
Chemical Profiling and Assessment of Analgesic and Anti-Inflammatory Activity of Ammoides verticillata Essential Oil: In Vitro, In Vivo, and In Silico Studies

 , , , ,
, , , ,
Abstract

1. Introduction
2. Results
2.1. Phytochemical Chemical Profiling of A. verticillata Essential Oil: Composition Analysis and Yield
2.2. Antioxidant Potential
2.3. Acute Toxicity Assessement
2.4. In Vivo Anti-Inflammatory Effects
2.5. Analgesic Properties
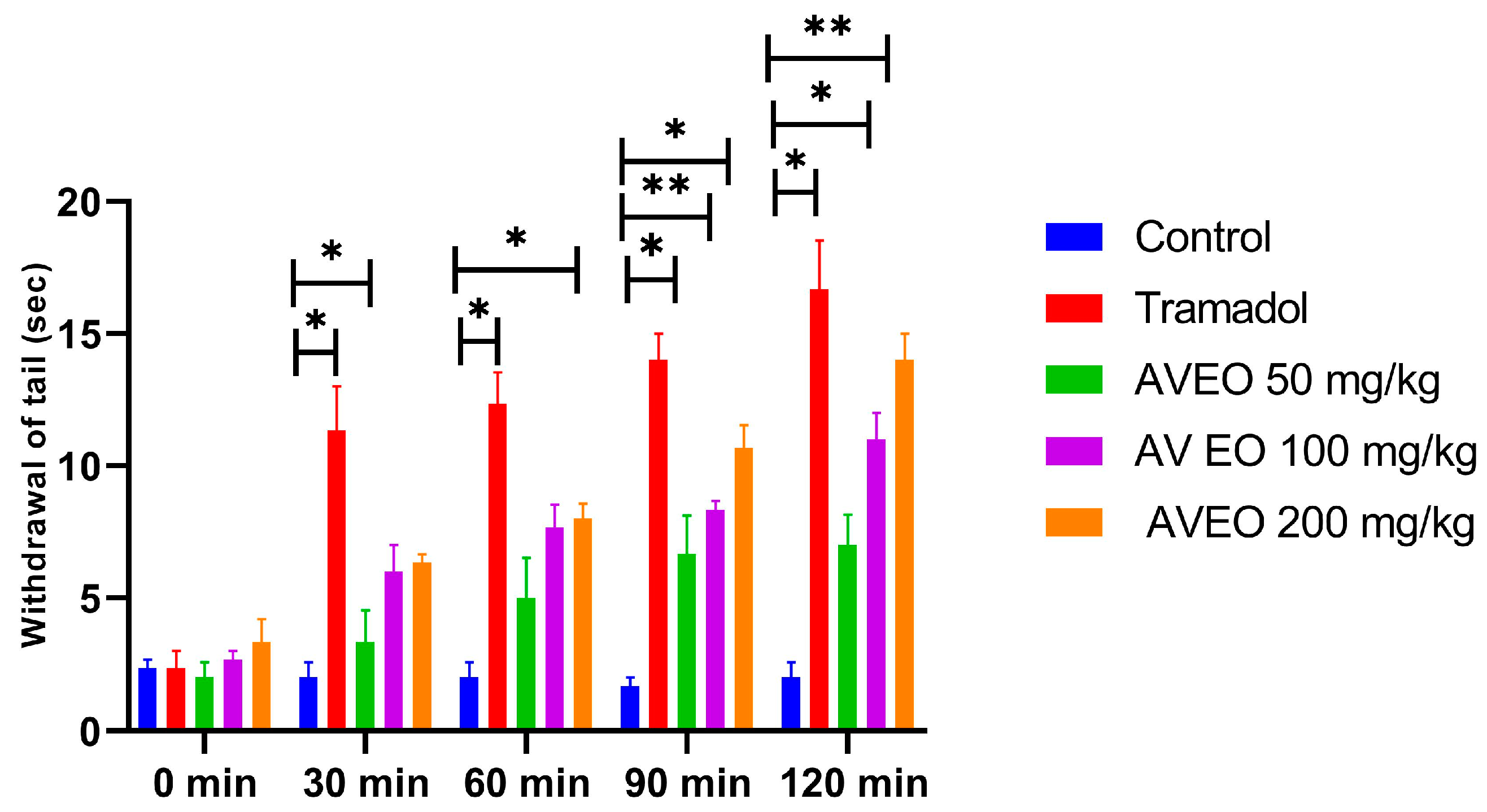
2.5.1. Tail Immersion Test: Central Anti-Nociceptive Activity
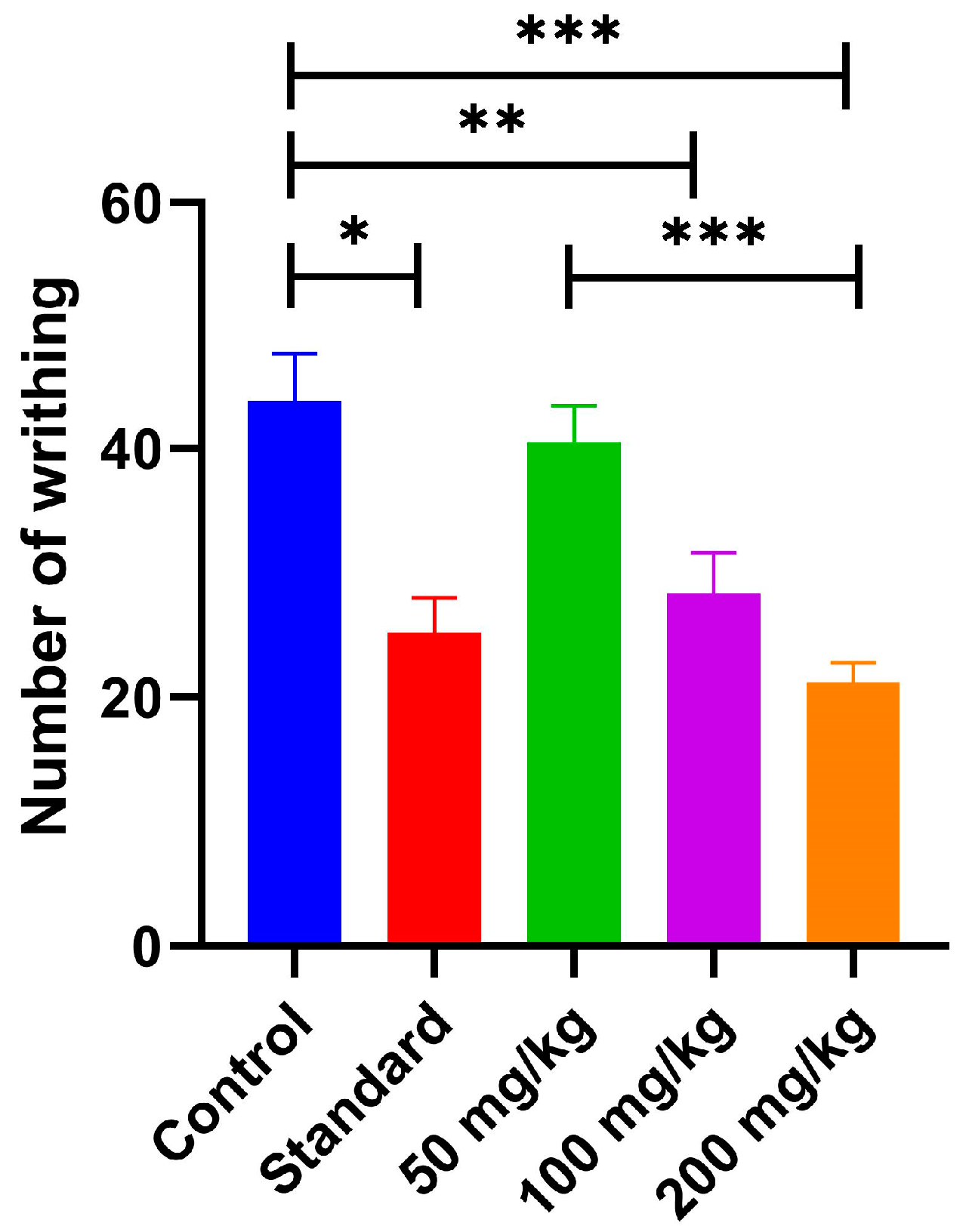
2.5.2. Acetic Acid-Induced Writhing Test: Peripheral Anti-Nociceptive Activity
2.6. In Vitro COX-1 and COX-2 Inhibition Assays
