
Geraniol Averts Methotrexate-Induced Acute Kidney Injury via Keap1/Nrf2/HO-1 and MAPK/NF-κB Pathways




Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Statement
2.2. Experimental Design and Treatments
2.3. Serum and Tissue Homogenate Biochemical Markers
2.4. Determination of Kidney Index
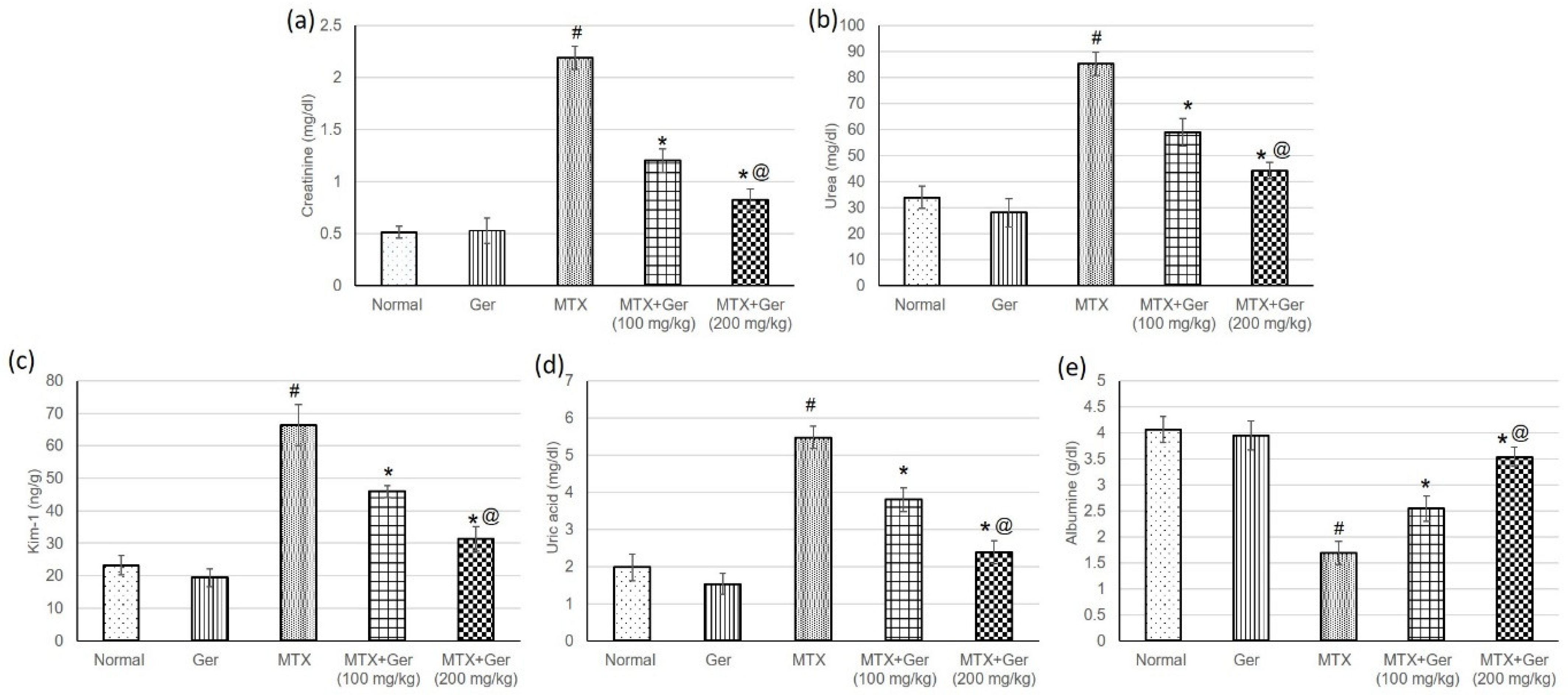
2.5. Determination of Renal Function
2.6. Determination of Oxidative Stress and Antioxidant Markers
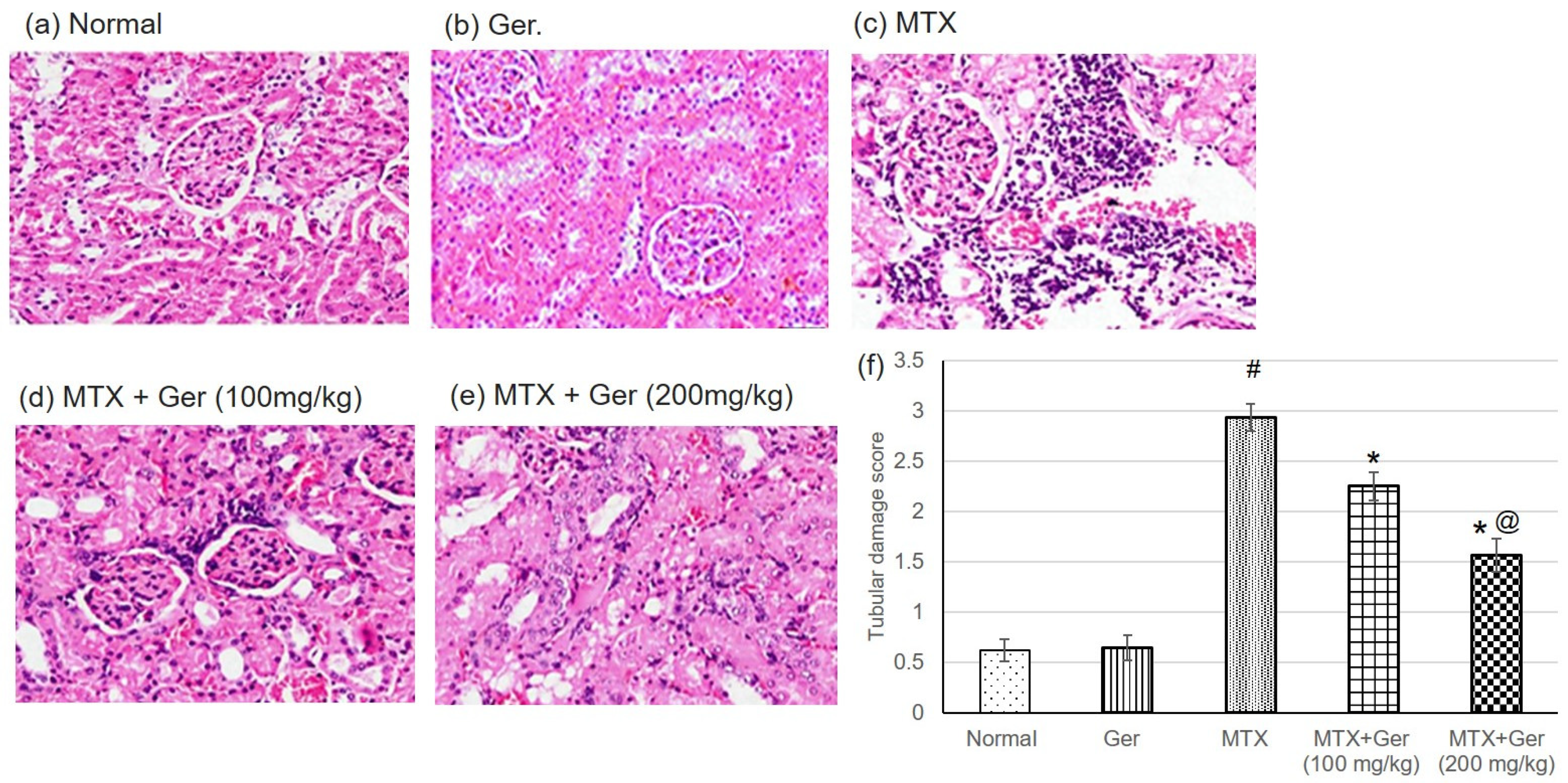
2.7. Histological Examination
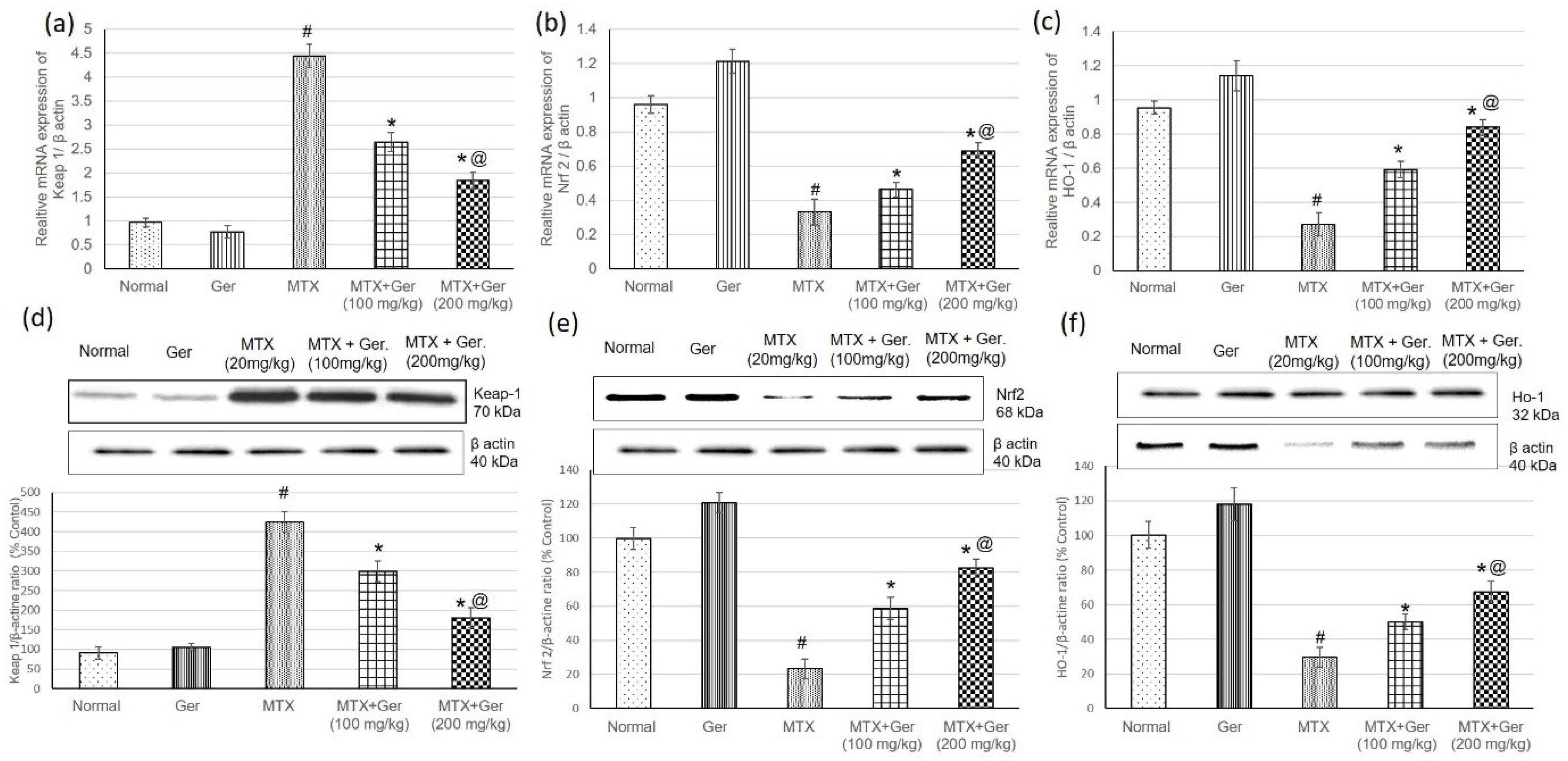
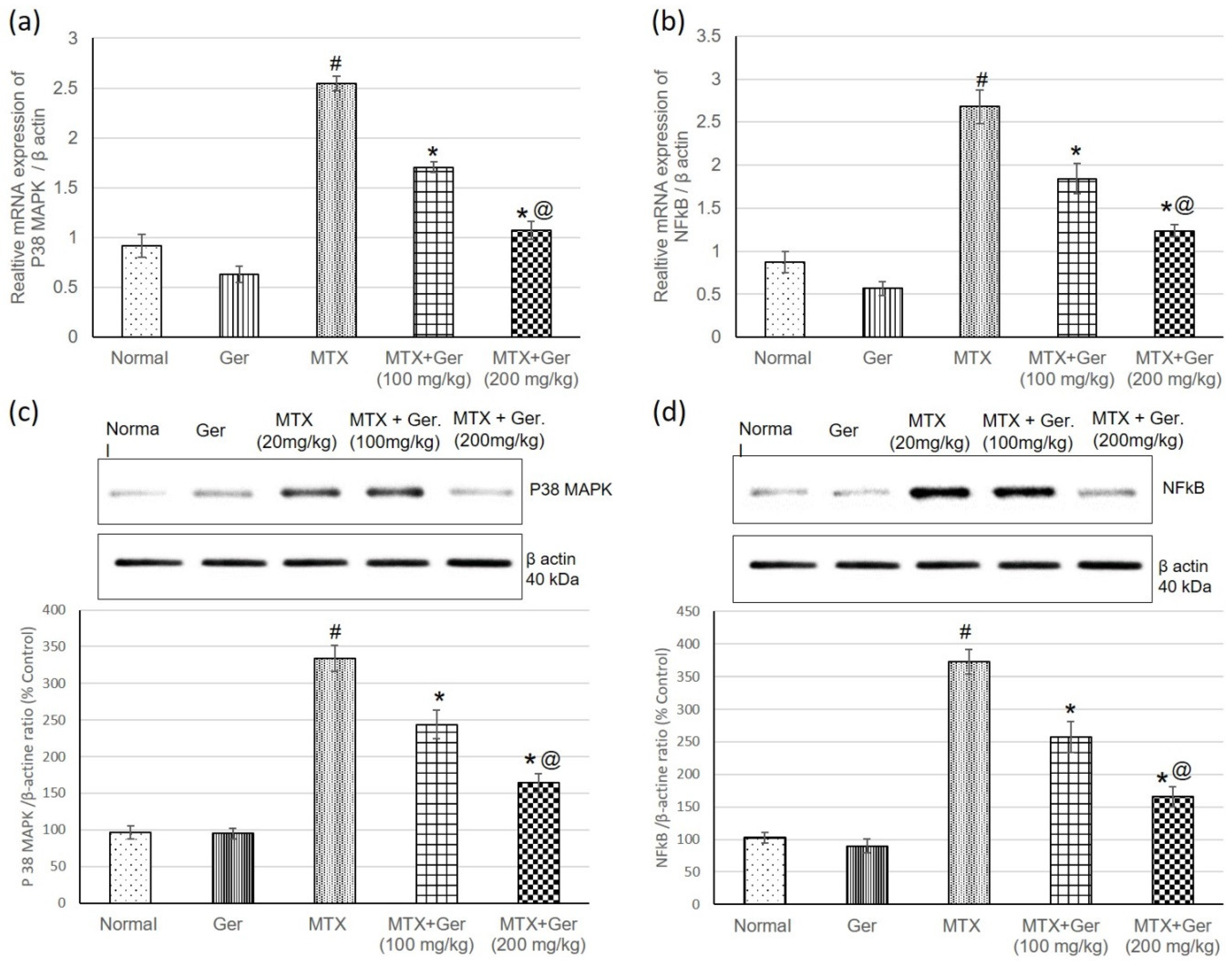
2.8. Real-Time PCR
2.9. Western Blot
2.10. Determination of Inflammatory Mediators
2.11. Determination of Apoptotic Signaling Markers
2.12. Statistical Analysis
3. Results
3.1. Ger Declined Kidney Index in MTX-Induced AKI in Rats
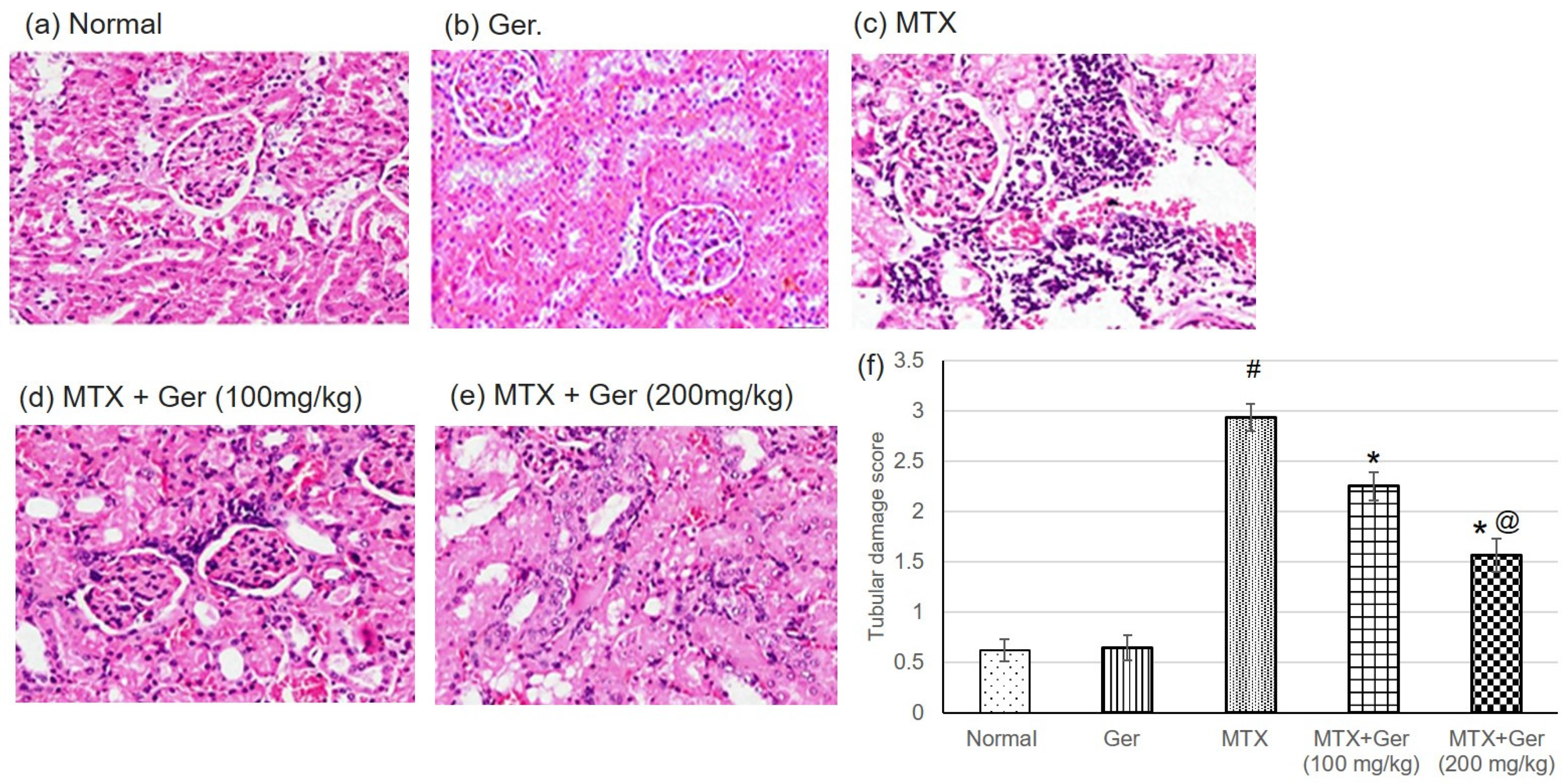
3.2. Ger Enhanced Renal Histopathological Alterations in MTX-Induced AKI in Rats
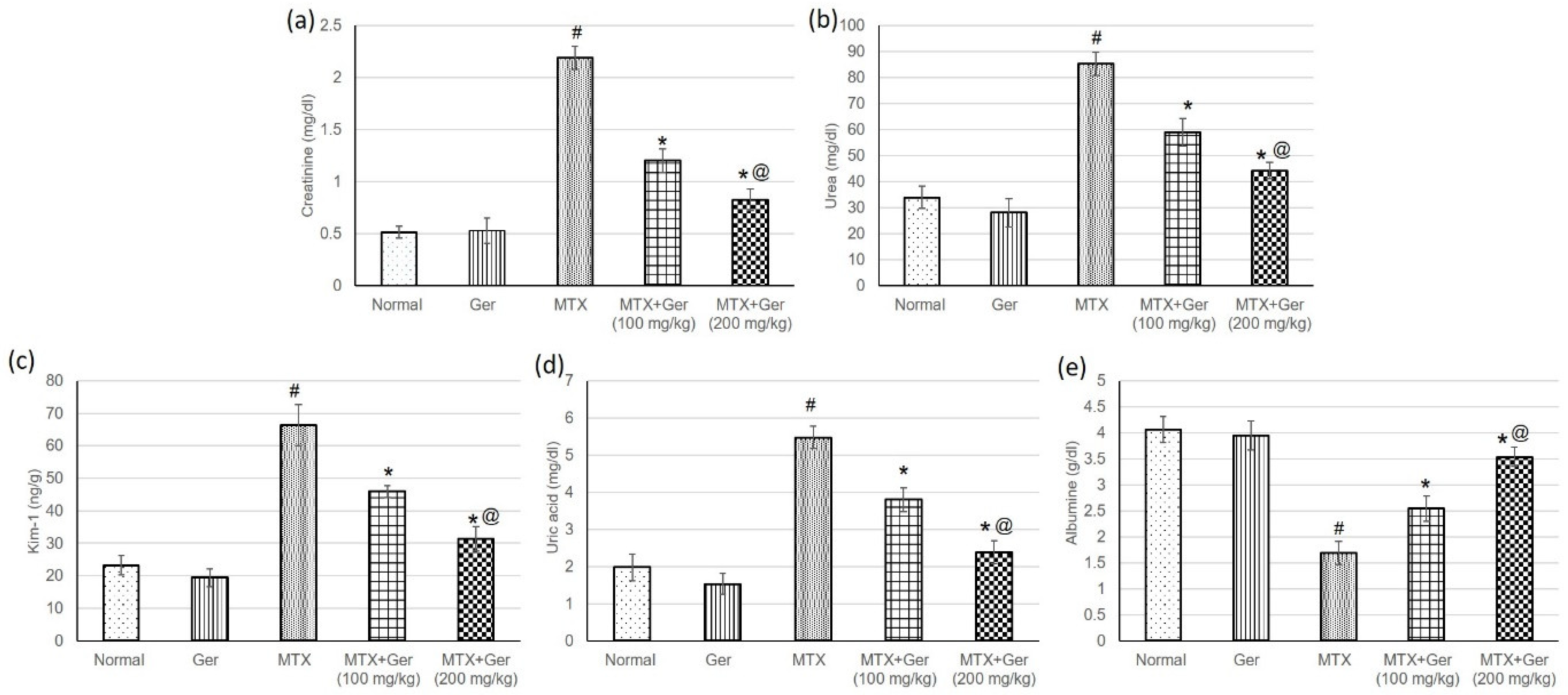
3.3. Ger Averted MTX-Induced Kidney Injury in Rats
3.4. Ger Regulated Renal Keap1/Nrf2/HO-1 Pathway in MTX-Induced AKI
3.5. Ger Enhanced Antioxidants Activities in MTX-Induced AKI
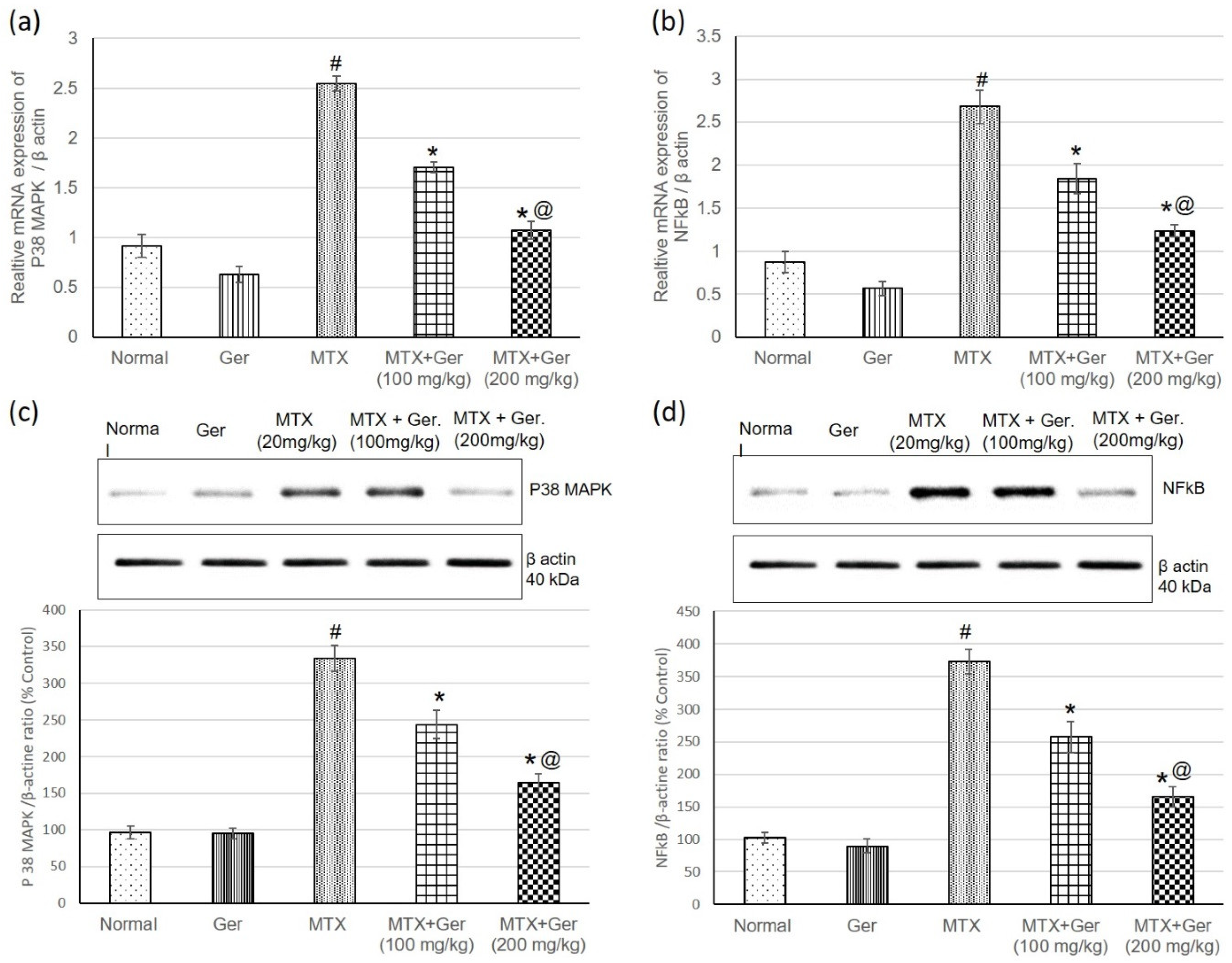
3.6. Ger Diminished Inflammatory Response via P38 MAPK/NF-κB Pathway in MTX-Induced AKI
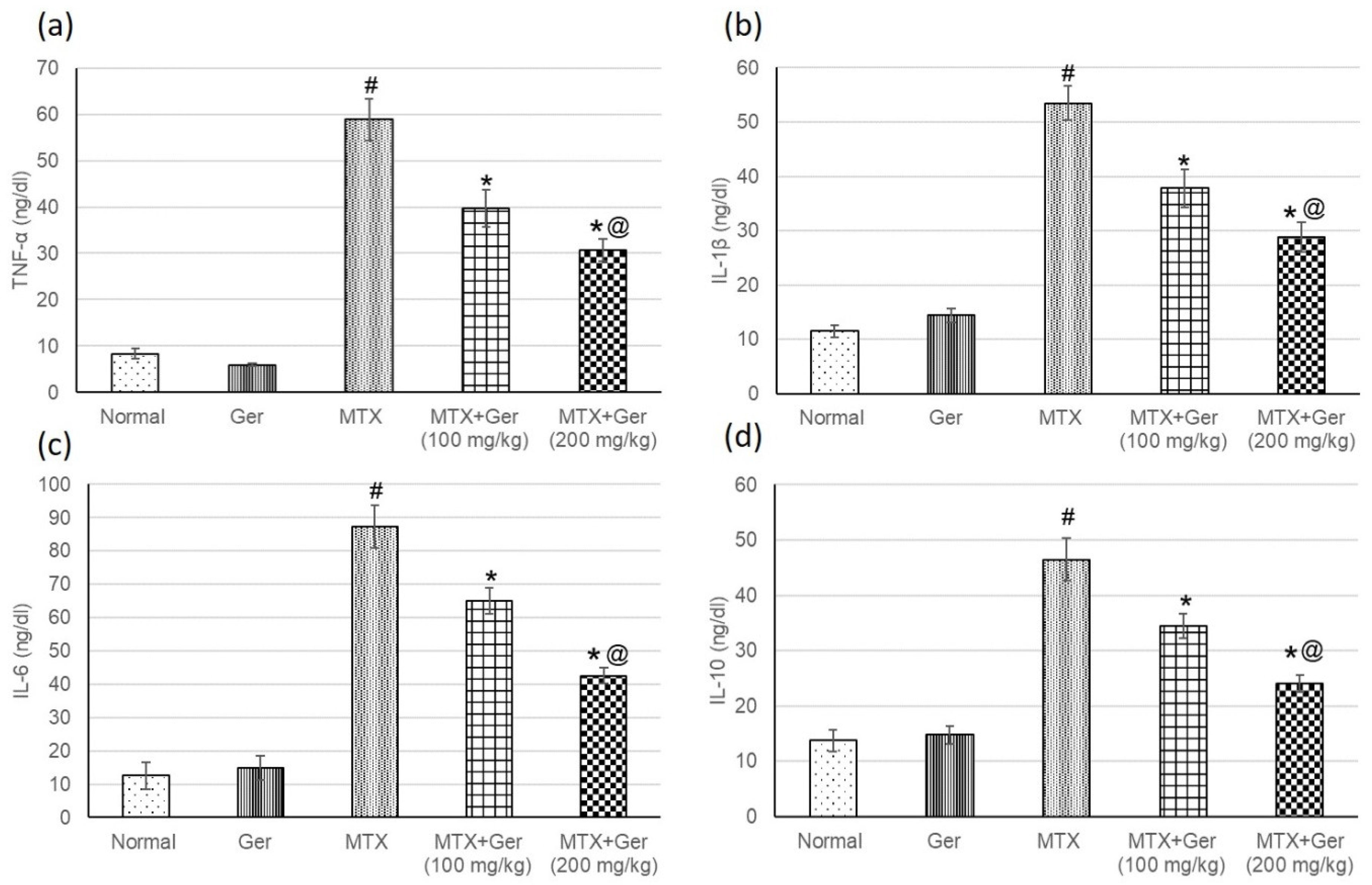
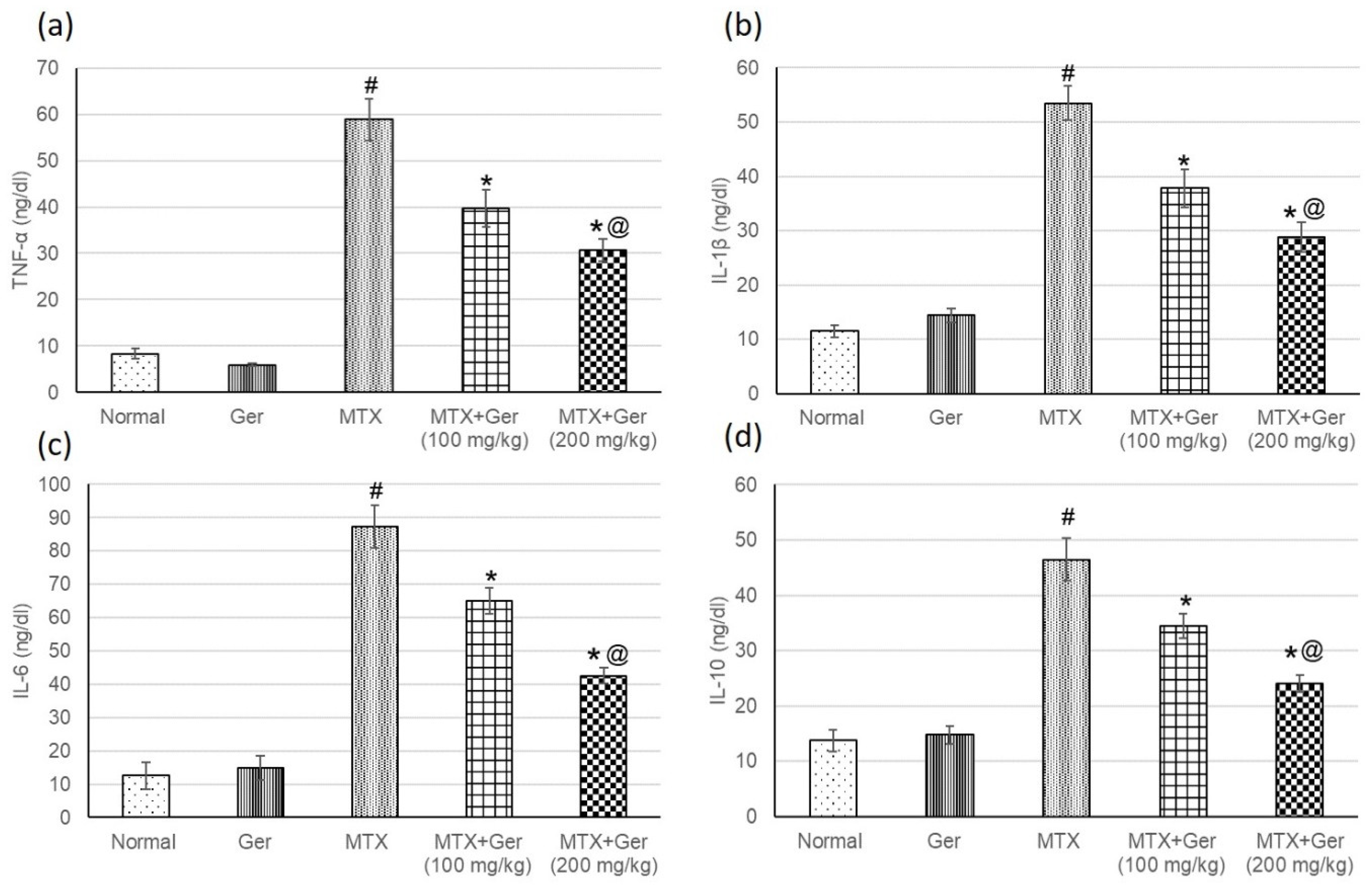
3.7. Ger Diminished Inflammatory Mediators in MTX-Induced AKI
3.8. Ger Mitigated Apoptotic Signaling in MTX-Induced AKI via Regulation of Bax/Bcl2/Caspase-3 and -9
4. Discussion
4.1. Ger and the Status of Renal Damage Induced by MTX
4.2. Ger and Oxidative Stress Parameters and Pathways
4.3. Ger and Inflammation Parameters and Pathways
4.4. Ger and Apoptotic Signaling
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abd El-Twab, S.M.; Hussein, O.E.; Hozayen, W.G.; Bin-Jumah, M.; Mahmoud, A.M. Chicoric acid prevents methotrexate-induced kidney injury by suppressing NF-κB/NLRP3 inflammasome activation and up-regulating Nrf2/ARE/HO-1 signaling. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2019, 68, 511–523. [Google Scholar] [CrossRef]
