broegger, T.; Andersson, K.-E.; Aalkjaer, C.; Forman, A.; Boedtkjer, D.B. Sensitivity to the Thromboxane A2 Analog U46619 Varies with Inner Diameter in Human Stem Villous Arteries. Placenta 2016, 39, 111–115. [Google Scholar] [CrossRef]
- Vetter, A.E.; O’Grady, S.M. Mechanisms of Electrolyte Transport across the Endometrium. I. Regulation by PGF2 Alpha and CAMP. Am. J. Physiol.-Cell Physiol. 1996, 270, C663–C672. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Long, A.; Fang, X.; Wood, S.L.; Slater, D.M.; Ni, X.; Olson, D.M. Effects of PGF2α on the Expression of Uterine Activation Proteins in Pregnant Human Myometrial Cells from Upper and Lower Segment. J. Clin. Endocrinol. Metab. 2013, 98, 2975–2983. [Google Scholar] [CrossRef]
- Xu, C.; You, X.; Liu, W.; Sun, Q.; Ding, X.; Huang, Y.; Ni, X. Prostaglandin F2 Regulates the Expression of Uterine Activation Proteins via Multiple Signalling Pathways. Reproduction 2015, 149, 139–146. [Google Scholar] [CrossRef]
- Kaczynski, P.; Kowalewski, M.P.; Waclawik, A. Prostaglandin F2α Promotes Angiogenesis and Embryo–Maternal Interactions during Implantation. Reproduction 2016, 151, 539–552. [Google Scholar] [CrossRef]
- Maybin, J.A.; Battersby, S.; Hirani, N.; Nikitenko, L.L.; Critchley, H.O.D.; Jabbour, H.N. The Expression and Regulation of Adrenomedullin in the Human Endometrium: A Candidate for Endometrial Repair. Endocrinology 2011, 152, 2845–2856. [Google Scholar] [CrossRef] [PubMed]
- Sztachelska, M.; Ponikwicka-Tyszko, D.; Sokolowska, G.; Anisimowicz, S.; Czerniecki, J.; Lebiedzinska, W.; Zbucka-Kretowska, M.; Zygmunt, M.; Wołczynski, S.; Pierzynski, P. Oxytocin Antagonism Reverses the Effects of High Oestrogen Levels and Oxytocin on Decidualization and Cyclooxygenase Activity in Endometrial Tissues. Reprod. Biomed. Online 2019, 39, 737–744. [Google Scholar] [CrossRef]
- Roth, K.; Zahradnik, H.P.; Schäfer, W.R. Effects of Different Progestins on Prostaglandin Biosynthesis in Human Endometrial Explants. Contraception 2019, 99, 61–66. [Google Scholar] [CrossRef]
- Kikut, J.; Komorniak, N.; Ziętek, M.; Palma, J.; Szczuko, M. Inflammation with the Participation of Arachidonic (AA) and Linoleic Acid (LA) Derivatives (HETEs and HODEs) Is Necessary in the Course of a Normal Reproductive Cycle and Pregnancy. J. Reprod. Immunol. 2020, 141, 103177. [Google Scholar] [CrossRef] [PubMed]
- Baryla, M.; Kaczynski, P.; Goryszewska, E.; Riley, S.C.; Waclawik, A. Prostaglandin F2α Stimulates Adhesion, Migration, Invasion and Proliferation of the Human Trophoblast Cell Line HTR-8/SVneo. Placenta 2019, 77, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.D.; Charlton, A.K.; Ayad, S. An Immunohistochemical Study of Human Endometrial Extracellular Matrix during the Menstrual Cycle and First Trimester of Pregnancy. Cell Tissue Res. 1988, 253, 231–240. [Google Scholar] [CrossRef]
- Béliard, A.; Donnez, J.; Nisolle, M.; Foidart, J.-M. Localization of Laminin, Fibronectin, E-Cadherin, and Integrins in Endometrium and Endometriosis. Fertil. Steril. 1997, 67, 266–272. [Google Scholar] [CrossRef]
- Irwin, J.C.; Kirk, D.; King, R.J.B.; Quigley, M.M.; Gwatkin, R.B.L. Hormonal Regulation of Human Endometrial Stromal Cells in Culture: An in Vitro Model for Decidualization. Fertil. Steril. 1989, 52, 761–768. [Google Scholar] [CrossRef]
- Tabibzadeh, S.; Kong, Q.F.; Babaknia, A.; May, L.T. Progressive Rise in the Expression of Interleukin-6 in Human Endometrium during Menstrual Cycle Is Initiated during the Implantation Window. MHR Basic Sci. Reprod. Med. 1995, 1, 407–413. [Google Scholar] [CrossRef]
- Downie, J.; Poyser, N.L.; Wunderlich, M. Levels of Prostaglandins in Human Endometrium during the Normal Menstrual Cycle. J. Physiol. 1974, 236, 465–472. [Google Scholar] [CrossRef]
- Woo, J.-H.; Yang, Y.-I.; Ahn, J.-H.; Choi, Y.S.; Choi, J.-H. Interleukin 6 Secretion from Alternatively Activated Macrophages Promotes the Migration of Endometriotic Epithelial Cells. Biol. Reprod. 2017, 97, 660–670. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jovanović, M.; Vićovac, L. Interleukin-6 Stimulates Cell Migration, Invasion and Integrin Expression in HTR-8/SVneo Cell Line. Placenta 2009, 30, 320–328. [Google Scholar] [CrossRef]