2.7. Molecular Insights into COX-2 Inhibition: Computational Findings
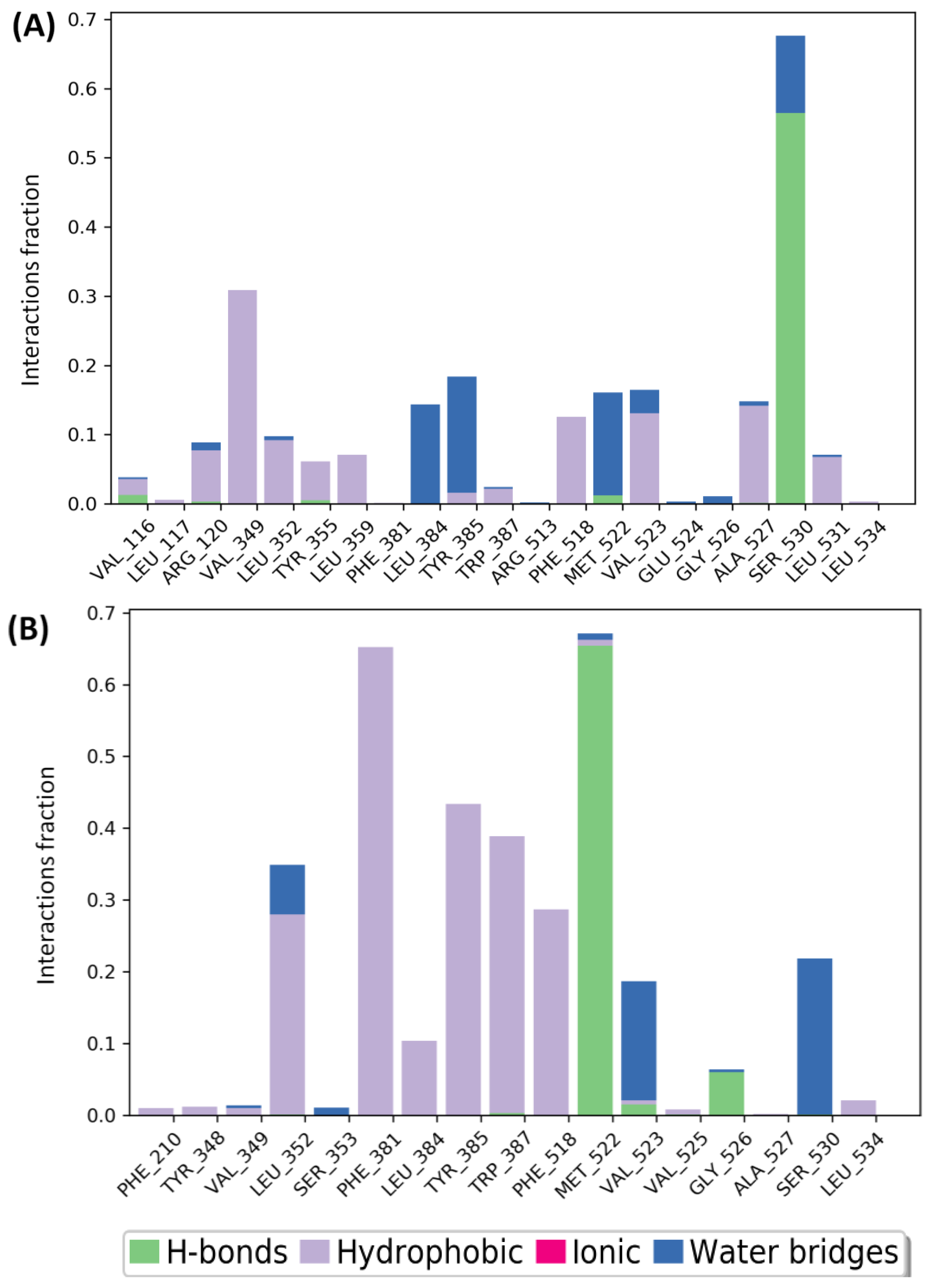
2.7.1. Molecular Docking and MMGBSA Findings
2.7.2. ADME Prediction
2.7.3. Molecular Dynamics Simulations
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Plant Collection and Identification
4.3. Experiment Animals
4.4. Extraction and Chemical Characterization of Essential Oil
4.5. Assessing Antioxidant Efficacy
4.5.1. DPPH Radical Scavenging Assay
4.5.2. ABTS Radical Scavenging Assay
4.5.3. Ferric Reducing Power Assay (FRAP)
4.5.4. Total Antioxidant Capacity (TAC)
4.6. Acute Oral Toxicity Assessement
4.7. Anti-Inflammatory Activity
paw thickness (0 h)]/average paw thickness (0 h).
