
Increased Production of Interleukin-10 and Tumor Necrosis Factor-Alpha in Stimulated Peripheral Blood Mononuclear Cells after Inhibition of S100A12

 , , , ,
, , , ,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Definitions
2.2. Plasma and Peripheral Blood Mononuclear Cell (PBMC) Preparation
2.3. Measurement of Plasma Cytokine Levels
2.4. Cell Culture
2.5. Measurement of Supernatant Cytokine Levels
2.6. Statistical Analysis
3. Results
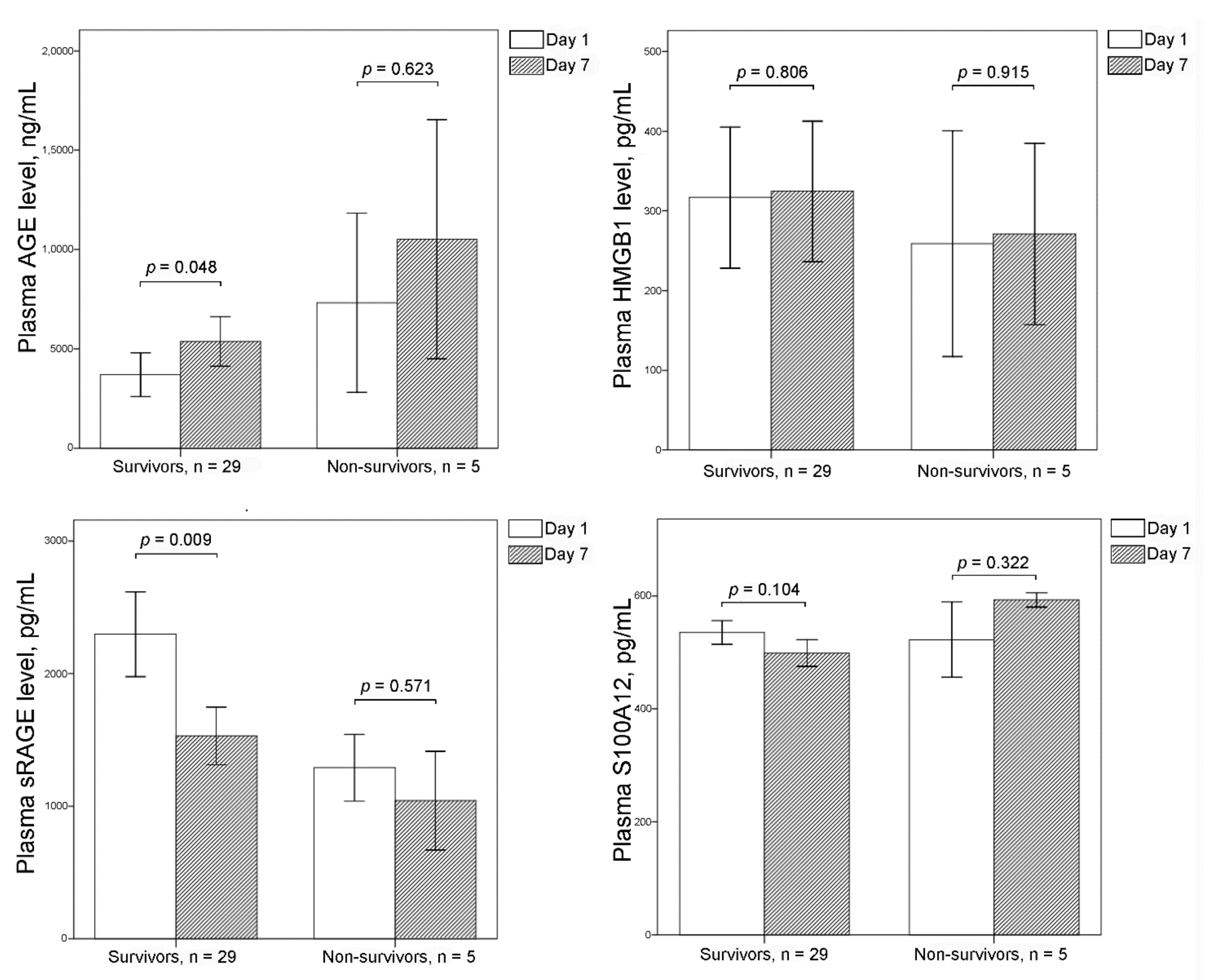
3.1. Plasma Levels of AGE, sRAGE, HMGB1, and S100A12 in Survivors, Non-Survivors, and Controls
3.2. Plasma Levels of AGE, sRAGE, HMGB1, and S100A12 between Days 1 and 7
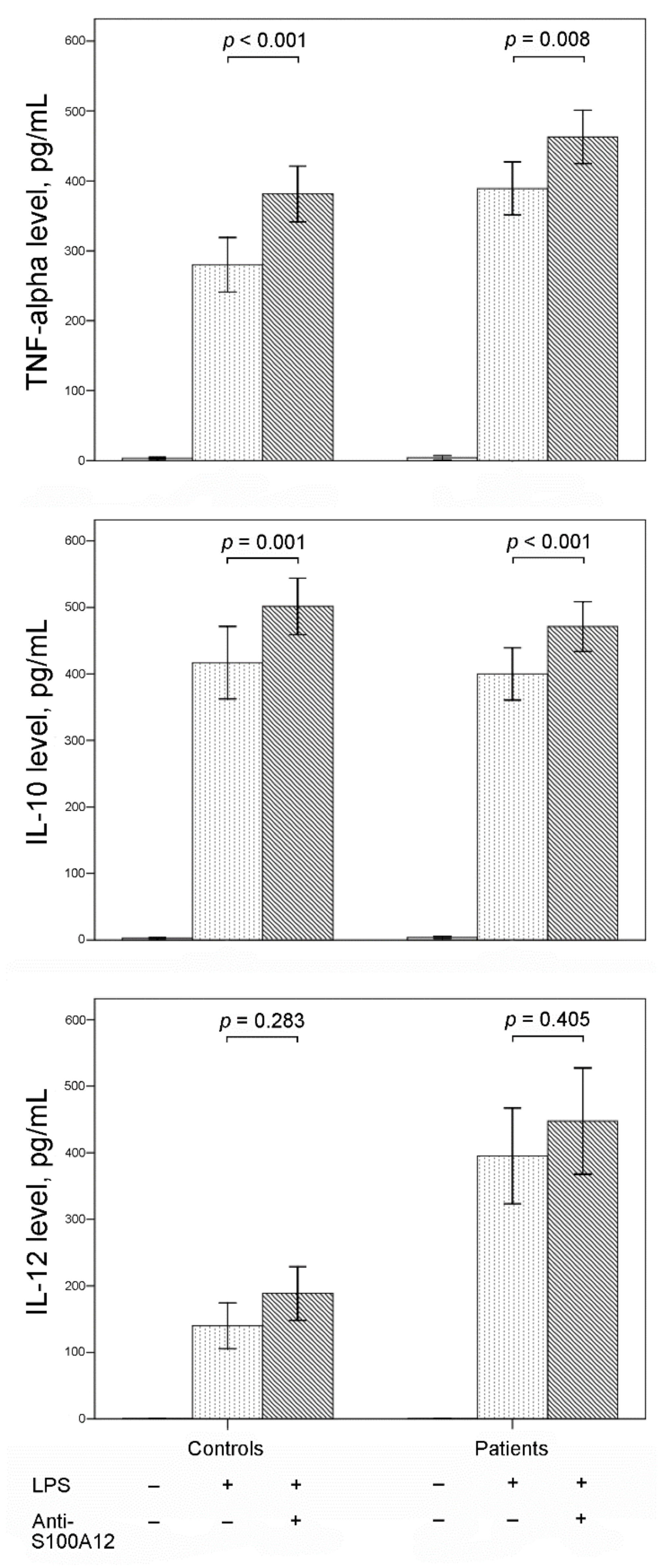
3.3. Effect of Inhibiting S100A12 on Cytokine Production in Stimulated PBMCs
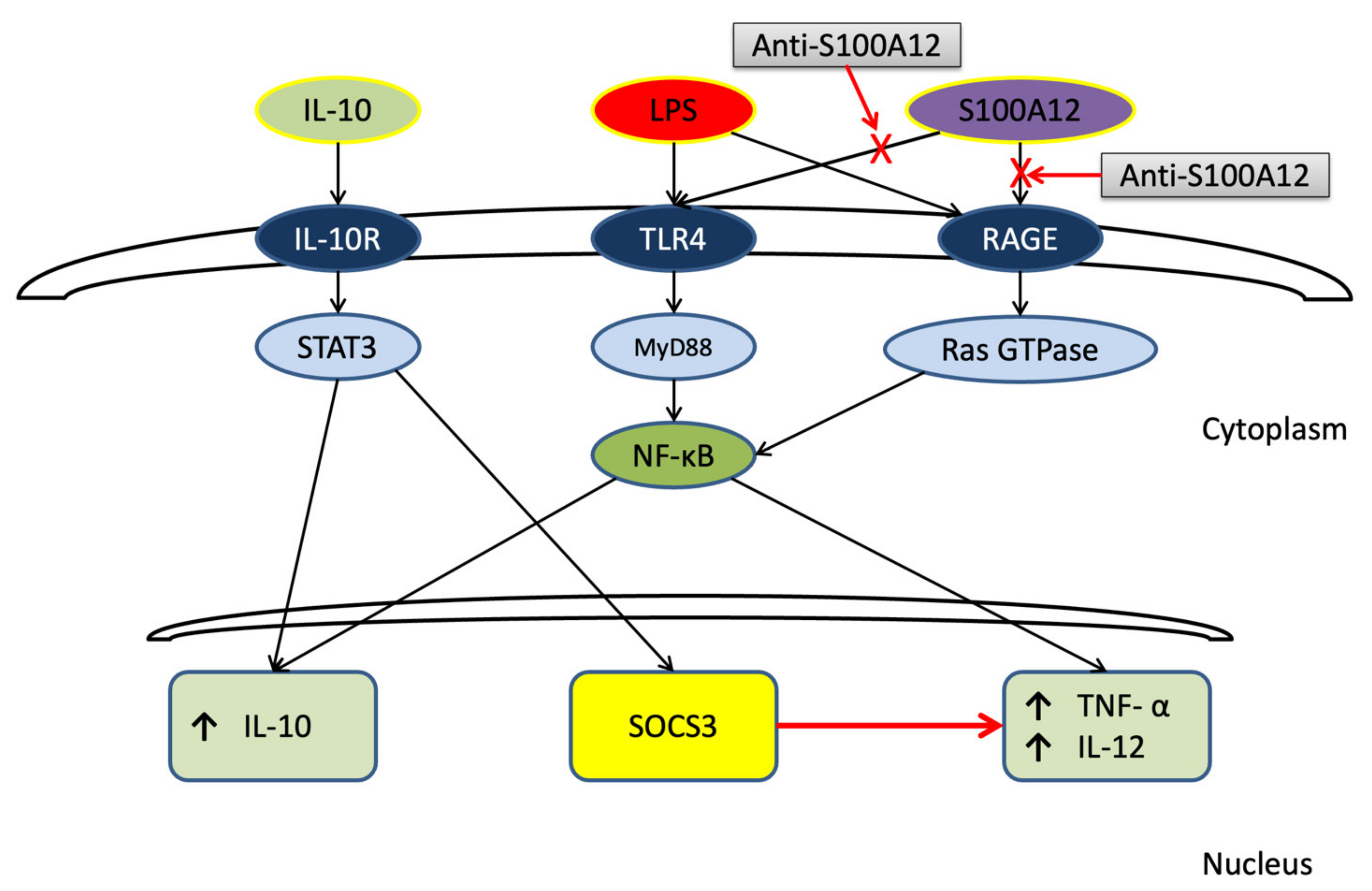
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.D.; Coopersmith, C.M.; et al. The Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). JAMA 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Venet, F.; Monneret, G. Advances in the understanding and treatment of sepsis-induced immunosuppression. Nat. Rev. Nephrol. 2018, 14, 121–137. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.P.; Chen, C.K.; Chung, K.; Tseng, J.C.; Hua, C.C.; Liu, Y.C.; Chuang, D.Y.; Yang, C.H. Serial cytokine levels in patients with severe sepsis. Inflamm. Res. 2009, 57, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.P.; Shih, C.C.; Lin, C.Y.; Hua, C.C.; Chuang, D.Y. Serial increase of IL-12 response and human leukocyte antigen-DR expression in severe sepsis survivors. Crit. Care 2011, 15, R224. [Google Scholar] [CrossRef] [PubMed]
- Frencken, J.F.; van Vught, L.A.; Peelen, L.M.; Ong, D.S.Y.; Klein Klouwenberg, P.M.C.; Horn, J.; Bonten, M.J.M.; van der Poll, T.; Cremer, O.L. An Unbalanced Inflammatory Cytokine Response Is Not Associated With Mortality Following Sepsis: A Prospective Cohort Study. Crit. Care Med. 2017, 45, e493–e499. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Harashima, A.; Saito, H.; Tsuneyama, K.; Munesue, S.; Motoyoshi, S.; Han, D.; Watanabe, T.; Asano, M.; Takasawa, S.; et al. Septic shock is associated with receptor for advanced glycation end products ligation of LPS. J. Immunol. 2011, 186, 3248–3257. [Google Scholar] [CrossRef] [PubMed]
- Rauvala, H.; Rouhiainen, A. Physiological and pathophysiological outcomes of the interactions of HMGB1 with cell surface receptors. Biochim. Biophys. Acta 2010, 1799, 164–170. [Google Scholar] [CrossRef]
- Bierhaus, A.; Stern, D.M.; Nawroth, P.P. RAGE in inflammation: A new therapeutic target? Curr. Opin. Investig. Drugs 2006, 7, 985–991. [Google Scholar]
- Park, L.; Raman, K.G.; Lee, K.J.; Lu, Y.; Ferran, L.J., Jr.; Chow, W.S.; Stern, D.; Schmidt, A.M. Suppression of accelerated diabetic atherosclerosis by the soluble receptor for advanced glycation endproducts. Nat. Med. 1998, 4, 1025–1031. [Google Scholar] [CrossRef]
- Cataldegirmen, G.; Zeng, S.; Feirt, N.; Ippagunta, N.; Dun, H.; Qu, W.; Lu, Y.; Rong, L.L.; Hofmann, M.A.; Kislinger, T.; et al. RAGE limits regeneration after massive liver injury by coordinated suppression of TNF-alpha and NF-kappaB. J. Exp. Med. 2005, 201, 473–484. [Google Scholar] [CrossRef]
- Goova, M.T.; Li, J.; Kislinger, T.; Qu, W.; Lu, Y.; Bucciarelli, L.G.; Nowygrod, S.; Wolf, B.M.; Caliste, X.; Yan, S.F.; et al. Blockade of receptor for advanced glycation end-products restores effective wound healing in diabetic mice. Am. J. Pathol. 2001, 159, 513–525. [Google Scholar] [CrossRef]
- Leclerc, E.; Fritz, G.; Vetter, S.W.; Heizmann, C.W. Binding of S100 proteins to RAGE: An update. Biochim. Biophys. Acta 2009, 1793, 993–1007. [Google Scholar] [CrossRef]
- Foell, D.; Wittkowski, H.; Kessel, C.; Luken, A.; Weinhage, T.; Varga, G.; Vogl, T.; Wirth, T.; Viemann, D.; Bjork, P.; et al. Proinflammatory S100A12 can activate human monocytes via Toll-like receptor 4. Am. J. Respir. Crit. Care Med. 2013, 187, 1324–1334. [Google Scholar] [CrossRef]
