

Intestinal Metabolism of Crocin and a Pharmacokinetics and Pharmacodynamics Study in the Chronic Social Defeat Stress Mouse Model

Abstract
1. Introduction
2. Results
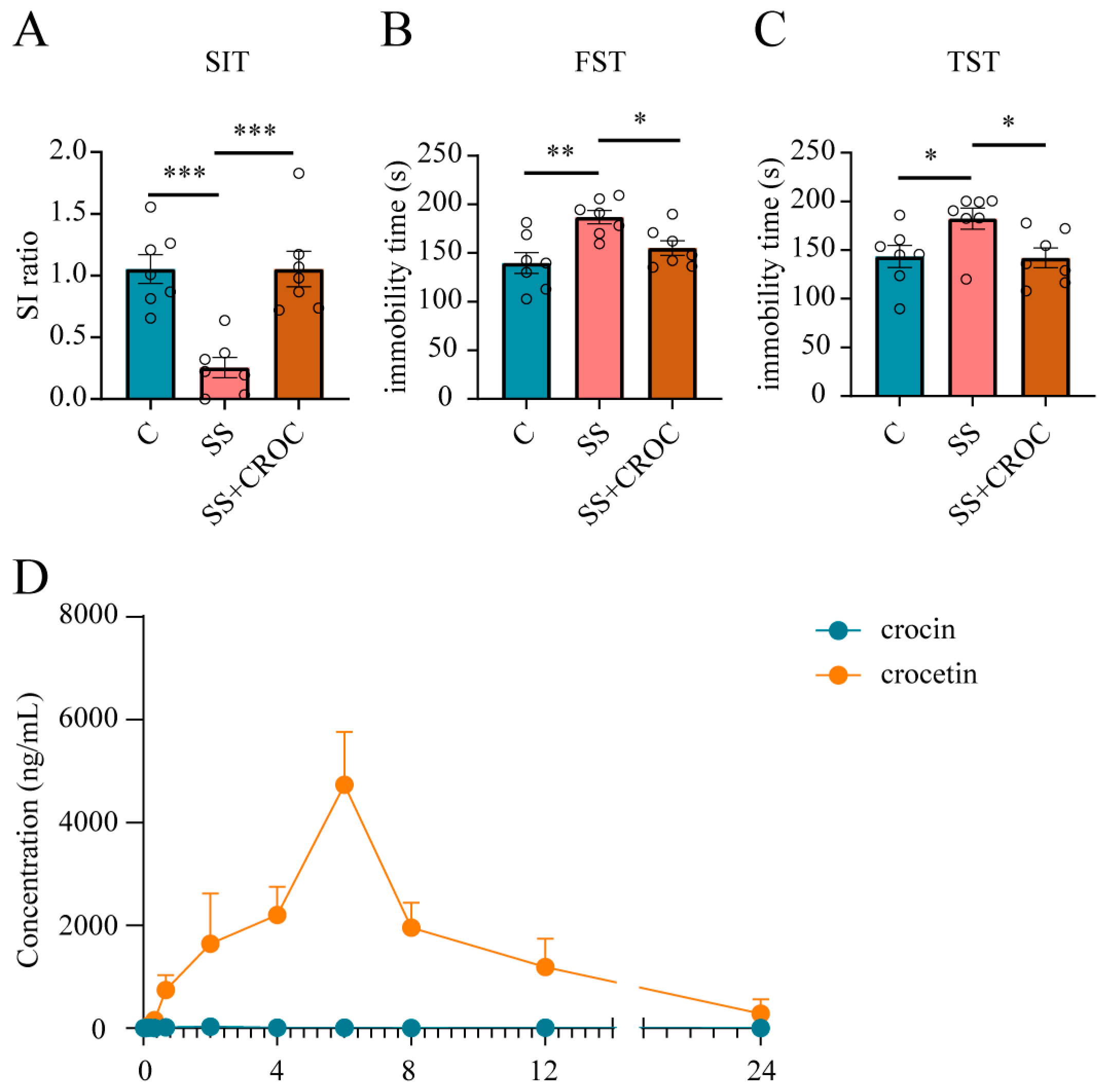
2.1. Pharmacokinetics and Pharmacodynamics Study of Crocin in CSDS Model Mice
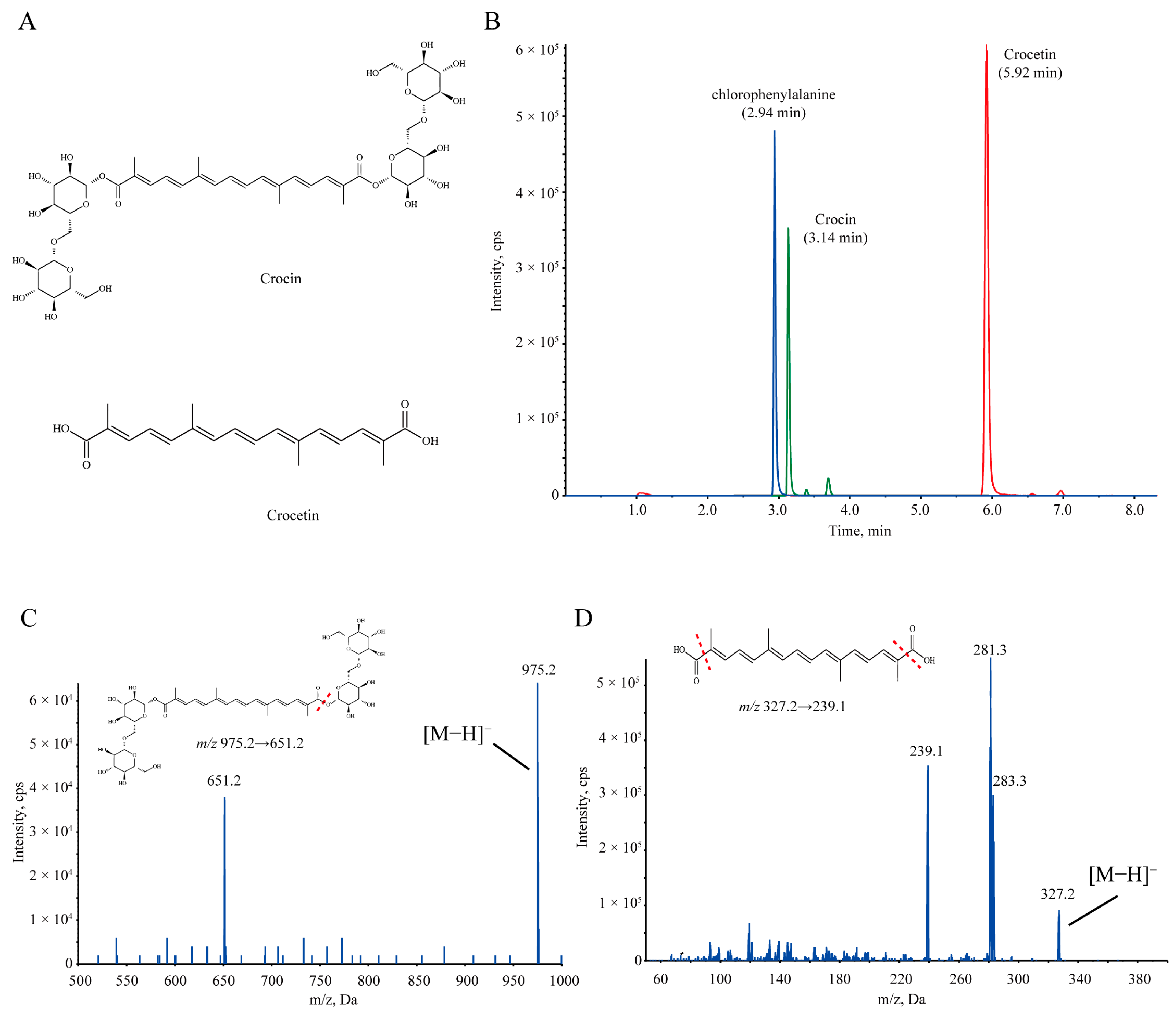
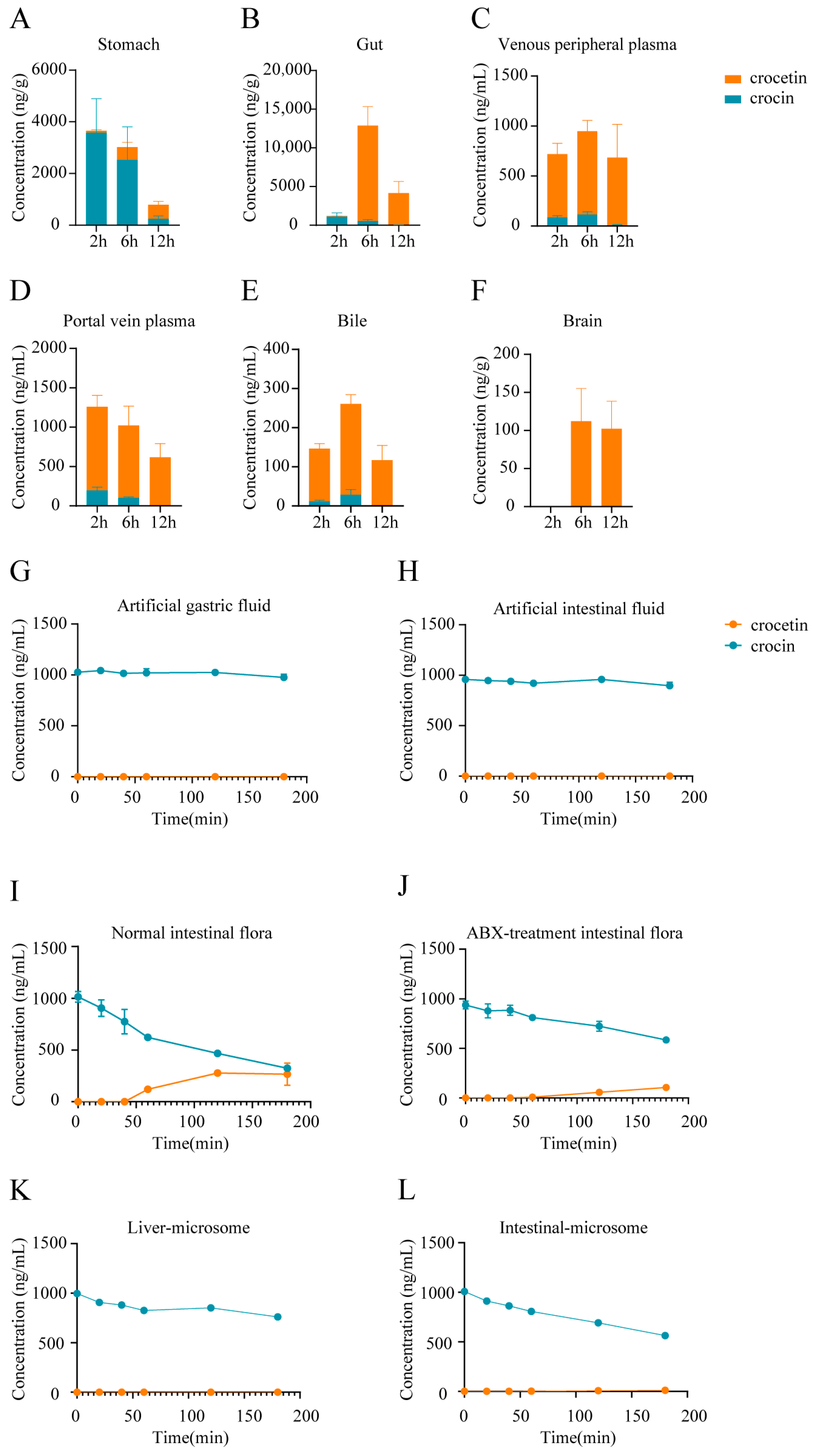
2.2. Metabolism of Orally Administered Crocin Both In Vivo and In Vitro
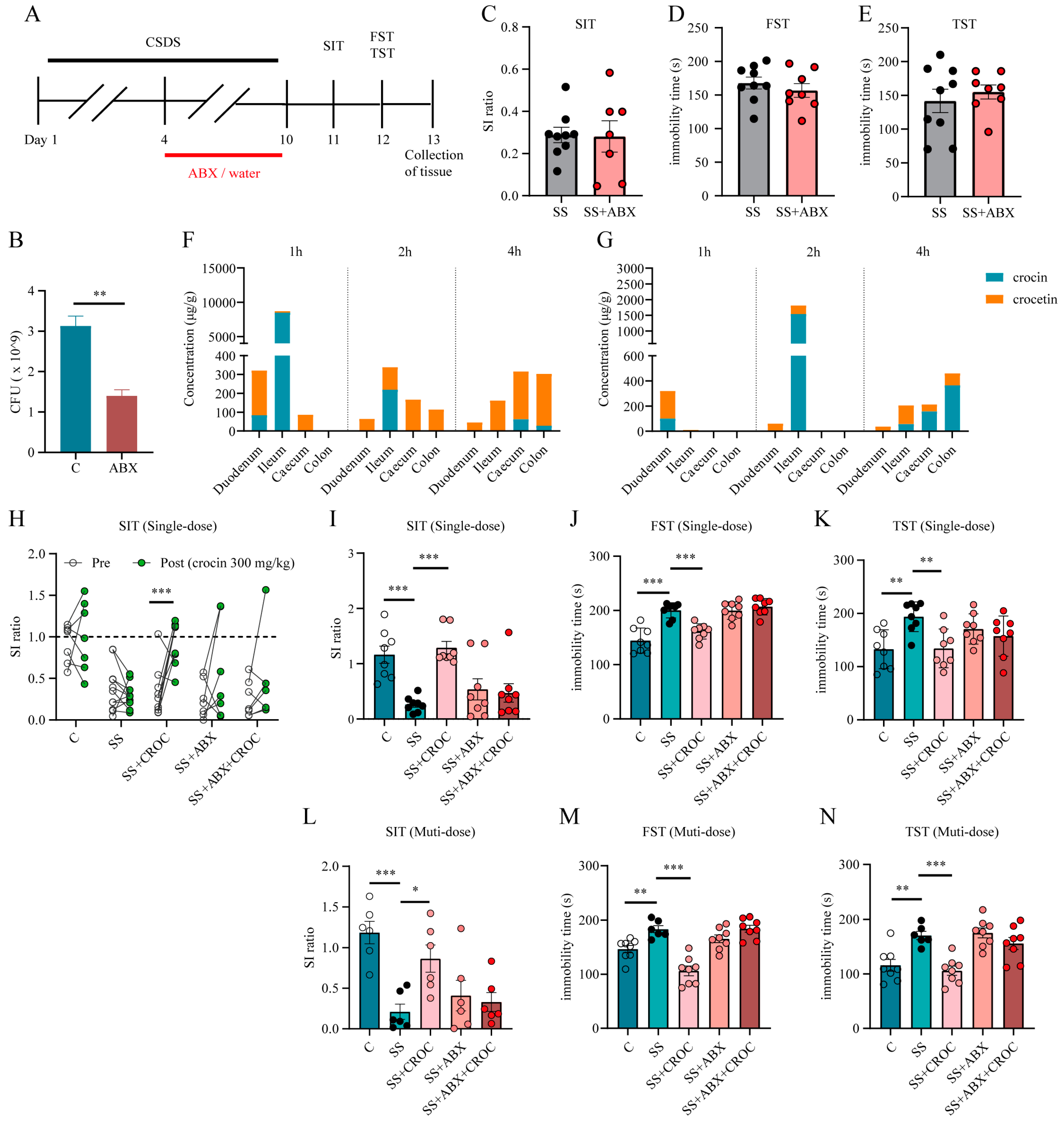
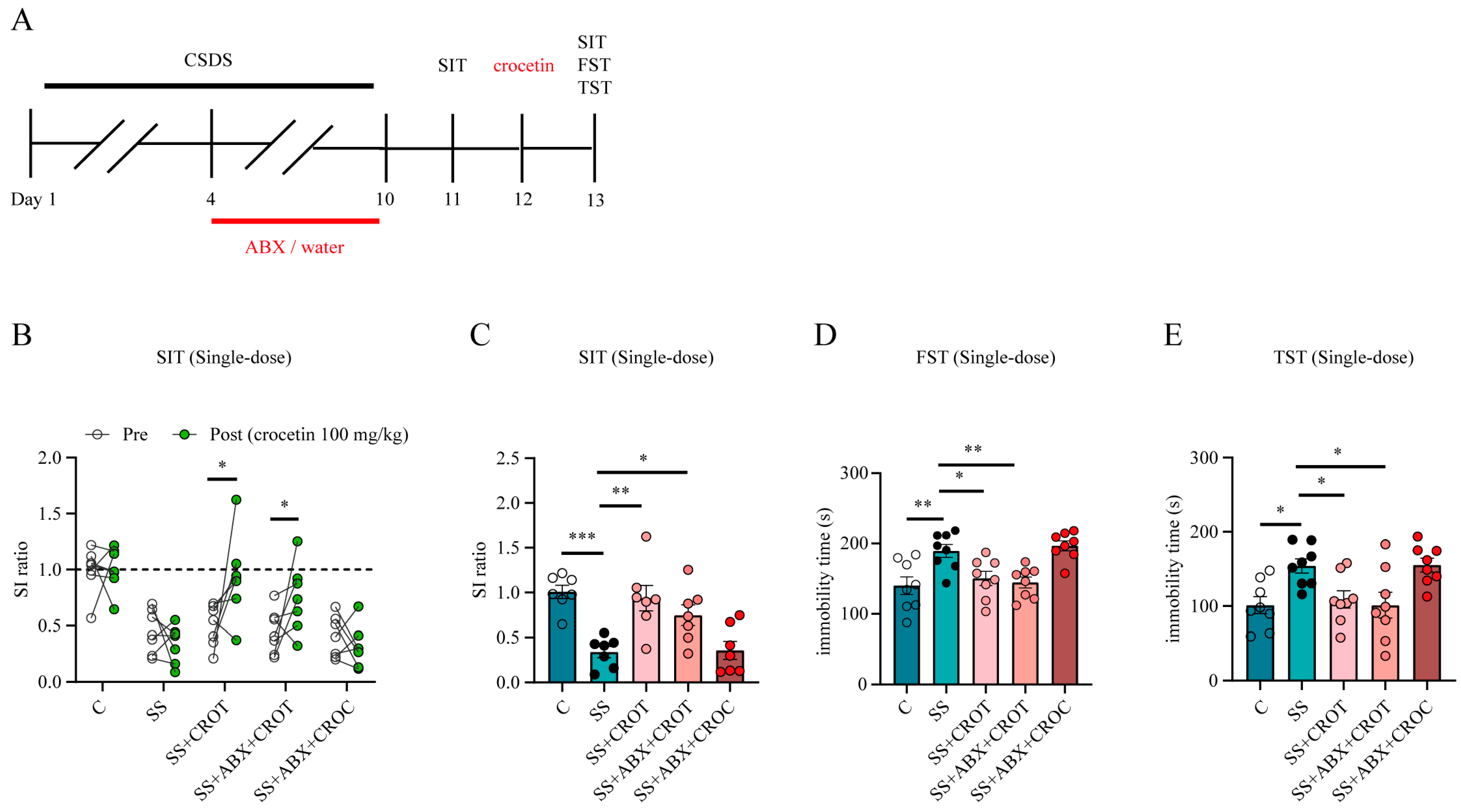
2.3. Inhibition of the Gut Flora Reduced the Production of Crocetin and Affected the Antidepressive Efficacy of Crocin
2.4. Crocetin Is an Actual Pharmacodynamic Substance That Has Rapid Antidepressant Efficacy
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. CSDS Model
4.4. Behavioural Testing
4.4.1. Social Interaction Test (SIT)
4.4.2. Forced Swimming Test (FST)
4.4.3. Tail Suspension Test (TST)
4.5. Antibiotic Cocktail Treatment
4.6. Chemical Analysis
4.7. In Vitro Metabolism of Crocin
4.8. Bacterial Flora Enumeration
4.9. Pharmacokinetics Study in Rats
4.10. Tissue Distribution Study in Rats
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arioz, B.I.; Tastan, B.; Tarakcioglu, E.; Tufekci, K.U.; Olcum, M.; Ersoy, N.; Bagriyanik, A.; Genc, K.; Genc, S. Melatonin Attenuates LPS-Induced Acute Depressive-Like Behaviors and Microglial NLRP3 Inflammasome Activation through the SIRT1/Nrf2 Pathway. Front. Immunol. 2019, 10, 1511. [Google Scholar] [CrossRef] [PubMed]
