

Bamboo Shoot and Artemisia capillaris Extract Mixture Ameliorates Dextran Sodium Sulfate-Induced Colitis
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Line Culture
2.3. Animal Experimental Design
2.4. Induction of Colitis and BA Treatment
2.5. Clinical Score
2.6. Histological Analysis
2.7. Western Blot Analysis
2.8. Detection of NF-κB Phosphorylation in Cultured Cells
2.9. Measurement of Reactive Oxygen Species Generation
2.10. RhoA Activation Assay
2.11. Quantification of Filamentous-Actin (F-Actin) Formation
2.12. Statistical Analysis
3. Results
3.1. Effects of the Combined Ratio of BA Extracts on Body Weight Loss, Stool Constancy, Bloody Stool, and DAI Score
3.2. The Combined Ratio of BA Extracts Ameliorates Colon Shortening in the DSS-Induced Colitis Model
3.3. The BA Extracts Ameliorate Histological Score in the DSS-Induced Colitis Model
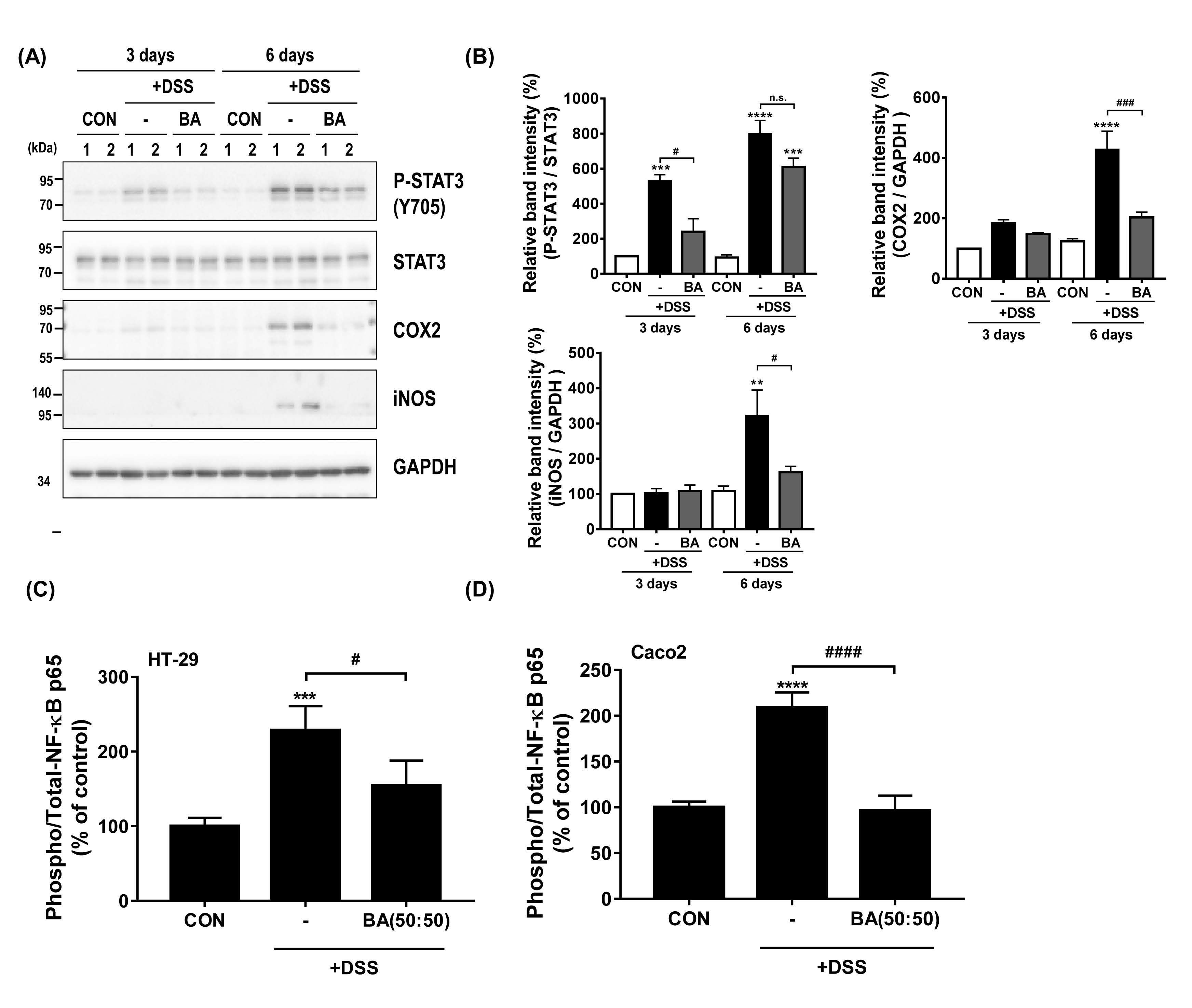
3.4. Treatment with BA Attenuated the Protein Expression of Pro-Inflammatory Mediators (STAT3, COX2, iNOS, and NF-κB) in DSS-Induced Colitis
3.5. Treatment with BA Reduces ROS Generation in DSS-Treated Human Colorectal Adenocarcinoma Cells
3.6. Treatment with BA Decreases F-Actin Formation and RhoA Activity in DSS-Treated Human Colorectal Adenocarcinoma Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaplan, G.G.; Windsor, J.W. The four epidemiological stages in the global evolution of inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Ronchetti, C.; Cirillo, F.; Di Segni, N.; Cristodoro, M.; Busnelli, A.; Levi-Setti, P.E. Inflammatory Bowel Disease and Reproductive Health: From Fertility to Pregnancy-A Narrative Review. Nutrients 2022, 14, 1591. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Inflammatory Bowel Disease (IBD). Available online: http://www.cdc.gov/ibd/#epidIBD (accessed on 6 August 2012).
- Yasmin, F.; Najeeb, H.; Shaikh, S.; Hasanain, M.; Naeem, U.; Moeed, A.; Koritala, T.; Hasan, S.; Surani, S. Novel drug delivery systems for inflammatory bowel disease. World J. Gastroenterol. 2022, 28, 1922–1933. [Google Scholar] [CrossRef]
- Cai, Z.; Wang, S.; Li, J. Treatment of Inflammatory Bowel Disease: A Comprehensive Review. Front. Med. 2021, 8, 765474. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.Y.; Kim, S.; Son, M.J.; Son, C.Y.; Kim, J.Y.; Kronbichler, A.; Lee, K.H.; Shin, J.I. Induction and maintenance treatment of inflammatory bowel disease: A comprehensive review. Autoimmun. Rev. 2019, 18, 439–454. [Google Scholar] [CrossRef] [PubMed]
- M’Koma, A.E. Inflammatory Bowel Disease: Clinical Diagnosis and Surgical Treatment-Overview. Medicina 2022, 58, 567. [Google Scholar] [CrossRef] [PubMed]
