
Alternaria alternata (Fr) Keissl Crude Extract Inhibits HIV Subtypes and Integrase Drug-Resistant Strains at Different Stages of HIV Replication

 ,
,  and
and 
Abstract
1. Introduction
2. Results
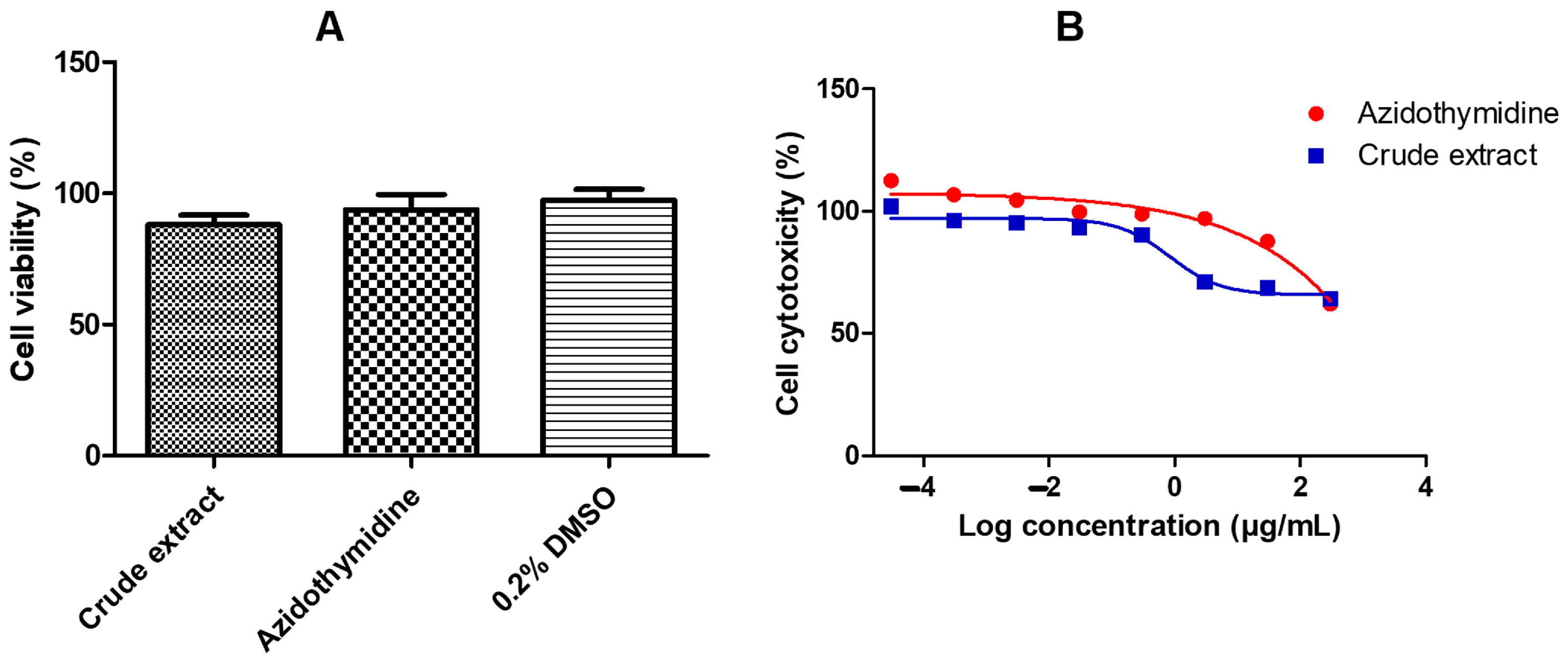
2.1. Cytotoxicity and Cell Viability of A. alternata CE Using MTT
2.2. Antiviral Activity of A. alternata Crude Extract Against Different HIV-1 Subtypes
2.3. Analysis of Half-Maximal Inhibitory Concentrations and Selective Indexes of A. alternata Crude Extract Against HIV-1 Subtypes A, B, C, and D Viruses
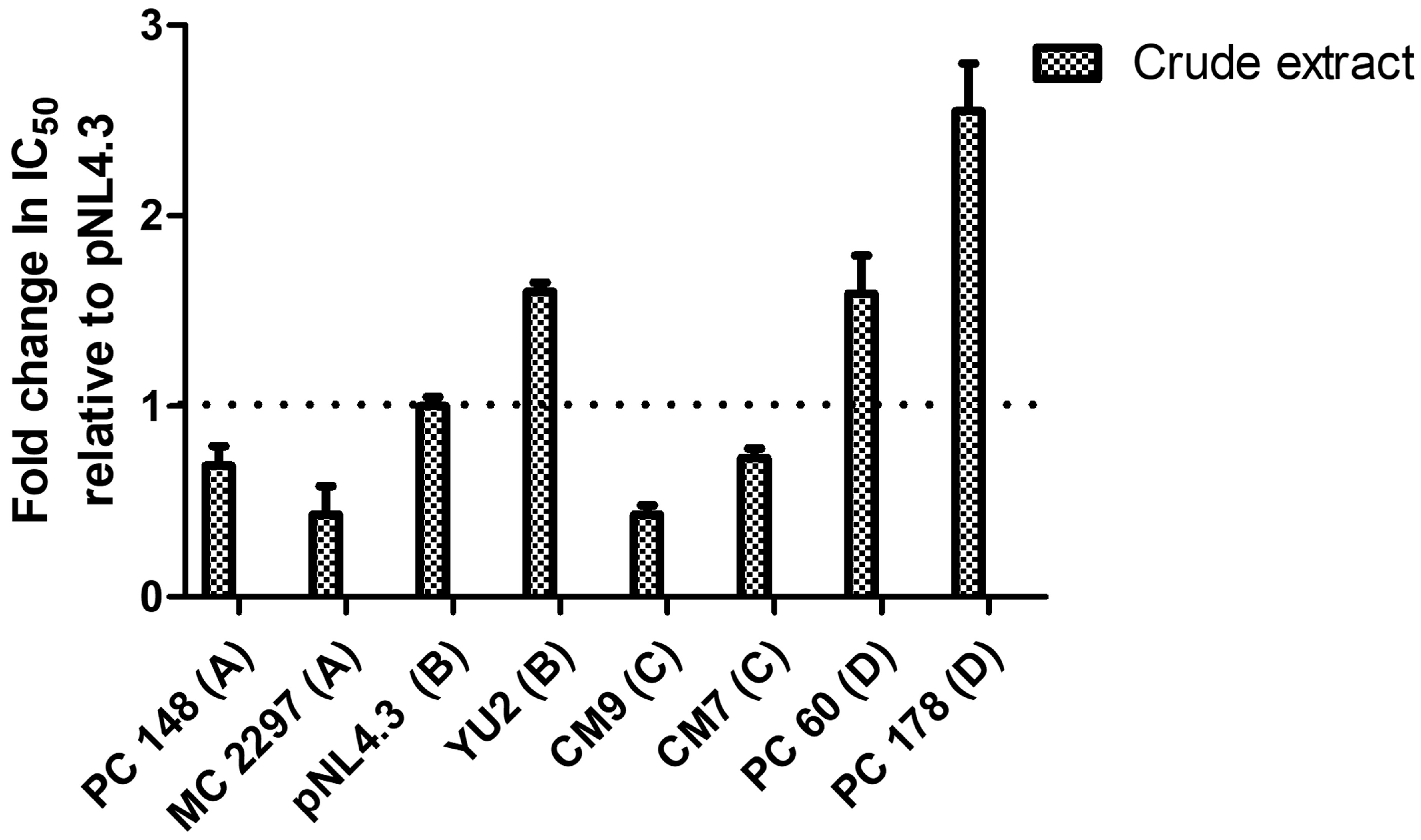
2.4. Relative Fold Change in IC50s of A. alternata Crude Extract Against Different HIV-1 Subtypes
2.5. Confirmation of Mutations Generated by Site-Directed Mutagenesis
2.6. Infectivity of HIV-1 Subtypes and Integrase Drug-Resistant Strains T66K, E92Q, S230R, and R263K
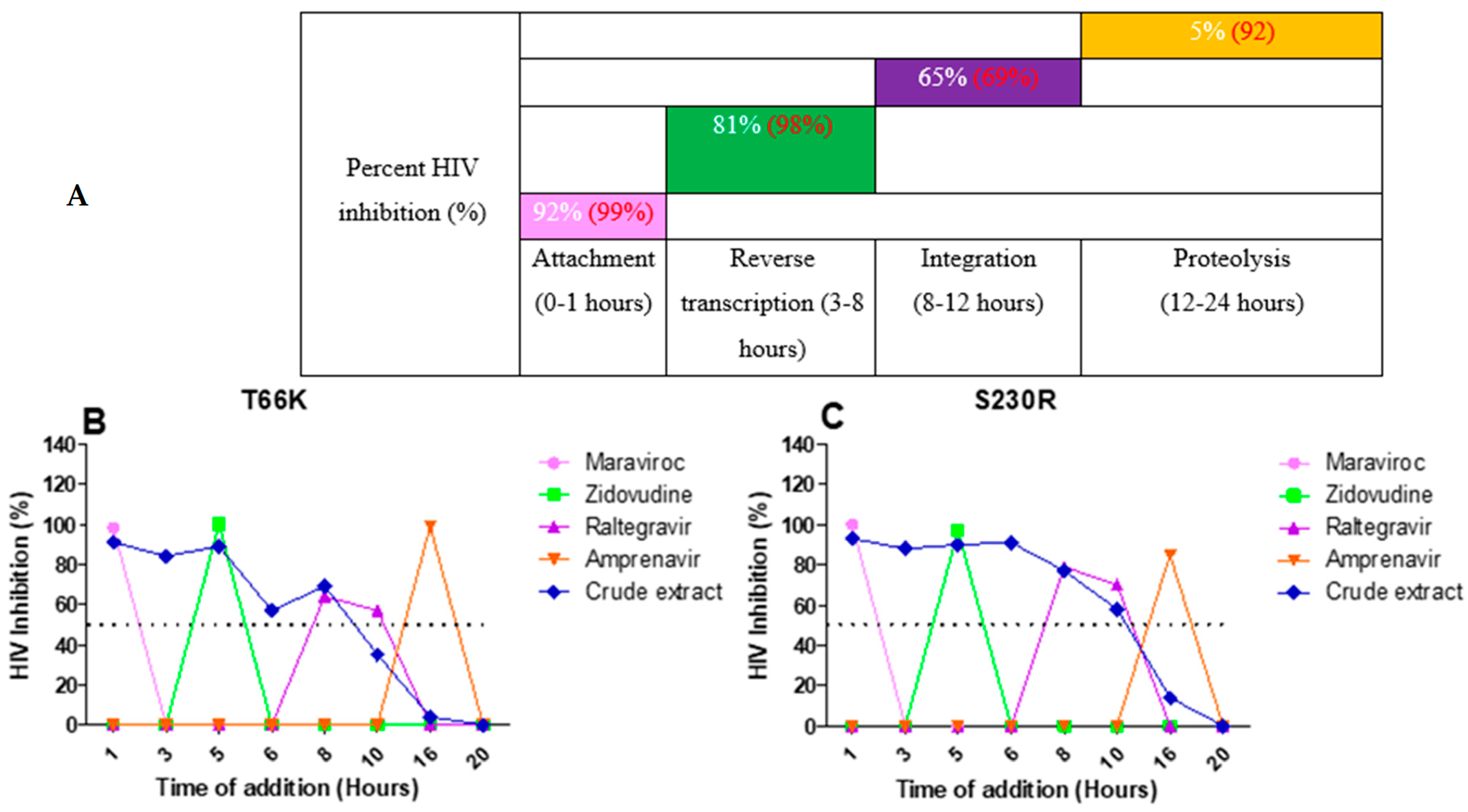
2.7. Antiviral Activity of A. alternata Crude Extract Against Integrase Drug-Resistant Mutants
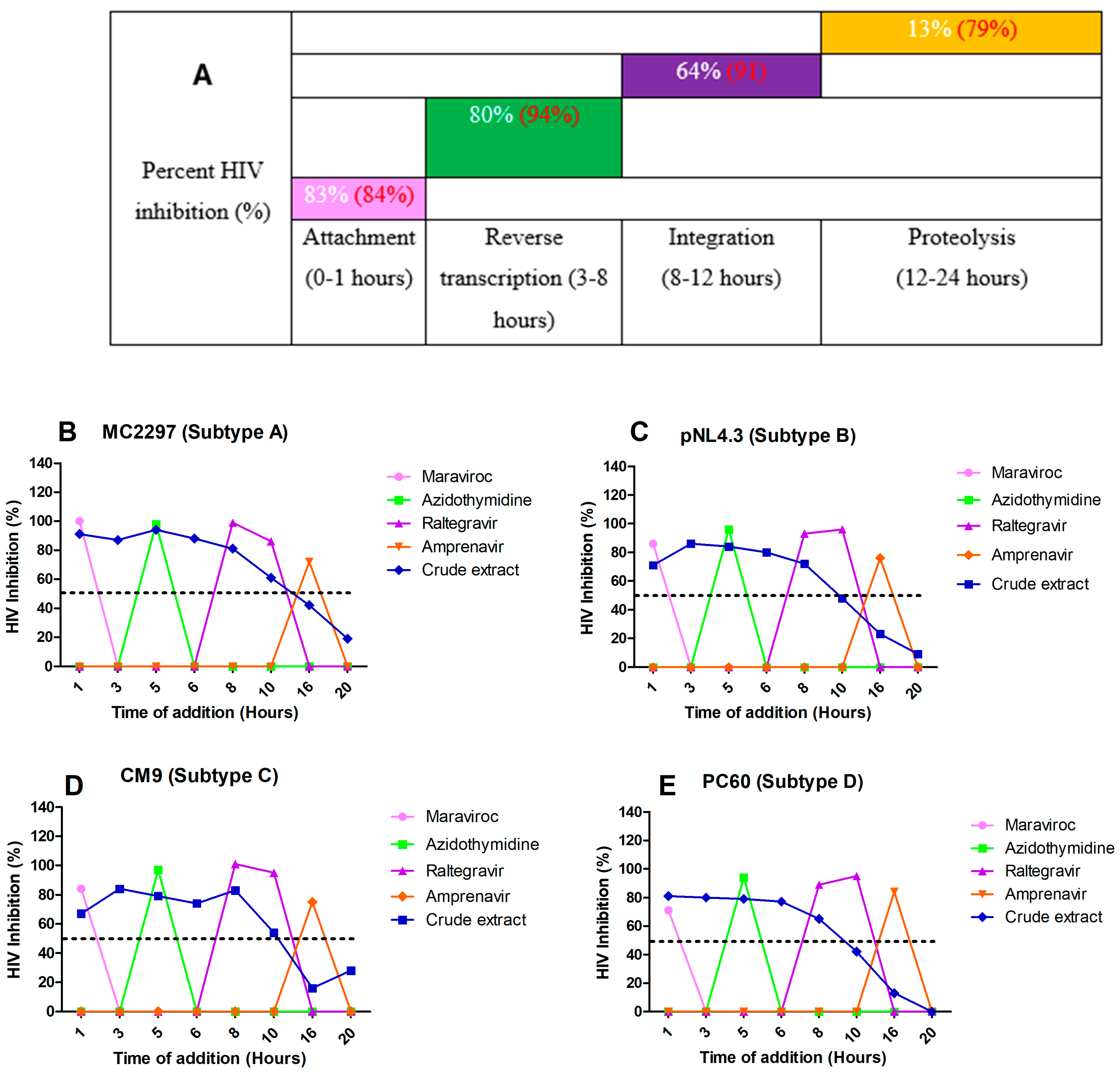
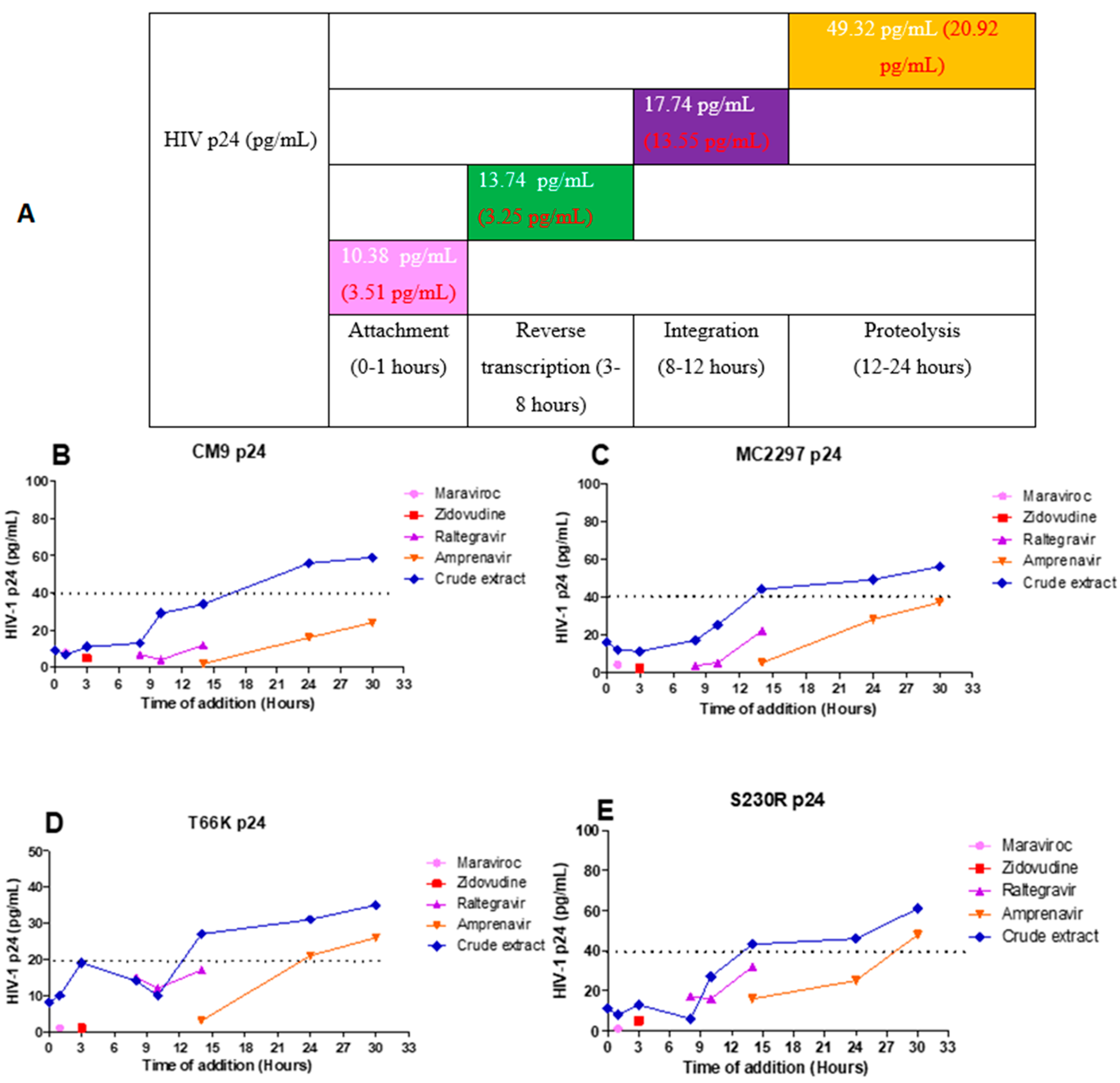
2.8. Luciferase-Based Time of Addition
2.9. HIV-1 p24-Based Time-of-Addition Assay
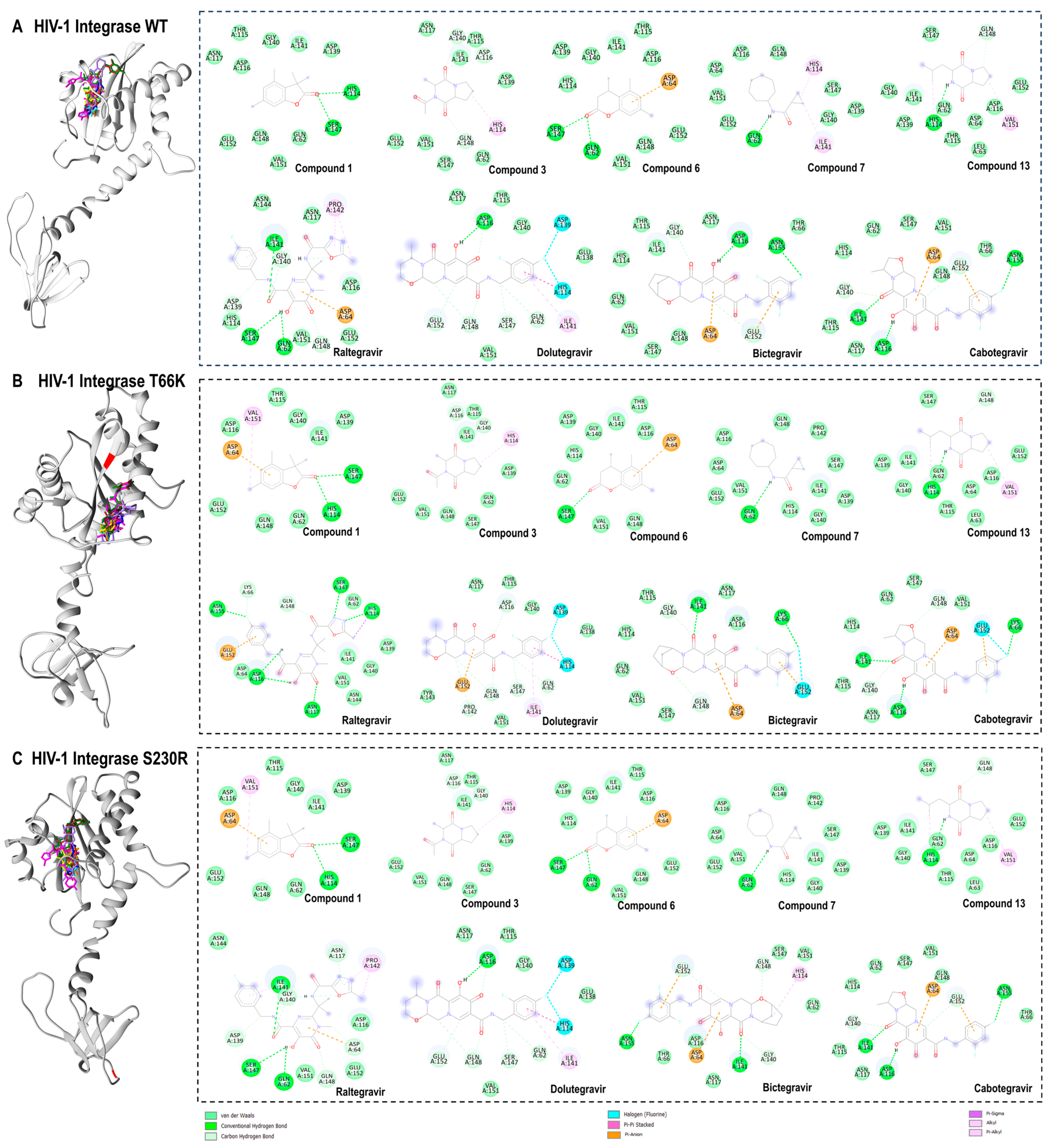
2.10. Docking Calculations of the Bioactive Compounds of A. alternata Against HIV-1 Integrase Strains (Wild Type, T66K, and S230R)
2.11. Protein–Ligand Interaction Framework Analyses
3. Discussion
4. Materials and Methods
4.1. Extraction of Crude Alternaria alternata Secondary Metabolites
4.2. Maintenance of Cell Lines
4.3. Cytotoxicity and Cell Viability Testing of A. alternata Crude Extract Using MTT Assay (3-[4,5-Dimethylthiazol-2-yl]-2,5 Diphenyl Tetrazolium Bromide)
4.4. Plasmid DNA Preparation and Purification
4.5. Site-Directed Mutagenesis
4.5.1. Design of Site-Directed Mutagenesis Primers
4.5.2. Site-Directed Mutagenesis PCR Reaction
4.6. Sequencing
4.6.1. Integrase PCR Amplification
4.6.2. Cycle Sequencing Reaction
4.7. Generation of Different HIV-1 Subtypes by Transfection
4.8. Titration of HIV-1 Viruses
4.9. Luciferase-Based Antiviral Assay
4.10. Time-of-Addition Assay-Based Inhibitory Mechanism in the HIV-1 Life Cycle
4.10.1. Luciferase-Based Time-of-Addition Assay
4.10.2. HIV-1 p24 Time-Based ELISA to Measure p24 Titre
4.11. Computational Methodology
4.11.1. Preparation and Validation of Protein Structures
4.11.2. Preparation of Ligand Structures
