
Dexpanthenol Promotes Cell Growth by Preventing Cell Senescence and Apoptosis in Cultured Human Hair Follicle Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Culture of Human Dermal Papilla Cells (hDPCs) and Outer Root Sheath Cells (hORSCs)
2.2. Cell Viability Assay
2.3. mRNA Analysis
2.4. Western Blot Analysis
2.5. Immunocytochemistry
2.6. Alkaline Phosphatase (ALP) Staining and Quantification
2.7. Statistical Analysis
3. Results
3.1. D-Panthenol Stimulated the Growth of Cultured hDPCs
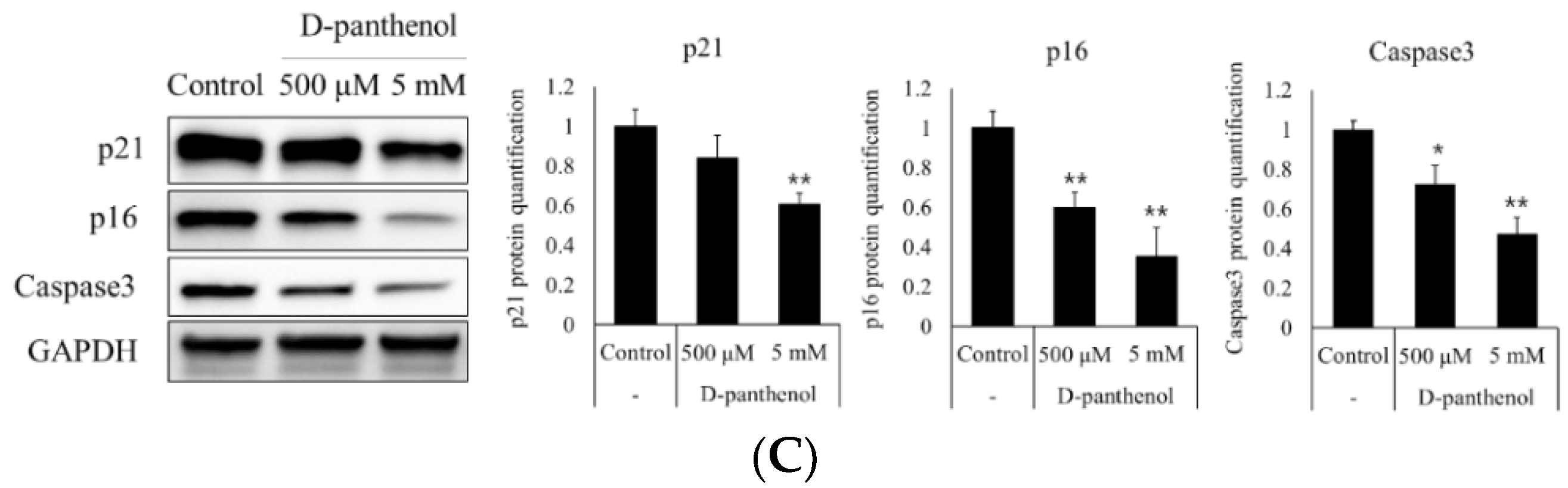
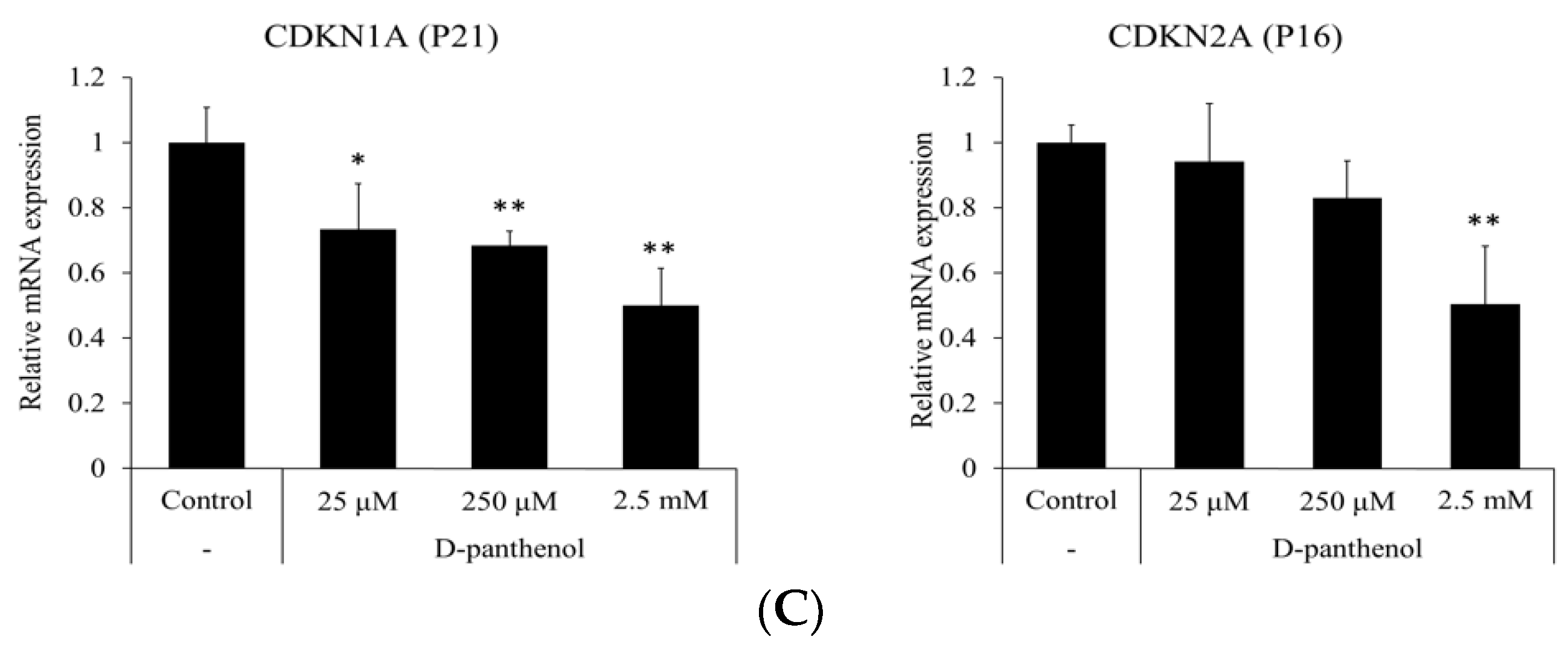
3.2. D-Panthenol Reduced Apoptotic and Senescence Markers in Cultured hDPCs
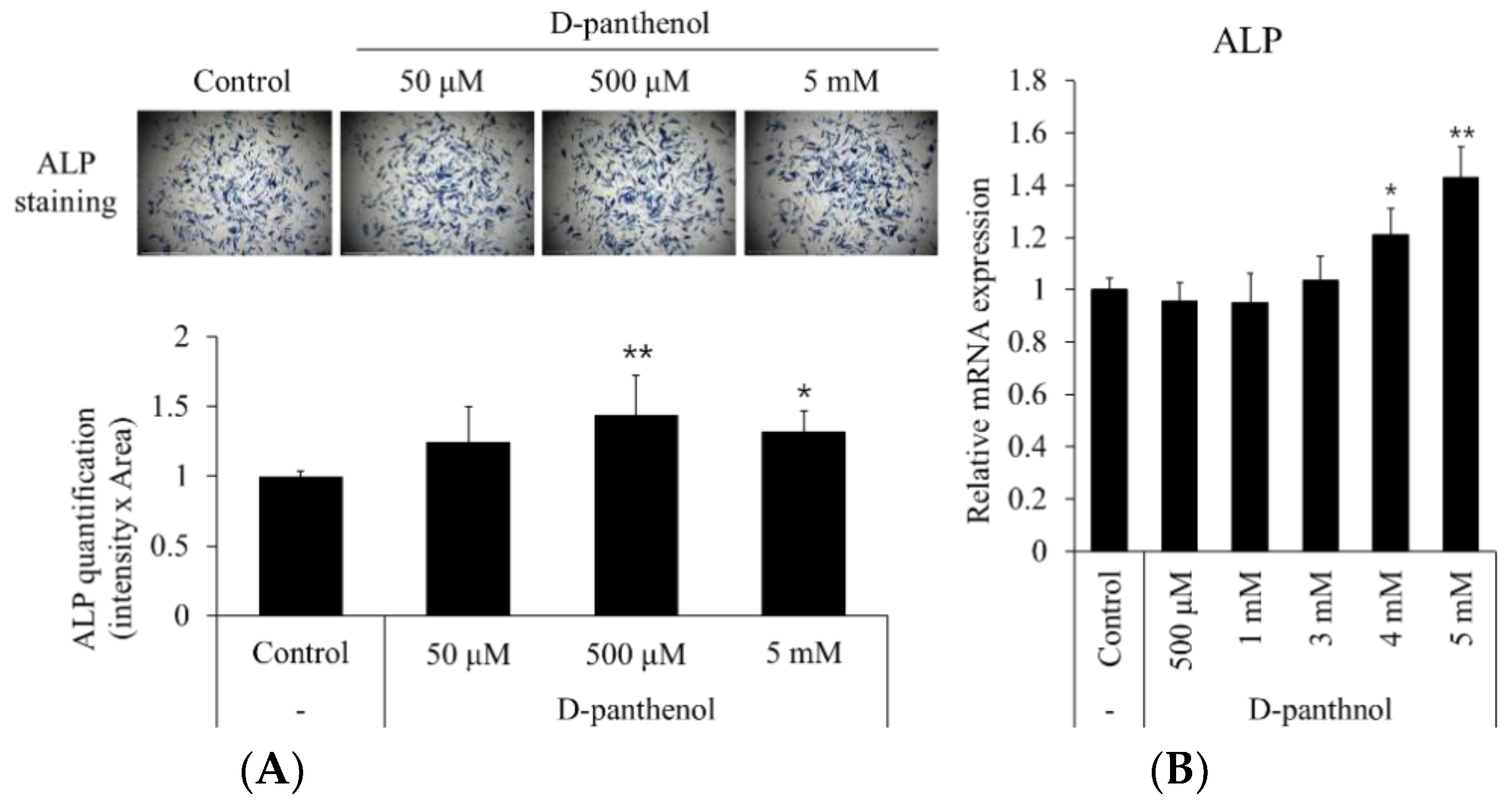
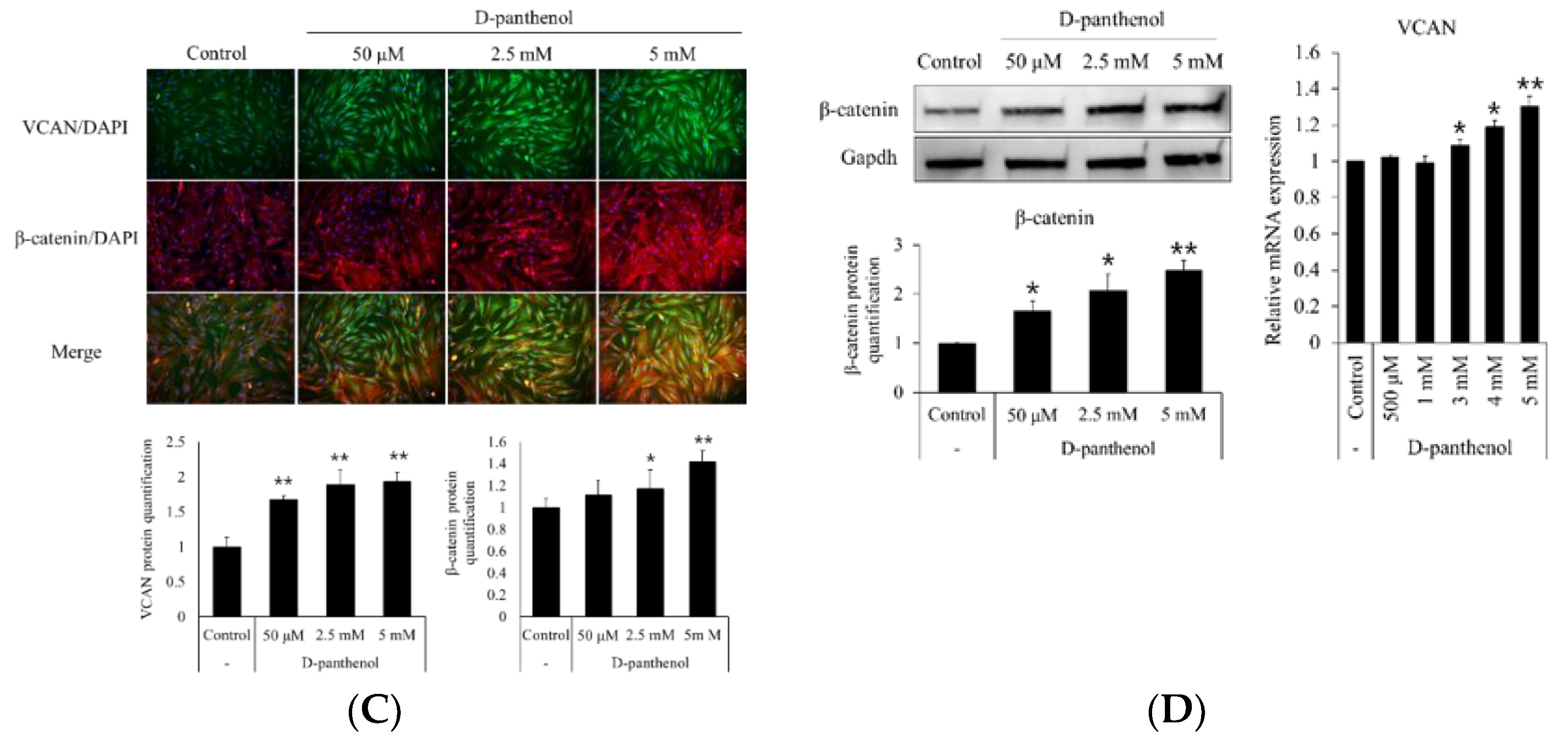
3.3. D-Panthenol Significantly Increased the Anagen Markers in Cultured hDPCs
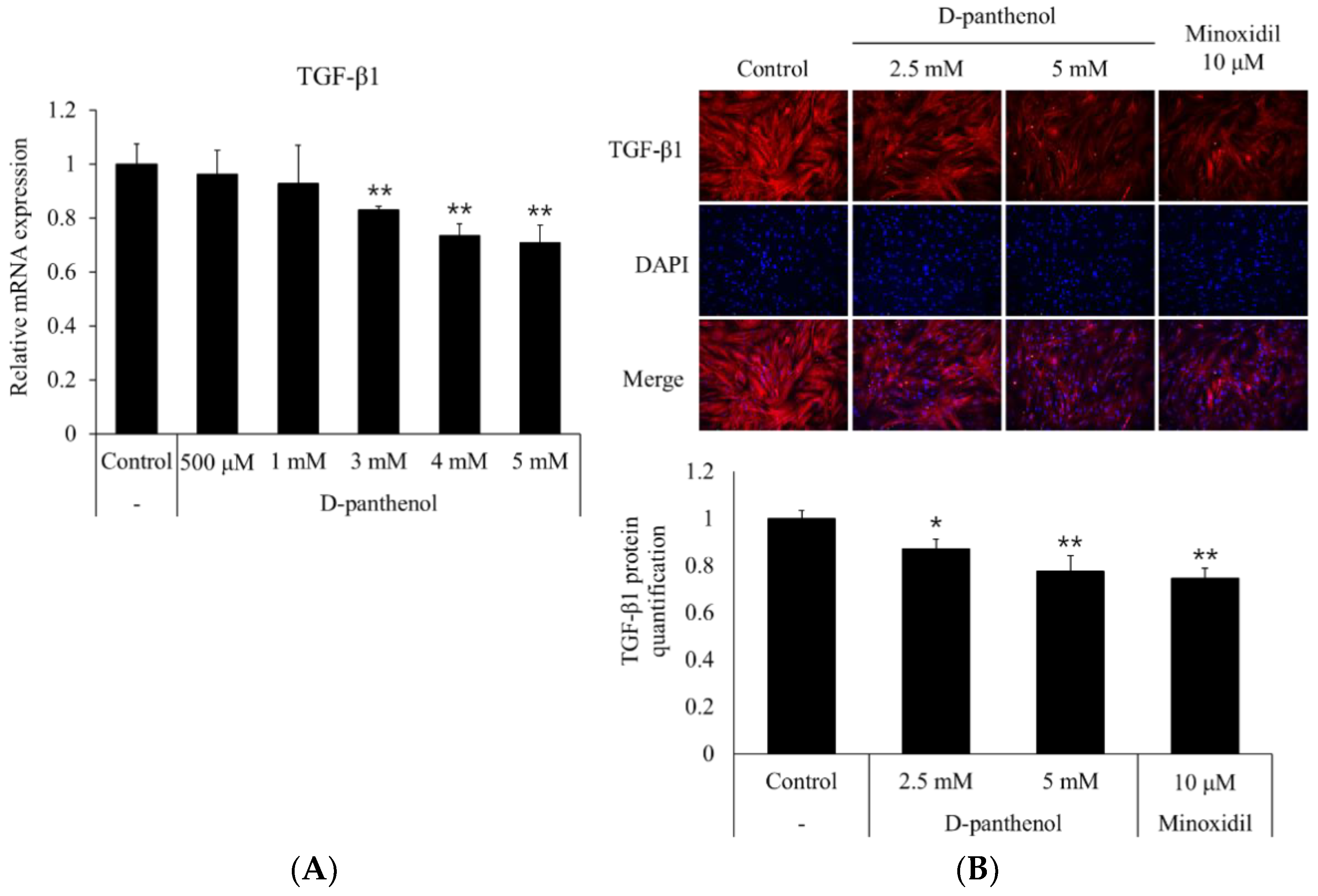
3.4. D-Panthenol Reduced TGF-β1 Expression in Cultured hDPCs
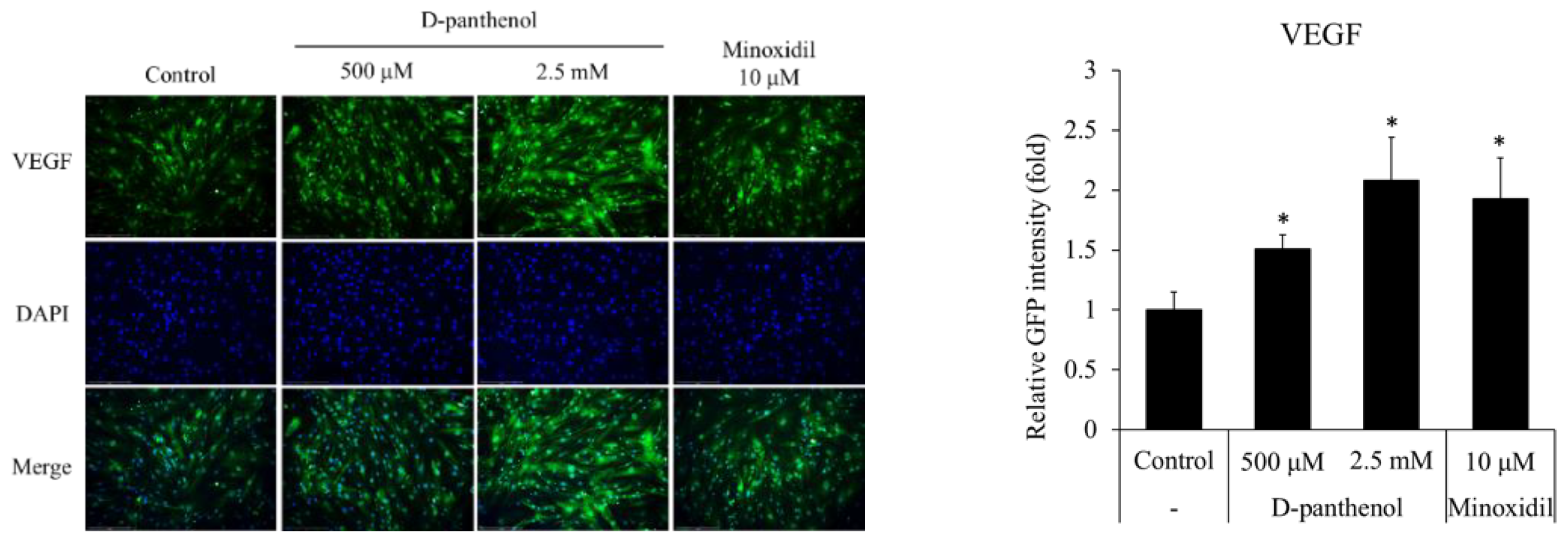
3.5. D-Panthenol Stimulated VEGF Expression in Cultured hDPCs
3.6. D-Panthenol Stimulated the Growth of Cultured hORSCs and Reduced the Senescence Markers
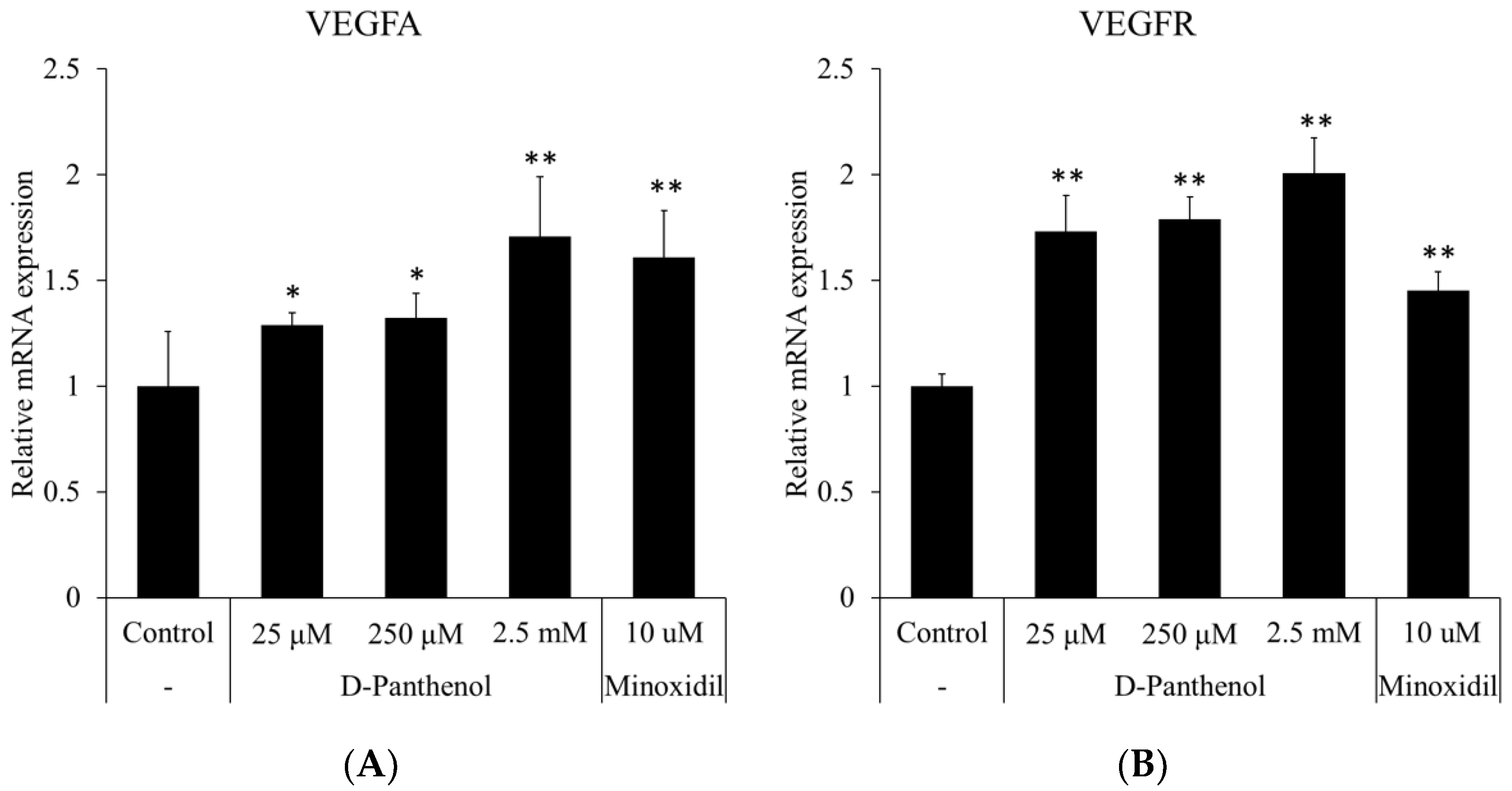
3.7. D-Panthenol Induced the Expression of VEGF and VEGFR in Cultured hORSCs
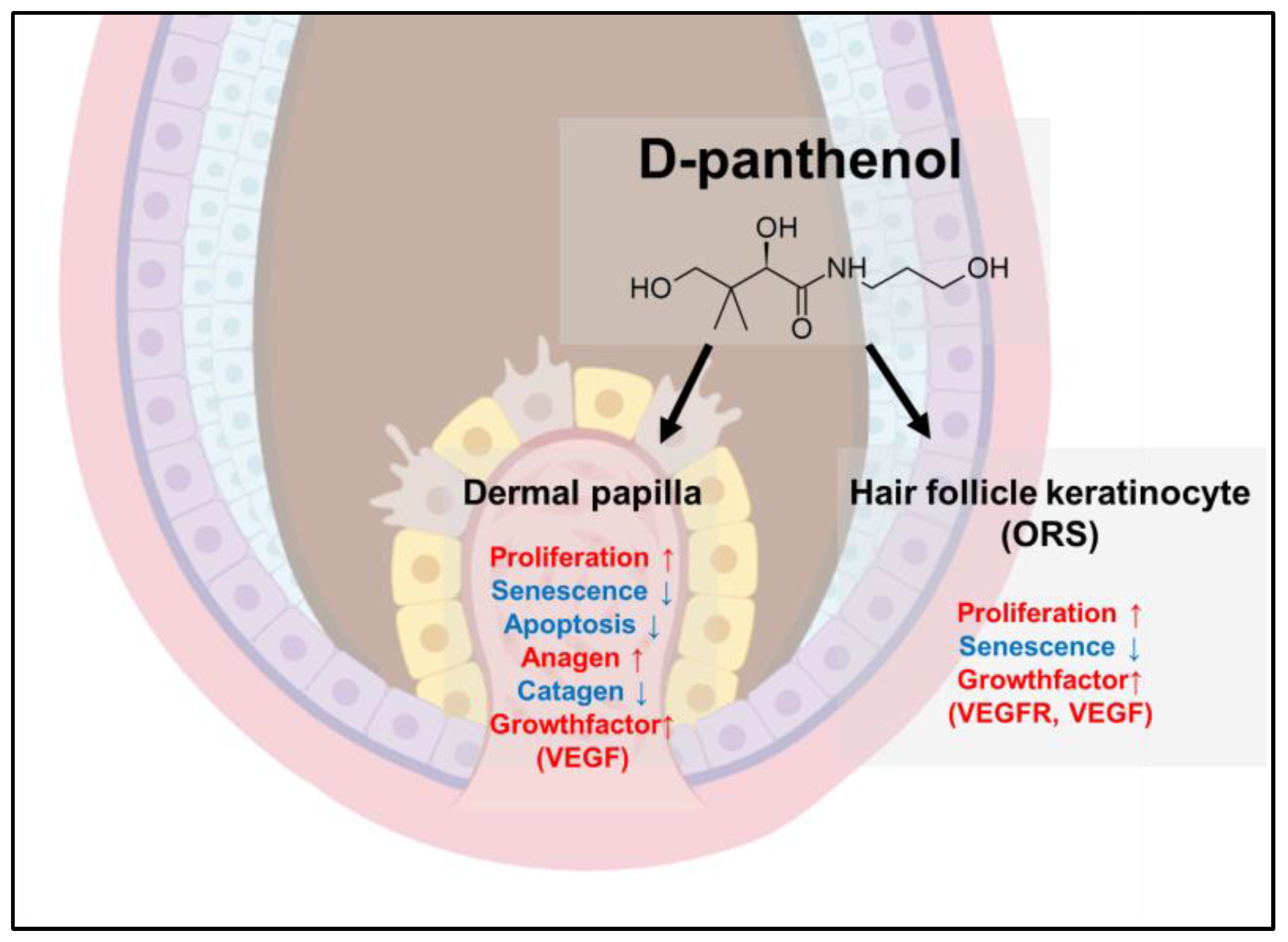
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Schneider, M.R.; Schmidt-Ullrich, R.; Paus, R. The hair follicle as a dynamic miniorgan. Curr. Biol. 2009, 19, R132–R142. [Google Scholar] [CrossRef]
- Whiting, D.A. Possible mechanisms of miniaturization during androgenetic alopecia or pattern hair loss. J. Am. Acad. Dermatol. 2001, 45, S81–S86. [Google Scholar] [CrossRef]
- Giacomini, F.; Starace, M.; Tosti, A. Short anagen syndrome. Pediatr. Dermatol. 2011, 28, 133–134. [Google Scholar] [CrossRef]