- Schechter, L.E.; Ring, R.H.; Beyer, C.E.; Hughes, Z.A.; Khawaja, X.; Malberg, J.E.; Rosenzweig-Lipson, S. Innovative approaches for the development of antidepressant drugs: Current and future strategies. NeuroRx J. Am. Soc. Exp. Neurother. 2005, 2, 590–611. [Google Scholar] [CrossRef] [PubMed]
- Tsuboi, H.; Tatsumi, A.; Yamamoto, K.; Kobayashi, F.; Shimoi, K.; Kinae, N. Possible connections among job stress, depressive symptoms, lipid modulation and antioxidants. J. Affect. Disord. 2006, 91, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Short, B.; Fong, J.; Galvez, V.; Shelker, W.; Loo, C.K. Side-effects associated with ketamine use in depression: A systematic review. Lancet. Psychiatry 2018, 5, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Bathaie, S.Z.; Mousavi, S.Z. New applications and mechanisms of action of saffron and its important ingredients. Crit. Rev. Food Sci. Nutr. 2010, 50, 761–786. [Google Scholar] [CrossRef] [PubMed]
- Amin, B.; Hosseinzadeh, H. Evaluation of aqueous and ethanolic extracts of saffron, Crocus sativus L., and its constituents, safranal and crocin in allodynia and hyperalgesia induced by chronic constriction injury model of neuropathic pain in rats. Fitoterapia 2012, 83, 888–895. [Google Scholar] [CrossRef] [PubMed]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [PubMed]