- Bazan, J.F. Structural Design and Molecular Evolution of a Cytokine Receptor Superfamily. Proc. Natl. Acad. Sci. USA 1990, 87, 6934–6938. [Google Scholar] [CrossRef]
- Norwitz, E.R.; Wilson, T. Secretory Component: A Potential Regulator of Endometrial-Decidual Prostaglandin Production in Early Human Pregnancy. Am. J. Obs. Gynecol. 2000, 183, 108–117. [Google Scholar]
- Bajekal, N.; Li, T.-C. Fibroids, Infertility and Pregnancy Wastage. Hum. Reprod. Update 2000, 6, 614–620. [Google Scholar] [CrossRef]
- Miura, S.; Khan, K.N.; Kitajima, M.; Hiraki, K.; Moriyama, S.; Masuzaki, H.; Samejima, T.; Fujishita, A.; Ishimaru, T. Differential Infiltration of Macrophages and Prostaglandin Production by Different Uterine Leiomyomas. Hum. Reprod. 2006, 21, 2545–2554. [Google Scholar] [CrossRef]
- Sugino, N.; Karube-Harada, A.; Kashida, S.; Takiguchi, S.; Kato, H. Reactive Oxygen Species Stimulate Prostaglandin F2α Production in Human Endometrial Stromal Cells in Vitro. Hum. Reprod. 2001, 16, 1797–1801. [Google Scholar] [CrossRef] [PubMed]
- Sugino, N.; Nakata, M.; Kashida, S.; Karube, A.; Takiguchi, S.; Kato, H. Decreased Superoxide Dismutase Expression and Increased Concentrations of Lipid Peroxide and Prostaglandin F2α in the Decidua of Failed Pregnancy. Mol. Hum. Reprod. 2000, 6, 642–647. [Google Scholar] [CrossRef]
- Keleş, I.D.; Ülgen, E.; Erkan, M.B.; Çelik, S.E.; Aydın, Y.; Önem, A.N.; Kandemir, H.; Arslanoğlu, T.; Apak, M.R.; Sezerman, U. Comparison of Endometrial Prostanoid Profiles in Three Infertile Subgroups: The Missing Part of Receptivity? Fertil. Steril. 2020, 113, 670–678. [Google Scholar] [CrossRef]
- Correa, F.; Wolfson, M.L.; Valchi, P.; Aisemberg, J.; Franchi, A.M. Endocannabinoid System and Pregnancy. Reproduction 2016, 152, R191–R200. [Google Scholar] [CrossRef]
- El-Talatini, M.R.; Taylor, A.H.; Konje, J.C. Fluctuation in Anandamide Levels from Ovulation to Early Pregnancy in In-Vitro Fertilization-Embryo Transfer Women, and Its Hormonal Regulation. Hum. Reprod. 2009, 24, 1989–1998. [Google Scholar] [CrossRef] [PubMed]
- Torella, M.; Bellini, G.; Punzo, F.; Argenziano, M.; Schiattarella, A.; Labriola, D.; Schettino, M.T.; Ambrosio, D.; Ammaturo, F.P.; de Franciscis, P. TNF-α Effect on Human Delivery Onset by CB1/TRPV1 Crosstalk: New Insights into Endocannabinoid Molecular Signaling in Preterm vs. Term Labor. Analysis of the EC/EV Pathway and Predictive Biomarkers for Early Diagnosis of Preterm Delivery. Minerva Ginecol. 2019, 71, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.D.; Sato, T.A.; Wang, A.; Keelan, J.A.; Ponnampalam, A.P.; Glass, M. Cannabinoids Stimulate Prostaglandin Production by Human Gestational Tissues through a Tissue-and CB1-Receptor-Specific Mechanism. Am. J. Physiol.-Endocrinol. Metab. 2008, 294, E352–E356. [Google Scholar] [CrossRef] [PubMed]
- Vercelli, C.A.; Aisemberg, J.; Cella, M.; Salazar, A.I.; Wolfson, M.L.; Franchi, A.M. Opposite Effects of Methanandamide on Lipopolysaccharide-Induced Prostaglandin E2 and F2α Synthesis in Uterine Explants from Pregnant Mice. PLoS ONE 2012, 7, e39532. [Google Scholar] [CrossRef]
- Dekel, N.; Gnainsky, Y.; Granot, I.; Racicot, K.; Mor, G. The Role of Inflammation for a Successful Implantation. Am. J. Reprod. Immunol. 2014, 72, 141–147. [Google Scholar] [CrossRef]
- Lessey, B.A.; Lebovic, D.I.; Taylor, R.N. Eutopic Endometrium in Women with Endometriosis: Ground Zero for the Study of Implantation Defects. In Proceedings of the Seminars in Reproductive Medicine; Thieme Medical Publishers: New York, NY, USA, 2013; Volume 31, pp. 109–124. [Google Scholar]
- Lessey, B.A.; Young, S.L. Homeostasis Imbalance in the Endometrium of Women with Implantation Defects: The Role of Estrogen and Progesterone. In Proceedings of the Seminars in Reproductive Medicine; Thieme Medical Publishers: New York, NY, USA, 2014; Volume 32, pp. 365–375. [Google Scholar]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and Treatment. Nat. Rev. Endocrinol. 2014, 10, 261. [Google Scholar] [CrossRef]