- Kim, J.; Shin, J.Y.; Choi, Y.H.; Jang, M.; Nam, Y.J.; Lee, S.Y.; Jeon, J.; Jin, M.H.; Lee, S. Hair Growth Promoting Effect of Hottuynia cordata Extract in Cultured Human Hair Follicle Dermal Papilla Cells. Biol. Pharm. Bull. 2019, 42, 1665–1673. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Yoon, J.; Shin, S.H.; Zahoor, M.; Kim, H.J.; Park, P.J.; Park, W.S.; Min do, S.; Kim, H.Y.; Choi, K.Y. Valproic acid induces hair regeneration in murine model and activates alkaline phosphatase activity in human dermal papilla cells. PLoS ONE 2012, 7, e34152. [Google Scholar] [CrossRef] [PubMed]
- Itami, S.; Kurata, S.; Takayasu, S. Androgen induction of follicular epithelial cell growth is mediated via insulin-like growth factor-I from dermal papilla cells. Biochem. Biophys. Res. Commun. 1995, 212, 988–994. [Google Scholar] [CrossRef] [PubMed]
- Lachgar, S.; Charveron, M.; Gall, Y.; Bonafe, J.L. Minoxidil upregulates the expression of vascular endothelial growth factor in human hair dermal papilla cells. Br. J. Dermatol. 1998, 138, 407–411. [Google Scholar] [CrossRef]
- Li, W.; Man, X.Y.; Li, C.M.; Chen, J.Q.; Zhou, J.; Cai, S.Q.; Lu, Z.F.; Zheng, M. VEGF induces proliferation of human hair follicle dermal papilla cells through VEGFR-2-mediated activation of ERK. Exp. Cell Res. 2012, 318, 1633–1640. [Google Scholar] [CrossRef]
- Inoue, K.; Aoi, N.; Yamauchi, Y.; Sato, T.; Suga, H.; Eto, H.; Kato, H.; Tabata, Y.; Yoshimura, K. TGF-beta is specifically expressed in human dermal papilla cells and modulates hair folliculogenesis. J. Cell. Mol. Med. 2009, 13, 4643–4656. [Google Scholar] [CrossRef]
- Shin, H.; Yoo, H.G.; Inui, S.; Itami, S.; Kim, I.G.; Cho, A.R.; Lee, D.H.; Park, W.S.; Kwon, O.; Cho, K.H.; et al. Induction of transforming growth factor-beta 1 by androgen is mediated by reactive oxygen species in hair follicle dermal papilla cells. BMB Rep. 2013, 46, 460–464. [Google Scholar] [CrossRef]