- Elsherbiny, N.M.; Salama, M.F.; Said, E.; El-Sherbiny, M.; Al-Gayyar, M.M. Crocin protects against doxorubicin-induced myocardial toxicity in rats through down-regulation of inflammatory and apoptic pathways. Chem.-Biol. Interact. 2016, 247, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Shahidani, S.; Rajaei, Z.; Alaei, H. Pretreatment with crocin along with treadmill exercise ameliorates motor and memory deficits in hemiparkinsonian rats by anti-inflammatory and antioxidant mechanisms. Metab. Brain Dis. 2019, 34, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Cai, X.; Hu, W.; Li, Z.; Kong, F.; Chen, X.; Wang, D. Investigation of the neuroprotective effects of crocin via antioxidant activities in HT22 cells and in mice with Alzheimer’s disease. Int. J. Mol. Med. 2019, 43, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Cerdá-Bernad, D.; Valero-Cases, E.; Pastor, J.J.; Frutos, M.J. Saffron bioactives crocin, crocetin and safranal: Effect on oxidative stress and mechanisms of action. Crit. Rev. Food Sci. Nutr. 2022, 62, 3232–3249. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, H.; Sadeghnia, H.R.; Ghaeni, F.A.; Motamedshariaty, V.S.; Mohajeri, S.A. Effects of saffron (Crocus sativus L.) and its active constituent, crocin, on recognition and spatial memory after chronic cerebral hypoperfusion in rats. Phytother. Res. PTR 2012, 26, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Previn, R.; Lu, L.; Liao, R.F.; Jin, Y.; Wang, R.K. Crocin, a natural product attenuates lipopolysaccharide-induced anxiety and depressive-like behaviors through suppressing NF-kB and NLRP3 signaling pathway. Brain Res. Bull. 2018, 142, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Shu, R.; Wu, C.; Tong, Y.; Xiong, Z.; Zhou, J.; Yu, C.; Xie, X.; Fu, Z. Crocin-I alleviates the depression-like behaviors probably via modulating “microbiota-gut-brain” axis in mice exposed to chronic restraint stress. J. Affect. Disord. 2020, 276, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Alsanie, W.F.; Alamri, A.S.; Abdulaziz, O.; Salih, M.M.; Alamri, A.; Asdaq, S.M.B.; Alhomrani, M.H.; Alhomrani, M. Antidepressant Effect of Crocin in Mice with Chronic Mild Stress. Molecules 2022, 27, 5462. [Google Scholar] [CrossRef]
- Zhang, F.; Zhu, X.; Yu, P.; Sheng, T.; Wang, Y.; Ye, Y. Crocin ameliorates depressive-like behaviors induced by chronic restraint stress via the NAMPT-NAD(+)-SIRT1 pathway in mice. Neurochem. Int. 2022, 157, 105343. [Google Scholar] [CrossRef] [PubMed]
- Kolahdooz, G.; Vosough, I.; Sepahi, S.; Mohajeri, S.A. The effect of crocin versus sertraline in treatment of mild to moderate postpartum depression: A double-blind, randomized clinical trial. Int. Clin. Psychopharmacol. 