
Associations of APOE Gene Variants rs429358 and rs7412 with Parameters of the Blood Lipid Profile and the Risk of Myocardial Infarction and Death in a White Population of Western Siberia

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Measures and Clinical Data
2.3. Genotyping and Quality Control
2.4. Statistical Analyses
3. Results and Discussion
3.1. Frequencies of Alleles and Genotypes of rs429358 & rs7412 (the APOE Gene)
3.2. Associations of rs429358 & rs7412 with Parameters of the Blood Lipid Profile
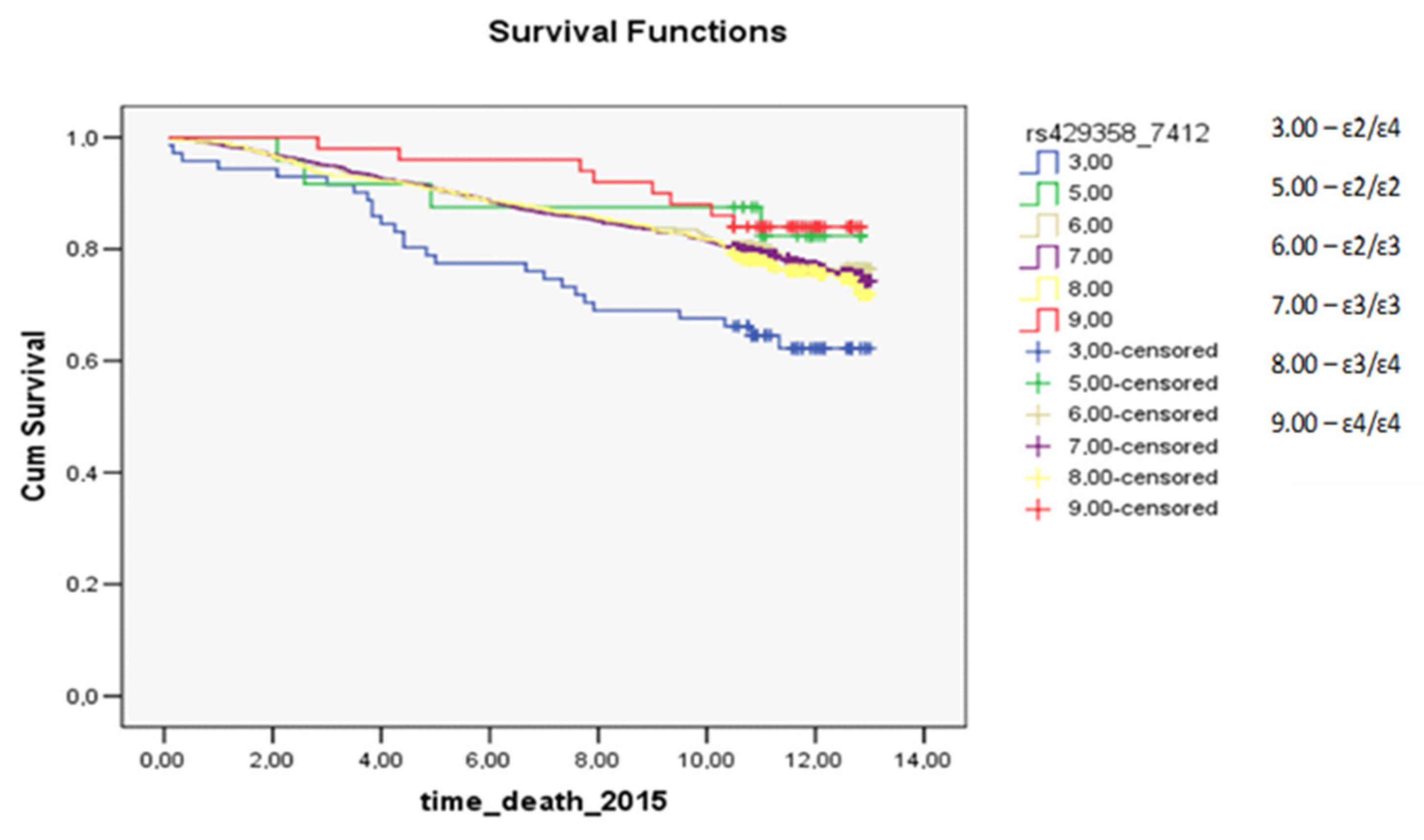
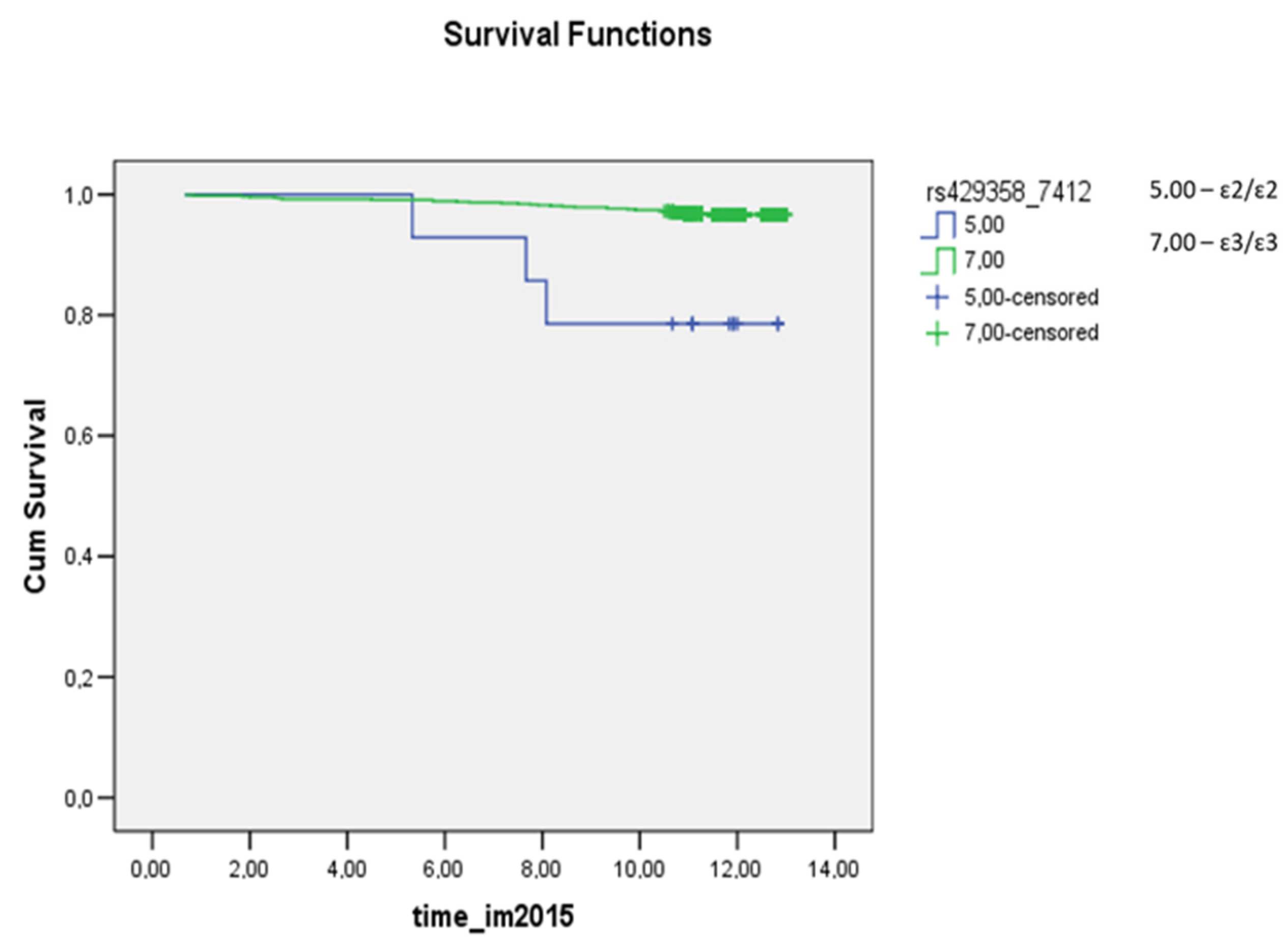
3.3. Survival Analysis for Carriers of Common Apolipoprotein E Isoforms
3.4. Discussion
3.5. Limitations
3.6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| HDL-C | high-density lipoprotein cholesterol |
| LDL-C | low-density lipoprotein cholesterol |
| PCR | polymerase chain reaction |
| TC | total cholesterol |
| TGs | triglycerides |
References
- Nielsen, S.H.; Mouton, A.J.; DeLeon-Pennell, K.Y.; Genovese, F.; Karsdal, M.; Lindsey, M. Understanding cardiac extracellular matrix remodeling to develop biomarkers of myocardial infarction outcomes. Matrix Biol. 2019, 75, 43–47. [Google Scholar] [CrossRef] [PubMed]
- Atar, D.; Jukema, J.W.; Molemans, B.; Taub, P.R.; Goto, S.; Mach, G.; CerezoOlmos, C.; Underberg, J.; Keech, A.; Tokgözoğlu, L.; et al. New cardiovascular prevention guidelines: How to optimally manage dyslipidaemia and cardiovascular risk in 2021 in patients needing secondary prevention? Atherosclerosis 2021, 319, 51–61. [Google Scholar] [CrossRef]
- Marias, A.D. Apolipoprotein E in lipoprotein metabolism, health and cardiovascular disease. Pathology 2019, 51, 165–176. [Google Scholar] [CrossRef]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef]
- Khalil, Y.A.; Rabès, J.P.; Boileau, C.; Varret, M. APOE gene variants in primary dyslipidemia. Atherosclerosis 2021, 328, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, A.L.; Mulugeta, A.; Zhou, A.; Hyppönen, E. Apolipoprotein E (APOE) genotype-associated disease risks: A phenome-wide, registry-based, case-control study utilising the UK Biobank. EBioMedicine 2020, 59, 102954. [Google Scholar] [CrossRef]
