
Gene Therapy in the Light of Lifestyle Diseases: Budesonide, Acetaminophen and Simvastatin Modulates rAAV Transduction Efficiency


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
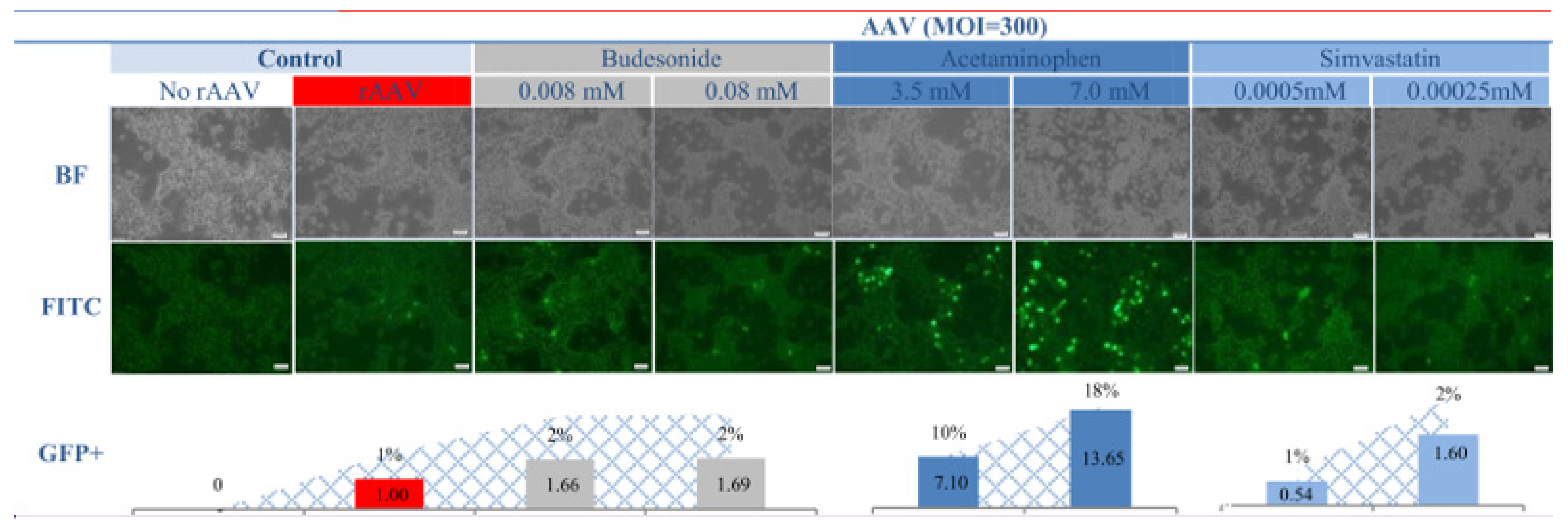
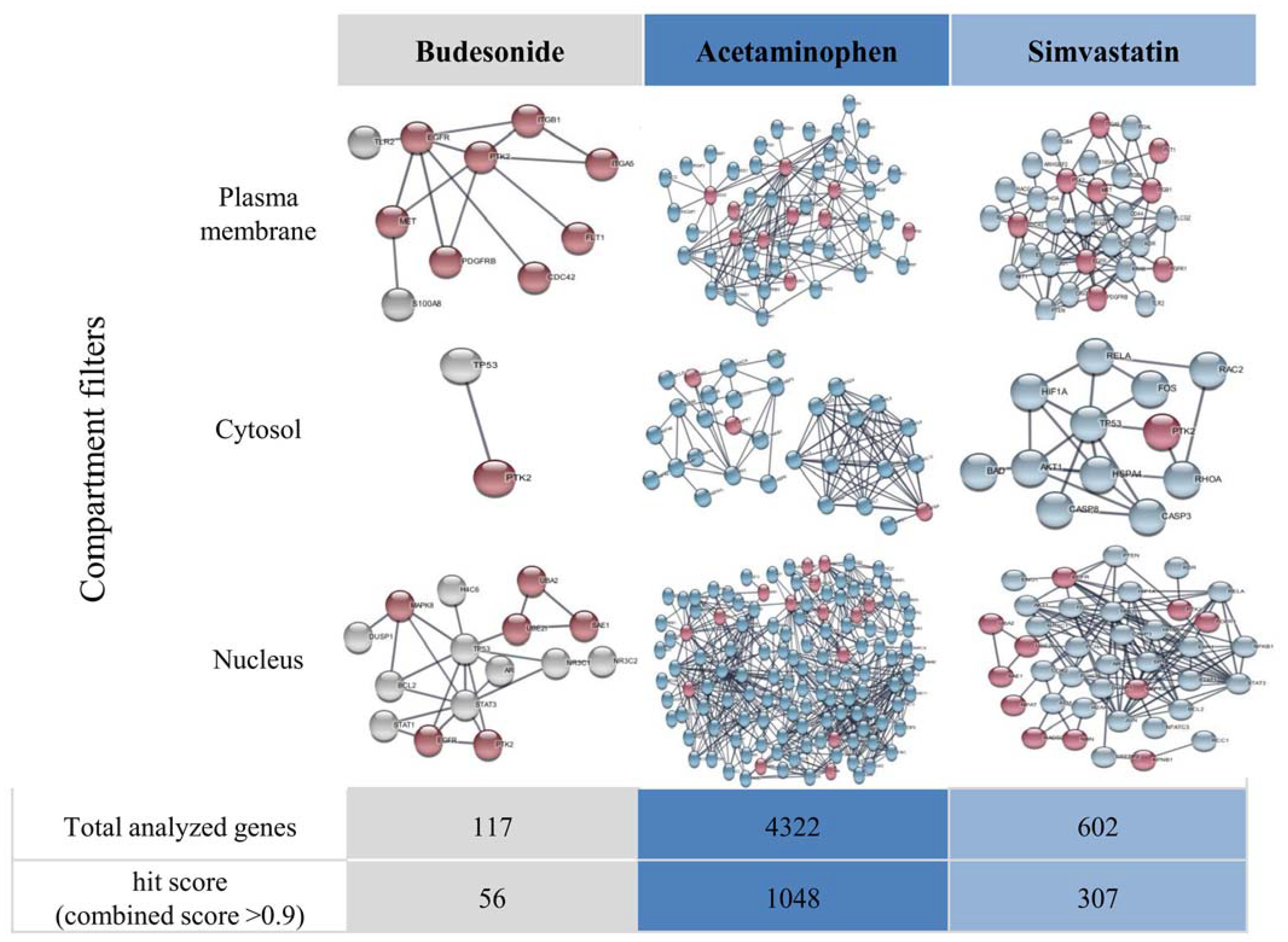
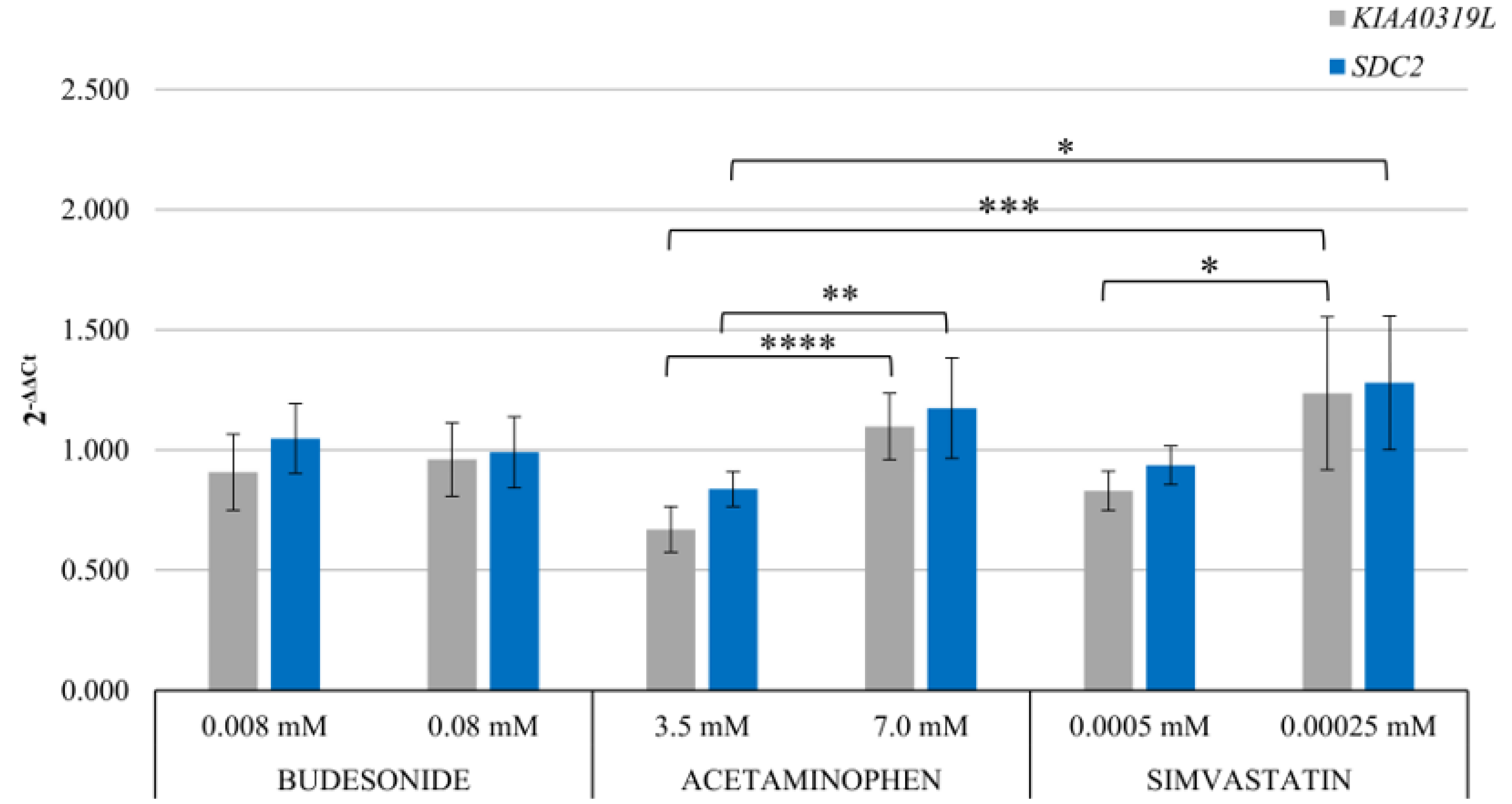
2. Results and Discussion
3. Conclusions
4. Materials and Methods
4.1. Reagents and Materials
Preparation of Drug Solutions
4.2. rAAV Transduction
4.3. Measurement of Transduction Efficiency
- A.
- Real-time PCR for examination of rAAV genome copy number
- B.
