

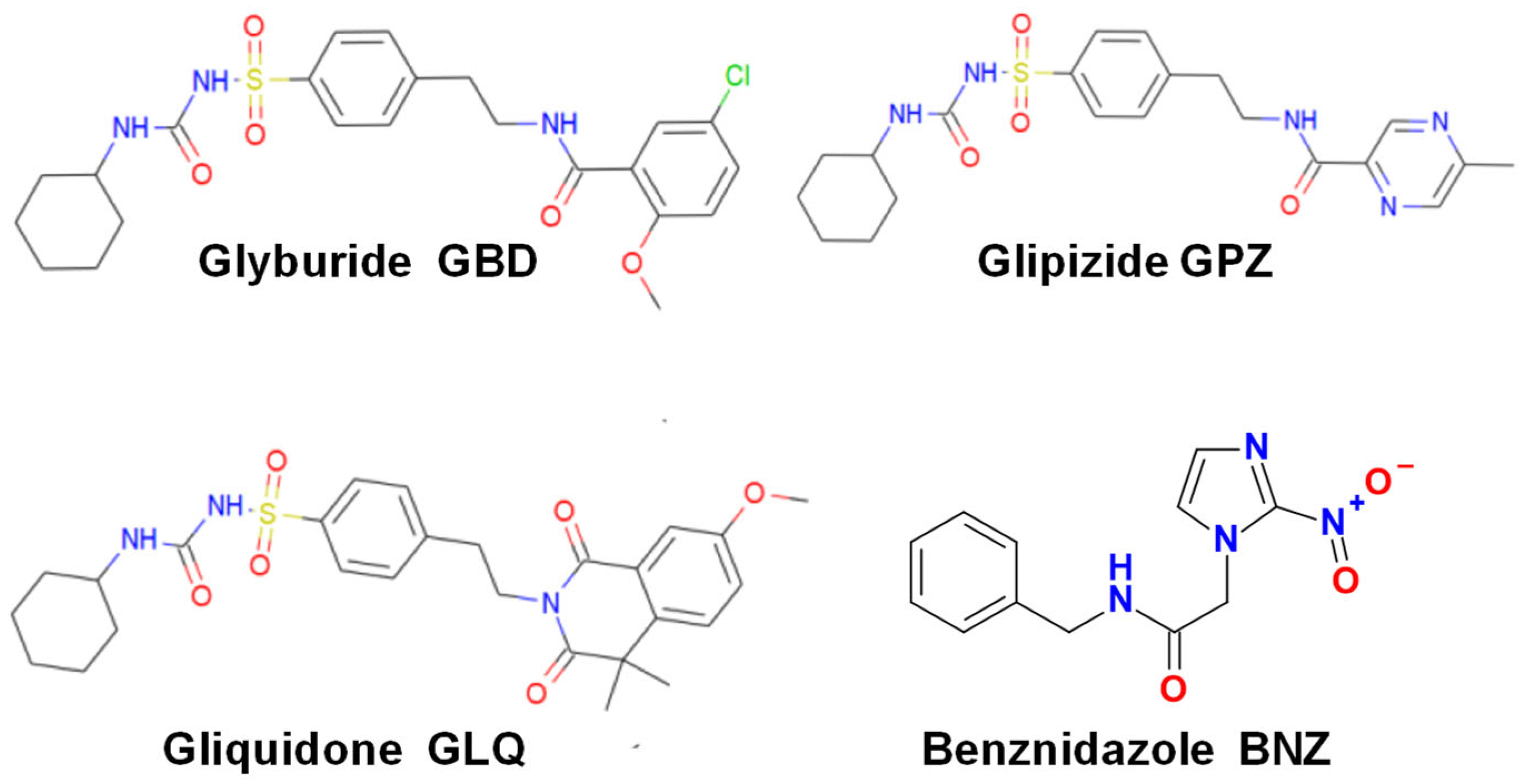
Repurposing the Antidiabetic Drugs Glyburide, Gliquidone, and Glipizide in Combination with Benznidazole for Trypanosoma cruzi Infection


 , and
, and
Abstract

1. Introduction
2. Results
2.1. Effect of BNZ and Antidiabetics in Monotherapy
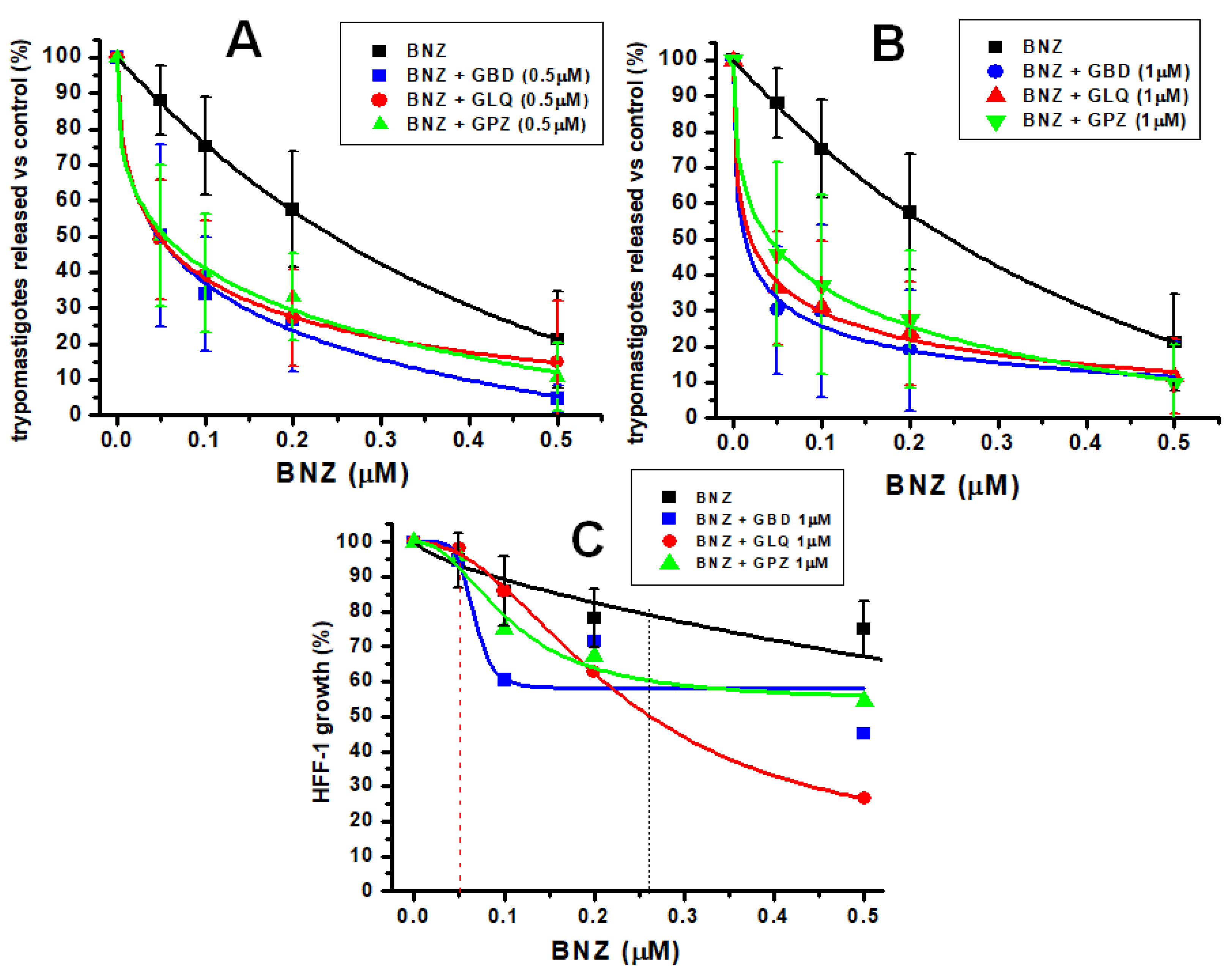
2.2. Effect of BNZ and Antidiabetics in Bi-Therapy
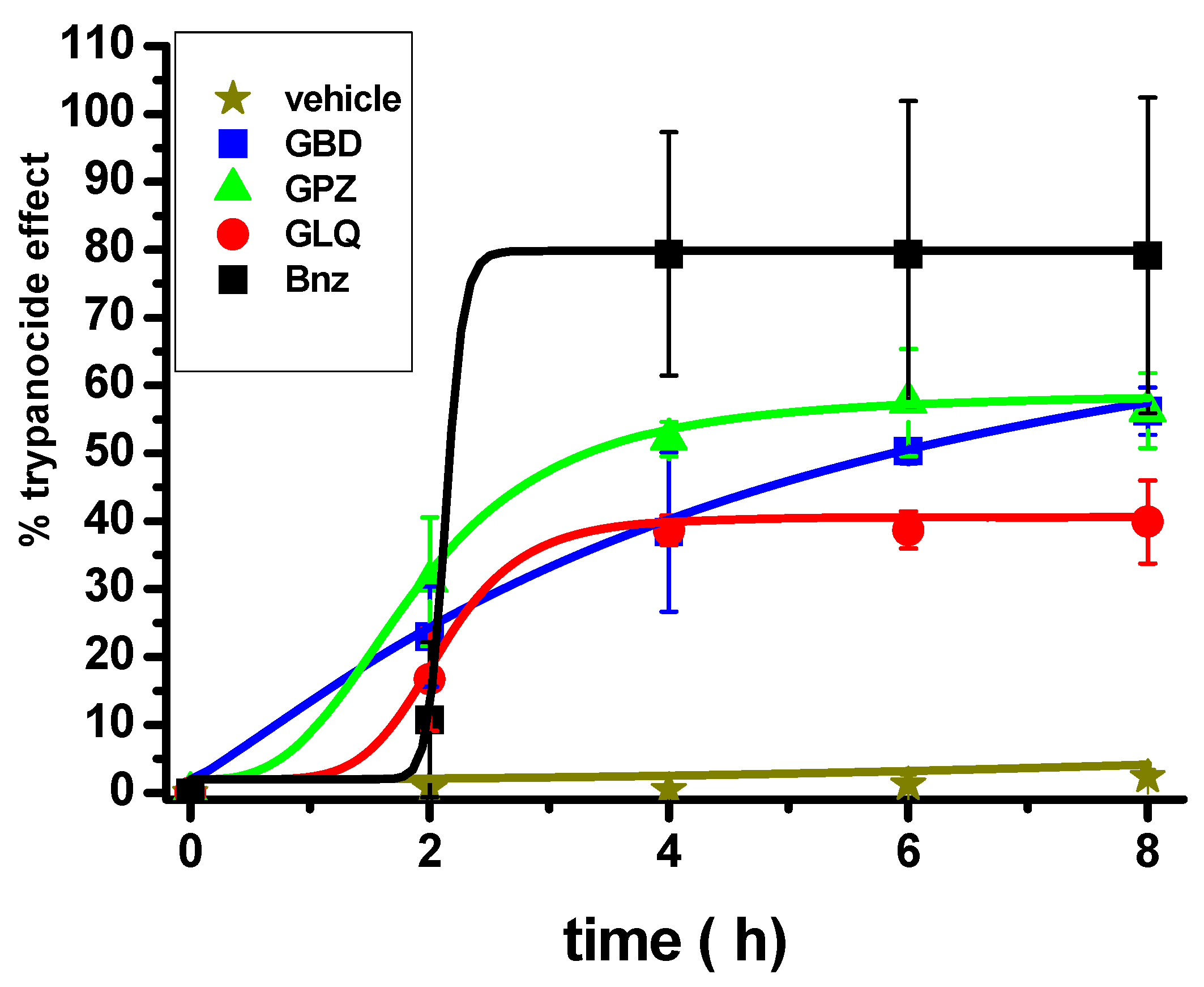
2.3. Effect of the Antidiabetic Drugs on an Acute Model of T. cruzi Infection in Mice
2.4. Effect on Thiol Metabolites
3. Discussion
3.1. Effectiveness of BNZ Alone and in Combination with Antidiabetics
3.2. Possible Mode of Action
4. Materials and Methods
4.1. Cell Culture
4.2. Trypomastigotes Obtention
4.3. Evaluation of the Mono and Bi-Therapy in the In Vitro Infection
4.4. Short-Term Antidiabetics Monotherapy in Animals Infected with T. cruzi
4.5. Effect of the Mono and Bi-Therapy on Thiol Metabolites
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Pérez-Molina, J.A.; Molina, I. Chagas Disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef] [PubMed]
- Echavarría, N.G.; Echeverría, L.E.; Stewart, M.; Gallego, C.; Saldarriaga, C. Chagas Disease: Chronic Chagas Cardiomyopathy. Curr. Probl. Cardiol. 2021, 46, 100507. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Chagas Disease (also Known as American Trypanosomiasis). 2024. Available online: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 8 October 2024).