- Peasey, A.; Bobak, M.; Kubinova, R.; Malyutina, S.; Pajak, A.; Tamosiunas, A.; Pikhart, H.; Nicholson, A.; Marmot, M. Determinants of cardiovascular disease and other non-communicable diseases in Central and Eastern Europe: Rationale and design of the HAPIEE study. BMC Public Health 2006, 6, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Russel, D.W. Purification of nucleic acids by extraction with phenol:chloroform. CSH Protoc. 2006, 1, pdb.prot4455. [Google Scholar] [CrossRef] [PubMed]
- Rich, J.T.; Neely, J.G.; Paniello, R.C.; Voelker, C.C.J.; Nussenbaum, B.; Wang, E.W. A practical guide to understanding Kaplan-Meier curves. Otolaryngol. Head Neck Surg. 2010, 143, 331–336. [Google Scholar] [CrossRef] [Green Version]
- Karjalainen, J.P.; Mononen, N.; Hutri-Kähönen, N.; Lehtimäki, M.; Juonala, M.; Ala-Korpela, M.; Kähönen, M.; Raitakari, O.; Lehtimäki, T. The effect of apolipoprotein E polymorphism on serum metabolome–a population-based 10-year follow-up study. Sci. Rep. 2019, 9, 458. [Google Scholar] [CrossRef] [Green Version]
- Karjalainen, J.P.; Mononen, N.; Hutri-Kähönen, N.; Lehtimäki, M.; Hilvo, M.; Kauhanen, D.; Juonala, M.; Viikari, J.; Kähönen, M.; Raitakari, O.; et al. New evidence from plasma ceramides links apoE polymorphism to greater risk of coronary artery disease in Finnish adults. J. Lipid Res. 2019, 60, 1622–1629. [Google Scholar] [CrossRef] [PubMed]
- Wolters, F.J.; Yang, Q.; Biggs, M.L.; Jakobsdottir, J.; Li, S.; Evans, D.S.; Bis, J.C.; Harris, T.B.; Vasan, R.S.; Zilhao, N.S.; et al. E2-CHARGE investigators. The impact of APOE genotype on survival: Results of 38,537 participants from six population-based cohorts (E2-CHARGE). PLoS ONE 2019, 14, e0219668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adler, G.; Adler, M.A.; Urbańska, A.; Skonieczna-Żydecka, K.; Kiseljakovic, E.; Valjevac, A.; Parczewski, M.; Hadzovic-Dzuvo, A. Bosnian study of APOE distribution (BOSAD): A comparison with other European populations. Ann. Hum. Biol. 2017, 44, 568–573. [Google Scholar] [CrossRef]