- Fluorescent imaging
4.4. Assessment of the Expression of rAAV Receptors
5. Visualization of Functional Gene Interactions in Drug-Exposed Cells
6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, D.; Tai, P.W.L.; Gao, G. Adeno-associated virus vector as a platform for gene therapy delivery. Nat. Rev. Drug Discov. 2019, 18, 358–378. [Google Scholar] [CrossRef] [PubMed]
- Au, H.K.E.; Isalan, M.; Mielcarek, M. Gene Therapy Advances: A Meta-Analysis of AAV Usage in Clinical Settings. Front. Med. 2021, 9, 809118. [Google Scholar] [CrossRef] [PubMed]
- Heo, Y.-A. Etranacogene dezaparvovec: First approval. Drugs 2023, 83, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Keam, S.J. Eladocagene exuparvovec: First approval. Drugs 2022, 82, 1427–1432. [Google Scholar] [CrossRef]
- Philippidis, A. BioMarin’s ROCTAVIAN wins food and drug administration approval as first gene therapy for severe hemophilia A. Hum. Gene Ther. 2023, 34, 665–668. [Google Scholar] [CrossRef]
- Zittersteijn, H.A.; Gonçalves, M.; Hoeben, R.C. A primer to gene therapy: Progress, prospects, and problems. J. Inherit. Metab. Dis. 2021, 44, 54–71. [Google Scholar] [CrossRef]
- Srivastava, A.; Mallela, K.M.G.; Deorkar, N.; Brophy, G. Manufacturing Challenges and Rational Formulation Development for AAV Viral Vectors. J. Pharm. Sci. 2021, 110, 2609–2624. [Google Scholar] [CrossRef]
- Pupo, A.; Fernández, A.; Low, S.H.; François, A.; Suárez-Amarán, L.; Samulski, R.J. AAV vectors: The Rubik’s cube of human gene therapy. Mol. Ther. 2022, 30, 3515–3541. [Google Scholar] [CrossRef]
- Stone, D.; Aubert, M.; Jerome, K.R. Adeno-associated virus vectors and neurotoxicity—Lessons from preclinical and human studies. Gene Ther. 2023, 1–14. [Google Scholar] [CrossRef]
- Duan, D. Lethal immunotoxicity in high-dose systemic AAV therapy. Mol. Ther. 2023, 31, 3123–3126. [Google Scholar] [CrossRef]
- Cao, D.; Byrne, B.J.; de Jong, Y.P.; Terhorst, C.; Duan, D.; Herzog, R.W.; Kumar, S.R. Innate Immune Sensing of Adeno-Associated Virus Vectors. Hum. Gene Ther. 2024, 35, 451–463. [Google Scholar] [CrossRef] [PubMed]
- Costa-Verdera, H.; Unzu, C.; Valeri, E.; Adriouch, S.; González Aseguinolaza, G.; Mingozzi, F.; Kajaste-Rudnitski, A. Understanding and tackling immune responses to adeno-associated viral vectors. Hum. Gene Ther. 2023, 34, 836–852. [Google Scholar] [CrossRef] [PubMed]
- Boutin, S.; Monteilhet, V.; Veron, P.; Leborgne, C.; Benveniste, O.; Montus, M.F.; Masurier, C. Prevalence of serum IgG and neutralizing factors against adeno-associated virus (AAV) types 1, 2, 5, 6, 8, and 9 in the healthy population: Implications for gene therapy using AAV vectors. Hum. Gene Ther. 2010, 21, 704–712. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.; Liu, S.; Ou, L. rAAV immunogenicity, toxicity, and durability in 255 clinical trials: A meta-analysis. Front. Immunol. 2022, 13, 1001263. [Google Scholar] [CrossRef]
- Srivastava, A. Rationale and strategies for the development of safe and effective optimized AAV vectors for human gene therapy. Mol. Ther. -Nucleic Acids 2023, 32, 949–959. [Google Scholar] [CrossRef]
- Tickner, Z.J.; Farzan, M. Riboswitches for Controlled Expression of Therapeutic Transgenes Delivered by Adeno-Associated Viral Vectors. Pharmaceuticals 2021, 14, 554. [Google Scholar] [CrossRef]
- Guilbaud, M.; Devaux, M.; Couzinié, C.; Le Duff, J.; Toromanoff, A.; Vandamme, C.; Jaulin, N.; Gernoux, G.; Larcher, T.; Moullier, P.; et al. Five Years of Successful Inducible Transgene Expression Following Locoregional Adeno-Associated Virus Delivery in Nonhuman Primates with No Detectable Immunity. Hum. Gene Ther. 2019, 30, 802–813. [Google Scholar] [CrossRef]