4.11.3. Molecular Docking Calculations
4.11.4. Protein–Ligand Interaction Assessment
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UNAIDS, R. Global AIDS report. Geneva Switzerland. 2024. Available online: www.unaids.org/en/resources/documents/2024/global-aids-update-2024 (accessed on 21 August 2024).
- Maeda, K.; Das, D.; Kobayakawa, T.; Tamamura, H.; Takeuchi, H. Discovery and Development of Anti-HIV Therapeutic Agents: Progress Towards Improved HIV Medication. Curr. Top. Med. Chem. 2019, 19, 1621–1649. [Google Scholar] [CrossRef] [PubMed]
- Phanuphak, N.; Gulick, R.M. HIV treatment and prevention 2019: Current standards of care. Curr. Opin. HIV AIDS 2020, 15, 4–12. [Google Scholar] [CrossRef]
- Patiño-Galindo, J.Á.; González-Candelas, F. The substitution rate of HIV-1 subtypes: A genomic approach. Virus Evol. 2017, 3, vex029. [Google Scholar] [CrossRef]
- Langs-Barlow, A.; Paintsil, E. Impact of human immunodeficiency virus type-1 sequence diversity on antiretroviral therapy outcomes. Viruses 2014, 6, 3855–3872. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, A.B.; Usman, M.; Kandi, V. Current scenario of HIV/AIDS, treatment options, and major challenges with compliance to antiretroviral therapy. Cureus 2016, 8, e515. [Google Scholar] [CrossRef] [PubMed]
- Hamers, R.L.; de Wit, T.F.R.; Holmes, C.B. HIV drug resistance in low-income and middle-income countries. Lancet HIV 2018, 5, e588–e596. [Google Scholar] [CrossRef]
- Lessells, R.; Katzenstein, D.; De Oliveira, T. Are subtype differences important in HIV drug resistance? Curr. Opin. Virol. 2012, 2, 636–643. [Google Scholar] [CrossRef]
- Chesney, M.A. Factors affecting adherence to antiretroviral therapy. Clin. Infect. Dis. 2000, 30 (Suppl. S2), S171–S176. [Google Scholar] [CrossRef] [PubMed]
- Bourgi, K.; Jenkins, C.A.; Rebeiro, P.F.; Shepherd, B.E.; Palella, F.; Moore, R.D.; Althoff, K.N.; Gill, J.; Rabkin, C.S.; Gange, S.J.; et al. Weight gain among treatment-naïve persons with HIV starting integrase inhibitors compared to non-nucleoside reverse transcriptase inhibitors or protease inhibitors in a large observational cohort in the United States and Canada. J. Int. AIDS Soc. 2020, 23, e25484. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The traditional medicine and modern medicine from natural products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Mofokeng, M.M.; Araya, H.T.; Amoo, S.O.; Sehlola, D.; du Plooy, C.P.; Bairu, M.W.; Venter, S.; Mashela, P.W. Diversity and Conservation through Cultivation of Hypoxis in Africa—A Case Study of Hypoxis hemerocallidea. Diversity 2020, 12, 122. [Google Scholar] [CrossRef]