- Saito, T.; Matsunaga, A.; Fukunaga, M.; Nagahama, K.; Hara, S.; Muso, E. Apolipoprotein E-related glomerular disorders. Kidney Int. 2020, 97, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Moriyama, K.; Sasaki, J.; Matsunaga, A.; Arakawa, F.; Takada, Y.; Araki, K.; Kaneko, S.; Arakawa, K. Apolipoprotein E1 Lys-146→Glu with type III hyperlipoproteinemia. Biochim. Biophys. Acta 1992, 1128, 58–64. [Google Scholar] [CrossRef]
- Limonova, A.S.; Ershova, A.I.; Meshkov, A.N.; Kiseleva, A.V.; Divashuk, M.G.; Kutsenko, V.A.; Drapkina, O.M. Case Report: Hypertriglyceridemia and Premature Atherosclerosis in a Patient With Apolipoprotein E Gene e2e1 Genotype. Front. Cardiovasc. Med. 2021, 7, 585779. [Google Scholar] [CrossRef] [PubMed]
- Koopal, C.; Geerlings, M.I.; Muller, M.; de Borst, G.J.; Algra, A.; van der Graaf, Y.; Visseren, F.L.J.; SMART Study Group. The relation between apolipoprotein E (APOE) genotype and peripheral artery disease in patients at high risk for cardiovascular disease. Atherosclerosis 2016, 246, 187–192. [Google Scholar] [CrossRef]
- Huebbe, P.; Rimbach, G. Evolution of human apolipoprotein E (APOE) isoforms: Gene structure, protein function and interaction with dietary factors. Ageing Res. Rev. 2017, 37, 146–161. [Google Scholar] [CrossRef]
- Bos, M.M.; de Vries, L.; Rensen, P.C.; van Dijk, K.W.; Blauw, G.J.; van Heemst, D.; Noordam, R. Apolipoprotein E genotype, lifestyle and coronary artery disease: Gene-environment interaction analyses in the UK Biobank population. Atherosclerosis 2021, 328, 33–37. [Google Scholar] [CrossRef]
- Bos, M.M.; Noordam, R.; Blauw, G.J.; Slagboom, P.E.; Rensen, P.C.N.; van Heemst, D. The ApoE ε4 Isoform: Can the Risk of Diseases be Reduced by Environmental Factors? J. Gerontol. Ser. A 2019, 74, 99–107. [Google Scholar] [CrossRef]