2023, 38, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Talaei, A.; Hassanpour Moghadam, M.; Sajadi Tabassi, S.A.; Mohajeri, S.A. Crocin, the main active saffron constituent, as an adjunctive treatment in major depressive disorder: A randomized, double-blind, placebo-controlled, pilot clinical trial. J. Affect. Disord. 2015, 174, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Ghajar, A.; Neishabouri, S.M.; Velayati, N.; Jahangard, L.; Matinnia, N.; Haghighi, M.; Ghaleiha, A.; Afarideh, M.; Salimi, S.; Meysamie, A.; et al. Crocus sativus L. versus Citalopram in the Treatment of Major Depressive Disorder with Anxious Distress: A Double-Blind, Controlled Clinical Trial. Pharmacopsychiatry 2017, 50, 152–160. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, J.; Li, N.; Xu, P.; Qu, S.; Cheng, J.; Wang, M.; Li, X.; Song, Y.; Xiao, F.; et al. Targeting cAMP in D1-MSNs in the nucleus accumbens, a new rapid antidepressant strategy. Acta Pharm. Sinica. B 2024, 14, 667–681. [Google Scholar] [CrossRef]
- Zhang, Y.; Lu, W.; Wang, Z.; Zhang, R.; Xie, Y.; Guo, S.; Jiao, L.; Hong, Y.; Di, Z.; Wang, G.; et al. Reduced Neuronal cAMP in the Nucleus Accumbens Damages Blood-Brain Barrier Integrity and Promotes Stress Vulnerability. Biol. Psychiatry 2020, 87, 526–537. [Google Scholar] [CrossRef] [PubMed]