4.8. Analgesic Activity
4.8.1. Tail Immersion Test
4.8.2. Acetic Acid-Induced Writhing Test
4.9. In Vitro Cyclooxygenas-1 and Cyclooxygenase-2 Inhibition Assays
4.10. Computational Studies
4.10.1. Molecular Docking
4.10.2. Prime MMGBSA Calculations
4.10.3. Drug-Likeness and ADME Prediction
4.10.4. Molecular Dynamics Simulations (MD)
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lee, J.S.; Lee, E.J.; Na, H.K.; Surh, Y.J. From Inflammation to Cancer: Opportunities for Chemoprevention via Dietary Intervention. In Immunity and Inflammation in Health and Disease; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2018; pp. 203–211. [Google Scholar]
- Greten, F.R.; Grivennikov, S.I. Inflammation and cancer: Triggers, mechanisms, and consequences. Immunity 2019, 51, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Aziz, N.; Kim, M.Y.; Cho, J.Y. Anti-inflammatory effects of luteolin: A review of in vitro, in vivo, and in silico studies. J. Ethnopharmacol. 2018, 225, 342–358. [Google Scholar] [CrossRef]
- Tu, Z.; Zhong, Y.; Hu, H.; Shao, D.; Haag, R.; Schirner, M.; Leong, K.W. Design of therapeutic biomaterials to control inflammation. Nat. Rev. Mater. 2022, 7, 557–574. [Google Scholar] [CrossRef] [PubMed]
- Noti, V.; Pontiki, E.; Hadjipavlou-Litina, D. Development of Novel Pyrrole Derivatives and Their Cinnamic Hybrids as Dual COX-2/LOX Inhibitors. Molecules 2023, 28, 7958. [Google Scholar] [CrossRef]
- Liu, B.; Wang, J.; Li, Y.Y.; Li, K.P.; Zhang, Q. The association between systemic immune-inflammation index and rheumatoid arthritis: Evidence from NHANES 1999–2018. Arthritis Res. Ther. 2023, 25, 34. [Google Scholar] [CrossRef]
- Yu, Y.; Yan, Y.; Niu, F.; Wang, Y.; Chen, X.; Su, G.; Xiong, Y. Ferroptosis: A cell death connecting oxidative stress, inflammation and cardiovascular diseases. Cell Death Discov. 2021, 7, 193. [Google Scholar] [CrossRef]
- Kim, T.K.; Bae, E.J.; Jung, B.C.; Choi, M.; Shin, S.J.; Park, S.J.; Lee, S.J. Inflammation promotes synucleinopathy propagation. Exp. Mol. Med. 2022, 54, 2148–2161. [Google Scholar] [CrossRef] [PubMed]
- Jiang, B.C.; Liu, T.; Gao, Y.J. Chemokines in chronic pain: Cellular and molecular mechanisms and therapeutic potential. Pharmacol. Ther. 2020, 212, 107581. [Google Scholar] [CrossRef]
- Chen, O.; Luo, X.; Ji, R.R. Macrophages and microglia in inflammation and neuroinflammation underlying different pain states. Med. Rev. 2023, 3, 381–407. [Google Scholar] [CrossRef]
- El-Miligy, M.M.; Al-Kubeisi, A.K.; Bekhit, M.G.; El-Zemity, S.R.; Nassra, R.A.; Hazzaa, A.A. Towards safer anti-inflammatory therapy: Synthesis of new thymol–pyrazole hybrids as dual COX-2/5-LOX inhibitors. J. Enzym. Inhib. Med. Chem. 2023, 38, 294–308. [Google Scholar] [CrossRef]
- Khan, S.; Andrews, K.L.; Chin-Dusting, J.P. Cyclo-oxygenase (COX) inhibitors and cardiovascular risk: Are non-steroidal anti-inflammatory drugs really anti-inflammatory? Int. J. Mol. Sci. 2019, 20, 4262. [Google Scholar] [CrossRef]
- Kassab, S.E.; Khedr, M.A.; Ali, H.I.; Abdalla, M.M. Discovery of new indomethacin-based analogs with potentially selective cyclooxygenase-2 inhibition and observed diminishing to PGE2 activities. Eur. J. Med. Chem. 2017, 141, 306–321. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhou, Y. Endothelium-dependent contraction: The non-classical action of endothelial prostacyclin, its underlying mechanisms, and implications. FASEB J. 2021, 35, e21877. [Google Scholar] [CrossRef] [PubMed]
- Riaz, A.; Ali, S.; Summer, M.; Noor, S.; Nazakat, L.; Aqsa; Sharjeel, M. Exploring the underlying pharmacological, immunomodulatory, and anti-inflammatory mechanisms of phytochemicals against wounds: A molecular insight. Inflammopharmacology 2024, 32, 2695–2727. [Google Scholar] [CrossRef]
- Benyoucef, F.; Dib, M.E.; Tabti, B.; Zoheir, A.; Costa, J.; Muselli, A. Synergistic effects of essential oils of Ammoides verticillata and Satureja candidissima against many pathogenic microorganisms. Anti-Infect. Agents 2020, 18, 72–78. [Google Scholar]
- Abdelli, I.; Hassani, F.; Bekkel Brikci, S.; Ghalem, S. In silico study the inhibition of angiotensin converting enzyme 2 receptor of COVID-19 by Ammoides verticillata components harvested from Western Algeria. J. Biomol. Struct. Dyn. 2021, 39, 3263–3276. [Google Scholar] [CrossRef]
- Jalloul, A.B.; Ayadi, N.; Klai, A.; Abderrabba, M. Functionalization of pasteurized milk using rosemary, thyme, and ammoides aqueous extracts for better microbial quality and an improved antioxidant activity. Molecules 2022, 27, 3725. [Google Scholar] [CrossRef]
- Ouslimani, S.; Bendahou, M.; Abedelmounaim, K.; Benbelaid, F.; Mahdi, F.; Yassine Benziane, M.; Costa, J. Antibacterial and anti-biofilm efficiency of twenty Algerian plants essential oils against resistant Acinetobacter baumannii. J. Essent. Oil Bear. Plants 2023, 26, 206–231. [Google Scholar] [CrossRef]
- Bnouham, M.; Merhfour, F.Z.; Legssyer, A.; Mekhfi, H.; Maâllem, S.; Ziyyat, A. Antihyperglycemic activity of Arbutus unedo, Ammoides pusilla and Thymelaea hirsuta. Die Pharm.-Int. J. Pharm. Sci. 2007, 62, 630–632. [Google Scholar]
- Bnouham, M.; Merhfour, F.Z.; Ziyyat, A.; Aziz, M.; Legssyer, A.; Mekhfi, H. Antidiabetic effect of some medicinal plants of Oriental Morocco in neonatal non-insulin-dependent diabetes mellitus rats. Hum. Exp. Toxicol. 2010, 29, 865–871. [Google Scholar] [CrossRef]
- Tefiani, C.; Riazi, A.; Youcefi, F.; Aazza, S.; Gago, C.; Faleiro, M.L.; Miguel, M.G. Ammoides pusilla (Apiaceae) and Thymus munbyanus (Lamiaceae) from Algeria essential oils: Chemical composition, antimicrobial, antioxidant and antiproliferative activities. J. Essent. Oil Res. 2015, 27, 131–139. [Google Scholar] [CrossRef]
- Senouci, H.; Benyelles, N.G.; Dib, M.E.; Costa, J.; Muselli, A. Ammoides verticillata essential oil as biocontrol agent of selected fungi and pest of olive tree. Recent Pat. Food Nutr. Agric. 2020, 11, 182–188. [Google Scholar] [CrossRef]