- Nongdam, P.; Tikendra, L. The Nutritional Facts of Bamboo Shoots and Their Usage as Important Traditional Foods of Northeast India. Int. Sch. Res. Not. 2014, 2014, 679073. [Google Scholar] [CrossRef] [PubMed]
- Singhal, P.; Bal, L.M.; Satya, S.; Sudhakar, P.; Naik, S.N. Bamboo shoots: A novel source of nutrition and medicine. Crit. Rev. Food Sci. Nutr. 2013, 53, 517–534. [Google Scholar] [CrossRef]
- Ren, Y.; Ma, Y.; Zhang, Z.; Qiu, L.; Zhai, H.; Gu, R.; Xie, Y. Total Alkaloids from Bamboo Shoots and Bamboo Shoot Shells of Pleioblastus amarus (Keng) Keng f. and Their Anti-Inflammatory Activities. Molecules 2019, 24, 2699. [Google Scholar] [CrossRef]
- Chen, C.C.; Lin, J.T.; Cheng, Y.F.; Kuo, C.Y.; Huang, C.F.; Kao, S.H.; Liang, Y.J.; Cheng, C.Y.; Chen, H.M. Amelioration of LPS-induced inflammation response in microglia by AMPK activation. BioMed Res. Int. 2014, 2014, 692061. [Google Scholar] [CrossRef]
- Li, Q.; Wu, W.; Fang, X.; Chen, H.; Han, Y.; Liu, R.; Niu, B.; Gao, H. Structural characterization of a polysaccharide from bamboo (Phyllostachys edulis) shoot and its prevention effect on colitis mouse. Food Chem. 2022, 387, 132807. [Google Scholar] [CrossRef] [PubMed]
- Jang, E.; Kim, B.J.; Lee, K.T.; Inn, K.S.; Lee, J.H. A Survey of Therapeutic Effects of Artemisia capillaris in Liver Diseases. Evid.-Based Complement. Altern. Med. eCAM 2015, 2015, 728137. [Google Scholar] [CrossRef]
- Bora, K.S.; Sharma, A. The genus Artemisia: A comprehensive review. Pharm. Biol. 2011, 49, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The artemisia L. Genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef]
- Hsueh, T.P.; Lin, W.L.; Dalley, J.W.; Tsai, T.H. The Pharmacological Effects and Pharmacokinetics of Active Compounds of Artemisia capillaris. Biomedicines 2021, 9, 1412. [Google Scholar] [CrossRef]
- Park, J.M.; Hahm, K.B.; Kwon, S.O.; Kim, E.H. The Anti-inflammatory Effects of Acidic Polysaccharide from Artemisia capillaris on Helicobacter pylori Infection. J. Cancer Prev. 2013, 18, 161–168. [Google Scholar] [CrossRef]
- Hall, A. Rho GTPases and the actin cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef]
- Aspenström, P. The Rho GTPases have multiple effects on the actin cytoskeleton. Exp. Cell Res. 1999, 246, 20–25. [Google Scholar] [CrossRef]
- Kim, J.G.; Islam, R.; Cho, J.Y.; Jeong, H.; Cap, K.C.; Park, Y.; Hossain, A.J.; Park, J.B. Regulation of RhoA GTPase and various transcription factors in the RhoA pathway. J. Cell. Physiol. 2018, 233, 6381–6392. [Google Scholar] [CrossRef]
- Johnson, L.A.; Rodansky, E.S.; Haak, A.J.; Larsen, S.D.; Neubig, R.R.; Higgins, P.D. Novel Rho/MRTF/SRF inhibitors block matrix-stiffness and TGF-β-induced fibrogenesis in human colonic myofibroblasts. Inflamm. Bowel Dis. 2014, 20, 154–165. [Google Scholar] [CrossRef]