- Wu, R.; Liu, Y.; Yan, R.; Liu, X.; Duan, L. Assessment of EN-RAGE, sRAGE and EN-RAGE/sRAGE as potential biomarkers in patients with autoimmune hepatitis. J. Transl. Med. 2020, 18, 384. [Google Scholar] [CrossRef]
- Achouiti, A.; Foll, D.; Vogl, T.; van Till, J.W.; Laterre, P.F.; Dugernier, T.; Wittebole, X.; Boermeester, M.A.; Roth, J.; van der Poll, T.; et al. S100A12 and soluble receptor for advanced glycation end products levels during human severe sepsis. Shock 2013, 40, 188–194. [Google Scholar] [CrossRef]
- Uhel, F.; Azzaoui, I.; Gregoire, M.; Pangault, C.; Dulong, J.; Tadie, J.M.; Gacouin, A.; Camus, C.; Cynober, L.; Fest, T.; et al. Early Expansion of Circulating Granulocytic Myeloid-derived Suppressor Cells Predicts Development of Nosocomial Infections in Patients with Sepsis. Am. J. Respir. Crit. Care Med. 2017, 196, 315–327. [Google Scholar] [CrossRef]
- Tosson, A.M.S.; Glaser, K.; Weinhage, T.; Foell, D.; Aboualam, M.S.; Edris, A.A.; El Ansary, M.; Lotfy, S.; Speer, C.P. Evaluation of the S100 protein A12 as a biomarker of neonatal sepsis. J. Matern. Fetal Neonatal Med. 2020, 33, 2768–2774. [Google Scholar] [CrossRef]
- Dubois, C.; Marce, D.; Faivre, V.; Lukaszewicz, A.C.; Junot, C.; Fenaille, F.; Simon, S.; Becher, F.; Morel, N.; Payen, D. High plasma level of S100A8/S100A9 and S100A12 at admission indicates a higher risk of death in septic shock patients. Sci. Rep. 2019, 9, 15660. [Google Scholar] [CrossRef]
- Bellomo, R.; Ronco, C.; Kellum, J.A.; Mehta, R.L.; Palevsky, P. Acute renal failure—Definition, outcome measures, animal models, fluid therapy and information technology needs: The Second International Consensus Conference of the Acute Dialysis Quality Initiative (ADQI) Group. Crit. Care 2004, 8, R204–R212. [Google Scholar] [CrossRef]