- Kwack, M.H.; Sung, Y.K.; Chung, E.J.; Im, S.U.; Ahn, J.S.; Kim, M.K.; Kim, J.C. Dihydrotestosterone-inducible dickkopf 1 from balding dermal papilla cells causes apoptosis in follicular keratinocytes. J. Invest. Dermatol. 2008, 128, 262–269. [Google Scholar] [CrossRef] [PubMed]
- Vidal, V.P.; Chaboissier, M.C.; Lutzkendorf, S.; Cotsarelis, G.; Mill, P.; Hui, C.C.; Ortonne, N.; Ortonne, J.P.; Schedl, A. Sox9 is essential for outer root sheath differentiation and the formation of the hair stem cell compartment. Curr. Biol. 2005, 15, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Rompolas, P.; Deschene, E.R.; Zito, G.; Gonzalez, D.G.; Saotome, I.; Haberman, A.M.; Greco, V. Live imaging of stem cell and progeny behaviour in physiological hair-follicle regeneration. Nature 2012, 487, 496–499. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Nan, W.; Wang, S.; Zhang, T.; Si, H.; Yang, F.; Li, G. Epidermal Growth Factor Promotes Proliferation and Migration of Follicular Outer Root Sheath Cells via Wnt/beta-Catenin Signaling. Cell Physiol. Biochem. 2016, 39, 360–370. [Google Scholar] [CrossRef] [PubMed]
- Biro, K.; Thaci, D.; Ochsendorf, F.R.; Kaufmann, R.; Boehncke, W.H. Efficacy of dexpanthenol in skin protection against irritation: A double-blind, placebo-controlled study. Contact Dermat. 2003, 49, 80–84. [Google Scholar] [CrossRef]
- Gehring, W.; Gloor, M. Effect of topically applied dexpanthenol on epidermal barrier function and stratum corneum hydration. Results of a human in vivo study. Arzneimittelforschung 2000, 50, 659–663. [Google Scholar] [CrossRef]
- Stettler, H.; Kurka, P.; Wagner, C.; Sznurkowska, K.; Czernicka, O.; Bohling, A.; Bielfeldt, S.; Wilhelm, K.P.; Lenz, H. A new topical panthenol-containing emollient: Skin-moisturizing effect following single and prolonged usage in healthy adults, and tolerability in healthy infants. J. Dermatolog. Treat 2017, 28, 251–257. [Google Scholar] [CrossRef]