- Lautenschläger, M.; Sendker, J.; Hüwel, S.; Galla, H.J.; Brandt, S.; Düfer, M.; Riehemann, K.; Hensel, A. Intestinal formation of trans-crocetin from saffron extract (Crocus sativus L.) and in vitro permeation through intestinal and blood brain barrier. Phytomedicine Int. J. Phytother. Phytopharm. 2015, 22, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Nakano, T.; Takahashi, M.; Nagao, A. Orally administered crocetin and crocins are absorbed into blood plasma as crocetin and its glucuronide conjugates in mice. J. Agric. Food Chem. 2005, 53, 7302–7306. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Qian, Z.; Du, P.; Fu, J. Pharmacokinetic properties of crocin (crocetin digentiobiose ester) following oral administration in rats. Phytomedicine Int. J. Phytother. Phytopharm. 2007, 14, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fei, F.; Zhen, L.; Zhu, X.; Wang, J.; Li, S.; Geng, J.; Sun, R.; Yu, X.; Chen, T.; et al. Sensitive analysis and simultaneous assessment of pharmacokinetic properties of crocin and crocetin after oral administration in rats. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1044–1045, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Geng, J.; Hong, Y.; Jiao, L.; Li, S.; Sun, R.; Xie, Y.; Yan, C.; Aa, J.; Wang, G. Orally Administered Crocin Protects Against Cerebral Ischemia/Reperfusion Injury Through the Metabolic Transformation of Crocetin by Gut Microbiota. Front. Pharmacol. 2019, 10, 440. [Google Scholar] [CrossRef] [PubMed]