- Mihaescu, A.; Santén, S.; Jeppsson, B.; Thorlacius, H. Rho kinase signalling mediates radiation-induced inflammation and intestinal barrier dysfunction. Br. J. Surg. 2011, 98, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Ma, L.; Zhao, Y.; Zhang, S.; Zhou, C.; Cai, Y. Inhibition of Rho kinase protects against colitis in mice by attenuating intestinal epithelial barrier dysfunction via MLC and the NF-κB pathway. Int. J. Mol. Med. 2018, 41, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Duan, X.; Liu, X.; Liu, Y.; Fan, H.; Xu, M.; Chen, Q.; Tang, Q. Rho kinase Blockade Ameliorates DSS-Induced Ulcerative Colitis in Mice Through Dual Inhibition of the NF-κB and IL-6/STAT3 Pathways. Inflammation 2020, 43, 857–867. [Google Scholar] [CrossRef]
- Huang, Y.; Xiao, S.; Jiang, Q. Role of Rho kinase signal pathway in inflammatory bowel disease. Int. J. Clin. Exp. Med. 2015, 8, 3089–3097. [Google Scholar]
- Wirtz, S.; Neufert, C.; Weigmann, B.; Neurath, M.F. Chemically induced mouse models of intestinal inflammation. Nat. Protoc. 2007, 2, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Friedman, D.J.; Künzli, B.M.; YI, A.R.; Sevigny, J.; Berberat, P.O.; Enjyoji, K.; Csizmadia, E.; Friess, H.; Robson, S.C. From the Cover: CD39 deletion exacerbates experimental murine colitis and human polymorphisms increase susceptibility to inflammatory bowel disease. Proc. Natl. Acad. Sci. USA 2009, 106, 16788–16793. [Google Scholar] [CrossRef] [PubMed]
- Le Naour, J.; Montégut, L.; Joseph, A.; Garbin, K.; Vacchelli, E.; Kroemer, G.; Pol, J.G.; Maiuri, M.C. Improved Swiss-rolling method for histological analyses of colon tissue. MethodsX 2022, 9, 101630. [Google Scholar] [CrossRef]
- Bauer, C.; Duewell, P.; Mayer, C.; Lehr, H.A.; Fitzgerald, K.A.; Dauer, M.; Tschopp, J.; Endres, S.; Latz, E.; Schnurr, M. Colitis induced in mice with dextran sulfate sodium (DSS) is mediated by the NLRP3 inflammasome. Gut 2010, 59, 1192–1199. [Google Scholar] [CrossRef]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef]
- Hendrickson, B.A.; Gokhale, R.; Cho, J.H. Clinical aspects and pathophysiology of inflammatory bowel disease. Clin. Microbiol. Rev. 2002, 15, 79–94. [Google Scholar] [CrossRef]
- Jin, B.R.; Chung, K.S.; Cheon, S.Y.; Lee, M.; Hwang, S.; Noh Hwang, S.; Rhee, K.J.; An, H.J. Rosmarinic acid suppresses colonic inflammation in dextran sulphate sodium (DSS)-induced mice via dual inhibition of NF-κB and STAT3 activation. Sci. Rep. 2017, 7, 46252. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.W.; Shin, J.S.; Chung, K.S.; Lee, Y.G.; Baek, N.I.; Lee, K.T. Anti-Inflammatory Mechanisms of Koreanaside A, a Lignan Isolated from the Flower of Forsythia koreana, against LPS-Induced Macrophage Activation and DSS-Induced Colitis Mice: The Crucial Role of AP-1, NF-κB, and JAK/STAT Signaling. Cells 2019, 8, 1163. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yu, T.; Jang, H.J.; Byeon, S.E.; Song, S.Y.; Lee, B.H.; Rhee, M.H.; Kim, T.W.; Lee, J.; Hong, S.; et al. In vitro and in vivo anti-inflammatory activities of Polygonum hydropiper methanol extract. J. Ethnopharmacol. 2012, 139, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Yu, Y.; Li, Z.; Cao, G.; Huang, S.; Yang, H. Bamboo Leaf Flavonoids Extracts Alleviate Oxidative Stress in HepG2 Cells via Naturally Modulating Reactive Oxygen Species Production and Nrf2-Mediated Antioxidant Defense Responses. J. Food Sci. 2019, 84, 1609–1620. [Google Scholar] [CrossRef]
- Luo, G.Y.; Luo, Y.G.; Zhou, R.; Zhou, M.; Gu, J.; Ye, Q.; Dai, Y.; Zhang, G.L. Antioxidant compounds from ethanol extracts of bamboo (Neosinocalamus affinis) leaves. J. Asian Nat. Prod. Res. 2015, 17, 248–255. [Google Scholar] [CrossRef]
- Yeo, D.; Hwang, S.J.; Kim, W.J.; Youn, H.J.; Lee, H.J. The aqueous extract from Artemisia capillaris inhibits acute gastric mucosal injury by inhibition of ROS and NF-kB. Biomed. Pharmacother. 2018, 99, 681–687. [Google Scholar] [CrossRef]
- Pradhan, R.; Ngo, P.A.; Martínez-Sánchez, L.D.; Neurath, M.F.; López-Posadas, R. Rho GTPases as Key Molecular Players within Intestinal Mucosa and GI Diseases. Cells 2021, 10, 66. [Google Scholar] [CrossRef]
- Spiering, D.; Hodgson, L. Dynamics of the Rho-family small GTPases in actin regulation and motility. Cell Adhes. Migr. 2011, 5, 170–180. [Google Scholar] [CrossRef]
- Tkach, V.; Bock, E.; Berezin, V. The role of RhoA in the regulation of cell morphology and motility. Cell Motil. Cytoskelet. 2005, 61, 21–33. [Google Scholar] [CrossRef]
- Grivennikov, S.I.; Karin, M. Dangerous liaisons: STAT3 and NF-kappaB collaboration and crosstalk in cancer. Cytokine Growth Factor Rev. 2010, 21, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.D.; Zhao, Y.H. Targeting NF-κB pathway for treating ulcerative colitis: Comprehensive regulatory characteristics of Chinese medicines. Chin. Med. 2020, 15, 15. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Akanda, M.R.; Nam, H.H.; Tian, W.; Islam, A.; Choo, B.K.; Park, B.Y. Regulation of JAK2/STAT3 and NF-κB signal transduction pathways; Veronica polita alleviates dextran sulfate sodium-induced murine colitis. Biomed. Pharmacother. 2018, 100, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Danese, S.; Mantovani, A. Inflammatory bowel disease and intestinal cancer: A paradigm of the Yin-Yang interplay between inflammation and cancer. Oncogene 2010, 29, 3313–3323. [Google Scholar] [CrossRef]