- Knaus, W.A.; Draper, E.A.; Wagner, D.P.; Zimmerman, J.E. APACHE II: A severity of disease classification system. Crit. Care Med. 1985, 13, 818–829. [Google Scholar] [CrossRef]
- Rhodes, A.; Evans, L.E.; Alhazzani, W.; Levy, M.M.; Antonelli, M.; Ferrer, R.; Kumar, A.; Sevransky, J.E.; Sprung, C.L.; Nunnally, M.E.; et al. Surviving Sepsis Campaign: International Guidelines for Management of Sepsis and Septic Shock: 2016. Intensive Care Med. 2017, 43, 304–377. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, H.; Zhou, J.; Zhong, Y.; Ali, M.M.; McGuire, F.; Nagarkatti, P.S.; Nagarkatti, M. Role of cytokines as a double-edged sword in sepsis. In Vivo 2013, 27, 669–684. [Google Scholar]
- Ingels, C.; Derese, I.; Wouters, P.J.; Van den Berghe, G.; Vanhorebeek, I. Soluble RAGE and the RAGE ligands HMGB1 and S100A12 in critical illness: Impact of glycemic control with insulin and relation with clinical outcome. Shock 2015, 43, 109–116. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Matsumoto, H.; Matsumoto, N.; Ogura, H.; Shimazaki, J.; Yamakawa, K.; Yamamoto, K.; Shimazu, T. The clinical significance of circulating soluble RAGE in patients with severe sepsis. J. Trauma Acute Care Surg. 2015, 78, 1086–1093. [Google Scholar] [CrossRef] [PubMed]
- Narvaez-Rivera, R.M.; Rendon, A.; Salinas-Carmona, M.C.; Rosas-Taraco, A.G. Soluble RAGE as a severity marker in community acquired pneumonia associated sepsis. BMC Infect. Dis. 2012, 12, 15. [Google Scholar] [CrossRef]
- Jabaudon, M.; Futier, E.; Roszyk, L.; Chalus, E.; Guerin, R.; Petit, A.; Mrozek, S.; Perbet, S.; Cayot-Constantin, S.; Chartier, C.; et al. Soluble form of the receptor for advanced glycation end products is a marker of acute lung injury but not of severe sepsis in critically ill patients. Crit. Care Med. 2011, 39, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, O.; Akira, S. Toll-like receptors; their physiological role and signal transduction system. Int. Immunopharmacol. 2001, 1, 625–635. [Google Scholar] [CrossRef]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, H.; Wang, P.; Wang, J.; Zou, L. The roles of SOCS3 and STAT3 in bacterial infection and inflammatory diseases. Scand. J. Immunol. 2018, 88, e12727. [Google Scholar] [CrossRef]
- Bode, J.G.; Ehlting, C.; Haussinger, D. The macrophage response towards LPS and its control through the p38(MAPK)-STAT3 axis. Cell. Signal. 2012, 24, 1185–1194. [Google Scholar] [CrossRef]
- Gargalionis, A.N.; Papavassiliou, K.A.; Papavassiliou, A.G. Targeting STAT3 Signaling Pathway in Colorectal Cancer. Biomedicines 2021, 9, 1016. [Google Scholar] [CrossRef]
- Polimeno, L.; Francavilla, A.; Piscitelli, D.; Fiore, M.G.; Polimeno, R.; Topi, S.; Haxhirexha, K.; Ballini, A.; Daniele, A.; Santacroce, L. The role of PIAS3, p-STAT3 and ALR in colorectal cancer: New translational molecular features for an old disease. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 10496–10511. [Google Scholar] [CrossRef]
- Van Zoelen, M.A.; Achouiti, A.; van der Poll, T. The role of receptor for advanced glycation endproducts (RAGE) in infection. Crit. Care 2011, 15, 208. [Google Scholar] [CrossRef]
- Reynolds, P.R.; Kasteler, S.D.; Schmitt, R.E.; Hoidal, J.R. Receptor for advanced glycation end-products signals through Ras during tobacco smoke-induced pulmonary inflammation. Am. J. Respir. Cell Mol. Biol. 2011, 45, 411–418. [Google Scholar] [CrossRef]
- Ahmad, A.; Ahsan, H. Ras-Mediated Activation of NF-kappaB and DNA Damage Response in Carcinogenesis. Cancer Investig. 2020, 38, 185–208. [Google Scholar] [CrossRef]
- Wen, X.; Han, X.R.; Wang, Y.J.; Fan, S.H.; Zhang, Z.F.; Wu, D.M.; Lu, J.; Zheng, Y.L. Effects of S100A12 gene silencing on serum levels of anti-inflammatory/pro-inflammatory cytokines in septic rats through the ERK signaling pathway. J. Cell. Biochem. 2018, 119, 4038–4049. [Google Scholar] [CrossRef]
- Chung, Y.M.; Goyette, J.; Tedla, N.; Hsu, K.; Geczy, C.L. S100A12 suppresses pro-inflammatory, but not pro-thrombotic functions of serum amyloid A. PLoS ONE 2013, 8, e62372. [Google Scholar] [CrossRef]
- Zhang, Z.; Han, N.; Shen, Y. S100A12 promotes inflammation and cell apoptosis in sepsis-induced ARDS via activation of NLRP3 in fl ammasome signaling. Mol. Immunol. 2020, 122, 38–48. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Survivors (n = 30) | Non-Survivors (n = 14) | All Patients (n = 44) | Controls (n = 27) | |
|---|---|---|---|---|
| Age (years old) | 75.9 ± 1.8 | 74.7 ± 3.1 | 75.5 ± 1.6 | 60.3 ± 1.3 * |
| Male (%) | 17 (56.7) | 9 (64.3) | 26 (50.1) | 17 (63.0) |
| APACHE II score | 20.8 ± 1.0 | 28.4 ± 2.0 † | 23.2 ± 1.1 | |
| History (%) | ||||
| COPD | 0 (0.0) | 2 (14.3) | 2 (4.5) | |
| Heart failure | 5 (16.7) | 0 (0.0) | 5 (11.4) | |
| Hypertension | 20 (66.7) | 10 (71.4) | 30 (68.2) | |
| Diabetes mellitus | 12 (40.0) | 7 (50.0) | 19 (43.2) | |
| Old CVA | 8 (26.7) | 2 (14.3) | 10 (22.7) | |
| ESRD | 4 (13.3) | 4 (28.6) | 8 (18.2) | |
| Liver cirrhosis | 3 (10.0) | 2 (14.3) | 5 (11.4) | |
| Active malignancy | 2 (6.7) | 0 (0.0) | 2 (4.5) | |
| Infection source | ||||
| Pneumonia | 21 (70.0) | 8 (57.1) | 29 (65.9) | |
| UTI | 4 (13.3) | 1 (7.1) | 5 (11.4) | |
| Others | 5 (16.7) | 5 (35.8) | 10 (22.7) | |
| Adverse event | ||||
| New arrhythmia | 3 (10.0) | 1 (7.1) | 4 (9.1) | |
| GI bleeding | 1 (3.3) | 3 (21.4) | 4 (9.1) | |
| Acute renal failure | 8 (26.7) | 10 (71.4) ‡ | 18 (40.9) | |
| Shock | 13 (43.3) | 12 (85.7) ¶ | 25 (56.8) | |
| Thrombocytopenia | 8 (26.7) | 7 (50.0) | 15 (34.1) | |
| Jaundice | 5 (16.7) | 3 (21.4) | 8 (18.2) | |
| Bacteremia | 5 (16.7) | 2 (14.3) | 7 (15.9) |
| Survivors | Non-Survivors | All Patients | Controls | |
|---|---|---|---|---|
| Day 1 | (n = 30) | (n = 14) | (n = 44) | (n = 27) |
| AGE, ng/mL | 4335.9 ± 1249.4 | 3839.5 ± 1715.1 | 4177.9 ± 1001.0 | 2117.8 ± 833.4 |
| sRAGE, pg/mL | 2355.2 ± 314.9 | 2751.5 ± 650.0 | 2481.3 ± 295.0 | 1273.0 ± 108.2 * |
| HMGB1, pg/mL | 306.3 ± 86.1 | 247.4 ± 56.8 | 287.6 ± 61.1 | 160.7 ± 54.0 |
| S100A12, pg/mL | 533.4 ± 20.2 | 523.6 ± 38.8 | 530.3 ± 18.2 | 310.1 ± 28.1 * |
| Day 7 | (n = 29) | (n = 5) | (n = 34) | |
| AGE, ng/mL | 5372.0 ± 1254.3 | 10517.0 ± 6020.9 | 6128.7 ± 1373.1 | |
| sRAGE, pg/mL | 1530.1 ± 219.1 | 1041.6 ± 371.9 | 1458.2 ± 195.2 | |
| HMGB1, pg/mL | 324.6 ± 88.1 | 271.0 ± 114.0 | 316.7 ± 76.6 | |
| S100A12, pg/mL | 499.3 ± 23.8 | 593.1 ± 12.7 † | 513.1 ± 21.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.-P.; Chu, C.-M.; Liu, P.-H.; Leu, S.-W.; Lin, S.-W.; Hu, H.-C.; Kao, K.-C.; Li, L.-F.; Yu, C.-C. Increased Production of Interleukin-10 and Tumor Necrosis Factor-Alpha in Stimulated Peripheral Blood Mononuclear Cells after Inhibition of S100A12. Curr. Issues Mol. Biol. 2022, 44, 1701-1712. https://doi.org/10.3390/cimb44040117
Wu H-P, Chu C-M, Liu P-H, Leu S-W, Lin S-W, Hu H-C, Kao K-C, Li L-F, Yu C-C. Increased Production of Interleukin-10 and Tumor Necrosis Factor-Alpha in Stimulated Peripheral Blood Mononuclear Cells after Inhibition of S100A12. Current Issues in Molecular Biology. 2022; 44(4):1701-1712. https://doi.org/10.3390/cimb44040117
Chicago/Turabian StyleWu, Huang-Pin, Chien-Ming Chu, Pi-Hua Liu, Shaw-Woei Leu, Shih-Wei Lin, Han-Chung Hu, Kuo-Chin Kao, Li-Fu Li, and Chung-Chieh Yu. 2022. "Increased Production of Interleukin-10 and Tumor Necrosis Factor-Alpha in Stimulated Peripheral Blood Mononuclear Cells after Inhibition of S100A12" Current Issues in Molecular Biology 44, no. 4: 1701-1712. https://doi.org/10.3390/cimb44040117
APA StyleWu, H.-P., Chu, C.-M., Liu, P.-H., Leu, S.-W., Lin, S.-W., Hu, H.-C., Kao, K.-C., Li, L.-F., & Yu, C.-C. (2022). Increased Production of Interleukin-10 and Tumor Necrosis Factor-Alpha in Stimulated Peripheral Blood Mononuclear Cells after Inhibition of S100A12. Current Issues in Molecular Biology, 44(4), 1701-1712. https://doi.org/10.3390/cimb44040117