- Shakya, R.; Nepal, M.R.; Kang, M.J.; Jeong, T.C. Effects of Intestinal Microbiota on Pharmacokinetics of Crocin and Crocetin in Male Sprague-Dawley Rats. Metabolites 2020, 10, 424. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Li, Q.; Xu, Z.; Chen, Z.; Tao, Y.; Tong, Y.; Wang, T.; Chen, S.; Wang, P. Detection of the role of intestinal flora and tryptophan metabolism involved in antidepressant-like actions of crocetin based on a multi-omics approach. Psychopharmacology 2022, 239, 3657–3677. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Li, Q.; Jiang, S.; Xu, Z.; Jiang, Y.; Liu, L.; Jiang, J.; Tong, Y.; Wang, P. Crocetin ameliorates chronic restraint stress-induced depression-like behaviors in mice by regulating MEK/ERK pathways and gut microbiota. J. Ethnopharmacol. 2021, 268, 113608. [Google Scholar] [CrossRef] [PubMed]
- Tao, W.; Ruan, J.; Wu, R.; Zhao, M.; Zhao, T.; Qi, M.; Yau, S.S.Y.; Yao, G.; Zhang, H.; Hu, Y.; et al. A natural carotenoid crocin exerts antidepressant action by promoting adult hippocampal neurogenesis through Wnt/β-catenin signaling. J. Adv. Res. 2023, 43, 219–231. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.; Lu, L.; Wang, Q.; Liu, H.; Xue, W.; Zhou, T.; Xu, L.; Wang, K.; Wu, D.; Wei, F.; et al. Crocin Reverses Depression-Like Behavior in Parkinson Disease Mice via VTA-mPFC Pathway. Mol. Neurobiol. 2020, 57, 3158–3170. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.; Wei, P.C.; Chen, Q.; Chen, X.; Wang, S.C.; Li, J.R.; Gao, C. Functional and structural characterization of a β-glucosidase involved in saponin metabolism from intestinal bacteria. Biochem. Biophys. Res. Commun. 2018, 496, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Zimmermann-Kogadeeva, M.; Wegmann, R.; Goodman, A.L. Mapping human microbiome drug metabolism by gut bacteria and their genes. Nature 2019, 570, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Abenavoli, L.; Scarpellini, E.; Colica, C.; Boccuto, L.; Salehi, B.; Sharifi-Rad, J.; Aiello, V.; Romano, B.; De Lorenzo, A.; Izzo, A.A.; et al. Gut Microbiota and Obesity: A Role for Probiotics. Nutrients 2019, 11, 2690. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. New Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Golden, S.A.; Covington, H.E., 3rd; Berton, O.; Russo, S.J. A standardized protocol for repeated social defeat stress in mice. Nat. Protoc. 2011, 6, 1183–1191. [Google Scholar] [CrossRef] [PubMed]
- Slattery, D.A.; Cryan, J.F. Using the rat forced swim test to assess antidepressant-like activity in rodents. Nat. Protoc. 2012, 7, 1009–1014. [Google Scholar] [CrossRef] [PubMed]
- Castagné, V.; Moser, P.; Roux, S.; Porsolt, R.D. Rodent models of depression: Forced swim and tail suspension behavioral despair tests in rats and mice. Curr. Protoc. Neurosci. 2011, 49, 5–8. [Google Scholar] [CrossRef]
- Gao, X.; Cao, Q.; Cheng, Y.; Zhao, D.; Wang, Z.; