
Temporal Modulation of Drug Desensitization Procedures


Abstract
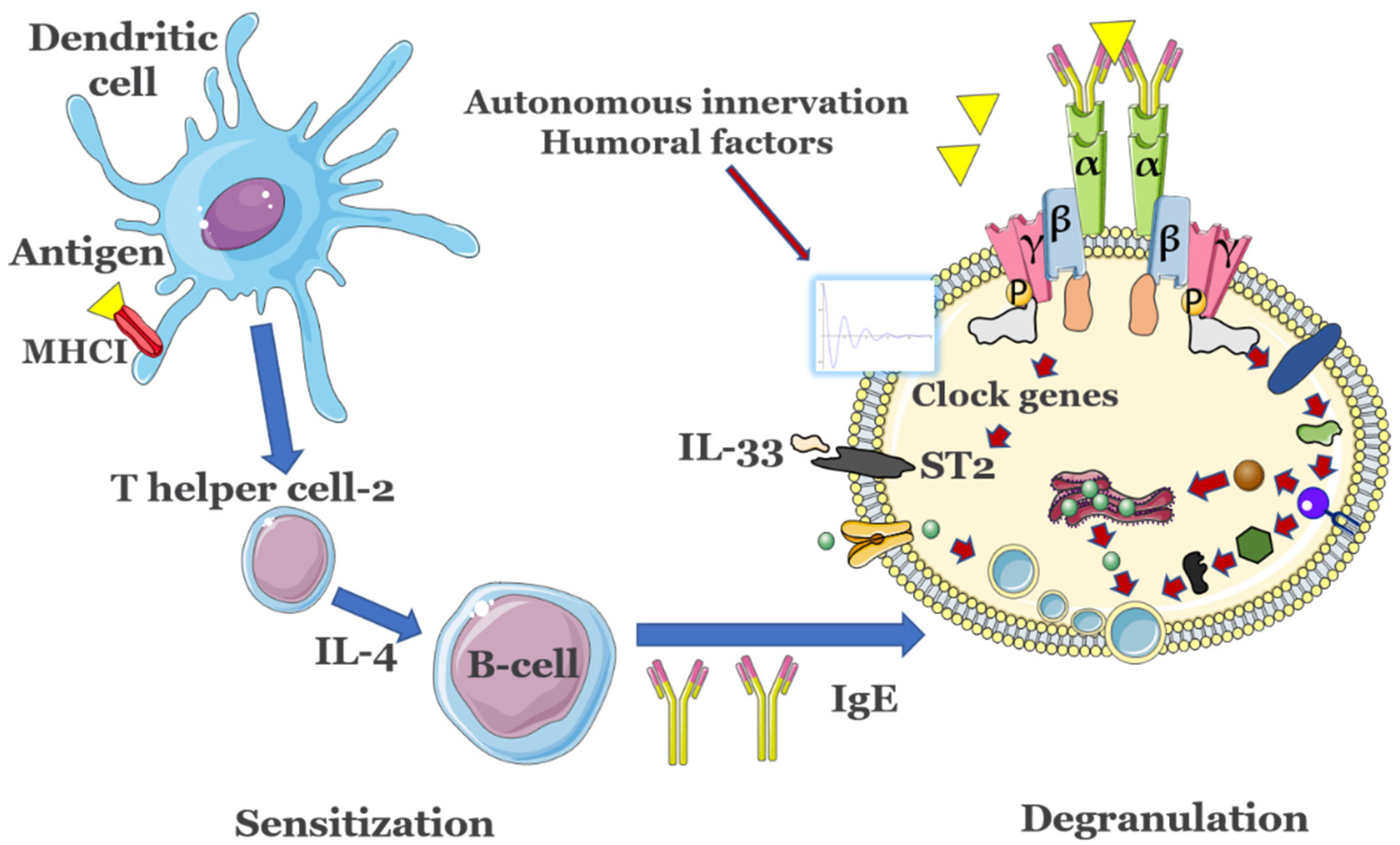
:1. Introduction
2. Drug Desensitization: Positive and Negative Role of Mast Cells
3. Temporal Aspects of DS Procedures
3.1. Circadian Clock Modulation of IgE Reactions
3.2. Time Interval Control in DS Steps
3.2.1. DS Is Directly Dependent on the Interval between Doses
3.2.2. Modulation of the Intervals between Doses
3.2.3. Unique Dose vs. Sequential Desensitization
3.2.4. Temporal Modulation of DS Procedures May Be Just as Important as the Amount of the Administered Drug Ex Vivo
3.2.5. Increasing the Dose Is Not as Effective as Increasing the Number and Duration of the Intervals
4. Conclusions and Outlook
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sánchez, L.D.L.V.; Alenazy, L.A.; Garcia-Neuer, M.; Castells, M.C. Drug Hypersensitivity and Desensitizations: Mechanisms and New Approaches. Int. J. Mol. Sci. 2017, 18, 1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vultaggio, A.; Matucci, A.; Nencini, F.; Bormioli, S.; Vivarelli, E.; Maggi, E. Mechanisms of Drug Desensitization: Not Only Mast Cells. Front. Pharmacol. 2020, 11, 991. [Google Scholar] [CrossRef] [PubMed]
- Cernadas, J.R.; Brockow, K.; Romano, A.; Aberer, W.; Torres, M.J.; Bircher, A.; Campi, P.; Sanz, M.L.; Castells, M.; Demoly, P.; et al. General considerations on rapid desensitization for drug hypersensitivity—a consensus statement. Allergy 2010, 65, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Akarsu, A.; Soyer, O.; Sekerel, B.E. Hypersensitivity Reactions to Biologicals: From Bench to Bedside. Curr. Treat. Options Allergy 2020, 7, 71–83. [Google Scholar] [CrossRef]
- Steinman, R.M.; Mellman, I.S.; Muller, W.A.; Cohn, Z.A. Endocytosis and the recycling of plasma membrane. J. Cell Biol. 1983, 96, 1–27. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Enríquez, E.; Hallgren, J. Mast Cells and Their Progenitors in Allergic Asthma. Front Immunol. 2019, 10, 821. [Google Scholar] [CrossRef] [Green Version]
- Demarest, S.J.; Hopp, J.; Chung, J.; Hathaway, K.; Mertsching, E.; Cao, X.; George, J.; Miatkowski, K.; LaBarre, M.J.; Shields, M.; et al. An Intermediate pH Unfolding Transition Abrogates the Ability of IgE to Interact with Its High Affinity Receptor FcϵRIα. J. Biol. Chem. 2006, 281, 30755–30767. [Google Scholar] [CrossRef] [Green Version]
- Wollman, R.; Meyer, T. Coordinated oscillations in cortical actin and Ca2+ correlate with cycles of vesicle secretion. Nat. Cell Biol. 2012, 14, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Blank, U.; Huang, H.; Kawakami, T. The high affinity IgE receptor: A signaling update. Curr. Opin. Immunol. 2021, 72, 51–58. [Google Scholar] [CrossRef]
- Smith, K.G.C.; Clatworthy, M.R. FcγRIIB in autoimmunity and infection: Evolutionary and therapeutic implications. Nat. Rev. Immunol. 2010, 10, 328–343. [Google Scholar] [CrossRef] [Green Version]
- Murphy-Schafer, A.R.; Paust, S. Divergent Mast Cell Responses Modulate Antiviral Immunity During Influenza Virus Infection. Front. Cell. Infect. Microbiol. 2021, 11, 580679. [Google Scholar] [CrossRef] [PubMed]