- Süntar, I. Importance of ethnopharmacological studies in drug discovery: Role of medicinal plants. Phytochem. Rev. 2020, 19, 1199–1209. [Google Scholar] [CrossRef]
- Albrecht, C. Hypoxoside: A putative, non-toxic prodrug for the possible treatment of certain malignancies, HIV infections, and inflammatory conditions. In Chemistry, Biological and Pharmacological Properties of African Medicinal Plants. Proceedings of the First International IOCD Symposium, Victoria Falls, Zimbabwe, 25–28 February 1996; University of Zimbabwe Press: Harare, Zimbabwe, 1996. [Google Scholar]
- Gail, H.; Tarryn, B.; Oluwaseyi, A.; Denver, D.; Oluchi, M.; Charlotte, V.K.; Joop, D.J.; Diana, G. An ethnobotanical survey of medicinal plants used by traditional health practitioners to manage HIV and its related opportunistic infections in Mpoza, Eastern Cape Province, South Africa. J. Ethnopharmacol. 2015, 171, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Ancheeva, E.; Daletos, G.; Proksch, P. Bioactive secondary metabolites from endophytic fungi. Curr. Med. Chem. 2020, 27, 1836–1854. [Google Scholar] [CrossRef] [PubMed]
- Banyal, A.; Thakur, V.; Thakur, R.; Kumar, P. Endophytic microbial diversity: A new hope for the production of novel anti-tumor and anti-HIV agents as future therapeutics. Curr. Microbiol. 2021, 78, 1699–1717. [Google Scholar] [CrossRef]
- Nzimande, B.; Kumalo, H.M.; Ndlovu, S.I.; Mkhwanazi, N.P. Secondary metabolites produced by endophytic fungi, Alternaria alternata, as potential inhibitors of the human immunodeficiency virus. Front. Genet. 2022, 13, 1077159. [Google Scholar] [CrossRef]
- Singh, S.B.; Zink, D.L.; Guan, Z.; Collado, J.; Pelaez, F.; Felock, P.J.; Hazuda, D.J. Isolation, structure, and HIV-1 integrase inhibitory activity of Xanthoviridicatin E and F, two novel fungal metabolites produced by Penicillium chrysogenum. Helv. Chim. Acta 2003, 86, 3380–3385. [Google Scholar] [CrossRef]
- Wellensiek, B.P.; Ramakrishnan, R.; Bashyal, B.P.; Eason, Y.; Gunatilaka, A.L.; Ahmad, N. Inhibition of HIV-1 replication by secondary metabolites from endophytic fungi of desert plants. Open Virol. J. 2013, 7, 72. [Google Scholar] [CrossRef] [PubMed]