- Maksym, R.B.; Hoffmann-Młodzianowska, M.; Skibińska, M.; Rabijewski, M.; Mackiewicz, A.; Kieda, C. Immunology and Immunotherapy of Endometriosis. J. Clin. Med. 2021, 10, 5879. [Google Scholar] [CrossRef]
- vander Borght, M.; Wyns, C. Fertility and Infertility: Definition and Epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef]
- Meuleman, C.; Vandenabeele, B.; Fieuws, S.; Spiessens, C.; Timmerman, D.; D’Hooghe, T. High Prevalence of Endometriosis in Infertile Women with Normal Ovulation and Normospermic Partners. Fertil. Steril. 2009, 92, 68–74. [Google Scholar] [CrossRef]
- Ahn, S.H.; Khalaj, K.; Young, S.L.; Lessey, B.A.; Koti, M.; Tayade, C. Immune-Inflammation Gene Signatures in Endometriosis Patients. Fertil. Steril. 2016, 106, 1420–1431. [Google Scholar] [CrossRef]
- Ahn, S.H.; Edwards, A.K.; Singh, S.S.; Young, S.L.; Lessey, B.A.; Tayade, C. IL-17A Contributes to the Pathogenesis of Endometriosis by Triggering Proinflammatory Cytokines and Angiogenic Growth Factors. J. Immunol. 2015, 195, 2591–2600. [Google Scholar] [CrossRef] [PubMed]
- Khoufache, K.; Michaud, N.; Harir, N.; Kibangou, P.B.; Akoum, A. Anomalies in the Inflammatory Response in Endometriosis and Possible Consequences: A Review. Minerva Endocrinol. 2012, 37, 75–92. [Google Scholar] [PubMed]
- Lessey, B.A. Implantation Defects in Infertile Women with Endometriosis. Ann. N. Y. Acad. Sci. 2002, 955, 265–280. [Google Scholar] [CrossRef] [PubMed]
- Lessey, B.A.; Castelbaum, A.J. Integrins and Implantation in the Human. Rev. Endocr. Metab. Disord. 2002, 3, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Margarit, L.; Gonzalez, D.; Lewis, P.D.; Hopkins, L.; Davies, C.; Conlan, R.S.; Joels, L.; White, J.O. L-Selectin Ligands in Human Endometrium: Comparison of Fertile and Infertile Subjects. Hum. Reprod. 2009, 24, 2767–2777. [Google Scholar] [CrossRef] [PubMed]
- Foulk, R.A.; Zdravkovic, T.; Genbacev, O.; Prakobphol, A. Expression of L-Selectin Ligand MECA-79 as a Predictive Marker of Human Uterine Receptivity. J. Assist. Reprod. Genet. 2007, 24, 316–321. [Google Scholar] [CrossRef][Green Version]
- Kao, L.C.; Germeyer, A.; Tulac, S.; Lobo, S.; Yang, J.P.; Taylor, R.N.; Osteen, K.; Lessey, B.A.; Giudice, L.C. Expression Profiling of Endometrium from Women with Endometriosis Reveals Candidate Genes for Disease-Based Implantation Failure and Infertility. Endocrinology 2003, 144, 2870–2881. [Google Scholar] [CrossRef]
- Du, H.; Taylor, H.S. The Role of Hox Genes in Female Reproductive Tract Development, Adult Function, and Fertility. Cold Spring Harb. Perspect. Med. 2016, 6, a023002. [Google Scholar] [CrossRef]
- Celik, O.; Unlu, C.; Otlu, B.; Celik, N.; Caliskan, E. Laparoscopic Endometrioma Resection Increases Peri-Implantation Endometrial HOXA-10 and HOXA-11 MRNA Expression. Fertil. Steril. 2015, 104, 356–365. [Google Scholar] [CrossRef]
- Wu, Y.; Halverson, G.; Basir, Z.; Strawn, E.; Yan, P.; Guo, S.-W. Aberrant Methylation at HOXA10 May Be Responsible for Its Aberrant Expression in the Endometrium of Patients with Endometriosis. Am. J. Obs. Gynecol. 2005, 193, 371–380. [Google Scholar] [CrossRef]
- Klemmt, P.A.B.; Carver, J.G.; Kennedy, S.H.; Koninckx, P.R.; Mardon, H.J. Stromal Cells from Endometriotic Lesions and Endometrium from Women with Endometriosis Have Reduced Decidualization Capacity. Fertil. Steril. 2006, 85, 564–572. [Google Scholar] [CrossRef]
- Ahn, J.I.; Yoo, J.-Y.; Kim, T.H.; Kim, Y.I.; Ferguson, S.D.; Fazleabas, A.T.; Young, S.L.; Lessey, B.A.; Ahn, J.Y.; Lim, J.M. cAMP-Response Element-Binding 3-like Protein 1 (CREB3L1) Is Required for Decidualization and Its Expression Is Decreased in Women with Endometriosis. Curr. Mol. Med. 2016, 16, 276–287. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yin, X.; Pavone, M.E.; Lu, Z.; Wei, J.; Kim, J.J. Increased Activation of the PI3K/AKT Pathway Compromises Decidualization of Stromal Cells from Endometriosis. J. Clin. Endocrinol. Metab. 2012, 97, E35–E43. [Google Scholar] [CrossRef] [PubMed]