- Di Capite, J.; Parekh, A.B. CRAC channels and Ca2+signaling in mast cells. Immunol. Rev. 2009, 231, 45–58. [Google Scholar] [CrossRef]
- Vasilev, F.; Ezhova, Y.; Chun, J.T. Signaling Enzymes and Ion Channels Being Modulated by the Actin Cytoskeleton at the Plasma Membrane. Int. J. Mol. Sci. 2021, 22, 10366. [Google Scholar] [CrossRef] [PubMed]
- Sancho-Serra, M.D.C.; Simarro, M.; Castells, M. Rapid IgE desensitization is antigen specific and impairs early and late mast cell responses targeting FcεRI internalization. Eur. J. Immunol. 2011, 41, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Ang, W.G.; Church, A.M.; Kulis, M.; Choi, H.W.; Burks, A.W.; Abraham, S.N. Anaphylaxis in the 21st century: Phenotypes, endotypes, and biomarkers. J. Clin. Investig. 2016, 126, 4103–4118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacGlashan, D. Subthreshold desensitization of human basophils re-capitulates the loss of Syk and FcεRI expression characterized by other methods of desensitization. Clin. Exp. Allergy 2012, 42, 1060–1070. [Google Scholar] [CrossRef] [Green Version]
- Nejad, A.S.; MacGlashan, D.W., Jr. Dependence of Optimal Histamine Release on Cell Surface IgE Density on Human Basophils: Nature of the Stimulus. Int. Arch. Allergy Immunol. 2018, 177, 181–191. [Google Scholar] [CrossRef]
- Mahajan, A.; Barua, D.; Cutler, P.; Lidke, D.S.; Espinoza, F.A.; Pehlke, C.; Grattan, R.; Kawakami, Y.; Tung, C.-S.; Bradbury, A.R.M.; et al. Optimal Aggregation of FcεRI with a Structurally Defined Trivalent Ligand Overrides Negative Regulation Driven by Phosphatases. ACS Chem. Biol. 2014, 9, 1508–1519. [Google Scholar] [CrossRef]
- MacGlashan, D.W., Jr.; Peters, S.P.; Warner, J. Lichtenstein LM. Characteristics of human basophil sulfidopeptide leukotriene re-lease: Releasability defined as the ability of the basophil to respond to dimeric cross-links. J. Immunol. 1986, 15, 2231–2239. [Google Scholar]
- Luecke, S.; Sheu, K.M.; Hoffmann, A. Stimulus-specific responses in innate immunity: Multilayered regulatory circuits. Immunity 2021, 54, 1915–1932. [Google Scholar] [CrossRef]
- Lewis, A.; MacGlashan, D.W.; Suvarna, S.K.; Peachell, P.T. Recovery from desensitization of IgE-dependent responses in human lung mast cells. Clin. Exp. Allergy 2017, 47, 1022–1031. [Google Scholar] [CrossRef] [PubMed]
- Pichler, W.J.; Hausmann, O. Classification of Drug Hypersensitivity into Allergic, p-i, and Pseudo-Allergic Forms. Int. Arch. Allergy Immunol. 2016, 171, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Tontini, C.; Bulfone-Paus, S. Novel Approaches in the Inhibition of IgE-Induced Mast Cell Reactivity in Food Allergy. Front. Immunol. 2021, 12, 613461. [Google Scholar] [CrossRef] [PubMed]