- Coura, J.R. The main sceneries of Chagas disease transmission. The vectors, blood and oral transmissions—A comprehensive review. Mem. Inst. Oswaldo Cruz. 2015, 110, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Rassi, A.; Marcondes de Rezende, J.; Luquetti, A.O.; Rassi, A. Clinical Phases and Forms of Chagas Disease. In American Trypanosomiasis; Telleria, J., Tibayrenc, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2010; pp. 709–741. [Google Scholar] [CrossRef]
- Echeverría, L.E.; Rojas, L.Z.; Rueda-Ochoa, O.L.; Gómez-Ochoa, S.A.; González Rugeles, C.I.; Díaz, M.L.; Marcus, R.; Morillo, C.A. Circulating Trypanosoma cruzi load and major cardiovascular outcomes in patients with chronic Chagas cardiomyopathy: A prospective cohort study. Trop. Med. Int. Health 2020, 25, 1534–1541. [Google Scholar] [CrossRef]
- Mills, R.M. Chagas Disease: Epidemiology and Barriers to Treatment. Am. J. Med. 2020, 133, 1262–1265. [Google Scholar] [CrossRef] [PubMed]
- Losada Galván, I.; Alonso-Padilla, J.; Cortés-Serra, N.; Alonso-Vega, C.; Gascón, J.; Pinazo, M.J. Benznidazole for the treatment of Chagas disease. Expert Rev. Anti-Infect. Ther. 2021, 19, 547–556. [Google Scholar] [CrossRef]
- Viotti, R.; Vigliano, C.; Lococo, B.; Alvarez, M.G.; Petti, M.; Bertocchi, G.; Armenti, A. Side effects of benznidazole as treatment in chronic Chagas disease: Fears and realities. Expert Rev. Anti-Infect. Ther. 2009, 7, 157–163. [Google Scholar] [CrossRef]
- Jackson, Y.; Alirol, E.; Getaz, L.; Wolff, H.; Combescure, C.; Chappuis, F. Tolerance and safety of nifurtimox in patients with chronic Chagas disease. Clin. Infect. Dis. 2010, 51, e69–e75. [Google Scholar] [CrossRef]
- Ramos, L.G.; de Souza, K.R.; Júnior, P.A.S.; Câmara, C.C.; Castelo-Branco, F.S.; Boechat, N.; Carvalho, S.A. Tackling the challenges of human Chagas disease: A comprehensive review of treatment strategies in the chronic phase and emerging therapeutic approaches. Acta Trop. 2024, 256, 107264. [Google Scholar] [CrossRef]
- Torchelsen, F.K.V.D.S.; Mazzeti, A.L.; Mosqueira, V.C.F. Drugs in preclinical and early clinical development for the treatment of Chagas’s disease: The current status. Expert Opin. Investig. Drugs 2024, 33, 575–590. [Google Scholar] [CrossRef]
- Porta, E.O.J.; Kalesh, K.; Steel, P.G. Navigating drug repurposing for Chagas disease: Advances, challenges, and opportunities. Front. Pharmacol. 2023, 14, 1233253. [Google Scholar] [CrossRef]
- Bellera, C.L.; Alberca, L.N.; Sbaraglini, M.L.; Talevi, A. In silico Drug Repositioning for Chagas Disease. Curr. Med. Chem. 2020, 27, 662–675. [Google Scholar] [CrossRef]
- Juárez-Saldivar, A.; Schroeder, M.; Salentin, S.; Haupt, V.J.; Saavedra, E.; Vázquez, C.; Reyes-Espinosa, F.; Herrera-Mayorga, V.; Villalobos-Rocha, J.C.; García-Pérez, C.A.; et al. Computational Drug Repositioning for Chagas Disease Using Protein-Ligand Interaction Profiling. Int. J. Mol. Sci. 2020, 21, 4270. [Google Scholar] [CrossRef]
- Proks, P.; Reimann, F.; Green, N.; Gribble, F.; Ashcroft, F. Sulfonylurea stimulation of insulin secretion. Diabetes 2002, 51 (Suppl. S3), S368–S376. [Google Scholar] [CrossRef]