- Abd El-Twab, S.M.; Hozayen, W.G.; Hussein, O.E.; Mahmoud, A.M. 18β-Glycyrrhetinic acid protects against methotrexate-induced kidney injury by up-regulating the Nrf2/ARE/HO-1 pathway and endogenous antioxidants. Ren. Fail. 2016, 38, 1516–1527. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Hussein, O.E.; Abd El-Twab, S.M.; Hozayen, W.G. Ferulic acid protects against methotrexate nephrotoxicity via activation of Nrf2/ARE/HO-1 signaling and PPARγ, and suppression of NF-κB/NLRP3 inflammasome axis. Food Funct. 2019, 10, 4593–4607. [Google Scholar] [CrossRef]
- Mehta, R.L.; Awdishu, L.; Davenport, A.; Murray, P.T.; Macedo, E.; Cerda, J.; Chakaravarthi, R.; Holden, A.L.; Goldstein, S.L. Phenotype standardization for drug-induced kidney disease. Kidney Int. 2015, 88, 226–234. [Google Scholar] [CrossRef]
- Heidari, R.; Ahmadi, A.; Mohammadi, H.; Ommati, M.M.; Azarpira, N.; Niknahad, H. Mitochondrial dysfunction and oxidative stress are involved in the mechanism of methotrexate-induced renal injury and electrolytes imbalance. Biomed. Pharmacother. 2018, 107, 834–840. [Google Scholar] [CrossRef]
- Kolli, V.K.; Abraham, P.; Isaac, B.; Selvakumar, D. Neutrophil infiltration and oxidative stress may play a critical role in methotrexate-induced renal damage. Chemotherapy 2009, 55, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Widemann, B.C.; Adamson, P.C. Understanding and managing methotrexate nephrotoxicity. Oncologist 2006, 11, 694–703. [Google Scholar] [CrossRef] [Green Version]
- Perazella, M.A. Drug-induced acute kidney injury: Diverse mechanisms of tubular injury. Curr. Opin. Crit. Care 2019, 25, 550–557. [Google Scholar] [CrossRef] [PubMed]
- Hassanein, E.H.M.; Mohamed, W.R.; Shalkami, A.S.; Khalaf, M.M.; Hemeida, R.A.M. Renoprotective effects of umbelliferone on methotrexate-induced renal injury through regulation of Nrf-2/Keap-1, P(38)MAPK/NF-κB, and apoptosis signaling pathways. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 116, 152–160. [Google Scholar] [CrossRef]