- Kubo, S.; Matsuoka, K.; Taya, C.; Kitamura, F.; Takai, T.; Yonekawa, H.; Karasuyama, H. Drastic Up-Regulation of FcεRI on Mast Cells Is Induced by IgE Binding Through Stabilization and Accumulation of FcεRI on the Cell Surface. J. Immunol. 2001, 167, 3427–3434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, G. Current Strategies to Inhibit High Affinity FcεRI-Mediated Signaling for the Treatment of Allergic Disease. Front. Immunol. 2019, 10, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacGlashan, D., Jr.; Xia, H.Z.; Schwartz, L.B.; Gong, J. IgE-regulated loss, not IgE-regulated synthesis, controls expression of FcepsilonRI in human basophils. J. Leukoc. Biol. 2001, 70, 207–218. [Google Scholar] [PubMed]
- Ali, E.A.; Kalli, M.; Wan, D.; Nakamura, R.; Onion, D.; Alanine, D.G.W.; Alcocer, M.J.C.; Falcone, F.H. Characterization of human FcεRIα chain expression and gene copy number in humanized rat basophilic leukaemia (RBL) reporter cell lines. PLoS ONE 2019, 14, e0221034. [Google Scholar] [CrossRef] [PubMed]
- Oetjen, L.K.; Noti, M.; Kim, B.S. New insights into basophil heterogeneity. Semin. Immunopathol. 2016, 38, 549–561. [Google Scholar] [CrossRef] [Green Version]
- Shamji, M.H.; Valenta, R.; Jardetzky, T.; Verhasselt, V.; Durham, S.R.; Würtzen, P.A.; van Neerven, R.J. The role of allergen-specific IgE, IgG and IgA in allergic disease. Allergy 2021, 76, 3627–3641. [Google Scholar] [CrossRef]
- Frieri, M. Mast Cell Activation Syndrome. Clin. Rev. Allergy Immunol. 2015, 54, 353–365. [Google Scholar] [CrossRef]
- Kepley, C.L. Antigen-Induced Reduction in Mast Cell and Basophil Functional Responses due to Reduced Syk Protein Levels. Int. Arch. Allergy Immunol. 2005, 138, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyman, O.; Comte, D.; Spertini, F. Adverse reactions to biologic agents and their medical management. Nat. Rev. Rheumatol. 2014, 10, 612–627. [Google Scholar] [CrossRef] [PubMed]
- Pichler, W.J. Anaphylaxis to drugs: Overcoming mast cell unresponsiveness by fake antigens. Allergy 2020, 76, 1340–1349. [Google Scholar] [CrossRef] [PubMed]
- Harmon, B.; Chylek, L.A.; Liu, Y.; Mitra, E.D.; Mahajan, A.; Saada, E.A.; Schudel, B.R.; Holowka, D.A.; Baird, B.A.; Wilson, B.S.; et al. Timescale Separation of Positive and Negative Signaling Creates History-Dependent Responses to IgE Receptor Stimulation. Sci. Rep. 2017, 7, 15586. [Google Scholar] [CrossRef] [Green Version]
- Ménasché, G.; Longé, C.; Bratti, M.; Blank, U. Cytoskeletal Transport, Reorganization, and Fusion Regulation in Mast Cell-Stimulus Secretion Coupling. Front. Cell Dev. Biol. 2021, 9, 652077. [Google Scholar] [CrossRef]
- MacGlashan, D.; Vilariño, N. Polymerization of actin does not regulate desensitization in human basophils. J. Leukoc. Biol. 2009, 85, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Hishinuma, S.; Nozawa, H.; Akatsu, C.; Shoji, M. C-terminal of human histamine H1receptors regulates their agonist-induced clathrin-mediated internalization and G-protein signaling. J. Neurochem. 2016, 139, 552–565. [Google Scholar] [CrossRef] [Green Version]
- Agier, J.; Różalska, S.; Wódz, K.; Brzezińska-Błaszczyk, E. Leukotriene receptor expression in mast cells is affected by their agonists. Cell. Immunol. 2017, 317, 37–47. [Google Scholar] [CrossRef]
- Listos, J.; Łupina, M.; Talarek, S.; Mazur, A.; Orzelska-Górka, J.; Kotlińska, J. The Mechanisms Involved in Morphine Addiction: An Overview. Int. J. Mol. Sci. 2019, 20, 4302. [Google Scholar] [CrossRef] [Green Version]
- Sigismund, S.; Lanzetti, L.; Scita, G.; Di Fiore, P.P. Endocytosis in the context-dependent regulation of individual and collective cell properties. Nat. Rev. Mol. Cell Biol. 2021, 22, 625–643. [Google Scholar] [CrossRef]
- Stan, R.C.; Bhatt, D.; Camargo, M. Cellular Adaptation Relies on Regulatory Proteins Having Episodic Memory: Proteins Modulate Cell Metabolism and Reproduction by Remembering, Transmitting, and Using Data on the Environment. BioEssays 2019, 42, e1900115. [Google Scholar] [CrossRef]
- Tanaka, S.; Furuta, K. Roles of IgE and Histamine in Mast Cell Maturation. Cells 2021, 10, 2170. [Google Scholar] [CrossRef] [PubMed]
- Kitaura, J.; Song, J.; Tsai, M.; Asai, K.; Maeda-Yamamoto, M.; Mócsai, A.; Kawakami, Y.; Liu, F.-T.; Lowell, C.A.; Barisas, B.G.; et al. Evidence that IgE molecules mediate a spectrum of effects on mast cell survival and activation via aggregation of the Fc RI. Proc. Natl. Acad. Sci. USA 2003, 100, 12911–12916. [Google Scholar] [CrossRef] [Green Version]
- Kalesnikoff, J.; Huber, M.; Lam, V.; Damen, J.E.; Zhang, J.; Siraganian, R.P.; Krystal, G. Monomeric IgE Stimulates Signaling Pathways in Mast Cells that Lead to Cytokine Production and Cell Survival. Immunity 2001, 14, 801–811. [Google Scholar] [CrossRef] [Green Version]
- Cantaruti, T.; Costa, R.A.; De Souza, K.S.; Vaz, N.M.; Carvalho, C.R. Indirect effects of immunological tolerance to a regular dietary protein reduce cutaneous scar formation. Immunology 2017, 151, 314–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalit, M.; Levi-Schaffer, F. Challenge of mast cells with increasing amounts of antigen induces desensitization. Clin. Exp. Allergy 1995, 25, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Sieber, J.; Renner, S.; Lakatos-Krepcik, A.; Szépfalusi, Z. Case Report: Maintenance of Desensitization to Nebulized Colomycin Over 10 Years. Front. Pediatr. 2021, 9, 663228. [Google Scholar] [CrossRef] [PubMed]
- Tsao, L.R.; Young, F.D.; Otani, I.M.; Castells, M.C. Hypersensitivity Reactions to Platinum Agents and Taxanes. Clin. Rev. Allergy Immunol. 2021, 1–17. [Google Scholar] [CrossRef]
- Walton, J.C.; Ii, W.H.W.; Bumgarner, J.R.; Meléndez-Fernández, O.H.; Liu, J.A.; Hughes, H.L.; Kaper, A.L.; Nelson, R.J. Circadian Variation in Efficacy of Medications. Clin. Pharmacol. Ther. 2021, 109, 1457–1488. [Google Scholar] [CrossRef]
- Panzer, S.E.; Dodge, A.M.; Kelly, E.; Jarjour, N.N. Circadian variation of sputum inflammatory cells in mild asthma. J. Allergy Clin. Immunol. 2003, 111, 308–312. [Google Scholar] [CrossRef]
- Waldmann, T.A.; Iio, A.; Ogawa, M.; McIntyre, O.R.; Strober, W. The metabolism of IgE. Studies in normal individuals and in a patient with IgE myeloma. J. Immunol. 1976, 117, 1139–1144. [Google Scholar]