- Su, R.-W.; Strug, M.R.; Joshi, N.R.; Jeong, J.-W.; Miele, L.; Lessey, B.A.; Young, S.L.; Fazleabas, A.T. Decreased Notch Pathway Signaling in the Endometrium of Women with Endometriosis Impairs Decidualization. J. Clin. Endocrinol. Metab. 2015, 100, E433–E442. [Google Scholar] [CrossRef]
- Pei, T.; Liu, C.; Liu, T.; Xiao, L.; Luo, B.; Tan, J.; Li, X.; Zhou, G.; Duan, C.; Huang, W. MiR-194-3p Represses the Progesterone Receptor and Decidualization in Eutopic Endometrium from Women with Endometriosis. Endocrinology 2018, 159, 2554–2562. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Fu, J.; Xiao, L.; Yang, S.; Song, Y.; Zhang, X.; Feng, X.; Sun, H.; Xu, W.; Huang, W. MiR-196a Overexpression Activates the MEK/ERK Signal and Represses the Progesterone Receptor and Decidualization in Eutopic Endometrium from Women with Endometriosis. Hum. Reprod. 2016, 31, 2598–2608. [Google Scholar] [CrossRef]
- Eldafira, E.; Prasasty, V.D.; Abinawanto, A.; Syahfirdi, L.; Pujianto, D.A. Polymorphisms of Estrogen Receptor-α and Estrogen Receptor-β Genes and Its Expression in Endometriosis. Turk. J. Pharm. Sci. 2021, 18, 91. [Google Scholar] [CrossRef]
- Kobayashi, H.; Kimura, M.; Maruyama, S.; Nagayasu, M.; Imanaka, S. Revisiting Estrogen-Dependent Signaling Pathways in Endometriosis: Potential Targets for Non-Hormonal Therapeutics. Eur. J. Obstet. Gynecol. Reprod. Biol. 2021, 258, 103–110. [Google Scholar] [CrossRef]
- Yilmaz, B.D.; Bulun, S.E. Endometriosis and Nuclear Receptors. Hum. Reprod. Update 2019, 25, 473–485. [Google Scholar] [CrossRef]
- Patel, B.G.; Rudnicki, M.; Yu, J.; Shu, Y.; Taylor, R.N. Progesterone Resistance in Endometriosis: Origins, Consequences and Interventions. Acta Obs. Gynecol. Scand. 2017, 96, 623–632. [Google Scholar] [CrossRef]
- Marquardt, R.M.; Kim, T.H.; Shin, J.-H.; Jeong, J.-W. Progesterone and Estrogen Signaling in the Endometrium: What Goes Wrong in Endometriosis? Int. J. Mol. Sci. 2019, 20, 3822. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Martínez, E.R.; Bello-Alvarez, C.; Hermenegildo-Molina, A.L.; Solís-Paredes, M.; Parra-Hernández, S.; Cruz-Orozco, O.; Silvestri-Tomassoni, J.R.; Escobar-Ponce, L.F.; Hernández-López, L.A.; Reyes-Mayoral, C. Expression of Membrane Progesterone Receptors in Eutopic and Ectopic Endometrium of Women with Endometriosis. BioMed Res. Int. 2020, 2020, 2196024. [Google Scholar] [CrossRef] [PubMed]
- Rocha-Junior, C.V.; da Broi, M.G.; Miranda-Furtado, C.L.; Navarro, P.A.; Ferriani, R.A.; Meola, J. Progesterone Receptor B (PGR-B) Is Partially Methylated in Eutopic Endometrium from Infertile Women with Endometriosis. Reprod. Sci. 2019, 26, 1568–1574. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.-Y.; Kim, T.H.; Fazleabas, A.T.; Palomino, W.A.; Ahn, S.H.; Tayade, C.; Schammel, D.P.; Young, S.L.; Jeong, J.-W.; Lessey, B.A. KRAS Activation and Over-Expression of SIRT1/BCL6 Contributes to the Pathogenesis of Endometriosis and Progesterone Resistance. Sci. Rep. 2017, 7, 6765. [Google Scholar] [CrossRef]
- Li, M.; Peng, J.; Shi, Y.; Sun, P. MiR-92a Promotes Progesterone Resistance in Endometriosis through PTEN/AKT Pathway. Life Sci. 2020, 242, 117190. [Google Scholar] [CrossRef]
- Bulun, S.E.; Yildiz, S.; Adli, M.; Wei, J.-J. Adenomyosis Pathogenesis: Insights from next-Generation Sequencing. Hum. Reprod. Update 2021, 27, 1086–1097. [Google Scholar] [CrossRef]
- Fox, C.; Morin, S.; Jeong, J.-W.; Scott, R.T., Jr.; Lessey, B.A. Local and Systemic Factors and Implantation: What Is the Evidence? Fertil. Steril. 2016, 105, 873–884. [Google Scholar] [CrossRef]
- Likes, C.E.; Cooper, L.J.; Efird, J.; Forstein, D.A.; Miller, P.B.; Savaris, R.; Lessey, B.A. Medical or Surgical Treatment before Embryo Transfer Improves Outcomes in Women with Abnormal Endometrial BCL6 Expression. J. Assist. Reprod. Genet. 2019, 36, 483–490. [Google Scholar] [CrossRef]