- Ishii, T.; Itoh, K.; Takahashi, S.; Sato, H.; Yanagawa, T.; Katoh, Y.; Bannai, S.; Yamamoto, M. Transcription factor Nrf2 coordinately regulates a group of oxidative stress-inducible genes in macrophages. J. Biol. Chem. 2000, 275, 16023–16029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aladaileh, S.H.; Hussein, O.E.; Abukhalil, M.H.; Saghir, S.A.M.; Bin-Jumah, M.; Alfwuaires, M.A.; Germoush, M.O.; Almaiman, A.A.; Mahmoud, A.M. Formononetin Upregulates Nrf2/HO-1 Signaling and Prevents Oxidative Stress, Inflammation, and Kidney Injury in Methotrexate-Induced Rats. Antioxidants 2019, 8, 430. [Google Scholar] [CrossRef] [Green Version]
- Younis, N.S.; Abduldaium, M.S.; Mohamed, M.E. Protective Effect of Geraniol on Oxidative, Inflammatory and Apoptotic Alterations in Isoproterenol-Induced Cardiotoxicity: Role of the Keap1/Nrf2/HO-1 and PI3K/Akt/mTOR Pathways. Antioxidants 2020, 9, 977. [Google Scholar] [CrossRef]
- Mohamed, M.E.; Abduldaium, Y.S.; Younis, N.S. Ameliorative Effect of Linalool in Cisplatin-Induced Nephrotoxicity: The Role of HMGB1/TLR4/NF-κB and Nrf2/HO1 Pathways. Biomolecules 2020, 10, 1488. [Google Scholar] [CrossRef]
- Mohamed, M.E.; Abduldaium, M.S.; Younis, N.S. Cardioprotective Effect of Linalool against Isoproterenol-Induced Myocardial Infarction. Life 2021, 11, 120. [Google Scholar] [CrossRef]
- Liu, M.; Grigoryev, D.N.; Crow, M.T.; Haas, M.; Yamamoto, M.; Reddy, S.P.; Rabb, H. Transcription factor Nrf2 is protective during ischemic and nephrotoxic acute kidney injury in mice. Kidney Int. 2009, 76, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Martindale, J.L.; Holbrook, N.J. Cellular response to oxidative stress: Signaling for suicide and survival. J. Cell. Physiol. 2002, 192, 1–15. [Google Scholar] [CrossRef]
- Hassanein, E.H.M.; Shalkami, A.S.; Khalaf, M.M.; Mohamed, W.R.; Hemeida, R.A.M. The impact of Keap1/Nrf2, P(38)MAPK/NF-κB and Bax/Bcl2/caspase-3 signaling pathways in the protective effects of berberine against methotrexate-induced nephrotoxicity. Biomed. Pharmacother. 2019, 109, 47–56. [Google Scholar] [CrossRef]