- Kader, H.; Azeem, M.; Jwayed, S.; Al-Shehhi, A.; Tabassum, A.; Ayoub, M.; Hetta, H.; Waheed, Y.; Iratni, R.; Al-Dhaheri, A.; et al. Current Insights into Immunology and Novel Therapeutics of Atopic Dermatitis. Cells 2021, 10, 1392. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, K.; Kitagawa, E.; Wada, M.; Haraguchi, A.; Orihara, K.; Tahara, Y.; Nakao, A.; Shibata, S. Antigen exposure in the late light period induces severe symptoms of food allergy in an OVA-allergic mouse model. Sci. Rep. 2015, 5, 14424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kolkhir, P.; Elieh-Ali-Komi, D.; Metz, M.; Siebenhaar, F.; Maurer, M. Understanding human mast cells: Lesson from therapies for allergic and non-allergic diseases. Nat. Rev. Immunol. 2021, 1–15. [Google Scholar] [CrossRef]
- Christ, P.; Sowa, A.S.; Froy, O.; Lorentz, A. The Circadian Clock Drives Mast Cell Functions in Allergic Reactions. Front. Immunol. 2018, 9, 1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pick, R.; He, W.; Chen, C.-S.; Scheiermann, C. Time-of-Day-Dependent Trafficking and Function of Leukocyte Subsets. Trends Immunol. 2019, 40, 524–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, Y.; Nakano, N.; Ishimaru, K.; Ando, N.; Katoh, R.; Suzuki-Inoue, K.; Koyanagki, S.; Ogawa, H.; Okumura, K.; Shibata, S.; et al. Inhibition of IgE-mediated allergic reactions by pharmacologically targeting the circadian clock. J. Allergy Clin. Immunol. 2015, 137, 1226–1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayasaka, N.; Yaita, T.; Kuwaki, T.; Honma, S.; Honma, K.-I.; Kudo, T.; Shibata, S. Optimization of Dosing Schedule of Daily Inhalant Dexamethasone to Minimize Phase Shifting of Clock Gene Expression Rhythm in the Lungs of the Asthma Mouse Model. Endocrinology 2007, 148, 3316–3326. [Google Scholar] [CrossRef] [Green Version]
- Duan, J.; Greenberg, E.N.; Karri, S.S.; Andersen, B. The circadian clock and diseases of the skin. FEBS Lett. 2021, 595, 2413–2436. [Google Scholar] [CrossRef]
- Baumann, A.; Gönnenwein, S.; Bischoff, S.C.; Sherman, H.; Chapnik, N.; Froy, O.; Lorentz, A. The circadian clock is functional in eosinophils and mast cells. Immunology 2013, 140, 465–474. [Google Scholar] [CrossRef]
- Wendell, S.G.; Fan, H.; Zhang, C. G Protein–Coupled Receptors in Asthma Therapy: Pharmacology and Drug Action. Pharmacol. Rev. 2019, 72, 1–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minaldi, E.; Phillips, E.J.; Norton, A. Immediate and Delayed Hypersensitivity Reactions to Beta-Lactam Antibiotics. Clin. Rev. Allergy Immunol. 2021, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Mawhirt, S.L.; Fonacier, L.S.; Calixte, R.; Davis-Lorton, M.; Aquino, M.R. Skin testing and desensitization outcomes among platinum-sensitive oncology patients. Ann. Allergy Asthma Immunol. 2018, 120, 437–439. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.R.; Shah, N.; Castells, M. Antigen-IgE desensitization in signal transducer and activator of transcription 6-deficient mast cells by suboptimal doses of antigen. Ann. Allergy Asthma Immunol. 2005, 94, 575–580. [Google Scholar] [CrossRef]
- Caiado, J.; Brás, R.; Paulino, M.; Costa, L.; Castells, M. Rapid desensitization to antineoplastic drugs in an outpatient immunoallergology clinic. Ann. Allergy Asthma Immunol. 2020, 125, 325–333. [Google Scholar] [CrossRef]