- Bashyal, B.P.; Wellensiek, B.P.; Ramakrishnan, R.; Faeth, S.H.; Ahmad, N.; Gunatilaka, A.L. Altertoxins with potent anti-HIV activity from Alternaria tenuissima QUE1Se, a fungal endophyte of Quercus emoryi. Bioorganic Med. Chem. 2014, 22, 6112–6116. [Google Scholar] [CrossRef] [PubMed]
- Stack, M.E.; Prival, M.J. Mutagenicity of the Alternaria metabolites altertoxins I, II, and III. Appl. Environ. Microbiol. 1986, 52, 718–722. [Google Scholar] [CrossRef] [PubMed]
- Govindappa, M.; Hemmanur, K.C.; Nithin, S.; Poojari, C.C.; Bhat, G.; Channabasava, K. In vitro anti-HIV activity of partially purified coumarin (s) isolated from fungal endophyte, Alternaria species of Calophyllum inophyllum. Pharmacol. Pharm. 2015, 6, 321. [Google Scholar] [CrossRef]
- Khalymbadzha, I.A.; Fatykhov, R.F.; Butorin, I.I.; Sharapov, A.D.; Potapova, A.P.; Muthipeedika, N.J.; Zyryanov, G.V.; Melekhin, V.V.; Tokhtueva, M.D.; Deev, S.L.; et al. Bioinspired Pyrano [2, 3-f] chromen-8-ones: Ring C-Opened Analogues of Calanolide A: Synthesis and Anti-HIV-1 Evaluation. Biomimetics 2024, 9, 44. [Google Scholar] [CrossRef] [PubMed]
- Kubheka, M.X.; Ndlovu, S.I.; Mkhwanazi, N.P. Anti-HIV Activity and Immunomodulatory Properties of Fractionated Crude Extracts of Alternaria alternata. Microorganisms 2024, 12, 1150. [Google Scholar] [CrossRef]
- Mogatle, S.; Skinner, M.; Mills, E.; Kanfer, I. Effect of African potato (Hypoxis hemerocallidea) on the pharmacokinetics of efavirenz. S. Afr. Med. J. 2008, 98, 945–949. [Google Scholar]
- Chiu, T.K.; Davies, D.R. Structure and function of HIV-1 integrase. Curr. Top. Med. Chem. 2004, 4, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Aslantürk, Ö.S. In vitro cytotoxicity and cell viability assays: Principles, advantages, and disadvantages. Genotoxicity-A Predict. Risk Our Actual World 2018, 2, 64–80. [Google Scholar]
- Berger, E.; Breznan, D.; Stals, S.; Jasinghe, V.J.; Gonçalves, D.; Girard, D.; Faucher, S.; Vincent, R.; Thierry, A.R.; Lavigne, C. Cytotoxicity assessment, inflammatory properties, and cellular uptake of Neutraplex lipid-based nanoparticles in THP-1 monocyte-derived macrophages. Nanobiomedicine 2017, 4, 1849543517746259. [Google Scholar] [CrossRef]
- Jimoh, M.A.; Jimoh, M.O.; Bello, M.; Raimi, I.O.; Okunlola, G.O.; Mkhwanazi, N.; Laubscher, C.P. In vitro anti-HIV, cytotoxicity and nutritional analysis of Trianthema portulacastrum L. (Aizoaceae). BMC Complement. Med. Ther. 2024, 24, 35. [Google Scholar] [CrossRef]
- Moga, D.K.; Adipo, N.; Matu, E.N.; Kirira, P.G.; Moga, D.K.; Adipo, N.; Matu, E.N.; Kirira, P.G. Antioxidant and antiproliferative activity of Azadirachta indica A. Juss Extracts against cancer cell lines: An experimental study. Afr. J. Health Sci. 2021, 34, 650–656. [Google Scholar]
- Indrayanto, G.; Putra, G.S.; Suhud, F. Validation of in-vitro bioassay methods: Application in herbal drug research. Profiles Drug Subst. Excip. Relat. Methodol. 2021, 46, 273–307. [Google Scholar]
- Adu-Ampratwum, D.; Pan, Y.; Koneru, P.C.; Antwi, J.; Hoyte, A.C.; Kessl, J.; Griffin, P.R.; Kvaratskhelia, M.; Fuchs, J.R.; Larue, R.C. Identification and optimization of a novel HIV-1 integrase inhibitor. ACS Omega 2022, 7, 4482–4491. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Nakahara, K.; Seki, T.; Miki, S.; Kawauchi, S.; Suyama, A.; Wakasamorimoto, C.; Kodama, M.; Endoh, T.; Oosugi, E.; et al. Selection of diverse and clinically relevant integrase inhibitor-resistant human immunodeficiency virus type 1 mutants. Antivir. Res. 2008, 80, 213–222. [Google Scholar] [CrossRef]
- Healthcare, V. Tivicay (Dolutegravir) [Package Insert]; ViiV Healthcare: Brentford, UK, 2013. [Google Scholar]
- Pham, H.T.; Labrie, L.; Wijting, I.E.A.; Hassounah, S.; Lok, K.Y.; Portna, I.; Goring, M.E.; Han, Y.; Lungu, C.; van der Ende, M.E.; et al. The S230R integrase substitution associated with virus load rebound during dolutegravir monotherapy confers low-level resistance to integrase strand-transfer inhibitors. J. Infect. Dis. 2018, 218, 698–706. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.T.; Isaacs, R.D.; Teppler, H.; Leavitt, R.Y.; Sklar, P.; Iwamoto, M.; Wenning, L.A.; Miller, M.D.; Chen, J.; Kemp, R.; et al. Raltegravir: The first HIV-1 integrase strand transfer inhibitor in the HIV armamentarium. Ann. N. Y. Acad. Sci. 2011, 1222, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Mouscadet, J.-F.; Tchertanov, L. Raltegravir: Molecular basis of its mechanism of action. Eur. J. Med. Res. 2009, 14, 1–12. [Google Scholar] [CrossRef]
- Wei, X.; Decker, J.M.; Liu, H.; Zhang, Z.; Arani, R.B.; Kilby, J.M.; Saag, M.S.; Wu, X.; Shaw, G.M.; Kappes, J.C. Emergence of resistant human immunodeficiency virus type 1 in patients receiving fusion inhibitor (T-20) monotherapy. Antimicrob. Agents Chemother. 2002, 46, 1896–1905. [Google Scholar] [CrossRef]
- Ikeda, Y.; Takeuchi, Y.; Martin, F.; Cosset, F.-L.; Mitrophanous, K.; Collins, M. Continuous high-titer HIV-1 vector production. Nat. Biotechnol. 2003, 21, 569–572. [Google Scholar] [CrossRef]
- Shafer, R.W. Rationale and uses of a public HIV drug-resistance database. J. Infect. Dis. 2006, 194 (Suppl. S1), S51–S58. [Google Scholar] [CrossRef]
- Chrysostomou, A.C.; Topcu, C.; Stylianou, D.C.; Hezka, J.; Kostrikis, L.G. Development of a new comprehensive HIV-1 genotypic drug resistance assay for all commercially available reverse transcriptase, protease and integrase inhibitors in patients infected with group M HIV-1 strains. Infect. Genet. Evol. 2020, 81, 104243. [Google Scholar] [CrossRef]
- Farinre, O.O. Gag-Protease Driven Viral Replication Capacity Among HIV-1 Subtypes: Implications for Disease Progression, Epidemic Spread and Vaccine Design. 2020. Available online: https://researchspace.ukzn.ac.za/items/fdddda1b-47df-4243-bc99-4a036a13e0a3 (accessed on 11 July 2023).
- Modi, M.; Goel, T.; Das, T.; Malik, S.; Suri, S.; Rawat, A.K.S.; Srivastava, S.K.; Tuli, R.; Malhotra, S.; Gupta, S.K. Ellagic acid & gallic acid from Lagerstroemia speciosa L. inhibit HIV-1 infection through inhibition of HIV-1 protease & reverse transcriptase activity. Indian J. Med. Res. 2013, 137, 540–548. [Google Scholar] [PubMed]
- Lara, H.H.; Trevino, E.N.G.; De Zamacona, M.E.; Mureyko, L.; Ixtepan-Turrent, L. Luciferase Time-based, High-throughput Screening Assay for the Discovery of HIV-1 Inhibitors. J. Hum. Virol. Retrovirol. 2014, 1, 00017. [Google Scholar]
- Daelemans, D.; Pauwels, R.; De Clercq, E.; Pannecouque, C. A time-of–drug addition approach to target identification of antiviral compounds. Nat. Protoc. 2011, 6, 925–933. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.-R.; Gao, Y.-D.; Ma, C.-H.; Zhang, X.-J.; Huang, C.-G.; Huang, J.-F.; Zheng, Y.-T. Mangiferin, an anti-HIV-1 agent targeting protease and effective against resistant strains. Molecules 2011, 16, 4264–4277. [Google Scholar] [CrossRef] [PubMed]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, R.; Šali, A. Comparative protein structure modeling: Introduction and practical examples with modeller. In Protein Structure Prediction: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2000; pp. 97–129. [Google Scholar]