- Sola, D.; Rossi, L.; Schianca, G.P.; Maffioli, P.; Bigliocca, M.; Mella, R.; Corlianò, F.; Fra, G.P.; Bartoli, E.; Derosa, G. Sulfonylureas and their use in clinical practice. Arch. Med. Sci. 2015, 11, 840–848. [Google Scholar] [CrossRef]
- Lv, W.; Wang, X.; Xu, Q.; Lu, W. Mechanisms and Characteristics of Sulfonylureas and Glinides. Curr. Top. Med. Chem. 2020, 20, 37–56. [Google Scholar] [CrossRef]
- Ponte-Sucre, A.; Campos, Y.; Fernandez, M.; Moll, H.; Mendoza-León, A. Leishmania sp.: Growth and survival are impaired by ion channel blockers. Exp. Parasitol. 1998, 88, 11–19. [Google Scholar] [CrossRef]
- Serrano-Martín, X.; Payares, G.; Mendoza-León, A. Glibenclamide, a blocker of K+ATP channels, shows antileishmanial activity in experimental murine cutaneous leishmaniasis. Antimicrob. Agents Chemother. 2006, 50, 4214–4216. [Google Scholar] [CrossRef]
- Padrón-Nieves, M.; Díaz, E.; Machuca, C.; Romero, A.; Ponte, A. Glibenclamide modulates glucantime activity and disposition in Leishmania major. Exp. Parasitol. 2009, 121, 331–337. [Google Scholar] [CrossRef]
- Carvalho, A.M.; Novais, F.O.; Paixão, C.S.; de Oliveira, C.I.; Machado, P.R.L.; Carvalho, L.P.; Scott, P.; Carvalho, E.M. Glyburide, a NLRP3 Inhibitor, Decreases Inflammatory Response and Is a Candidate to Reduce Pathology in Leishmania braziliensis Infection. J. Investig. Dermatol. 2020, 140, 246–249.e2. [Google Scholar] [CrossRef]
- Rub, A.; Shaker, K.; Kashif, M.; Arish, M.; Dukhyil, A.A.B.; Alshehri, B.M.; Alaidarous, M.A.; Banawas, S.; Amir, K. Repurposing Glyburide as Antileishmanial Agent to Fight Against Leishmaniasis. Protein Pept. Lett. 2019, 26, 371–376. [Google Scholar] [CrossRef]
- Talevi, A.; Carrillo, C.; Comini, M. The Thiol-polyamine Metabolism of Trypanosoma cruzi: Molecular Targets and Drug Repurposing Strategies. Curr. Med. Chem. 2019, 26, 6614–6635. [Google Scholar] [CrossRef]
- González-Chávez, Z.; Vázquez, C.; Mejia-Tlachi, M.; Márquez-Dueñas, C.; Manning-Cela, R.; Encalada, R.; Rodríguez-Enríquez, S.; Michels, P.A.M.; Moreno-Sánchez, R.; Saavedra, E. Gamma-glutamylcysteine synthetase and tryparedoxin 1 exert high control on the antioxidant system in Trypanosoma cruzi contributing to drug resistance and infectivity. Redox Biol. 2019, 26, 101231. [Google Scholar] [CrossRef]
- Saavedra, E.; González-Chávez, Z.; Moreno-Sánchez, R.; Michels, P.A.M. Drug Target Selection for Trypanosoma cruzi Metabolism by Metabolic Control Analysis and Kinetic Modeling. Curr. Med. Chem. 2019, 26, 6652–6671. [Google Scholar] [CrossRef]
- Martínez, I.; Nogueda, B.; Martínez-Hernández, F.; Espinoza, B. Microsatellite and mini-exon analysis of Mexican human DTU I Trypanosoma cruzi strains and their susceptibility to nifurtimox and benznidazole. Vector Borne Zoonotic Dis. 2013, 13, 181–187. [Google Scholar] [CrossRef]
- Espinoza, B.; Rico, T.; Sosa, S.; Oaxaca, E.; Vizcaino-Castillo, A.; Caballero, M.L.; Martínez, I. Mexican Trypanosoma cruzi T. cruzi I strains with different degrees of virulence induce diverse humoral and cellular immune responses in a murine experimental infection model. J. Biomed. Biotechnol. 2010, 2010, 890672. [Google Scholar] [CrossRef]
- Pfizer. Material Safety Data Sheet: Glyburide. Available online: https://cdn.pfizer.com/pfizercom/products/material_safety_data/MICRONASE.pdf (accessed on 26 December 2024).