- Benziane, M.Y.; Bendahou, M.; Benbelaïd, F.; Khadir, A.; Belhadef, H.; Benaissa, A.; Muselli, A. Efficacy of endemic Algerian essential oils against single and mixed biofilms of Candida albicans and Candida glabrata. Arch. Oral Biol. 2023, 145, 105584. [Google Scholar] [CrossRef] [PubMed]
- Belaiba, M.; Saoudi, M.M.; Abedrabba, M.; Bouajila, J. Ammoides pusilla Aerial Part: GC-MS Profiling and Evaluation of In Vitro Antioxidant and Biological Activities. Processes 2024, 12, 1274. [Google Scholar] [CrossRef]
- Bendjabeur, S.; Bensouici, C.; Hazzit, M. Investigation of chemical composition, anticholinesterase, antioxidant, antihemolytic and antibacterial activities of essential oil and ethanol extract from aerial parts of Algerian Ammoides verticillata (Brot.) Breistr. J. Essent. Oil Res. 2024, 26, 185–199. [Google Scholar] [CrossRef]
- Rebai, R.; Carmena-Bargueño, M.; Toumi, M.E.; Derardja, I.; Jasmin, L.; Pérez-Sánchez, H.; Boudah, A. Identification of potent inhibitors of kynurenine-3-monooxygenase from natural products: In silico and in vitro approaches. Heliyon 2024, 10, e30287. [Google Scholar] [CrossRef]
- Senouci, H.; Benyelles, N.G.; Dib, M.E.; Costa, J.; Muselli, A. Chemical composition and combinatory antifungal activities of Ammoides verticillata, Allium sativum and Curcuma longa essential oils against four fungi responsible for tomato diseases. Comb. Chem. High Throughput Screen. 2020, 23, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Tefiani, C.; Riazi, A.; Belbachir, B.; Lahmar, H.; Aazza, S.; Figueiredo, A.C.; Miguel, M.G. Ammoides pusilla (Brot.) Breistr. from Algeria: Effect of harvesting place and plant part (leaves and flowers) on the essential oils chemical composition and antioxidant activity. Open Chem. 2016, 14, 343–350. [Google Scholar] [CrossRef]
- Attou, A.; Davenne, D.; Benmansour, A.; Lazouni, H.A. Chemical composition and biological activities of Ammoides verticillata essential oil from west Algeria. Phytothérapie 2019, 17, 2. [Google Scholar] [CrossRef]
- Souhaiel, N.; Sifaoui, I.; Hassine, D.B.; Bleton, J.; Bonose, M.; Moussa, F.; Abderrabba, M. Ammoides pusilla (Apiaceae) essential oil: Activity against Acanthamoeba castellanii Neff. Exp. Parasitol. 2017, 183, 99–103. [Google Scholar] [CrossRef]
- Chakroun, Y.; Oueslati, S.; Atanasova, V.; Richard-Forget, F.; Abderrabba, M.; Savoie, J.M. Ammoides pusilla essential oil: A potent inhibitor of the growth of Fusarium avenaceum and its enniatin production. Molecules 2021, 26, 6906. [Google Scholar] [CrossRef]
- Platzer, M.; Kiese, S.; Herfellner, T.; Schweiggert-Weisz, U.; Miesbauer, O.; Eisner, P. Common trends and differences in antioxidant activity analysis of phenolic substances using single electron transfer based assays. Molecules 2021, 26, 1244. [Google Scholar] [CrossRef]
- Rumpf, J.; Burger, R.; Schulze, M. Statistical evaluation of DPPH, ABTS, FRAP, and Folin-Ciocalteu assays to assess the antioxidant capacity of lignins. Int. J. Biol. Macromol. 2023, 233, 123470. [Google Scholar] [CrossRef]
- Yildiz, S.; Turan, S.; Kiralan, M.; Ramadan, M.F. Antioxidant properties of thymol, carvacrol, and thymoquinone and its efficiencies on the stabilization of refined and stripped corn oils. J. Food Meas. Charact. 2021, 15, 621–632. [Google Scholar] [CrossRef]
- Fakharian, P.; Taghavi, F.; Kianmehr, Z.; Atashian, M. Inhibitory effects of thymol and carvacrol on heme degradation and oxidative products due to tartrazine: In silico and in vitro studies. Heliyon 2024, 10, e24576. [Google Scholar] [CrossRef]
- Brewer, M.S. Natural antioxidants: Sources, compounds, mechanisms of action, and potential applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, e13394. [Google Scholar] [CrossRef]
- Bello, I.; Bakkouri, A.S.; Tabana, Y.M.; Al-Hindi, B.; Al-Mansoub, M.A.; Mahmud, R.; Asmawi, M.Z. Acute and sub-acute toxicity evaluation of the methanolic extract of Alstonia scholaris stem bark. Med. Sci. 2016, 4, 4. [Google Scholar] [CrossRef]
- Dzoyem, J.P.; McGaw, L.J.; Kuete, V.; Bakowsky, U. Anti-inflammatory and anti-nociceptive activities of African medicinal spices and vegetables. In Medicinal Spices and Vegetables from Africa; Elsevier: Amsterdam, The Netherlands, 2017; pp. 239–270. [Google Scholar]
- Patil, K.R.; Mahajan, U.B.; Unger, B.S.; Goyal, S.N.; Belemkar, S.; Surana, S.J.; Patil, C.R. Animal models of inflammation for screening of anti-inflammatory drugs: Implications for the discovery and development of phytopharmaceuticals. Int. J. Mol. Sci. 2019, 20, 4367. [Google Scholar] [CrossRef]
- Vazquez, E.; Navarro, M.; Salazar, Y.; Crespo, G.; Bruges, G.; Osorio, C.; López, M. Systemic changes following carrageenan-induced paw inflammation in rats. Inflamm. Res. 2015, 64, 333–342. [Google Scholar] [CrossRef]
- Da Silva Lima, M.; Quintans-Junior, L.J.; de Santana, W.A.; Kaneto, C.M.; Soares, M.B.P.; Villarreal, C.F. Anti-inflammatory effects of carvacrol: Evidence for a key role of interleukin-10. Eur. J. Pharmacol. 2013, 699, 112–117. [Google Scholar] [CrossRef]
- Somensi, N.; Rabelo, T.K.; Guimarães, A.G.; Quintans-Junior, L.J.; de Souza Araújo, A.A.; Moreira, J.C.F.; Gelain, D.P. Carvacrol suppresses LPS-induced pro-inflammatory activation in RAW 264.7 macrophages through ERK1/2 and NF-kB pathway. Int. Immunopharmacol. 2019, 75, 105743. [Google Scholar] [CrossRef]
- Cui, Z.W.; Xie, Z.X.; Wang, B.F.; Zhong, Z.H.; Chen, X.Y.; Sun, Y.H.; Bian, L.G. Carvacrol protects neuroblastoma SH-SY5Y cells against Fe2+-induced apoptosis by suppressing activation of MAPK/JNK-NF-κB signaling pathway. Acta Pharmacol. Sin. 2015, 36, 1426–1436. [Google Scholar] [CrossRef]
- Li, Z.; Hua, C.; Pan, X.; Fu, X.; Wu, W. Carvacrol exerts neuroprotective effects via suppression of the inflammatory response in middle cerebral artery occlusion rats. Inflammation 2016, 39, 1566–1572. [Google Scholar] [CrossRef]