- Shapovalov, M.V.; Dunbrack, R.L. A smoothed backbone-dependent rotamer library for proteins derived from adaptive kernel density estimates and regressions. Structure 2011, 19, 844–858. [Google Scholar] [CrossRef] [PubMed]
- Goldgur, Y.; Craigie, R.; Cohen, G.H.; Fujiwara, T.; Yoshinaga, T.; Fujishita, T.; Sugimoto, H.; Endo, T.; Murai, H.; Davies, D.R. Structure of the HIV-1 integrase catalytic domain complexed with an inhibitor: A platform for antiviral drug design. Proc. Natl. Acad. Sci. USA 1999, 96, 13040–13043. [Google Scholar] [CrossRef] [PubMed]
- Hanwell, M.D.; Curtis, D.E.; Lonie, D.C.; Vandermeersch, T.; Zurek, E.; Hutchison, G.R. Avogadro: An advanced semantic chemical editor, visualization, and analysis platform. J. Cheminf. 2012, 4, 17. [Google Scholar] [CrossRef]
- Oduro-Kwateng, E.; Soliman, M.E. Unveiling therapeutic frontiers: DON/DRP-104 as innovative Plasma kallikrein inhibitors against carcinoma-associated hereditary angioedema shocks-a comprehensive molecular dynamics exploration. Cell Biochem. Biophys. 2024, 82, 1159–1177. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Subtype | IC50 µg/mL (Crude Extract) | IC50 µg/mL (Azidothymidine) | SI (Crude Extract) | SI (Azidothymidine) |
|---|---|---|---|---|---|
| PC 148 | A | 0.1650 ± 0.035 | 0.04025 ± 0.002 | 1818 | 9565 |
| MC 2297 | A | 0.1029 ± 0.024 | 0.01383 ± 0.001 | 2915 | 27,838 |
| pNL4.3 | B | 0.2383 ± 0.015 | 0.03377 ± 0.004 | 1259 | 11,401 |
| YU2 | B | 0.3821 ± 0.056 | 0.04226 ± 0.004 | 785 | 9110 |
| CM7 | C | 0.1739 ± 0.032 | 0.05798 ± 0.009 | 1725 | 6640 |
| CM9 | C | 0.1027 ± 0.019 | 0.09241 ± 0.007 | 2921 | 37,487 |
| PC 60 | D | 0.3796 ± 0.037 | 0.07795 ± 0.005 | 790 | 4939 |
| PC 178 | D | 0.6086 ± 0.025 | 0.08129 ± 0.003 | 493 | 4736 |
| ID | Compound Name | Docking Score (kcal/mol) | ||
|---|---|---|---|---|
| HIV-1 Integrase Wild Type | HIV-1 Integrase T66K | HIV-1 Integrase S230R | ||
| 1 | 2,3-2H-Benzofuran-2-one, 3,3,4,6-tetramethyl- | −6.8 | −6.8 | −6.8 |
| 2 | 2-Acetylcyclopentanone | −4.6 | −4.6 | −4.6 |
| 3 | 3-Methyl-1,4-diazabicyclo[4.3.0]nonan-2,5-dione, N-acetyl | −6.2 | −6.2 | −6.2 |
| 4 | Asarone | −5.7 | −5.6 | −5.6 |
| 5 | Citraconic anhydride | −4.4 | −4.4 | −4.4 |
| 6 | Coumarin, 3,4-dihydro-4,5,7-trimethyl- | −6.6 | −6.6 | −6.6 |