- Song, Y.; Stampfer, M.J.; Liu, S. Meta-analysis: Apolipoprotein E genotypes and risk for coronary heart disease. Ann. Intern. Med. 2004, 141, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Dudley, W.N.; Wickham, R.; Coombs, N. An Introduction to Survival Statistics: Kaplan-Meier Analysis. J. Adv. Pract. Oncol. 2016, 7, 91–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, K.L.; Tybjærg-Hansen, A.; Nordestgaard, B.G.; Frikke-Schmidt, R. Data on plasma levels of apolipoprotein E, correlations with lipids and lipoproteins stratified by APOE genotype, and risk of ischemic heart disease. Data Brief 2016, 6, 923–932. [Google Scholar] [CrossRef] [Green Version]
- Frikke-Schmidt, R.; Tybjaerg-Hansen, A.; Steffensen, R.; Jensen, G.; Nordestgaard, B.G. Apolipoprotein E genotype: Epsilon32 women are protected while epsilon43 and epsilon44 men are susceptible to ischemic heart disease: The Copenhagen City Heart Study. J. Am. Coll. Cardiol. 2000, 35, 1192–1199. [Google Scholar] [CrossRef] [Green Version]
- Kulminski, A.M.; Raghavachari, N.; Arbeev, K.G.; Culminskaya, I.; Arbeeva, L.; Wu, D.; Ukraintseva, S.V.; Christensen, K.; Yashin, A.I. Protective role of the apolipoprotein E2 allele in age-related disease traits and survival: Evidence from the Long Life Family Study. Biogerontology 2016, 17, 893–905. [Google Scholar] [CrossRef]
- Zeng, Y.; Nie, C.; Min, J.; Liu, X.; Li, M.; Chen, H.; Xu, H.; Wang, M.; Ni, T.; Li, Y.; et al. Novel loci and pathways significantly associated with longevity. Sci. Rep. 2016, 25, 21243. [Google Scholar] [CrossRef]
- Deelen, J.; Beekman, M.; Uh, H.W.; Broer, L.; Ayers, K.L.; Tan, Q.; Kamatani, Y.; Bennet, A.M.; Tamm, R.; Trompet, S.; et al. Genome-wide association meta-analysis of human longevity identifies a novel locus conferring survival beyond 90 years of age. Hum. Mol. Genet. 2014, 23, 4420–4432. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, H.; Yu, Z.; Huang, Q.; Zhong, Z. APOE gene ɛ4 allele (388C-526C) effects on serum lipids and risk of coronary artery disease in southern Chinese Hakka population. J. Clin. Lab. Anal. 2021, 35, e23925. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| rs429358 | rs7412 | rs429358 & rs7412 Genotype | ID for rs429358 & rs7412 Genotype in Survival Plots |
|---|---|---|---|
| (C;C) | (T;T) | ɛ1/ɛ1 | 1 |
| (C;T) | (T;T) | ɛ1/ɛ2 | 2 |
| (C;T) | (C;T) | ɛ2/ɛ4 | 3 |
| (C;C) | (C;T) | ɛ1/ɛ4 | 4 |
| (T;T) | (T;T) | ɛ2/ɛ2 | 5 |
| (T;T) | (C;T) | ɛ2/ɛ3 | 6 |
| (T;T) | (C;C) | ɛ3/ɛ3 | 7 |
| (C;T) | (C;C) | ɛ3/ɛ4 | 8 |
| (C;C) | (C;C) | ɛ4/ɛ4 | 9 |
| Males | Females | Both Sexes | |
|---|---|---|---|
| Number of subjects, n | 1263 | 1446 | 2709 |
| Age, years | 56.7 ± 0.2 | 56.6 ± 0.2 | 56.6 ± 0.1 |
| TC, mg/dL | 241.5 ± 1.4 | 259.2 ± 1.5 | 250.9 ± 1.1 |
| HDL-C, mg/dL | 58.3 ± 0.4 | 61.4 ± 0.5 | 59.9 ± 0.3 |
| LDL-C, mg/dL | 119.5 ± 1.3 | 132.4 ± 1.3 | 126.4 ± 0.9 |
| TGs, mg/dL | 141.6 ± 2.2 | 144.8 ± 2.2 | 143.3 ± 1.6 |
| Index of atherogenicity | 3.4 ± 0.04 | 3.4 ± 0.04 | 3.4 ± 0.03 |
| Body–mass index, kg/m2 | 27.2 ± 0.1 | 30.3 ± 0.2 | 28.8 ± 0.1 |
| Among Males | Among Females | In Both Sexes | |
|---|---|---|---|
| Frequency, Number of Subjects | Frequency, Number of Subjects | Frequency, Number of Subjects | |
| Genotypes | |||
| ɛ1/ɛ1 | - n = 0 | - n = 0 | - n = 0 |
| ɛ1/ɛ2 | 0.001 n = 1 | - n = 0 | 0.0005 n = 1 |
| ɛ2/ɛ4 | 0.032 n = 40 | 0.021 n = 31 | 0.026 n = 71 |
| ɛ1/ɛ4 | - n = 0 | 0.001 n = 1 | 0.0005 n = 1 |
| ɛ2/ɛ2 | 0.008 n = 10 | 0.010 n = 14 | 0.009 n = 24 |
| ɛ2/ɛ3 | 0.144 n = 182 | 0.127 n = 183 | 0.135 n = 365 |
| ɛ3/ɛ3 | 0.610 n = 771 | 0.618 n = 893 | 0.614 n = 1664 |
| ɛ3/ɛ4 | 0.188 n = 237 | 0.205 n = 296 | 0.197 n = 533 |
| ɛ4/ɛ4 | 0.017 n = 22 | 0.019 n = 28 | 0.018 n = 50 |
| Allele frequencies | |||
| ε1 | 0.0004 | 0.0004 | 0.0004 |
| ε2 | 0.0962 | 0.0837 | 0.0895 |
| ε3 | 0.7763 | 0.7832 | 0.7799 |
| ε4 | 0.1271 | 0.1328 | 0.1301 |