- Cayman Chemical. Safety Data Sheet: Glipizide. Available online: https://s3-us-west-2.amazonaws.com/drugbank/msds/DB01067.pdf?1561061159 (accessed on 6 December 2024).
- Vázquez, C.; Mejia-Tlachi, M.; González-Chávez, Z.; Silva, A.; Rodríguez-Zavala, J.S.; Moreno-Sánchez, R.; Saavedra, E. Buthionine sulfoximine is a multitarget inhibitor of trypanthione synthesis in Trypanosoma cruzi. FEBS Lett. 2017, 591, 3881–3894. [Google Scholar] [CrossRef]
- Vázquez, C.; Encalada, R.; Belmont-Díaz, J.; Rivera, M.; Alvarez, S.; Nogueda-Torres, B.; Saavedra, E. Metabolic control analysis of the transsulfuration pathway and the compensatory role of the cysteine transport in Trypanosoma cruzi. Biosystems 2023, 234, 105066. [Google Scholar] [CrossRef]
- Rodriques Coura, J.; de Castro, S.L. A critical review on Chagas disease chemotherapy. Mem. Inst. Oswaldo Cruz. 2002, 97, 3–24. [Google Scholar] [CrossRef]
- Guedes, P.M.; Silva, G.K.; Gutierrez, F.R.; Silva, J.S. Current status of Chagas disease chemotherapy. Expert Rev. Anti-Infect. Ther. 2011, 9, 609–620. [Google Scholar] [CrossRef]
- Pérez-Molina, J.A.; Crespillo-Andújar, C.; Bosch-Nicolau, P.; Molina, I. Trypanocidal treatment of Chagas disease. Enferm. Infecc. Microbiol. Clin. 2020, 39, 458–470. [Google Scholar] [CrossRef]
- Beltran-Hortelano, I.; Alcolea, V.; Font, M.; Pérez-Silanes, S. Examination of multiple Trypanosoma cruzi targets in a new drug discovery approach for Chagas disease. Bioorg. Med. Chem. 2022, 58, 116577. [Google Scholar] [CrossRef] [PubMed]
- Laureano de Souza, M.; Lapierre, T.J.W.J.D.; Vitor de Lima Marques, G.; Ferraz, W.R.; Penteado, A.B.; Henrique Goulart Trossini, G.; Murta, S.M.F.; de Oliveira, R.B.; de Oliveira Rezende, C., Jr.; Ferreira, R.S. Molecular targets for Chagas disease: Validation, challenges and lead compounds for widely exploited targets. Expert Opin. Ther. Targets 2023, 27, 911–925. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, C.; Matus-Meza, A.S.; Nuñez-Moreno, O.; Barbosa-Sánchez, B.M.; Farías-Gutiérrez, V.M.; Mendoza-Conde, M.; Hernández-Luis, F.; Saavedra, E. Exploring Quinazoline Nitro-Derivatives as Potential Antichagasic Agents: Synthesis and In vitro Evaluation. Molecules 2024, 29, 4501. [Google Scholar] [CrossRef]
- Maciel Diogo, G.; Andrade, J.S.; Sales Junior, P.A.; Maria Fonseca Murta, S.; Dos Santos, V.M.R.; Taylor, J.G. Trypanocidal Activity of Flavanone Derivatives. Molecules 2020, 25, 397. [Google Scholar] [CrossRef]
- Acosta, N.; Yaluff, G.; López, E.; Bobadilla, C.; Ramírez, A.; Fernández, I.; Escobar, P. In vitro susceptibility to benznidazole, nifurtimox and posaconazole of Trypanosoma cruzi isolates from Paraguay (Sensibilidad in vitro a benznidazol, nifurtimox y posaconazol de cepas de Trypanosoma cruzi de Paraguay). Biomed. Rev. Inst. Nac. Salud 2020, 40, 749–763. [Google Scholar] [CrossRef]