- Słyk, Ż.; Stachowiak, N.; Małecki, M. Recombinant Adeno-Associated Virus Vectors for Gene Therapy of the Central Nervous System: Delivery Routes and Clinical Aspects. Biomedicines 2024, 12, 1523. [Google Scholar] [CrossRef]
- Coroadinha, A.S. Host Cell Restriction Factors Blocking Efficient Vector Transduction: Challenges in Lentiviral and Adeno-Associated Vector Based Gene Therapies. Cells 2023, 12, 732. [Google Scholar] [CrossRef]
- Mano, M.; Ippodrino, R.; Zentilin, L.; Zacchigna, S.; Giacca, M. Genome-wide RNAi screening identifies host restriction factors critical for in vivo AAV transduction. Proc. Natl. Acad. Sci. USA 2015, 112, 11276–11281. [Google Scholar] [CrossRef]
- Zajkowska, A.; Czajka, M.; Gulik, K.; Gawrychowski, K.; Małecki, M. Profiling of microRNA as a tool to introduce rAAV vectors in gene therapy of breast cancer: A preliminary report. Adv. Clin. Exp. Med. Off. Organ Wroc. Med. Univ. 2023, 32, 1201–1210. [Google Scholar] [CrossRef] [PubMed]
- Bieńkowska, A.; Ducher, M.; Orzechowska, M.; Słyk, Ż.; Ciepiela, O.; Jaworowski, J.; Małecki, M. Increased temperature-related adeno-associated virus vectors transduction of ovarian cancer cells—Essential signatures of AAV receptor and heat shock proteins. Exp. Ther. Med. 2019, 18, 4718–4732. [Google Scholar] [CrossRef] [PubMed]
- Bieńkowska, A.; Kuźmicka, W.; Ciepiela, O.; Ochocki, J.; Małecki, M. Increased Temperature Facilitates Adeno-Associated Virus Vector Transduction of Colorectal Cancer Cell Lines in a Manner Dependent on Heat Shock Protein Signature. BioMed Res. Int. 2020, 1, 9107140. [Google Scholar] [CrossRef] [PubMed]
- Czajka, M.; Zajkowska, A.; Gawlak, M.; Bujalska-Zadrozny, M.; Malecki, M. Mosaic Recombinant Adeno-associated Virus Vector rAAV/DJ/CAG for Targeted Gene Delivery to Melanoma Cells Metastasized to the Lung. Anticancer. Res. 2020, 40, 4425–4444. [Google Scholar] [CrossRef]
- Nathwani, A.C.; Cochrane, M.; McIntosh, J.; Ng, C.Y.C.; Zhou, J.; Gray, J.T.; Davidoff, A.M. Enhancing transduction of the liver by adeno-associated viral vectors. Gene Ther. 2009, 16, 60–69. [Google Scholar] [CrossRef]
- Hölscher, C.; Sonntag, F.; Henrich, K.; Chen, Q.; Beneke, J.; Matula, P.; Rohr, K.; Kaderali, L.; Beil, N.; Erfle, H.; et al. The SUMOylation Pathway Restricts Gene Transduction by Adeno-Associated Viruses. PLoS Pathog. 2015, 11, e1005281. [Google Scholar] [CrossRef]
- Chai, Z.; Zhang, X.; Dobbins, A.L.; Samulski, R.J.; Merricks, E.P.; Nichols, T.C.; Li, C. Dexamethasone Transiently Enhances Transgene Expression in the Liver When Administered at Late-Phase Post Long-Term Adeno-Associated Virus Transduction. Hum. Gene Ther. 2022, 33, 119–130. [Google Scholar] [CrossRef]
- Business Research Insights. Paracetamol Market Size. 2024. Available online: https://www.businessresearchinsights.com/market-reports/paracetamol-market-102048 (accessed on 11 September 2024).
- Business Research Insights. Budesonide Market Size 2024. Available online: https://www.businessresearchinsights.com/market-reports/budesonide-market-102187 (accessed on 11 September 2024).
- Business Research Insights. Statin Market Size. 2024. Available online: https://www.businessresearchinsights.com/market-reports/statin-market-108449 (accessed on 11 September 2024).
- Freo, U.; Ruocco, C.; Valerio, A.; Scagnol, I.; Nisoli, E. Paracetamol: A Review of Guideline Recommendations. J. Clin. Med. 2021, 10, 3420. [Google Scholar] [CrossRef]
- Kalola, U.K.; Ambati, S. StatPearls; StatPearls Publishing: Treasure Island, Finland, 2023. Available online: https://www.ncbi.nlm.nih.gov/books/NBK563201/ (accessed on 11 September 2024).
- Issa, S.S.; Shaimardanova, A.A.; Solovyeva, V.V.; Rizvanov, A.A. Various AAV Serotypes and Their Applications in Gene Therapy: An Overview. Cells 2023, 12, 785. [Google Scholar] [CrossRef]
- Chowdhury, E.A.; Meno-Tetang, G.; Chang, H.Y.; Wu, S.; Huang, H.W.; Jamier, T.; Chandran, J.; Shah, D.K. Current progress and limitations of AAV mediated delivery of protein therapeutic genes and the importance of developing quantitative pharmacokinetic/pharmacodynamic (PK/PD) models. Adv. Drug Deliv. Rev. 2021, 170, 214–237. [Google Scholar] [CrossRef]
- Liu, S.; Chowdhury, E.A.; Xu, V.; Jerez, A.; Mahmood, L.; Ly, B.Q.; Le, H.K.; Nguyen, A.; Rajwade, A.; Meno-Tetang, G.; et al. Whole-Body Disposition and Physiologically Based Pharmacokinetic Modeling of Adeno-Associated Viruses and the Transgene Product. J. Pharm. Sci. 2024, 113, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhu, M.; Zhang, Y.; Diao, Y. Crossing the blood-brain barrier with AAV vectors. Metab. Brain Dis. 2021, 36, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, T.K.; Samulski, R.J. Addressing high dose AAV toxicity—‘one and done’ or ‘slower and lower’? Expert Opin. Biol. Ther. 2022, 22, 1067–1071. [Google Scholar] [CrossRef] [PubMed]
- Touahri, Y.; Dixit, R.; Kofoed, R.H.; Miloska, K.; Park, E.; Raeisossadati, R.; Markham-Coultes, K.; David, L.A.; Rijal, H.; Zhao, J.; et al. Focused ultrasound as a novel strategy for noninvasive gene delivery to retinal Müller glia. Theranostics 2020, 10, 2982–2999. [Google Scholar] [CrossRef]
- Corti, M.; Cleaver, B.; Clément, N.; Conlon, T.J.; Faris, K.J.; Wang, G.; Benson, J.; Tarantal, A.F.; Fuller, D.; Herzog, R.W.; et al. Evaluation of Readministration of a Recombinant Adeno-Associated Virus Vector Expressing Acid Alpha-Glucosidase in Pompe Disease: Preclinical to Clinical Planning. Hum. Gene Therapy. Clin. Dev. 2015, 26, 185–193. [Google Scholar] [CrossRef]
- Musa, A.; Ghoraie, L.S.; Zhang, S.D.; Glazko, G.; Yli-Harja, O.; Dehmer, M.; Haibe-Kains, B.; Emmert-Streib, F. A review of connectivity map and computational approaches in pharmacogenomics. Brief. Bioinform. 2018, 19, 506–523. [Google Scholar] [CrossRef]
- Nicolson, S.C.; Li, C.; Hirsch, M.L.; Setola, V.; Samulski, R.J. Identification and Validation of Small Molecules That Enhance Recombinant Adeno-associated Virus Transduction following High-Throughput Screens. J. Virol. 2016, 90, 7019–7031. [Google Scholar] [CrossRef]
- Grimm, D.; Lee Joyce, S.; Wang, L.; Desai, T.; Akache, B.; Storm Theresa, A.; Kay Mark, A. In Vitro and In Vivo Gene Therapy Vector Evolution via Multispecies Interbreeding and Retargeting of Adeno-Associated Viruses. J. Virol. 2008, 82, 5887–5911. [Google Scholar] [CrossRef]
- Katada, Y.; Kobayashi, K.; Tsubota, K.; Kurihara, T. Evaluation of AAV-DJ vector for retinal gene therapy. PeerJ 2019, 7, e6317. [Google Scholar] [CrossRef]