| 7 | Cyclopropanecarboxamide, N-cycloheptyl | −5.9 | −5.9 | −5.9 |
| 8 | Ethyl phenylpropiolate | −5.5 | −5.5 | −5.6 |
| 9 | Hexahydropyrrolo[1,2-a]pyrazine-1,4-dione | −5.7 | −5.7 | −5.7 |
| 10 | Hexamethylcyclotrisiloxane | −3.2 | −3.2 | −3.2 |
| 11 | Levoglucosenone | −4.6 | −4.6 | −4.6 |
| 12 | Octamethylcyclotetrasiloxane | −3.4 | −3.4 | −3.4 |
| 13 | Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro-3-(2-methylpropyl)- | −6.5 | −6.5 | −6.5 |
| 14 | Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro- | −5.5 | −5.5 | −5.5 |
| Raltegravir | −8.1 | −8.4 | −8.2 | |
| Dolutegravir | −7.5 | −7.6 | −7.6 | |
| Bictegravir | −8.3 | −8.5 | −8.2 | |
| Cabotegravir | −8.1 | −8.3 | −8.1 | |
| Primer Name | Sequence (5′-3′) |
|---|---|
| T66K_FWD | ATTAGATTGTAAACATTTAGAAGGAAAGA |
| T66K_REV | TGCCATATCCCTGGACTAC |
| E92Q_FWD | TATCCCAGCACAAACAGGACAAG |
| E92Q_REV | ACCTCTGCCTCTATATATCC |
| S230R_RWD | TTACAGAGACCGCAGAAATCCAC |
| S230R_REV | TAAACCCGAAAATTTTGAATTTTTG |
| R263K_FWD | AGTACCAAGGAAAAAAGTAAAAATC |
| R263K_REV | ACCTTTATGTCACTGTTATCTTGTATTAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Naidu, D.; Oduro-Kwateng, E.; Soliman, M.E.S.; Ndlovu, S.I.; Mkhwanazi, N.P. Alternaria alternata (Fr) Keissl Crude Extract Inhibits HIV Subtypes and Integrase Drug-Resistant Strains at Different Stages of HIV Replication. Pharmaceuticals 2025, 18, 189. https://doi.org/10.3390/ph18020189
Naidu D, Oduro-Kwateng E, Soliman MES, Ndlovu SI, Mkhwanazi NP. Alternaria alternata (Fr) Keissl Crude Extract Inhibits HIV Subtypes and Integrase Drug-Resistant Strains at Different Stages of HIV Replication. Pharmaceuticals. 2025; 18(2):189. https://doi.org/10.3390/ph18020189
Chicago/Turabian StyleNaidu, Darian, Ernest Oduro-Kwateng, Mahmoud E. S. Soliman, Sizwe I. Ndlovu, and Nompumelelo P. Mkhwanazi. 2025. "Alternaria alternata (Fr) Keissl Crude Extract Inhibits HIV Subtypes and Integrase Drug-Resistant Strains at Different Stages of HIV Replication" Pharmaceuticals 18, no. 2: 189. https://doi.org/10.3390/ph18020189
APA StyleNaidu, D., Oduro-Kwateng, E., Soliman, M. E. S., Ndlovu, S. I., & Mkhwanazi, N. P. (2025). Alternaria alternata (Fr) Keissl Crude Extract Inhibits HIV Subtypes and Integrase Drug-Resistant Strains at Different Stages of HIV Replication. Pharmaceuticals, 18(2), 189. https://doi.org/10.3390/ph18020189