- Patel, B.; Elguero, S.; Thakore, S.; Dahoud, W.; Bedaiwy, M.; Mesiano, S. Role of Nuclear Progesterone Receptor Isoforms in Uterine Pathophysiology. Hum. Reprod. Update 2015, 21, 155–173. [Google Scholar] [CrossRef]
- Kapoor, R.; Stratopoulou, C.A.; Dolmans, M.-M. Pathogenesis of Endometriosis: New Insights into Prospective Therapies. Int. J. Mol. Sci. 2021, 22, 11700. [Google Scholar] [CrossRef]
- Kalaitzopoulos, D.R.; Samartzis, N.; Kolovos, G.N.; Mareti, E.; Samartzis, E.P.; Eberhard, M.; Dinas, K.; Daniilidis, A. Treatment of Endometriosis: A Review with Comparison of 8 Guidelines. BMC Women’s Health 2021, 21, 397. [Google Scholar] [CrossRef]
- Koninckx, P.R.; Fernandes, R.; Ussia, A.; Schindler, L.; Wattiez, A.; Al-Suwaidi, S.; Amro, B.; Al-Maamari, B.; Hakim, Z.; Tahlak, M. Pathogenesis Based Diagnosis and Treatment of Endometriosis. Front. Endocrinol. 2021, 12, 745548. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.W.; Savaris, R.F.; Jeong, J.-W.; Kim, T.H.; Miller, P.B.; Likes, C.E.; Schammel, D.P.; Young, S.L.; Lessey, B.A. Unexplained Recurrent Pregnancy Loss and Unexplained Infertility: Twins in Disguise. Hum. Reprod. Open 2020, 2020, hoz021. [Google Scholar] [CrossRef]
- Kimura, F.; Takebayashi, A.; Ishida, M.; Nakamura, A.; Kitazawa, J.; Morimune, A.; Hirata, K.; Takahashi, A.; Tsuji, S.; Takashima, A. Chronic Endometritis and Its Effect on Reproduction. J. Obstet. Gynaecol. Res. 2019, 45, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Park, H.J.; Kim, Y.S.; Yoon, T.K.; Lee, W.S. Chronic Endometritis and Infertility. Clin. Exp. Reprod. Med. 2016, 43, 185–192. [Google Scholar] [CrossRef]
- Cicinelli, E.; de Ziegler, D.; Nicoletti, R.; Colafiglio, G.; Saliani, N.; Resta, L.; Rizzi, D.; de Vito, D. Chronic Endometritis: Correlation among Hysteroscopic, Histologic, and Bacteriologic Findings in a Prospective Trial with 2190 Consecutive Office Hysteroscopies. Fertil. Steril. 2008, 89, 677–684. [Google Scholar] [CrossRef]
- Haggerty, C.L.; Hillier, S.L.; Bass, D.C.; Ness, R.B.; PID Evaluation and Clinical Health (PEACH) Study Investigators. Bacterial Vaginosis and Anaerobic Bacteria Are Associated with Endometritis. Clin. Infect. Dis. 2004, 39, 990–995. [Google Scholar] [CrossRef]
- Polisseni, F.; Bambirra, E.A.; Camargos, A.F. Detection of Chronic Endometritis by Diagnostic Hysteroscopy in Asymptomatic Infertile Patients. Gynecol. Obs. Investig. 2003, 55, 205–210. [Google Scholar] [CrossRef]
- Kitaya, K.; Matsubayashi, H.; Takaya, Y.; Nishiyama, R.; Yamaguchi, K.; Takeuchi, T.; Ishikawa, T. Live Birth Rate Following Oral Antibiotic Treatment for Chronic Endometritis in Infertile Women with Repeated Implantation Failure. Am. J. Reprod. Immunol. 2017, 78, e12719. [Google Scholar] [CrossRef]
- Janošević, D.R.; Trandafilović, M.; Krtinić, D.; Čolović, H.; Stevanović, J.M.; Dinić, S.P.-T. Endometrial Immunocompetent Cells in Proliferative and Secretory Phase of Normal Menstrual Cycle. Folia Morphol. 2020, 79, 296–302. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, X.; Huang, J.; Wang, C.-C.; Yu, M.-Y.; Laird, S.; Li, T.-C. Comparison of the Prevalence of Chronic Endometritis as Determined by Means of Different Diagnostic Methods in Women with and without Reproductive Failure. Fertil. Steril. 2018, 109, 832–839. [Google Scholar] [CrossRef]
- Kitaya, K.; Yasuo, T. Immunohistochemistrical and Clinicopathological Characterization of Chronic Endometritis. Am. J. Reprod. Immunol. 2011, 66, 410–415. [Google Scholar] [CrossRef] [PubMed]
- Kasius, J.C.; Fatemi, H.M.; Bourgain, C.; Sie-Go, D.M.D.S.; Eijkemans, R.J.C.; Fauser, B.C.; Devroey, P.; Broekmans, F.J.M. The Impact of Chronic Endometritis on Reproductive Outcome. Fertil. Steril. 2011, 96, 1451–1456. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Li, T.-C.; Zhang, Y.; Feng, X.; Xia, E.; Huang, X.; Xiao, Y. Correlation between Hysteroscopy Findings and Chronic Endometritis. Fertil. Steril. 2019, 111, 772–779. [Google Scholar] [CrossRef] [PubMed]
- Kitaya, K.; Yasuo, T. Aberrant Expression of Selectin E, CXCL1, and CXCL13 in Chronic Endometritis. Mod. Pathol. 2010, 23, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yu, S.; Huang, C.; Lian, R.; Chen, C.; Liu, S.; Li, L.; Diao, L.; Markert, U.R.; Zeng, Y. Evaluation of Peripheral and Uterine Immune Status of Chronic Endometritis in Patients with Recurrent Reproductive Failure. Fertil. Steril. 2020, 113, 187–196. [Google Scholar] [CrossRef]
- Kosei, N.; Zakharenko, N.; Herman, D. Endometrial Polyps in Women of Reproductive Age: Clinical and Pathogene-Tic Variations. Georgian Med. News 2017, 273, 16–22. [Google Scholar]
- Tsonis, O.; Gkrozou, F.; Dimitriou, E.; Paschopoulos, M. Hysteroscopic Detection of Chronic Endometritis: Evaluating Proposed Hysteroscopic Features Suggestive of Chronic Endometritis. J. Gynecol. Obstet. Hum. Reprod. 2021, 50, 102182. [Google Scholar] [CrossRef]
- Kitazawa, J.; Kimura, F.; Nakamura, A.; Morimune, A.; Hanada, T.; Amano, T.; Tsuji, S.; Kasahara, K.; Satooka, H.; Hirata, T. Alteration in Endometrial Helper T-cell Subgroups in Chronic Endometritis. Am. J. Reprod. Immunol. 2021, 85, e13372. [Google Scholar] [CrossRef]
- Wang, W.-J.; Zhang, H.; Chen, Z.-Q.; Zhang, W.; Liu, X.-M.; Fang, J.-Y.; Liu, F.-J.; Kwak-Kim, J. Endometrial TGF-β, IL-10, IL-17 and Autophagy Are Dysregulated in Women with Recurrent Implantation Failure with Chronic Endometritis. Reprod. Biol. Endocrinol. 2019, 17, 2. [Google Scholar] [CrossRef]
- di Pietro, C.; Cicinelli, E.; Guglielmino, M.R.; Ragusa, M.; Farina, M.; Palumbo, M.A.; Cianci, A. Altered Transcriptional Regulation of Cytokines, Growth Factors, and Apoptotic Proteins in the Endometrium of Infertile Women with Chronic Endometritis. Am. J. Reprod. Immunol. 2013, 69, 509–517. [Google Scholar] [CrossRef] [PubMed]
- di Pietro, C.; Caruso, S.; Battaglia, R.; Iraci Sareri, M.; la Ferlita, A.; Strino, F.; Bonaventura, G.; di Mauro, M.; Barcellona, M.L.; Perciavalle, V. MiR-27a-3p and MiR-124-3p, Upregulated in Endometrium and Serum from Women Affected by Chronic Endometritis, Are New Potential Molecular Markers of Endometrial Receptivity. Am. J. Reprod. Immunol. 2018, 80, e12858. [Google Scholar] [CrossRef] [PubMed]
- Cicinelli, E.; Resta, L.; Nicoletti, R.; Tartagni, M.; Marinaccio, M.; Bulletti, C.; Colafiglio, G. Detection of Chronic Endometritis at Fluid Hysteroscopy. J. Minim. Invasive Gynecol. 2005, 12, 514–518. [Google Scholar] [CrossRef] [PubMed]
- George, A.F.; Jang, K.S.; Nyegaard, M.; Neidleman, J.; Spitzer, T.L.; Xie, G.; Chen, J.C.; Herzig, E.; Laustsen, A.; Marques de Menezes, E.G. Seminal Plasma Promotes Decidualization of Endometrial Stromal Fibroblasts in Vitro from Women with and without Inflammatory Disorders in a Manner Dependent on Interleukin-11 Signaling. Hum. Reprod. 2020, 35, 617–640. [Google Scholar] [CrossRef]
- Brünnert, D.; Shekhawat, I.; Chahar, K.R.; Ehrhardt, J.; Pandey, J.; Yadav, J.K.; Zygmunt, M.; Goyal, P. Thrombin Stimulates Gene Expression and Secretion of IL-11 via Protease-Activated Receptor-1 and Regulates Extravillous Trophoblast Cell Migration. J. Reprod. Immunol. 2019, 132, 35–41. [Google Scholar] [CrossRef]
- Dimitriadis, E.; White, C.A.; Jones, R.L.; Salamonsen, L.A. Cytokines, Chemokines and Growth Factors in Endometrium Related to Implantation. Hum. Reprod. Update 2005, 11, 613–630. [Google Scholar] [CrossRef]
- von Rango, U.; Alfer, J.; Kertschanska, S.; Kemp, B.; Müller-Newen, G.; Heinrich, P.C.; Beier, H.M.; Classen-Linke, I. Interleukin-11 Expression: Its Significance in Eutopic and Ectopic Human Implantation. Mol. Hum. Reprod. 2004, 10, 783–792. [Google Scholar] [CrossRef]
- Guzeloglu-Kayisli, O.; Kayisli, U.A.; Taylor, H.S. The Role of Growth Factors and Cytokines during Implantation: Endocrine and Paracrine Interactions. In Proceedings of the Seminars in Reproductive Medicine; © Thieme Medical Publishers: New York, NY, USA, 2009; Volume 27, pp. 62–79. [Google Scholar]
- Sansone, A.M.; Hisrich, B.V.; Young, R.B.; Abel, W.F.; Bowens, Z.; Blair, B.B.; Funkhouser, A.T.; Schammel, D.P.; Green, L.J.; Lessey, B.A. Evaluation of BCL6 and SIRT1 as Non-Invasive Diagnostic Markers of Endometriosis. Curr. Issues Mol. Biol. 2021, 43, 1350–1360. [Google Scholar] [CrossRef]
- Evans-Hoeker, E.; Lessey, B.A.; Jeong, J.W.; Savaris, R.F.; Palomino, W.A.; Yuan, L.; Schammel, D.P.; Young, S.L. Endometrial BCL6 Overexpression in Eutopic Endometrium of Women with Endometriosis. Reprod. Sci. 2016, 23, 1234–1241. [Google Scholar] [CrossRef]
- Li, X.; Large, M.J.; Creighton, C.J.; Lanz, R.B.; Jeong, J.-W.; Young, S.L.; Lessey, B.A.; Palomino, W.A.; Tsai, S.Y.; DeMayo, F.J. COUP-TFII Regulates Human Endometrial Stromal Genes Involved in Inflammation. Mol. Endocrinol. 2013, 27, 2041–2054. [Google Scholar] [CrossRef]
- Fu, J.-L.; Hsiao, K.-Y.; Lee, H.-C.; Li, W.-N.; Chang, N.; Wu, M.-H.; Tsai, S.-J. Suppression of COUP-TFII Upregulates Angiogenin and Promotes Angiogenesis in Endometriosis. Hum. Reprod. 2018, 33, 1517–1527. [Google Scholar] [CrossRef] [PubMed]
- Dimitriadis, E.; Robb, L.; Liu, Y.X.; Enders, A.C.; Martin, H.; Stoikos, C.; Wallace, E.; Salamonsen, L.A. IL-11 and IL-11Rα Immunolocalisation at Primate Implantation Sites Supports a Role for IL-11 in Placentation and Fetal Development. Reprod. Biol. Endocrinol. 2003, 1, 34. [Google Scholar] [CrossRef] [PubMed]
- Fest, S.; Aldo, P.B.; Abrahams, V.M.; Visintin, I.; Alvero, A.; Chen, R.; Chavez, S.L.; Romero, R.; Mor, G. Trophoblast–Macrophage Interactions: A Regulatory Network for the Protection of Pregnancy. Am. J. Reprod. Immunol. 2007, 57, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Anupa, G.; Bhat, M.A.; Srivastava, A.K.; Sharma, J.B.; Mehta, N.; Patil, A.; Sengupta, J.; Ghosh, D. Cationic Antimicrobial Peptide, Magainin down-Regulates Secretion of pro-Inflammatory Cytokines by Early Placental Cytotrophoblasts. Reprod. Biol. Endocrinol. 2015, 13, 121. [Google Scholar] [CrossRef]
- Gnainsky, Y.; Granot, I.; Aldo, P.; Barash, A.; Or, Y.; Mor, G.; Dekel, N. Biopsy-Induced Inflammatory Conditions Improve Endometrial Receptivity: The Mechanism of Action. Reproduction 2015, 149, 75–85. [Google Scholar] [CrossRef]
- de Ziegler, D.; Pirtea, P.; Galliano, D.; Cicinelli, E.; Meldrum, D. Optimal Uterine Anatomy and Physiology Necessary for Normal Implantation and Placentation. Fertil. Steril. 2016, 105, 844–854. [Google Scholar] [CrossRef]
- Kitaya, K.; Takeuchi, T.; Mizuta, S.; Matsubayashi, H.; Ishikawa, T. Endometritis: New Time, New Concepts. Fertil. Steril. 2018, 110, 344–350. [Google Scholar] [CrossRef]
- Ness, R.B.; Soper, D.E.; Holley, R.L.; Peipert, J.; Randall, H.; Sweet, R.L.; Sondheimer, S.J.; Hendrix, S.L.; Amortegui, A.; Trucco, G.; et al. Effectiveness of Inpatient and Outpatient Treatment Strategies for Women with Pelvic Inflammatory Disease: Results from the Pelvic Inflammatory Disease Evaluation and Clinical Health (Peach) Randomized Trial. Am. J. Obstet. Gynecol. 2002, 186, 929–937. [Google Scholar] [CrossRef]
- Song, D.; He, Y.; Wang, Y.; Liu, Z.; Xia, E.; Huang, X.; Xiao, Y.; Li, T.-C. Impact of Antibiotic Therapy on the Rate of Negative Test Results for Chronic Endometritis: A Prospective Randomized Control Trial. Fertil. Steril. 2021, 115, 1549–1556. [Google Scholar] [CrossRef]
- Hong, X.; Ding, W.; Yuan, R.; Ding, J.; Jin, J. Effect of Interventional Embolization Treatment for Hydrosalpinx on the Outcome of in Vitro Fertilization and Embryo Transfer. Medicine 2018, 97, e13143. [Google Scholar] [CrossRef]
- Palagiano, A.; Cozzolino, M.; Ubaldi, F.M.; Palagiano, C.; Coccia, M.E. Effects of Hydrosalpinx on Endometrial Implantation Failures: Evaluating Salpingectomy in Women Undergoing in Vitro Fertilization. Rev. Bras. Ginecol. Obs. 2021, 43, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Li, T.; Wang, Q.-L.; Zhou, H.-L.; Duan, L.; Cai, X. Effects of Hydrosalpinx on Ultrasonographic Parameters for Endometrial Receptivity during the Window of Implantation Measured by Power Color Doppler Ultrasound. Int. J. Clin. Exp. Med. 2015, 8, 6103. [Google Scholar] [PubMed]
- El-Mazny, A.; Ramadan, W.; Kamel, A.; Gad-Allah, S. Effect of Hydrosalpinx on Uterine and Ovarian Hemodynamics in Women with Tubal Factor Infertility. Eur. J. Obstet. Gynecol. Reprod. Biol. 2016, 199, 55–59. [Google Scholar] [CrossRef]
- Ng, E.H.Y.; Chan, C.C.W.; Tang, O.S.; Ho, P.C. Comparison of Endometrial and Subendometrial Blood Flows among Patients with and without Hydrosalpinx Shown on Scanning during in Vitro Fertilization Treatment. Fertil. Steril. 2006, 85, 333–338. [Google Scholar] [CrossRef]
- Bao, H.; Wang, G.; Huang, X.; Wang, M.; Wang, X.; Hao, C. The Impact of HSF on Endometrium. Rev. Assoc. Méd. Bras. 2017, 63, 1069–1075. [Google Scholar] [CrossRef]
- Ersahin, A.A.; Ersahin, S.; Gungor, N.D. Surgical Removal of Hydrosalpinx Improves Endometrium Receptivity by Decreasing Nuclear Factor-Kappa B Expression. Reprod. Sci. 2020, 27, 787–792. [Google Scholar] [CrossRef]
- Holzer, I.; Ott, J.; Kurz, C.; Hofstetter, G.; Hager, M.; Kuessel, L.; Parry, J.P. Is Chronic Endometritis Associated with Tubal Infertility? A Prospective Cohort Study. J. Minim. Invasive Gynecol. 2021, 28, 1876–1881. [Google Scholar] [CrossRef]
- Daftary, G.S.; Kayisli, U.; Seli, E.; Bukulmez, O.; Arici, A.; Taylor, H.S. Salpingectomy Increases Peri-Implantation Endometrial HOXA10 Expression in Women with Hydrosalpinx. Fertil. Steril. 2007, 87, 367–372. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pregnancy | Secreted Factor | Role | Reference |
|---|---|---|---|
| Preimplantation | TNFα | Induction of IL-1β secretion. | [6,7] |
| IL-1β | Promotion/propagation of decidualization and modulation of maternal NK cells, secretion of chemokines, and other factors required for implantation. Enhanced glycoprotein fucosylation. Regulation of the synthesis/secretion of trophoblastic matrix metalloproteinases MMP-2, MMP-3, and MMP-9 involved in trophoblast invasion. | [15,16,17,18,19,20,21] | |
| Implantation | IL-1β | Promotion/propagation of decidualization and modulation of maternal NK cells, secretion of chemokines, and other factors required for implantation. | [15,18,20] |
| IL-6 | Stimulation of migration and trophoblast invasion. | [22,23] | |
| IL-8 | Stimulation of migration and trophoblast invasion. | [24,25] | |
| TNFα | Protection of the maternal tissue against excessive trophoblast invasion through the mechanism based on trophoblastic cell apoptosis. Regulation of synthesis/secretion of trophoblastic matrix metalloproteinases MMP-2, MMP-3, and MMP-9 participating in trophoblast invasion. | [26,27,28,29] | |
| IFNγ | Protection of the maternal tissue against excessive trophoblast invasion through the mechanism based on trophoblastic cells apoptosis. | [9,29,30,31] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sieg, W.; Kiewisz, J.; Podolak, A.; Jakiel, G.; Woclawek-Potocka, I.; Lukaszuk, J.; Lukaszuk, K. Inflammation-Related Molecules at the Maternal–Fetal Interface during Pregnancy and in Pathologically Altered Endometrium. Curr. Issues Mol. Biol. 2022, 44, 3792-3808. https://doi.org/10.3390/cimb44090260
Sieg W, Kiewisz J, Podolak A, Jakiel G, Woclawek-Potocka I, Lukaszuk J, Lukaszuk K. Inflammation-Related Molecules at the Maternal–Fetal Interface during Pregnancy and in Pathologically Altered Endometrium. Current Issues in Molecular Biology. 2022; 44(9):3792-3808. https://doi.org/10.3390/cimb44090260
Chicago/Turabian StyleSieg, Wlodzimierz, Jolanta Kiewisz, Amira Podolak, Grzegorz Jakiel, Izabela Woclawek-Potocka, Jakub Lukaszuk, and Krzysztof Lukaszuk. 2022. "Inflammation-Related Molecules at the Maternal–Fetal Interface during Pregnancy and in Pathologically Altered Endometrium" Current Issues in Molecular Biology 44, no. 9: 3792-3808. https://doi.org/10.3390/cimb44090260
APA StyleSieg, W., Kiewisz, J., Podolak, A., Jakiel, G., Woclawek-Potocka, I., Lukaszuk, J., & Lukaszuk, K. (2022). Inflammation-Related Molecules at the Maternal–Fetal Interface during Pregnancy and in Pathologically Altered Endometrium. Current Issues in Molecular Biology, 44(9), 3792-3808. https://doi.org/10.3390/cimb44090260