- Xue, Z.; Zhang, X.G.; Wu, J.; Xu, W.C.; Li, L.Q.; Liu, F.; Yu, J.E. Effect of treatment with geraniol on ovalbumin-induced allergic asthma in mice. Ann. Allergy Asthma Immunol. Off. Publ. Am. Coll. Allergy Asthma Immunol. 2016, 116, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Kakkar, P. Plant derived antioxidants–Geraniol and camphene protect rat alveolar macrophages against t-BHP induced oxidative stress. Toxicol. In Vitro 2009, 23, 295–301. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, K.I.; Bonamin, F.; Dos Santos, R.C.; Périco, L.L.; Beserra, F.P.; de Sousa, D.P.; Filho, J.M.; da Rocha, L.R.; Hiruma-Lima, C.A. Geraniol-a flavoring agent with multifunctional effects in protecting the gastric and duodenal mucosa. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2014, 387, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Cho, M.; So, I.; Chun, J.N.; Jeon, J.H. The antitumor effects of geraniol: Modulation of cancer hallmark pathways (Review). Int. J. Oncol. 2016, 48, 1772–1782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, X.Y.; Xue, J.S.; Li, H.Y.; Ma, Z.Q.; Fu, Q.; Qu, R.; Ma, S.P. Geraniol produces antidepressant-like effects in a chronic unpredictable mild stress mice model. Physiol. Behav. 2015, 152, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Kandeil, M.A.; Mahmoud, M.O.; Abdel-Razik, A.H.; Gomaa, S.B. Thymoquinone and geraniol alleviate cisplatin-induced neurotoxicity in rats through downregulating the p38 MAPK/STAT-1 pathway and oxidative stress. Life Sci. 2019, 228, 145–151. [Google Scholar] [CrossRef]
- El-Said, Y.A.M.; Sallam, N.A.A.; Ain-Shoka, A.A.; Abdel-Latif, H.A. Geraniol ameliorates diabetic nephropathy via interference with miRNA-21/PTEN/Akt/mTORC1 pathway in rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 2325–2337. [Google Scholar] [CrossRef] [PubMed]
- Armagan, I.; Bayram, D.; Candan, I.A.; Yigit, A.; Celik, E.; Armagan, H.H.; Uğuz, A.C. Effects of pentoxifylline and alpha lipoic acid on methotrexate-induced damage in liver and kidney of rats. Environ. Toxicol. Pharmacol. 2015, 39, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Grisham, M.B.; Johnson, G.G.; Lancaster, J.R., Jr. Quantitation of nitrate and nitrite in extracellular fluids. Methods Enzymol. 1996, 268, 237–246. [Google Scholar] [CrossRef]
- Hazelhoff, M.H.; Bulacio, R.P.; Chevalier, A.; Torres, A.M. Renal expression of organic anion transporters is modified after mercuric chloride exposure: Gender-related differences. Toxicol. Lett. 2018, 295, 390–396. [Google Scholar] [CrossRef]
- Bonventre, J.V. Kidney Injury Molecule-1 (KIM-1): A specific and sensitive biomarker of kidney injury. Scand. J. Clin. Lab. Investig. Suppl. 2008, 241, 78–83. [Google Scholar] [CrossRef]
- Mateos, R.; Lecumberri, E.; Ramos, S.; Goya, L.; Bravo, L. Determination of malondialdehyde (MDA) by high-performance liquid chromatography in serum and liver as a biomarker for oxidative stress: Application to a rat model for hypercholesterolemia and evaluation of the effect of diets rich in phenolic antioxidants from fruits. J. Chromatogr. B 2005, 827, 76–82. [Google Scholar] [CrossRef]
- Gowda, S.; Desai, P.B.; Kulkarni, S.S.; Hull, V.V.; Math, A.A.; Vernekar, S.N. Markers of renal function tests. N. Am. J. Med. Sci. 2010, 2, 170–173. [Google Scholar]
- Mahmoud, A.M.; Germoush, M.O.; Al-Anazi, K.M.; Mahmoud, A.H.; Farah, M.A.; Allam, A.A. Commiphora molmol protects against methotrexate-induced nephrotoxicity by up-regulating Nrf2/ARE/HO-1 signaling. Biomed. Pharmacother. 2018, 106, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Smeland, E.; Fuskevåg, O.M.; Nymann, K.; Svendesn, J.S.; Olsen, R.; Lindal, S.; Bremnes, R.M.; Aarbakke, J. High-dose 7-hydromethotrexate: Acute toxicity and lethality in a rat model. Cancer Chemother. Pharmacol. 1996, 37, 415–422. [Google Scholar] [CrossRef]
- Dierkes, J.; Westphal, S. Effect of drugs on homocysteine concentrations. Semin. Vasc. Med. 2005, 5, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Su, B.; Zhu, H.; Chen, C.; Zhao, G. Protective effect of geraniol inhibits inflammatory response, oxidative stress and apoptosis in traumatic injury of the spinal cord through modulation of NF-κB and p38 MAPK. Exp. Ther. Med. 2016, 12, 3607–3613. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, S.M.; Hejazian, L.B.; Amani, R.; Siahchehreh Badeli, N. Geraniol attenuates oxidative stress, bioaccumulation, serological and histopathological changes during aluminum chloride-hepatopancreatic toxicity in male Wistar rats. Environ. Sci. Pollut. Res. Int. 2020, 27, 20076–20089. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Z.; Fu, X.; Lin, Z.; Yu, K. Geraniol-mediated osteoarthritis improvement by down-regulating PI3K/Akt/NF-κB and MAPK signals: In vivo and in vitro studies. Int. Immunopharmacol. 2020, 86, 106713. [Google Scholar] [CrossRef]
- Bhattamisra, S.K.; Yean Yan, V.L.; Koh Lee, C.; Hui Kuean, C.; Candasamy, M.; Liew, Y.K.; Sahu, P.S. Protective activity of geraniol against acetic acid and Helicobacter pylori- induced gastric ulcers in rats. J. Tradit. Complement. Med. 2019, 9, 206–214. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Hozayen, W.G.; Ramadan, S.M. Berberine ameliorates methotrexate-induced liver injury by activating Nrf2/HO-1 pathway and PPARγ, and suppressing oxidative stress and apoptosis in rats. Biomed. Pharmacother. 2017, 94, 280–291. [Google Scholar] [CrossRef]
- El-Emam, S.Z.; Soubh, A.A.; Al-Mokaddem, A.K.; Abo El-Ella, D.M. Geraniol activates Nrf-2/HO-1 signaling pathway mediating protection against oxidative stress-induced apoptosis in hepatic ischemia-reperfusion injury. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2020, 393, 1849–1858. [Google Scholar] [CrossRef]
- Jayachandran, M.; Chandrasekaran, B.; Namasivayam, N. Geraniol attenuates oxidative stress by Nrf2 activation in diet-induced experimental atherosclerosis. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 335–346. [Google Scholar] [CrossRef]
- Mohammed, M.J.; Tadros, M.G.; Michel, H.E. Geraniol protects against cyclophosphamide-induced hepatotoxicity in rats: Possible role of MAPK and PPAR-γ signaling pathways. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2020, 139, 111251. [Google Scholar] [CrossRef] [PubMed]
- Soubh, A.A.; Abdallah, D.M.; El-Abhar, H.S. Geraniol ameliorates TNBS-induced colitis: Involvement of Wnt/β-catenin, p38MAPK, NFκB, and PPARγ signaling pathways. Life Sci. 2015, 136, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Zhang, T.; Yin, N.; Ma, X.; Zhao, G.; Wu, H.; Qiu, C.; Deng, G. Geraniol alleviates LPS-induced acute lung injury in mice via inhibiting inflammation and apoptosis. Oncotarget 2017, 8, 71038–71053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rekha, K.R.; Selvakumar, G.P. Gene expression regulation of Bcl2, Bax and cytochrome-C by geraniol on chronic MPTP/probenecid induced C57BL/6 mice model of Parkinson’s disease. Chem. Biol. Interact. 2014, 217, 57–66. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Markers | Primer Sequence (5′ to 3′) | |
|---|---|---|
| Forward Primer | Reverse Primers | |
| Bcl-2 | 5′-CCGGGAGATCGTGATGAAGT-3′ | 3′-ATCCCAGCCTCCGTTATCCT-5′ |
| Bax | 5′-GTGGTTGCCCTCTTCTACTTTG-3′ | 3′-CACAAAGATGGTCACTGTCTGC-5′ |
| Nrf2 | 5′-CATTTGTAGATGACCATGAGTCGC-3′ | 3′-ATCAGGGGTGGTGAAGACTG-5′ |
| Keap-1 | 5′-CTTCGGGGAGGAGGAGTTCT-3′ | 3′-CGTTCAGATCATCGCGGCTG-5′ |
| HO-1 | 5′-GTGCACATCCGTGCAGAGAA-3′ | 3′-GTGCACATCCGTGCAGAGAA-5′ |
| P38 MAPK | 5′-AGAGTCTCTGTCGACCTGCT-3′ | 3′-CCTGCTTTCAAAGGACTGGT-5′ |
| NF-κB | 5′-TGGGACGACACCTCTACACA-3′ | 3′-GGAGCTCATCTCATAGTTGTCC-5′ |
| β-actin | 5′-TGACAGGATGCAGAAGGAGA-3′ | 3′-TAGAGCCACCAATCCACACA-5′ |
| Treated Groups | Body Weight (gm) | Kidney Index | |
|---|---|---|---|
| Initial Body Weight | Final Body Weight | ||
| Normal | 190 ± 8.68 | 275 ± 14.5 | 0.71 ± 0.04 |
| Geraniol | 200 ± 11.46 | 280 ± 17.6 | 0.72 ± 0.03 |
| MTX (20 mg/kg) | 195.72 ± 19.22 | 140.62 ± 8.96 # | 1.19 ± 0.08 # |