- Yoo, M.; Shin, J.; Kim, J.; Ryall, K.A.; Lee, K.; Lee, S.; Jeon, M.; Kang, J.; Tan, A.C. DSigDB: Drug signatures database for gene set analysis. Bioinformatics 2015, 31, 3069–3071. [Google Scholar] [CrossRef]
- Chen, Y.W.; Diamante, G.; Ding, J.; Nghiem, T.X.; Yang, J.; Ha, S.M.; Cohn, P.; Arneson, D.; Blencowe, M.; Garcia, J.; et al. PharmOmics: A species- and tissue-specific drug signature database and gene-network-based drug repositioning tool. iScience 2022, 25, 104052. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S.; Feunang, Y.D.; Guo, A.C.; Lo, E.J.; Marcu, A.; Grant, J.R.; Sajed, T.; Johnson, D.; Li, C.; Sayeeda, Z.; et al. DrugBank 5.0: A major update to the DrugBank database for 2018. Nucleic Acids Res. 2018, 46, D1074–D1082. [Google Scholar] [CrossRef] [PubMed]
- Freshour, S.L.; Kiwala, S.; Cotto, K.C.; Coffman, A.C.; McMichael, J.F.; Song, J.J.; Griffith, M.; Griffith, O.L.; Wagner, A.H. Integration of the Drug-Gene Interaction Database (DGIdb 4.0) with open crowdsource efforts. Nucleic Acids Res. 2021, 49, D1144–D1151. [Google Scholar] [CrossRef] [PubMed]
- Pillay, S.; Meyer, N.L.; Puschnik, A.S.; Davulcu, O.; Diep, J.; Ishikawa, Y.; Jae, L.T.; Wosen, J.E.; Nagamine, C.M.; Chapman, M.S.; et al. An essential receptor for adeno-associated virus infection. Nature 2016, 530, 108–112. [Google Scholar] [CrossRef]
- Zhong, L.; Zhao, W.; Wu, J.; Li, B.; Zolotukhin, S.; Govindasamy, L.; Agbandje-McKenna, M.; Srivastava, A. A dual role of EGFR protein tyrosine kinase signaling in ubiquitination of AAV2 capsids and viral second-strand DNA synthesis. Mol. Ther. J. Am. Soc. Gene Ther. 2007, 15, 1323–1330. [Google Scholar] [CrossRef]
- Aurnhammer, C.; Haase, M.; Muether, N.; Hausl, M.; Rauschhuber, C.; Huber, I.; Nitschko, H.; Busch, U.; Sing, A.; Ehrhardt, A.; et al. Universal real-time PCR for the detection and quantification of adeno-associated virus serotype 2-derived inverted terminal repeat sequences. Hum. Gene Ther. Methods 2012, 23, 18–28. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Słyk, Ż.; Stachowiak, N.; Małecki, M. Gene Therapy in the Light of Lifestyle Diseases: Budesonide, Acetaminophen and Simvastatin Modulates rAAV Transduction Efficiency. Pharmaceuticals 2024, 17, 1213. https://doi.org/10.3390/ph17091213
Słyk Ż, Stachowiak N, Małecki M. Gene Therapy in the Light of Lifestyle Diseases: Budesonide, Acetaminophen and Simvastatin Modulates rAAV Transduction Efficiency. Pharmaceuticals. 2024; 17(9):1213. https://doi.org/10.3390/ph17091213
Chicago/Turabian StyleSłyk, Żaneta, Natalia Stachowiak, and Maciej Małecki. 2024. "Gene Therapy in the Light of Lifestyle Diseases: Budesonide, Acetaminophen and Simvastatin Modulates rAAV Transduction Efficiency" Pharmaceuticals 17, no. 9: 1213. https://doi.org/10.3390/ph17091213
APA StyleSłyk, Ż., Stachowiak, N., & Małecki, M. (2024). Gene Therapy in the Light of Lifestyle Diseases: Budesonide, Acetaminophen and Simvastatin Modulates rAAV Transduction Efficiency. Pharmaceuticals, 17(9), 1213. https://doi.org/10.3390/ph17091213