- Grothaus, J.S.; Ares, G.; Yuan, C.; Wood, D.R.; Hunter, C.J. Rho kinase inhibition maintains intestinal and vascular barrier function by upregulation of occludin in experimental necrotizing enterocolitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G514–G528. [Google Scholar] [CrossRef]
- Citalán-Madrid, A.F.; Vargas-Robles, H.; García-Ponce, A.; Shibayama, M.; Betanzos, A.; Nava, P.; Salinas-Lara, C.; Rottner, K.; Mennigen, R.; Schnoor, M. Cortactin deficiency causes increased RhoA/ROCK1-dependent actomyosin contractility, intestinal epithelial barrier dysfunction, and disproportionately severe DSS-induced colitis. Mucosal Immunol. 2017, 10, 1237–1247. [Google Scholar] [CrossRef]
- Tian, Y.M.; Tian, S.Y.; Wang, D.; Cui, F.; Zhang, X.J.; Zhang, Y. Elevated expression of the leptin receptor ob-R may contribute to inflammation in patients with ulcerative colitis. Mol. Med. Rep. 2019, 20, 4706–4712. [Google Scholar] [CrossRef]
- Chen, W.; Nyuydzefe, M.S.; Weiss, J.M.; Zhang, J.; Waksal, S.D.; Zanin-Zhorov, A. ROCK2, but not ROCK1 interacts with phosphorylated STAT3 and co-occupies TH17/TFH gene promoters in TH17-activated human T cells. Sci. Rep. 2018, 8, 16636. [Google Scholar] [CrossRef]
- Ivanov, A.I.; Parkos, C.A.; Nusrat, A. Cytoskeletal regulation of epithelial barrier function during inflammation. Am. J. Pathol. 2010, 177, 512–524. [Google Scholar] [CrossRef]
- Hotulainen, P.; Paunola, E.; Vartiainen, M.K.; Lappalainen, P. Actin-depolymerizing factor and cofilin-1 play overlapping roles in promoting rapid F-actin depolymerization in mammalian nonmuscle cells. Mol. Biol. Cell 2005, 16, 649–664. [Google Scholar] [CrossRef]
- Lin, T.; Zeng, L.; Liu, Y.; DeFea, K.; Schwartz, M.A.; Chien, S.; Shyy, J.Y. Rho-ROCK-LIMK-cofilin pathway regulates shear stress activation of sterol regulatory element binding proteins. Circ. Res. 2003, 92, 1296–1304. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Luan, Y.; Wang, T.; Zhuan, L.; Rao, K.; Wang, S.G.; Ye, Z.Q.; Liu, J.H.; Wang, D.W. Reduced corporal fibrosis to protect erectile function by inhibiting the Rho-kinase/LIM-kinase/cofilin pathway in the aged transgenic rat harboring human tissue kallikrein 1. Asian J. Androl. 2017, 19, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.M.; Lin, S.Z.; Chang, N.C. Membrane ERα attenuates myocardial fibrosis via RhoA/ROCK-mediated actin remodeling in ovariectomized female infarcted rats. J. Mol. Med. 2014, 92, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Moon, M.Y.; Kim, H.J.; Kim, M.J.; Uhm, S.; Park, J.W.; Suk, K.T.; Park, J.B.; Kim, D.J.; Kim, S.E. Rap1 regulates hepatic stellate cell migration through the modulation of RhoA activity in response to TGF-β1. Int. J. Mol. Med. 2019, 44, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Lai, D.; Gao, J.; Bi, X.; He, H.; Shi, X.; Weng, S.; Chen, Y.; Yang, Y.; Ye, Y.; Fu, G. The Rho kinase inhibitor, fasudil, ameliorates diabetes-induced cardiac dysfunction by improving calcium clearance and actin remodeling. J. Mol. Med. 2017, 95, 155–165. [Google Scholar] [CrossRef]