- Marconi, G.D.; Della Rocca, Y.; Fonticoli, L.; Guarnieri, S.; Carradori, S.; Rajan, T.S.; Diomede, F. The beneficial effect of carvacrol in HL-1 cardiomyocytes treated with LPS-G: Anti-inflammatory pathway investigations. Antioxidants 2022, 11, 386. [Google Scholar] [CrossRef]
- Xu, L.; Yang, X.; Liu, X.T.; Li, X.Y.; Zhu, H.Z.; Xie, Y.H.; Zhao, Y. Carvacrol alleviates LPS-induced myocardial dysfunction by inhibiting the TLR4/MyD88/NF-κB and NLRP3 inflammasome in cardiomyocytes. J. Inflamm. 2024, 21, 47. [Google Scholar] [CrossRef]
- Joshi, S.; Kundu, S.; Priya, V.V.; Kulhari, U.; Mugale, M.N.; Sahu, B.D. Anti-inflammatory activity of carvacrol protects the heart from lipopolysaccharide-induced cardiac dysfunction by inhibiting pyroptosis via NLRP3/Caspase1/Gasdermin D signaling axis. Life Sci. 2023, 324, 121743. [Google Scholar] [CrossRef]
- Hotta, M.; Nakata, R.; Katsukawa, M.; Hori, K.; Takahashi, S.; Inoue, H. Carvacrol, a component of thyme oil, activates PPARα and γ and suppresses COX-2 expression. J. Lipid Res. 2010, 51, 132–139. [Google Scholar] [CrossRef]
- Choi, M.J.; Lee, E.J.; Park, J.S.; Kim, S.N.; Park, E.M.; Kim, H.S. Anti-inflammatory mechanism of galangin in lipopolysaccharide-stimulated microglia: Critical role of PPAR-γ signaling pathway. Biochem. Pharmacol. 2017, 144, 120–131. [Google Scholar] [CrossRef]
- Aslam, H.; Albaqami, F.; Rehman, N.U.; Shah, F.A. Carvacrol attenuated myocardial infarction through NLRP3-mediated pyroptosis and mTOR/Nrf2/PPARγ-dependent autophagic signaling. Toxicol. Appl. Pharmacol. 2025, 498, 117281. [Google Scholar] [CrossRef]
- Braga, P.C.; Dal Sasso, M.; Culici, M.; Bianchi, T.; Bordoni, L.; Marabini, L. Anti-inflammatory activity of thymol: Inhibitory effect on the release of human neutrophil elastase. Pharmacology 2006, 77, 130–136. [Google Scholar] [CrossRef]
- Liang, D.; Li, F.; Fu, Y.; Cao, Y.; Song, X.; Wang, T.; Zhang, N. Thymol inhibits LPS-stimulated inflammatory response via down-regulation of NF-κB and MAPK signaling pathways in mouse mammary epithelial cells. Inflammation 2014, 37, 214–222. [Google Scholar] [CrossRef]
- Safarbalou, A.; Abbasi, A. Oral administration of liposome-encapsulated thymol could alleviate the inflammatory parameters in serum and hippocampus in a rat model of Alzheimer’s disease. Exp. Gerontol. 2024, 193, 112473. [Google Scholar] [CrossRef]
- Peirovy, Y.; Asle-Rousta, M. Thymol and p-Cymene Protect the Liver by Mitigating Oxidative Stress, Suppressing TNF-α/NF-κ B, and Enhancing Nrf2/HO-1 Expression in Immobilized Rats. Chem. Biol. Drug Des. 2024, 104, e14618. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, L.; Zhang, Y.; Shu, X.; Hu, Y.; Chen, D.; Xu, Y. Thymol improves ischemic brain injury by inhibiting microglia-mediated neuroinflammation. Brain Res. Bull. 2024, 215, 111029. [Google Scholar] [CrossRef]
- Dou, X.; Yan, D.; Liu, S.; Gao, L.; Shan, A. Thymol alleviates LPS-induced liver inflammation and apoptosis by inhibiting NLRP3 inflammasome activation and the AMPK-mTOR-autophagy pathway. Nutrients 2022, 14, 2809. [Google Scholar] [CrossRef]
- Wu, H.; Jiang, K.; Yin, N.; Ma, X.; Zhao, G.; Qiu, C.; Deng, G. Thymol mitigates lipopolysaccharide-induced endometritis by regulating the TLR4-and ROS-mediated NF-κB signaling pathways. Oncotarget 2017, 8, 20042. [Google Scholar] [CrossRef]
- Yu, L.; Yan, J.; Sun, Z. D-limonene exhibits anti-inflammatory and antioxidant properties in an ulcerative colitis rat model via regulation of iNOS, COX-2, PGE2 and ERK signaling pathways. Mol. Med. Rep. 2017, 15, 2339–2346. [Google Scholar] [CrossRef]
- Kathem, S.H.; Nasrawi, Y.S.; Mutlag, S.H.; Nauli, S.M. Limonene Exerts Anti-Inflammatory Effect on LPS-Induced Jejunal Injury in Mice by Inhibiting NF-κB/AP-1 Pathway. Biomolecules 2024, 14, 334. [Google Scholar] [CrossRef]
- Chen, J.; Ou, G.; Gu, W.; Shi, J.; Lyu, R.; Wu, X.; Liu, C. Role in Preventing Alcoholic Liver Disease Progression: A Comparative Study of Whole-Component Finger Citron Essential Oil and Its Major Component D-Limonene. Nutrients 2025, 17, 1255. [Google Scholar] [CrossRef]
- Chu, T.W.; Ho, C.C.; Chiu, H.P.; Hsu, Y.J.; Hung, C.T.; Sung, C.H.; Hung, C.F. d-Limonene inhibits cytokines and chemokines expression by regulating NF-kappaB and STAT in HaCat cells and DNCB-induced atopic dermatitis in BALB/c mice. Int. Immunopharmacol. 2025, 148, 114082. [Google Scholar] [CrossRef]
- De Oliveira Ramalho, T.R.; de Oliveira, M.T.P.; de Araujo Lima, A.L.; Bezerra-Santos, C.R.; Piuvezam, M.R. Gamma-terpinene modulates acute inflammatory response in mice. Planta Medica 2015, 81, 1248–1254. [Google Scholar]
- Khoshnazar, S.M.; Kazemi, M.; Amirheidari, B. Neuroprotective Effects of gamma-Terpinene in Rats with Acute Cerebral Ischemia: Modulation of Inflammation, Apoptosis, and Oxidation. Neurochem. Res. 2024, 49, 1863–1878. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Chi, G.; Jiang, L.; Soromou, L.W.; Chen, N.; Huo, M.; Feng, H. p-Cymene modulates in vitro and in vivo cytokine production by inhibiting MAPK and NF-κB activation. Inflammation 2013, 36, 529–537. [Google Scholar] [CrossRef]
- Formiga, R.; Junior, E.A.; Vasconcelos, R.C.; Guerra, G.C.B.; de Araujo, A.A.; Carvalho, T.G.; Vieira, G.C. p-Cymene and rosmarinic acid ameliorate TNBS-induced intestinal inflammation upkeeping ZO-1 and MUC-2: Role of antioxidant system and immunomodulation. Int. J. Mol. Sci 2020, 21, 5870. [Google Scholar] [CrossRef] [PubMed]
- El-Miligy, M.M.; Al-Kubeisi, A.K.; Nassra, R.A.; El-Zemity, S.R.; Hazzaa, A.A. Discovery of new thymol-3, 4-disubstituted thiazole hybrids as dual COX-2/5-LOX inhibitors with in vivo proof. J. Enzym. Inhib. Med. Chem. 2024, 39, 2309171. [Google Scholar] [CrossRef]