- Groop, L.C.; Barzilai, N.; Ratheiser, K.; Luzi, L.; Wåhlin-Boll, E.; Melander, A.; DeFronzo, R.A. Dose-dependent effects of glyburide on insulin secretion and glucose uptake in humans. Diabetes Care 1991, 14, 724–727. [Google Scholar] [CrossRef] [PubMed]
- Palos, I.; Lara-Ramirez, E.E.; Lopez-Cedillo, J.C.; Garcia-Perez, C.; Kashif, M.; Bocanegra-Garcia, V.; Nogueda-Torres, B.; Rivera, G. Repositioning FDA Drugs as Potential Cruzain Inhibitors from Trypanosoma cruzi: Virtual Screening, In Vitro and In Vivo Studies. Molecules 2017, 22, 1015. [Google Scholar] [CrossRef]
- Gómez-Escobedo, R.; Méndez-Álvarez, D.; Vázquez, C.; Saavedra, E.; Vázquez, K.; Alcántara-Farfán, V.; Cordero-Martínez, J.; Gonzalez-Gonzalez, A.; Rivera, G.; Nogueda-Torres, B. Molecular Docking-Based Virtual Screening of FDA-Approved Drugs Using Trypanothione Reductase Identified New Trypanocidal Agents. Molecules 2024, 29, 3796. [Google Scholar] [CrossRef]
- Al-Saleh, Y.; Sabico, S.; Al-Furqani, A.; Jayyousi, A.; Alromaihi, D.; Ba-Essa, E.; Alawadi, F.; Alkaabi, J.; Hassanein, M.; Al-Sifri, S.; et al. Sulfonylureas in the Current Practice of Type 2 Diabetes Management: Are They All the Same? Consensus from the Gulf Cooperation Council (GCC) Countries Advisory Board on Sulfonylureas. Diabetes Ther. 2021, 12, 2115–2132. [Google Scholar] [CrossRef]
- Loos, J.A.; Negro, P.S.; Ortega, H.H.; Salinas, F.J.; Arán, M.; Pellizza, L.; Salerno, G.L.; Cumino, A.C. Anti-echinococcal effect of metformin in advanced experimental cystic echinococcosis: Reprogrammed intermediary carbon metabolism in the parasite. Antimicrob. Agents Chemother. 2024, 68, e0094124. [Google Scholar] [CrossRef]
- Gomaa, M.M.; Nabil El Achy, S.; Hezema, N.N. Could metformin modulate the outcome of chronic murine toxoplasmosis? Acta Trop. 2024, 258, 107339. [Google Scholar] [CrossRef]
- Ozpinar, N.; Karaman, U.; Ozpinar, H.; Dag, S. Do antidiabetic drugs prevent the transformation of Acanthamoeba trophozoite into cyst form? Pathog. Glob. Health 2023, 117, 674–680. [Google Scholar] [CrossRef]
- Uzcategui, N.L.; Figarella, K.; Camacho, N.; Ponte-Sucre, A. Substrate preferences and glucose uptake in glibenclamide-resistant Leishmania parasites. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2005, 140, 395–402. [Google Scholar] [CrossRef]
- Loos, J.A.; Churio, M.S.; Cumino, A.C. Anthelminthic activity of glibenclamide on secondary cystic echinococcosis in mice. PLoS Negl. Trop. Dis. 2017, 11, e0006111. [Google Scholar] [CrossRef]
- López-Olmos, V.; Pérez-Nasser, N.; Piñero, D.; Ortega, E.; Hernandez, R.; Espinoza, B. Biological characterization and genetic diversity of Mexican isolates of Trypanosoma cruzi. Acta Trop. 1998, 69, 239–254. [Google Scholar] [CrossRef]
- Romanha, A.J.; de Castro, S.L.; de Nazaré Correia Soeiro, M.; Lannes-Vieira, J.; Ribeiro, I.; Talvani, A.; Bourdin, B.; Blum, B.; Olivieri, B.; Zan, C.; et al. In Vitro and In Vivo Experimental Models for Drug Screening and Development for Chagas Disease. Mem. Inst. Oswaldo Cruz 2010, 105, 233–238. [Google Scholar] [CrossRef]
- Bustamante, J.M.; Craft, J.M.; Crowe, B.D.; Ketchie, S.A.; Tarleton, R.L. New, combined, and reduced dosing treatment protocols cure Trypanosoma cruzi infection in mice. J. Infect. Dis. 2014, 209, 150–162. [Google Scholar] [CrossRef]
- Brener, Z. Therapeutic activity and criterion of cure on mice experimentally infected with Trypanosoma cruzi. Rev. Inst. Med. Trop. Sao Paulo 1962, 4, 389–396. [Google Scholar]
- Simões-Silva, M.R.; De Araújo, J.S.; Oliveira, G.M.; Demarque, K.C.; Peres, R.B.; D’Almeida-Melo, I.; Batista DG, J.; Da Silva, C.F.; Cardoso-Santos, C.; Da Silva, P.B.; et al. Drug repurposing strategy against Trypanosoma cruzi infection: In vitro and in vivo assessment of the activity of metronidazole in mono- and combined therapy. Biochem. Pharmacol. 2017, 145, 46–53. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Monotherapy | Bi-Therapy (IC50 of BNZ) | |||
|---|---|---|---|---|---|
| +1 µM Antidiabetic | +0.5 µM Antidiabetic | +1 µM Antidiabetic | |||
| HFF-1 (n = 3) | T. cruzi i.v. Infection | HFF-1 (n = 2) | T. cruzi i.v. Infection (n = 8) | T. cruzi i.v. Infection (n = 8) | |
| BNZ | 0.9 ± 0.09 | 0.26 ± 0.01 (10) | ‒ | ||
| GBD | 12 ± 8 | 8.4 ± 6.3 (4) | ‒ | ||
| BNZ + GBD | ‒ | >0.5 | 0.04 * ± 0.007 | 0.03 * ± 0.007 | |
| GLQ | 15 ± 11 | 7.6 ± 2 (4) | ‒ | ||
| BNZ + GLQ | ‒ | 0.25 | 0.05 * ± 0.02 | 0.04 * ± 0.01 | |
| GPZ | 35 ±19 | 14.3 ± 7 (4) | ‒ | ||
| BNZ + GPZ | ‒ | >0.5 | 0.055 * ± 0.02 | 0.04 * ± 0.01 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vázquez, C.; Encalada, R.; Jiménez-Galicia, I.; Gómez-Escobedo, R.; Rivera, G.; Nogueda-Torres, B.; Saavedra, E. Repurposing the Antidiabetic Drugs Glyburide, Gliquidone, and Glipizide in Combination with Benznidazole for Trypanosoma cruzi Infection. Pharmaceuticals 2025, 18, 21. https://doi.org/10.3390/ph18010021
Vázquez C, Encalada R, Jiménez-Galicia I, Gómez-Escobedo R, Rivera G, Nogueda-Torres B, Saavedra E. Repurposing the Antidiabetic Drugs Glyburide, Gliquidone, and Glipizide in Combination with Benznidazole for Trypanosoma cruzi Infection. Pharmaceuticals. 2025; 18(1):21. https://doi.org/10.3390/ph18010021
Chicago/Turabian StyleVázquez, Citlali, Rusely Encalada, Isabel Jiménez-Galicia, Rogelio Gómez-Escobedo, Gildardo Rivera, Benjamín Nogueda-Torres, and Emma Saavedra. 2025. "Repurposing the Antidiabetic Drugs Glyburide, Gliquidone, and Glipizide in Combination with Benznidazole for Trypanosoma cruzi Infection" Pharmaceuticals 18, no. 1: 21. https://doi.org/10.3390/ph18010021
APA StyleVázquez, C., Encalada, R., Jiménez-Galicia, I., Gómez-Escobedo, R., Rivera, G., Nogueda-Torres, B., & Saavedra, E. (2025). Repurposing the Antidiabetic Drugs Glyburide, Gliquidone, and Glipizide in Combination with Benznidazole for Trypanosoma cruzi Infection. Pharmaceuticals, 18(1), 21. https://doi.org/10.3390/ph18010021