| Sex | Genotype | TC, mg/dL | HDL-C, mg/dL | LDL-C, mg/dL | TGs, mg/dL | Index of Atherogenicity |
|---|---|---|---|---|---|---|
| Males | ɛ1/ɛ1 | - | - | - | - | - |
| ɛ1/ɛ2 | 206.9 ± 49.2 | 52.6 ± 14.8 | 124.2 ± 43.8 | 67.0 ± 72.7 | 3.0 ± 1.3 | |
| ɛ2/ɛ4 | 254.3 ± 7.8 | 56.1 ± 2.4 | 119.7 ± 6.9 | 174.4 ± 11.5 | 3.8 ± 0.2 | |
| ɛ1/ɛ4 | - | - | - | - | - | |
| ɛ2/ɛ2 | 227.9 ± 15.6 | 52.3 ± 4.7 | 93.5 ± 13.9 | 182.7 ± 23.0 | 3.5 ± 0.4 | |
| ɛ2/ɛ3 | 230.4 ± 3.7 | 58.3 ± 1.1 | 103.2 ± 3.3 | 153.3 ± 5.4 | 3.2 ± 0.1 | |
| ɛ3/ɛ3 | 241.3 ± 1.8 | 59.2 ± 0.5 | 121.1 ± 1.6 | 135.5 ± 2.6 | 3.3 ± 0.1 | |
| ɛ3/ɛ4 | 244.6 ± 3.2 | 56.3 ± 1.0 | 125.7 ± 2.9 | 139.1 ± 4.8 | 3.6 ± 0.1 | |
| ɛ4/ɛ4 | 245.0 ± 10.5 | 55.3 ± 3.2 | 125.9 ± 9.4 | 141.8 ± 15.5 | 3.6 ± 0.3 | |
| p | 0.033 | 0.100 | <0.0001 * | 0.001 * | 0.003 * | |
| Females | ɛ1/ɛ1 | - | - | - | - | - |
| ɛ1/ɛ2 | - | - | - | - | - | |
| ɛ2/ɛ4 | 254.0 ± 9.9 | 60.0 ± 3.5 | 127.7 ± 8.9 | 137.7 ± 14.4 | 3.4 ± 0.3 | |
| ɛ1/ɛ4 | 359.6 ± 55.2 | 58.0 ± 19.2 | 214.3 ± 48.9 | 194.0 ± 78.9 | 5.3 ± 1.4 | |
| ɛ2/ɛ2 | 319.8 ± 14.7 | 58.3 ± 5.2 | 142.8 ± 13.1 | 264.0 ± 21.1 | 4.7 ± 0.4 | |
| ɛ2/ɛ3 | 245.0 ± 4.1 | 63.9 ± 1.4 | 114.7 ± 3.6 | 147.7 ± 5.9 | 3.1 ± 0.1 | |
| ɛ3/ɛ3 | 258.2 ± 1.9 | 61.6 ± 0.7 | 133.6 ± 1.7 | 139.6 ± 2.7 | 3.4 ± 0.1 | |
| ɛ3/ɛ4 | 264.5 ± 3.2 | 59.5 ± 1.1 | 138.5 ± 2.9 | 148.0 ± 4.6 | 3.7 ± 0.1 | |
| ɛ4/ɛ4 | 255.1 ± 10.6 | 63.1 ± 1.7 | 128.8 ± 9.4 | 140.3 ± 15.2 | 3.3 ± 0.3 | |
| p | <0.0001 * | 0.346 | <0.0001 * | <0.0001 * | <0.0001 * | |
| Both sexes | ɛ1/ɛ1 | - | - | - | - | - |
| ɛ1/ɛ2 | 215.8 ± 52.9 | 53.4 ± 17.4 | 130.4 ± 46.7 | 70.9 ± 76.5 | 3.1 ± 1.3 | |
| ɛ2/ɛ4 | 255.3 ± 6.3 | 58.3 ± 2.1 | 124.1 ± 5.6 | 158.2 ± 9.1 | 3.6 ± 0.2 | |
| ɛ1/ɛ4 | 356.8 ± 52.9 | 55.5 ± 17.4 | 212.0 ± 46.7 | 198.8 ± 76.5 | 5.4 ± 1.3 | |
| ɛ2/ɛ2 | 281.3 ± 10.8 | 55.2 ± 3.6 | 122.0 ± 9.5 | 231.4 ± 15.6 | 4.3 ± 0.3 | |
| ɛ2/ɛ3 | 237.8 ± 2.8 | 61.3 ± 0.9 | 109.0 ± 2.5 | 150.0 ± 4.0 | 3.1 ± 0.1 | |
| ɛ3/ɛ3 | 250.3 ± 1.3 | 60.5 ± 0.4 | 127.7 ± 1.2 | 137.7 ± 1.9 | 3.4 ± 0.03 | |
| ɛ3/ɛ4 | 255.5 ± 2.3 | 58.0 ± 0.8 | 132.7 ± 2.0 | 144.2 ± 3.3 | 3.6 ± 0.1 | |
| ɛ4/ɛ4 | 251.7 ± 7.6 | 59.1 ± 2.5 | 128.2 ± 6.7 | 143.0 ± 10.9 | 3.5 ± 0.2 | |
| p | <0.0001 * | 0.071 | <0.0001 * | <0.0001 * | <0.0001 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semaev, S.; Shakhtshneider, E.; Shcherbakova, L.; Ivanoshchuk, D.; Orlov, P.; Malyutina, S.; Gafarov, V.; Ragino, Y.; Voevoda, M. Associations of APOE Gene Variants rs429358 and rs7412 with Parameters of the Blood Lipid Profile and the Risk of Myocardial Infarction and Death in a White Population of Western Siberia. Curr. Issues Mol. Biol. 2022, 44, 1713-1724. https://doi.org/10.3390/cimb44040118
Semaev S, Shakhtshneider E, Shcherbakova L, Ivanoshchuk D, Orlov P, Malyutina S, Gafarov V, Ragino Y, Voevoda M. Associations of APOE Gene Variants rs429358 and rs7412 with Parameters of the Blood Lipid Profile and the Risk of Myocardial Infarction and Death in a White Population of Western Siberia. Current Issues in Molecular Biology. 2022; 44(4):1713-1724. https://doi.org/10.3390/cimb44040118
Chicago/Turabian StyleSemaev, Sergey, Elena Shakhtshneider, Liliya Shcherbakova, Dinara Ivanoshchuk, Pavel Orlov, Sophia Malyutina, Valery Gafarov, Yuliya Ragino, and Mikhail Voevoda. 2022. "Associations of APOE Gene Variants rs429358 and rs7412 with Parameters of the Blood Lipid Profile and the Risk of Myocardial Infarction and Death in a White Population of Western Siberia" Current Issues in Molecular Biology 44, no. 4: 1713-1724. https://doi.org/10.3390/cimb44040118
APA StyleSemaev, S., Shakhtshneider, E., Shcherbakova, L., Ivanoshchuk, D., Orlov, P., Malyutina, S., Gafarov, V., Ragino, Y., & Voevoda, M. (2022). Associations of APOE Gene Variants rs429358 and rs7412 with Parameters of the Blood Lipid Profile and the Risk of Myocardial Infarction and Death in a White Population of Western Siberia. Current Issues in Molecular Biology, 44(4), 1713-1724. https://doi.org/10.3390/cimb44040118