- De Gaetano, G.; Donati, M.B.; Cerletti, C. Prevention of thrombosis and vascular inflammation: Benefits and limitations of selective or combined COX-1, COX-2 and 5-LOX inhibitors. Trends Pharmacol. Sci. 2003, 24, 245–252. [Google Scholar] [CrossRef]
- Gudis, K.; Sakamoto, C. The role of cyclooxygenase in gastric mucosal protection. Dig. Dis. Sci. 2005, 50, S16–S23. [Google Scholar] [CrossRef]
- Mancini, J.A.; Vickers, P.J.; O’Neill, G.P.; Boily, C.; Falgueyret, J.P.; Riendeau, D. Altered sensitivity of aspirin-acetylated prostaglandin G/H synthase-2 to inhibition by nonsteroidal anti-inflammatory drugs. Mol. Pharmacol. 1997, 51, 52–60. [Google Scholar] [CrossRef]
- Rowlinson, S.W.; Kiefer, J.R.; Prusakiewicz, J.J.; Pawlitz, J.L.; Kozak, K.R.; Kalgutkar, A.S.; Marnett, L.J. A novel mechanism of cyclooxygenase-2 inhibition involving interactions with Ser-530 and Tyr-385. J. Biol. Chem. 2003, 278, 45763–45769. [Google Scholar] [CrossRef]
- Blobaum, A.L.; Marnett, L.J. Structural and functional basis of cyclooxygenase inhibition. J. Med. Chem. 2007, 50, 1425–1441. [Google Scholar] [CrossRef] [PubMed]
- Rouzer, C.A.; Marnett, L.J. Structural and chemical biology of the interaction of cyclooxygenase with substrates and non-steroidal anti-inflammatory drugs. Chem. Rev. 2020, 120, 7592–7641. [Google Scholar] [CrossRef]
- Phong, N.V.; Anh, D.T.N.; Chae, H.Y.; Yang, S.Y.; Kwon, M.J.; Min, B.S.; Kim, J.A. Anti-inflammatory activity and cytotoxicity against ovarian cancer cell lines by amide alkaloids and piperic esters isolated from Piper longum fruits: In vitro assessments and molecular docking simulation. Bioorganic Chem. 2022, 128, 106072. [Google Scholar]
- Landa, P.; Kokoska, L.; Pribylova, M.; Vanek, T.; Marsik, P. In vitro anti-inflammatory activity of carvacrol: Inhibitory effect on COX-2 catalyzed prostaglandin E2 biosynthesisb. Arch. Pharmacal Res. 2009, 32, 75–78. [Google Scholar] [CrossRef]
- Marsik, P.; Kokoska, L.; Landa, P.; Nepovim, A.; Soudek, P.; Vanek, T. In vitro inhibitory effects of thymol and quinones of Nigella sativa seeds on cyclooxygenase-1-and-2-catalyzed prostaglandin E2 biosyntheses. Planta Medica 2005, 71, 739–742. [Google Scholar] [CrossRef]
- Krylova, S.G.; Povet’eva, T.N.; Zueva, E.P.; Suslov, N.I.; Amosova, E.N.; Razina, T.G.; Zhdanov, V.V. Analgesic activity of hexaazaisowurtzitane derivatives. Bull. Exp. Biol. Med. 2019, 166, 461–465. [Google Scholar] [CrossRef]
- Franzotti, E.M.; Santos, C.V.F.; Rodrigues, H.M.S.L.; Mourao, R.H.V.; Andrade, M.R.; Antoniolli, A.R. Anti-inflammatory, analgesic activity and acute toxicity of Sida cordifolia L. (Malva-branca). J. Ethnopharmacol. 2000, 72, 273–277. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Nasir, A.; Shah, S.W.A.; Khalil, A.A.K.; Ahn, M.J.; Shah, S.M.M.; Nisar, M. Evaluation of antinociceptive activity of Ilex dipyrena Wall. in mice. BMC Complement. Med. Ther. 2021, 21, 184. [Google Scholar] [CrossRef]
- Kundu, P.; Debnath, S.L.; Devnath, H.S.; Saha, L.; Sadhu, S.K. Analgesic, Anti-inflammatory, Antipyretic, and In Silico Measurements of Sonneratia caseolaris (L.) Fruits from Sundarbans, Bangladesh. BioMed Res. Int. 2022, 2022, 1405821. [Google Scholar] [CrossRef]
- Guimarães, A.G.; Oliveira, G.F.; Melo, M.S.; Cavalcanti, S.C.; Antoniolli, A.R.; Bonjardim, L.R.; Quintans-Júnior, L.J. Bioassay-guided evaluation of antioxidant and antinociceptive activities of carvacrol. Basic Clin. Pharmacol. Toxicol. 2010, 107, 949–957. [Google Scholar] [CrossRef]
- Cavalcante Melo, F.H.; Rios, E.R.V.; Rocha, N.F.M.; Citó, M.D.C.D.O.; Fernandes, M.L.; de Sousa, D.P.; de Sousa, F.C.F. Antinociceptive activity of carvacrol (5-isopropyl-2-methylphenol) in mice. J. Pharm. Pharmacol. 2012, 64, 1722–1729. [Google Scholar] [CrossRef]
- Joca, H.C.; Cruz-Mendes, Y.; Oliveira-Abreu, K.; Maia-Joca, R.P.M.; Barbosa, R.; Lemos, T.L.; Leal-Cardoso, J.H. Carvacrol decreases neuronal excitability by inhibition of voltage-gated sodium channels. J. Nat. Prod. 2012, 75, 1511–1517. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.T.; Fujita, T.; Jiang, C.Y.; Kumamoto, E. Carvacrol presynaptically enhances spontaneous excitatory transmission and produces outward current in adult rat spinal substantia gelatinosa neurons. Brain Res. 2014, 1592, 44–54. [Google Scholar] [CrossRef] [PubMed]
- De Cássia da Silveira e Sá, R.; Lima, T.C.; da Nobrega, F.R.; de Brito, A.E.M.; de Sousa, D.P. Analgesic-like activity of essential oil constituents: An update. Int. J. Mol. Sci. 2017, 18, 2392. [Google Scholar] [CrossRef] [PubMed]
- Melo, F.H.C.; Moura, B.A.; de Sousa, D.P.; de Vasconcelos, S.M.M.; Macedo, D.S.; Fonteles, M.M.D.F.; de Sousa, F.C.F. Antidepressant-like effect of carvacrol (5-Isopropyl-2-methylphenol) in mice: Involvement of dopaminergic system. Fundam. Clin. Pharmacol. 2011, 25, 362–367. [Google Scholar] [CrossRef]
- Haeseler, G.; Maue, D.; Grosskreutz, J.; Bufler, J.; Nentwig, B.; Piepenbrock, S.; Leuwer, M. Voltage-dependent block of neuronal and skeletal muscle sodium channels by thymol and menthol. Eur. J. Anaesthesiol. 2002, 19, 571–579. [Google Scholar] [CrossRef]
- Barbosa, R.; Cruz-Mendes, Y.; Silva-Alves, K.S.; Ferreira-da-Silva, F.W.; Ribeiro, N.M.; Morais, L.P.; Leal-Cardoso, J.H. Effects of Lippia sidoides essential oil, thymol, p-cymene, myrcene and caryophyllene on rat sciatic nerve excitability. Braz. J. Med. Biol. Res. 2017, 50, e6351. [Google Scholar] [CrossRef]
- Piccinelli, A.C.; Santos, J.A.; Konkiewitz, E.C.; Oesterreich, S.A.; Formagio, A.S.N.; Croda, J.; Kassuya, C.A.L. Antihyperalgesic and antidepressive actions of (R)-(+)-limonene, α-phellandrene, and essential oil from Schinus terebinthifolius fruits in a neuropathic pain model. Nutr. Neurosci. 2015, 18, 217–224. [Google Scholar] [CrossRef]