- Aviello, G.; Knaus, U.G. ROS in gastrointestinal inflammation: Rescue Or Sabotage? Br. J. Pharmacol. 2017, 174, 1704–1718. [Google Scholar] [CrossRef]
- Tian, T.; Wang, Z.; Zhang, J. Pathomechanisms of Oxidative Stress in Inflammatory Bowel Disease and Potential Antioxidant Therapies. Oxidative Med. Cell. Longev. 2017, 2017, 4535194. [Google Scholar] [CrossRef]
- Chiurchiù, V.; Maccarrone, M. Chronic inflammatory disorders and their redox control: From molecular mechanisms to therapeutic opportunities. Antioxid. Redox Signal. 2011, 15, 2605–2641. [Google Scholar] [CrossRef]
- Peng, Y.C.; Hsu, C.L.; Tung, C.F.; Chou, W.K.; Huang, L.R.; Hung, D.Z.; Hu, W.H.; Yang, D.Y. Chemiluminescence assay of mucosal reactive oxygen species in gastric cancer, ulcer and antral mucosa. Hepato-Gastroenterol. 2008, 55, 770–773. [Google Scholar]
- Grisham, M.B. Oxidants and free radicals in inflammatory bowel disease. Lancet 1994, 344, 859–861. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disease Activity Index (DAI) | |||
|---|---|---|---|
| Score | Weight Loss | Stool Consistency | Bleeding |
| 0 | None | Normal | Normal |
| 1 | 0–10% | Soft but still formed | |
| 2 | 10–15% | Loose stools | Hemocult+ |
| 3 | 15–20% | ||
| 4 | >20% | Diarrhea | Gross Bleeding |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-J.; Kim, B.; Lee, M.-R.; Ra, M.; Lee, Y. Bamboo Shoot and Artemisia capillaris Extract Mixture Ameliorates Dextran Sodium Sulfate-Induced Colitis. Curr. Issues Mol. Biol. 2022, 44, 5086-5103. https://doi.org/10.3390/cimb44100345
Kim H-J, Kim B, Lee M-R, Ra M, Lee Y. Bamboo Shoot and Artemisia capillaris Extract Mixture Ameliorates Dextran Sodium Sulfate-Induced Colitis. Current Issues in Molecular Biology. 2022; 44(10):5086-5103. https://doi.org/10.3390/cimb44100345
Chicago/Turabian StyleKim, Hee-Jun, Bohye Kim, Mi-Ra Lee, Moonjin Ra, and Yongjun Lee. 2022. "Bamboo Shoot and Artemisia capillaris Extract Mixture Ameliorates Dextran Sodium Sulfate-Induced Colitis" Current Issues in Molecular Biology 44, no. 10: 5086-5103. https://doi.org/10.3390/cimb44100345
APA StyleKim, H.-J., Kim, B., Lee, M.-R., Ra, M., & Lee, Y. (2022). Bamboo Shoot and Artemisia capillaris Extract Mixture Ameliorates Dextran Sodium Sulfate-Induced Colitis. Current Issues in Molecular Biology, 44(10), 5086-5103. https://doi.org/10.3390/cimb44100345