| MTX + Ger (100 mg/kg) | 195.48 ± 21.15 | 185.00 ± 12.76 | 0.87 ± 0.10 * |
| MTX + Ger (200 mg/kg) | 210.00 ± 20.87 | 198.00 ± 13.7 | 0.85 ± 0.08 * |
| Normal | Ger | MTX (20 mg/kg) | MTX + Ger (100 mg/kg) | MTX + Ger (200 mg/kg) | |
|---|---|---|---|---|---|
| MDA (nmol/100 mg) | 22.60 ± 3.65 | 25.34 ± 5.32 | 92.83 ± 8.37 # | 34.96 ± 5.04 * | 30.64 ± 4.15 * |
| NO (nmol/100 mg) | 21.39 ± 0.30 | 20.76 ± 3.59 | 79.24 ± 0.38 # | 52.46 ± 0.22 * | 32.01 ± 0.18 *,@ |
| Reduced GSH (nmol/100 mg) | 69.56 ± 8.48 | 67.12 ± 13.76 | 22.91 ± 2.12 # | 32.27 ± 3.51 * | 48.97 ± 6.51 *,@ |
| GSHPx (U/g protein) | 0.060 ± 0.01 | 0.062 ± 0.0093 | 0.045 ± 0.02 | 0.051 ± 0.01 * | 0.055 ± 0.02 *,@ |
| SOD (U/g protein) | 292.92 ± 22.43 | 300.32 ± 24.61 | 156.28 ± 0.69 # | 205.33 ± 22.4 * | 253.87 ± 11.31 *,@ |
| CAT (nmol/g tissue) | 76.13 ± 5.34 | 82.23 ± 15.28 | 22.70 ± 0.66 # | 34.80 ± 5.16 * | 48.75 ± 4.38 *,@ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Younis, N.S.; Elsewedy, H.S.; Shehata, T.M.; Mohamed, M.E. Geraniol Averts Methotrexate-Induced Acute Kidney Injury via Keap1/Nrf2/HO-1 and MAPK/NF-κB Pathways. Curr. Issues Mol. Biol. 2021, 43, 1741-1755. https://doi.org/10.3390/cimb43030123
Younis NS, Elsewedy HS, Shehata TM, Mohamed ME. Geraniol Averts Methotrexate-Induced Acute Kidney Injury via Keap1/Nrf2/HO-1 and MAPK/NF-κB Pathways. Current Issues in Molecular Biology. 2021; 43(3):1741-1755. https://doi.org/10.3390/cimb43030123
Chicago/Turabian StyleYounis, Nancy S., Heba S. Elsewedy, Tamer M. Shehata, and Maged E. Mohamed. 2021. "Geraniol Averts Methotrexate-Induced Acute Kidney Injury via Keap1/Nrf2/HO-1 and MAPK/NF-κB Pathways" Current Issues in Molecular Biology 43, no. 3: 1741-1755. https://doi.org/10.3390/cimb43030123
APA StyleYounis, N. S., Elsewedy, H. S., Shehata, T. M., & Mohamed, M. E. (2021). Geraniol Averts Methotrexate-Induced Acute Kidney Injury via Keap1/Nrf2/HO-1 and MAPK/NF-κB Pathways. Current Issues in Molecular Biology, 43(3), 1741-1755. https://doi.org/10.3390/cimb43030123