- Pereira, E.W.; Heimfarth, L.; Santos, T.K.; Passos, F.R.; Siqueira-Lima, P.; Scotti, L.; Quintans, J.S. Limonene, a citrus monoterpene, non-complexed and complexed with hydroxypropyl-β-cyclodextrin attenuates acute and chronic orofacial nociception in rodents: Evidence for involvement of the PKA and PKC pathway. Phytomedicine 2022, 96, 153893. [Google Scholar] [CrossRef]
- Kaimoto, T.; Hatakeyama, Y.; Takahashi, K.; Imagawa, T.; Tominaga, M.; Ohta, T. Involvement of transient receptor potential A1 channel in algesic and analgesic actions of the organic compound limonene. Eur. J. Pain 2016, 20, 1155–1165. [Google Scholar] [CrossRef]
- Araújo-Filho, H.G.; Pereira, E.W.; Rezende, M.M.; Menezes, P.P.; Araújo, A.A.; Barreto, R.S.; Quintans, J.S. D-limonene exhibits superior antihyperalgesic effects in a β-cyclodextrin-complexed form in chronic musculoskeletal pain reducing Fos protein expression on spinal cord in mice. Neuroscience 2017, 358, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Bonjardim, L.R.; Cunha, E.S.; Guimarães, A.G.; Santana, M.F.; Oliveira, M.G.; Serafini, M.R.; Quintans-Júnior, L.J. Evaluation of the anti-inflammatory and antinociceptive properties of p-cymene in mice. Z. Für Naturforschung C 2012, 67, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Quintans, J.D.S.S.; Menezes, P.P.; Santos, M.R.V.; Bonjardim, L.R.; Almeida, J.R.G.S.; Gelain, D.P.; Quintans-Júnior, L.J. Improvement of p-cymene antinociceptive and anti-inflammatory effects by inclusion in β-cyclodextrin. Phytomedicine 2013, 20, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Quintans-Júnior, L.; Moreira, J.C.; Pasquali, M.A.; Rabie, S.M.; Pires, A.S.; Schröder, R.; Gelain, D.P. Antinociceptive activity and redox profile of the monoterpenes (+)-camphene, p-cymene, and geranyl acetate in experimental models. Int. Sch. Res. Not. 2013, 2013, 459530. [Google Scholar] [CrossRef]
- De Santana, M.F.; Guimarães, A.G.; Chaves, D.O.; Silva, J.C.; Bonjardim, L.R.; Lucca Júnior, W.D.; Quintans-Júnior, L.J. The anti-hyperalgesic and anti-inflammatory profiles of p-cymene: Evidence for the involvement of opioid system and cytokines. Pharm. Biol. 2015, 53, 1583–1590. [Google Scholar] [CrossRef]
- Santana, M.F.; Quintans-Júnior, L.J.; Cavalcanti, S.C.; Oliveira, M.G.; Guimarães, A.G.; Cunha, E.S.; Bonjardim, L.R. p-Cymene reduces orofacial nociceptive response in mice. Rev. Bras. Farmacogn. 2011, 21, 1138–1143. [Google Scholar] [CrossRef]
- Santos, W.B.; Melo, M.A.; Alves, R.S.; de Brito, R.G.; Rabelo, T.K.; Prado, L.D.S.; Guimaraes, A.G. p-Cymene attenuates cancer pain via inhibitory pathways and modulation of calcium currents. Phytomedicine 2019, 61, 152836. [Google Scholar] [CrossRef]
- Passos, F.F.D.B.; Lopes, E.M.; de Araújo, J.M.; de Sousa, D.P.; Veras, L.M.C.; Leite, J.R.S.; Almeida, F.R.D.C. Involvement of Cholinergic and Opioid System in γ-Terpinene-Mediated Antinociception. Evid.-Based Complement. Altern. Med. 2015, 2015, 829414. [Google Scholar] [CrossRef]
- Pina, L.T.; Rabelo, T.K.; Trindade, G.G.; Almeida, I.K.; Oliveira, M.A.; Dos Santos, P.L.; Guimarães, A.G. γ-Terpinene complexed with β-cyclodextrin attenuates spinal neuroactivity in animals with cancer pain by Ca2+ channel block. J. Pharm. Pharmacol. 2022, 74, 1629–1639. [Google Scholar] [CrossRef]
- Pharmacopée, E. 5ème Edition-Direction Européenne de la Qualité du Médicament; Council of Europe: Strasbourg, France, 2005. [Google Scholar]
- NIST/EPA/NIH Mass Spectral Library, PC version 1.7; National Institute of Standards and Technology Standard Reference Data: Gaithersburg, MD, USA, 1999.
- Konig, W.A.; Joulain, D.; Hochmuth, D. Terpenoids and Related Constituents of Essential Oils; Library of Mass Finder: Hamburg, Germany, 2004.
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatoghraphy/Mass Spectrometry; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Hazzit, M.; Baaliouamer, A.; Veríssimo, A.R.; Faleiro, M.L.; Miguel, M.G. Chemical composition and biological activities of Algerian Thymus oils. Food Chem. 2009, 116, 714–721. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Oyaizu, M. Studies on products of browning reaction antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef] [PubMed]
- OECD. Guideline for Testing of Chemicals. Acute Oral Toxicity; Organisation for Economic Co-Operation and Development: Paris, France, 2001. [Google Scholar]
- Da Fonseca, A.G.; Fernandes Ribeiro Dantas, L.L.D.S.; Rodrigues, J.P.; Alencar Filho, M.P.D.C.; De Melo Rêgo, M.J.B.; Da Rocha Pitta, M.G.; Moura Lemos, T.M.A. PA-Int5: An isatin-thiosemicarbazone derivative that exhibits anti-nociceptive and anti-inflammatory effects in Swiss mice. Biomed. Rep. 2021, 15, 61. [Google Scholar] [CrossRef]
- Aziz, M.A.; Mehedi, M.; Akter, M.I.; Sajon, S.R.; Mazumder, K.; Rana, M.S. In vivo and in silico evaluation of analgesic activity of Lippia alba. Clin. Phytoscience 2019, 5, 38. [Google Scholar] [CrossRef]
- Tabassum, S.; Ahmad, S.; Ali, B.; Usman, F.; Jabeen, Q.; Sajid-ur-Rehman, M.; Basit, A. Chemical profiling and evaluation of toxicological, antioxidant, anti-inflammatory, anti-nociceptive and Tyrosinase inhibitory potential of Portulacariaafra using in-vitro, in-vivo and in-silico studies. Arab. J. Chem. 2023, 16, 104784. [Google Scholar] [CrossRef]
- Bošković, J.; Dobričić, V.; Mihajlović, M.; Kotur-Stevuljević, J.; Čudina, O. Synthesis, evaluation of enzyme inhibition and redox properties of potential dual COX-2 and 5-LOX inhibitors. Pharmaceuticals 2023, 16, 549. [Google Scholar] [CrossRef]
- Ahmad, S.; Bhanu, P.; Kumar, J.; Pathak, R.K.; Mallick, D.; Uttarkar, A.; Mishra, V. Molecular dynamics simulation and docking analysis of NF-κB protein binding with sulindac acid. Bioinformation 2022, 18, 170. [Google Scholar] [CrossRef]
- Orlando, B.J.; Malkowski, M.G. Crystal structure of rofecoxib bound to human cyclooxygenase-2. Struct. Biol. Cryst. Commun. 2016, 72, 772–776. [Google Scholar] [CrossRef]