- Hemmings, O.; Kwok, M.; McKendry, R.; Santos, A.F. Basophil Activation Test: Old and New Applications in Allergy. Curr. Allergy Asthma Rep. 2018, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- MacGlashan, D. Loss of receptors and IgE in vivo during treatment with anti-IgE antibody. J. Allergy Clin. Immunol. 2004, 114, 1472–1474. [Google Scholar] [CrossRef]
- Norton, A.E.; Konvinse, K.; Phillips, E.J.; Broyles, A.D. Antibiotic Allergy in Pediatrics. Pediatrics 2018, 141, e20172497. [Google Scholar] [CrossRef] [Green Version]
- Duan, S.; Arlian, B.M.; Nycholat, C.M.; Wei, Y.; Tateno, H.; Smith, S.A.; Macauley, M.S.; Zhu, Z.; Bochner, B.S.; Paulson, J.C. Nanoparticles Displaying Allergen and Siglec-8 Ligands Suppress IgE-FcεRI–Mediated Anaphylaxis and Desensitize Mast Cells to Subsequent Antigen Challenge. J. Immunol. 2021, 206, 2290–2300. [Google Scholar] [CrossRef]
- Wen, S.; Krohn, I.K.; Thuraiaiyah, J.; Skjold, T.; Schmid, J.M.; Hoffmann, H.J.H. Increasing time interval and decreasing allergen dose interval improves ex vivo desensitization of human blood basophils. Cytom. Part B Clin. Cytom. 2016, 92, 340–347. [Google Scholar] [CrossRef]
- Oka, T.; Rios, E.J.; Tsai, M.; Kalesnikoff, J.; Galli, S.J. Rapid desensitization induces internalization of antigen-specific IgE on mouse mast cells. J. Allergy Clin. Immunol. 2013, 132, 922–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burches, E.; Ferriols, F.; Pérez-Fidalgo, J.-A.; González-Barrallo, I.; Cervantes, A. A logarithmic rapid desensitization protocol: Initial experience in carboplatin hypersensitivity reactions. J. Cancer Metastasis Treat. 2019, 5, 63. [Google Scholar] [CrossRef] [Green Version]
- Madrigal-Burgaleta, R.; Rubio, L.B.; Berges-Gimeno, M.P.; Carpio-Escalona, L.V.; Gehlhaar, P.; Alvarez-Cuesta, E. A Large Single-Hospital Experience Using Drug Provocation Testing and Rapid Drug Desensitization in Hypersensitivity to Antineoplastic and Biological Agents. J. Allergy Clin. Immunol. Pract. 2018, 7, 618–632. [Google Scholar] [CrossRef] [PubMed]
- Diaferio, L.; Giovannini, M.; Clark, E.; Castagnoli, R.; Caimmi, D. Protocols for drug allergy desensitization in children. Expert Rev. Clin. Immunol. 2019, 16, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Ishimaru, K.; Shibata, S.; Nakao, A. Regulation of plasma histamine levels by the mast cell clock and its modulation by stress. Sci. Rep. 2017, 7, 39934. [Google Scholar] [CrossRef] [Green Version]
- Barnes, P.; FitzGerald, G.; Brown, M.; Dollery, C. Nocturnal Asthma and Changes in Circulating Epinephrine, Histamine, and Cortisol. N. Engl. J. Med. 1980, 303, 263–267. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, H.J.; Kim, J.H.; Kim, T.H.; Lee, S.H. Asymmetric expression level of clock genes in left vs. right nasal mucosa in humans with and without allergies and in rats: Circadian characteristics and possible contribution to nasal cycle. PLoS ONE 2018, 13, e0194018. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-C.; Chen, Y.-C.; Wang, T.-N.; Fang, W.-F.; Chang, Y.-C.; Chen, Y.-M.; Chen, I.-Y.; Lin, M.-C.; Yang, M.-Y. Disrupted Expression of Circadian Clock Genes in Patients with Bronchial Asthma. J. Asthma Allergy 2021, ume 14, 371–380. [Google Scholar] [CrossRef]