- Stamatas, G.N.; Tierney, N.K. Diaper dermatitis: Etiology, manifestations, prevention, and management. Pediatr. Dermatol. 2014, 31, 1–7. [Google Scholar] [CrossRef]
- Adam, R. Skin care of the diaper area. Pediatr. Dermatol. 2008, 25, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Heise, R.; Skazik, C.; Marquardt, Y.; Czaja, K.; Sebastian, K.; Kurschat, P.; Gan, L.; Denecke, B.; Ekanayake-Bohlig, S.; Wilhelm, K.P.; et al. Dexpanthenol modulates gene expression in skin wound healing in vivo. Skin Pharmacol. Physiol. 2012, 25, 241–248. [Google Scholar] [CrossRef]
- Marquardt, Y.; Amann, P.M.; Heise, R.; Czaja, K.; Steiner, T.; Merk, H.F.; Skazik-Voogt, C.; Baron, J.M. Characterization of a novel standardized human three-dimensional skin wound healing model using non-sequential fractional ultrapulsed CO2 laser treatments. Lasers Surg. Med. 2015, 47, 257–265. [Google Scholar] [CrossRef]
- Kutlu, O.; Metin, A. Systemic dexpanthenol as a novel treatment for female pattern hair loss. J. Cosmet Dermatol. 2021, 20, 1325–1330. [Google Scholar] [CrossRef]
- Kutlu, O. Dexpanthenol may be a novel treatment for male androgenetic alopecia: Analysis of nine cases. Dermatol. Ther. 2020, 33, e13381. [Google Scholar] [CrossRef]
- Wojtczak, L.; Slyshenkov, V.S. Protection by pantothenic acid against apoptosis and cell damage by oxygen free radicals—The role of glutathione. Biofactors 2003, 17, 61–73. [Google Scholar] [CrossRef]
- Kim, M.H.; Kim, S.H.; Yang, W.M. Beneficial effects of Astragaloside IV for hair loss via inhibition of Fas/Fas L-mediated apoptotic signaling. PLoS ONE 2014, 9, e92984. [Google Scholar] [CrossRef]
- Fujita, E.; Egashira, J.; Urase, K.; Kuida, K.; Momoi, T. Caspase-9 processing by caspase-3 via a feedback amplification loop in vivo. Cell Death Differ. 2001, 8, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Iida, M.; Ihara, S.; Matsuzaki, T. Hair cycle-dependent changes of alkaline phosphatase activity in the mesenchyme and epithelium in mouse vibrissal follicles. Dev. Growth Differ. 2007, 49, 185–195. [Google Scholar] [CrossRef]