- Derardja, I.; Rebai, R.; Toumi, M.E.; Kebaili, F.F.; Boudah, A. Identification of New Potential Cyclooxygenase-2 Inhibitors Using Structure-Based Virtual Screening, Molecular Dynamics and Pharmacokinetic Modelling. Biol. Life Sci. Forum 2024, 35, 6. [Google Scholar] [CrossRef]
- Alzain, A.A.; Elbadwi, F.A.; Al-Karmalawy, A.A.; Elhag, R.; Osman, W.; Mothana, R.A. Utilization of computational methods for the identification of new natural inhibitors of human neutrophil elastase in inflammation therapy. Open Chem. 2023, 21, 20230161. [Google Scholar] [CrossRef]
- Kim, J.; Jin, H.; Kim, J.; Cho, S.Y.; Moon, S.; Wang, J.; No, K.T. Leveraging the Fragment Molecular Orbital and MM-GBSA Methods in Virtual Screening for the Discovery of Novel Non-Covalent Inhibitors Targeting the TEAD Lipid Binding Pocket. Int. J. Mol. Sci. 2024, 25, 5358. [Google Scholar] [CrossRef] [PubMed]
- Rebai, R.; Jasmin, L.; Boudah, A. Identification of two flavonoids as new and safe inhibitors of kynurenine aminotransferase II via computational and in vitro Study. Pharmaceuticals 2025, 18, 76. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Component | nRI a | nRI b | pRI c | % |
|---|---|---|---|---|---|
| 1 | alpha-Thujene | 925 | 922 | 1023 | 0.26 |
| 2 | alpha–Pinene | 933 | 931 | 1022 | 0.77 |
| 3 | beta–Pinene | 973 | 970 | 1110 | 0.51 |
| 4 | Myrcene | 982 | 979 | 1159 | 0.89 |
| 5 | para-Cymene | 1015 | 1011 | 1268 | 15.45 |
| 6 | Limonene | 1024 | 1020 | 1199 | 15.22 |
| 7 | gamma-Terpinene | 1051 | 1047 | 1243 | 17.59 |
| 8 | Linalool | 1081 | 1080 | 1544 | 0.22 |
| 9 | (E)–Sabinene hydrate | 1085 | 1083 | 1541 | 0.23 |
| 10 | Camphor | 1126 | 1123 | 1517 | 0.14 |
| 11 | Iso-Menthone | 1143 | 1142 | 1658 | 0.12 |
| 12 | Borneol | 1153 | 1148 | 1698 | 0.4 |
| 13 | Terpinen–4–ol | 1164 | 1161 | 1600 | 0.68 |
| 14 | alpha–Terpineol | 1179 | 1179 | 1700 | 0.1 |
| 15 | Thymyl methyl oxide | 1215 | 1215 | 1586 | 0.32 |
| 16 | Carvacrol methyl ether | 1226 | 1231 | 1603 | 0.24 |
| 17 | Thymol | 1270 | 1266 | 2189 | 12.49 |
| 18 | Carvacrol | 1283 | 1278 | 2219 | 32.51 |
| Monoterpene hydrocarbons (%) | 49.92 | ||||
| Oxygenated monoterpenes (%) | 46.66 | ||||
| Total identified (%) | 96.58 | ||||
| DPPH | ABTS | FRAP | TAC | |
|---|---|---|---|---|
| IC50 (μg/mL) | µg AAE/mgEO | |||
| AVEO | 83.11 ± 0.34 bc | 3.52 ± 0.02 bc | 57.81 ± 1.88 bc | 7.689 ± 0.082 |
| Ascorbic acid | 3.45 ± 0.16 a | 7.43 ± 0.29 ac | 24.62 ± 1.20 ac | - |
| α-Tocopherol | 4.36 ± 0.68 a | 9.59 ± 0.08 ab | 37.39 ± 2.14 ac | 75.640 ± 0.075 |
| Sample | IC50 Data (μg/mL) | ||
|---|---|---|---|
| COX-1 | COX-2 | SI | |
| AVEO | 8.39 ± 0.50 bc | 1.51 ± 0.20 c | 5.56 |
| Diclofenac sodium | 4.02 ± 0.60 a | 1.06 ± 0.38 c | 3.79 |
| Celecoxib | 3.30 ± 0.47 a | 0.17 ± 0.04 ab | 19.41 |
| Compound | PubChem CID | Docking Score (Kcal/mol) | Glide Score (Kcal/mol) | MMGBSA (Kcal/mol) |
|---|---|---|---|---|
| Diclofenac (standard drug) | 3033 | −7.837 | −7.837 | −34.62 |
| Carvacrol | 10,364 | −7.381 | −7.381 | −51.50 |
| Thymol | 6989 | −6.939 | −6.939 | −42.35 |
| gamma-Terpinene | 7461 | −6.370 | −6.370 | −61.64 |
| para-Cymene | 7463 | −6.255 | −6.255 | −46.95 |
| Limonene | 22,311 | −5.666 | −5.666 | −53.05 |
| Protein-Ligand Interactions | Hydrogen Bonds (Distance) | Hydrophobic Bonds | Polar Interactions | Charged (+) Interactions | Pi-Pi Stacking |
|---|---|---|---|---|---|
| Interacting Residues | |||||
| 5 KIR-Diclofenac | - | Tyr355, Leu352, Val349, Tyr348, Phe518, Val344, Leu534, Leu531, Ala527, Val523, Met522, Phe381, Leu384, Tyr385, Trp387 | His90, Ser353, Ser530 | Arg120, Arg513 | Tyr355 |
| 5KIR-Carvacrol | Ser530 (1.80 Å) | Leu352, Phe518, Val349, Tyr348, Ala527, Val523, Met522, Phe381, Leu384, Tyr385, Trp387 | Ser353, Ser530 | - | Phe518 |
| 5KIR-Thymol | Ser530 (1.93 Å) | Val349, Leu352, Phe518, Met522, Val523, Ala527, Phe381, Leu384, Tyr385, Trp387 | Ser353, Ser530 | - | Trp387 |
| 5KIR-gamma-Terpinene | - | Leu352, Phe518, Val349, Tyr348, Ala527, Val523, Met522, Phe381, Leu384, Tyr385, Trp387 | Ser353, Ser530 | - | - |
| 5KIR-para- Cymene | - | Val349, Leu352, Phe518, Met522, Val523, Ala527, Phe381, Leu384, Tyr385, Trp387 | Ser353, Ser530 | - | - |
| 5KIR-Limonene | - | Val349, Leu352, Phe518, Met522, Val523, Ala527, Phe529, Trp387, Tyr385, Leu384, Phe381 | Ser353, Ser530 | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derardja, I.; Rebai, R.; Benbelaïd, F.; Jasmin, L.; Boudah, A.; Toumi, M.E.; Mebarki, S.; Kebaili, F.F.; Bellebcir, L.; Muselli, A. Chemical Profiling and Assessment of Analgesic and Anti-Inflammatory Activity of Ammoides verticillata Essential Oil: In Vitro, In Vivo, and In Silico Studies. Pharmaceuticals 2025, 18, 635. https://doi.org/10.3390/ph18050635
Derardja I, Rebai R, Benbelaïd F, Jasmin L, Boudah A, Toumi ME, Mebarki S, Kebaili FF, Bellebcir L, Muselli A. Chemical Profiling and Assessment of Analgesic and Anti-Inflammatory Activity of Ammoides verticillata Essential Oil: In Vitro, In Vivo, and In Silico Studies. Pharmaceuticals. 2025; 18(5):635. https://doi.org/10.3390/ph18050635
Chicago/Turabian StyleDerardja, Imene, Redouane Rebai, Fethi Benbelaïd, Luc Jasmin, Abdennacer Boudah, Mohammed Esseddik Toumi, Salsabil Mebarki, Fethi Farouk Kebaili, Leila Bellebcir, and Alain Muselli. 2025. "Chemical Profiling and Assessment of Analgesic and Anti-Inflammatory Activity of Ammoides verticillata Essential Oil: In Vitro, In Vivo, and In Silico Studies" Pharmaceuticals 18, no. 5: 635. https://doi.org/10.3390/ph18050635
APA StyleDerardja, I., Rebai, R., Benbelaïd, F., Jasmin, L., Boudah, A., Toumi, M. E., Mebarki, S., Kebaili, F. F., Bellebcir, L., & Muselli, A. (2025). Chemical Profiling and Assessment of Analgesic and Anti-Inflammatory Activity of Ammoides verticillata Essential Oil: In Vitro, In Vivo, and In Silico Studies. Pharmaceuticals, 18(5), 635. https://doi.org/10.3390/ph18050635