- Krakowiak, K.; Downton, P.; Blaikley, J.; Durrington, H. L1 Blood immune cells clock mechanism is altered in asthma, with a time-of-day dependent response to steroids in vitro. BMJ 2021, 76, A230. [Google Scholar] [CrossRef]
- Yang, S.; Chew, H.; Jiang, Y.; Cheng, L.; Guo, X.; Che, H.; Sun, N. Lentinan Inhibited the Activation of Th2 Cells in Allergic Mice by Reducing the Amplitude of Changes in Biological Rhythm. Int. Arch. Allergy Immunol. 2020, 1–15. [Google Scholar] [CrossRef]
- Chastain, D.B.; Hutzley, V.J.; Parekh, J.; Alegro, J.V.G. Antimicrobial Desensitization: A Review of Published Protocols. Pharmacy 2019, 7, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakao, A. Circadian Regulation of the Biology of Allergic Disease: Clock Disruption Can Promote Allergy. Front. Immunol. 2020, 11, 1237. [Google Scholar] [CrossRef] [PubMed]
- Long, J.E.; Drayson, M.; Taylor, A.E.; Toellner, K.-M.; Lord, J.; Phillips, A.C. Morning vaccination enhances antibody response over afternoon vaccination: A cluster-randomised trial. Vaccine 2016, 34, 2679–2685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Liu, Y.; Liu, D.; Zeng, Q.; Li, L.; Zhou, Q.; Li, M.; Mei, J.; Yang, N.; Mo, S.; et al. Time of day influences immune response to an inactivated vaccine against SARS-CoV-2. Cell Res. 2021, 31, 1215–1217. [Google Scholar] [CrossRef]
- Reijers, J.A.A.; Moerland, M.; Burggraaf, J. Remarkable Pharmacokinetics of Monoclonal Antibodies: A Quest for an Explanation. Clin. Pharmacokinet. 2016, 56, 1081–1089. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Interval between Doses | DS Efficacy |
| 1–5 min | low [65] |
| 10–15 min | high [66,68] |
| 20 min | highest [70] |
| accelerated last step | high [74] |
| Number of doses | |
| single (suboptimal) | low [69] |
| multiple | high [69] |
| Duration of 1 DS cycle | |
| minutes (min. 30 min) | low [62] |
| hours (max. 4 h) | high [68] |
| Dosage | |
| geometric progression | high [70] |
| logarithmic progression (accelerated regimen) | high [72] |
| geometric progression, last dose much higher (accelerated regimen) | high [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stan, R.C. Temporal Modulation of Drug Desensitization Procedures. Curr. Issues Mol. Biol. 2022, 44, 833-844. https://doi.org/10.3390/cimb44020057
Stan RC. Temporal Modulation of Drug Desensitization Procedures. Current Issues in Molecular Biology. 2022; 44(2):833-844. https://doi.org/10.3390/cimb44020057
Chicago/Turabian StyleStan, Razvan Costin. 2022. "Temporal Modulation of Drug Desensitization Procedures" Current Issues in Molecular Biology 44, no. 2: 833-844. https://doi.org/10.3390/cimb44020057
APA StyleStan, R. C. (2022). Temporal Modulation of Drug Desensitization Procedures. Current Issues in Molecular Biology, 44(2), 833-844. https://doi.org/10.3390/cimb44020057