- Jin, Y.H.; Kim, H.; Ki, H.; Yang, I.; Yang, N.; Lee, K.Y.; Kim, N.; Park, H.S.; Kim, K. Beta-catenin modulates the level and transcriptional activity of Notch1/NICD through its direct interaction. Biochim. Biophys. Acta 2009, 1793, 290–299. [Google Scholar] [CrossRef]
- Handjiski, B.K.; Eichmuller, S.; Hofmann, U.; Czarnetzki, B.M.; Paus, R. Alkaline phosphatase activity and localization during the murine hair cycle. Br. J. Dermatol. 1994, 131, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Foitzik, K.; Lindner, G.; Mueller-Roever, S.; Maurer, M.; Botchkareva, N.; Botchkarev, V.; Handjiski, B.; Metz, M.; Hibino, T.; Soma, T.; et al. Control of murine hair follicle regression (catagen) by TGF-beta1 in vivo. FASEB J. 2000, 14, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Hodgins, M.B.; Choudhry, R.; Parker, G.; Oliver, R.F.; Jahoda, C.A.; Withers, A.P.; Brinkmann, A.O.; van der Kwast, T.H.; Boersma, W.J.; Lammers, K.M.; et al. Androgen receptors in dermal papilla cells of scalp hair follicles in male pattern baldness. Ann. N. Y. Acad. Sci. 1991, 642, 448–451. [Google Scholar] [CrossRef]
- Yano, K.; Brown, L.F.; Detmar, M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J. Clin. Invest. 2001, 107, 409–417. [Google Scholar] [CrossRef]
- Park, P.J.; Moon, B.S.; Lee, S.H.; Kim, S.N.; Kim, A.R.; Kim, H.J.; Park, W.S.; Choi, K.Y.; Cho, E.G.; Lee, T.R. Hair growth-promoting effect of Aconiti Ciliare Tuber extract mediated by the activation of Wnt/beta-catenin signaling. Life Sci. 2012, 91, 935–943. [Google Scholar] [CrossRef]
- Wu, X.J.; Jing, J.; Lu, Z.F.; Zheng, M. VEGFR-2 Is in a State of Activation in Hair Follicles, Sebaceous Glands, Eccrine Sweat Glands, and Epidermis from Human Scalp: An In Situ Immunohistochemistry Study of Phosphorylated VEGFR-2. Med. Sci. Monit. Basic Res. 2019, 25, 107–112. [Google Scholar] [CrossRef]
- Tanaka, T.; Narisawa, Y.; Misago, N.; Hashimoto, K. The innermost cells of the outer root sheath in human anagen hair follicles undergo specialized keratinization mediated by apoptosis. J. Cutan Pathol. 1998, 25, 316–321. [Google Scholar] [CrossRef]
- Li, H.; Masieri, F.F.; Schneider, M.; Bartella, A.; Gaus, S.; Hahnel, S.; Zimmerer, R.; Sack, U.; Maksimovic-Ivanic, D.; Mijatovic, S.; et al. The Middle Part of the Plucked Hair Follicle Outer Root Sheath Is Identified as an Area Rich in Lineage-Specific Stem Cell Markers. Biomolecules 2021, 11, 154. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Sohn, K.C.; Kim, S.Y.; Ryu, E.K.; Park, Y.S.; Lee, Y.; Seo, Y.J.; Lee, J.H.; Kim, C.D. Sox9 Increases the Proliferation and Colony-forming Activity of Outer Root Sheath Cells Cultured In Vitro. Ann. Dermatol. 2011, 23, 138–143. [Google Scholar] [CrossRef][Green Version]
- Li, J.; Yang, Z.; Li, Z.; Gu, L.; Wang, Y.; Sung, C. Exogenous IGF-1 promotes hair growth by stimulating cell proliferation and down regulating TGF-beta1 in C57BL/6 mice in vivo. Growth Horm. IGF Res. 2014, 24, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Namekata, M.; Yamamoto, M.; Goitsuka, R. Nuclear localization of Meis1 in dermal papilla promotes hair matrix cell proliferation in the anagen phase of hair cycle. Biochem. Biophys. Res. Commun. 2019, 519, 727–733. [Google Scholar] [CrossRef]
- Oh, H.S.; Smart, R.C. An estrogen receptor pathway regulates the telogen-anagen hair follicle transition and influences epidermal cell proliferation. Proc. Natl. Acad. Sci. USA 1996, 93, 12525–12530. [Google Scholar] [CrossRef]
- Qiu, W.; Lei, M.; Zhou, L.; Bai, X.; Lai, X.; Yu, Y.; Yang, T.; Lian, X. Hair follicle stem cell proliferation, Akt and Wnt signaling activation in TPA-induced hair regeneration. Histochem. Cell Biol. 2017, 147, 749–758. [Google Scholar] [CrossRef]
- Kelly, Y.; Blanco, A.; Tosti, A. Androgenetic Alopecia: An Update of Treatment Options. Drugs 2016, 76, 1349–1364. [Google Scholar] [CrossRef]
- Matsumura, H.; Mohri, Y.; Binh, N.T.; Morinaga, H.; Fukuda, M.; Ito, M.; Kurata, S.; Hoeijmakers, J.; Nishimura, E.K. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science 2016, 351, aad4395. [Google Scholar] [CrossRef]
- Fernandez-Flores, A.; Saeb-Lima, M.; Cassarino, D.S. Histopathology of aging of the hair follicle. J. Cutan Pathol. 2019, 46, 508–519. [Google Scholar] [CrossRef]
- Soma, T.; Ogo, M.; Suzuki, J.; Takahashi, T.; Hibino, T. Analysis of apoptotic cell death in human hair follicles in vivo and in vitro. J. Invest. Dermatol. 1998, 111, 948–954. [Google Scholar] [CrossRef]
- Han, J.H.; Kwon, O.S.; Chung, J.H.; Cho, K.H.; Eun, H.C.; Kim, K.H. Effect of minoxidil on proliferation and apoptosis in dermal papilla cells of human hair follicle. J. Dermatol. Sci. 2004, 34, 91–98. [Google Scholar] [CrossRef]
- Lindner, G.; Botchkarev, V.A.; Botchkareva, N.V.; Ling, G.; van der Veen, C.; Paus, R. Analysis of apoptosis during hair follicle regression (catagen). Am. J. Pathol. 1997, 151, 1601–1617. [Google Scholar] [PubMed]
- Matsuo, K.; Mori, O.; Hashimoto, T. Apoptosis in murine hair follicles during catagen regression. Arch. Dermatol. Res 1998, 290, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Ferraris, C.; Cooklis, M.; Polakowska, R.R.; Haake, A.R. Induction of apoptosis through the PKC pathway in cultured dermal papilla fibroblasts. Exp. Cell. Res. 1997, 234, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Seiberg, M.; Wisniewski, S.; Cauwenbergh, G.; Shapiro, S.S. Trypsin-induced follicular papilla apoptosis results in delayed hair growth and pigmentation. Dev. Dyn. 1997, 208, 553–564. [Google Scholar] [CrossRef]
- Narita, M.; Nunez, S.; Heard, E.; Narita, M.; Lin, A.W.; Hearn, S.A.; Spector, D.L.; Hannon, G.J.; Lowe, S.W. Rb-mediated heterochromatin formation and silencing of E2F target genes during cellular senescence. Cell 2003, 113, 703–716. [Google Scholar] [CrossRef]
- Collado, M.; Blasco, M.A.; Serrano, M. Cellular senescence in cancer and aging. Cell 2007, 130, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Garza, L.A.; Yang, C.C.; Zhao, T.; Blatt, H.B.; Lee, M.; He, H.; Stanton, D.C.; Carrasco, L.; Spiegel, J.H.; Tobias, J.W.; et al. Bald scalp in men with androgenetic alopecia retains hair follicle stem cells but lacks CD200-rich and CD34-positive hair follicle progenitor cells. J. Clin. Invest. 2011, 121, 613–622. [Google Scholar] [CrossRef] [PubMed]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, J.Y.; Kim, J.; Choi, Y.-H.; Kang, N.-G.; Lee, S. Dexpanthenol Promotes Cell Growth by Preventing Cell Senescence and Apoptosis in Cultured Human Hair Follicle Cells. Curr. Issues Mol. Biol. 2021, 43, 1361-1373. https://doi.org/10.3390/cimb43030097
Shin JY, Kim J, Choi Y-H, Kang N-G, Lee S. Dexpanthenol Promotes Cell Growth by Preventing Cell Senescence and Apoptosis in Cultured Human Hair Follicle Cells. Current Issues in Molecular Biology. 2021; 43(3):1361-1373. https://doi.org/10.3390/cimb43030097
Chicago/Turabian StyleShin, Jae Young, Jaeyoon Kim, Yun-Ho Choi, Nae-Gyu Kang, and Sanghwa Lee. 2021. "Dexpanthenol Promotes Cell Growth by Preventing Cell Senescence and Apoptosis in Cultured Human Hair Follicle Cells" Current Issues in Molecular Biology 43, no. 3: 1361-1373. https://doi.org/10.3390/cimb43030097
APA StyleShin, J. Y., Kim, J., Choi, Y.-H., Kang, N.-G., & Lee, S. (2021). Dexpanthenol Promotes Cell Growth by Preventing Cell Senescence and Apoptosis in Cultured Human Hair Follicle Cells. Current Issues in Molecular Biology, 43(3), 1361-1373. https://doi.org/10.3390/cimb43030097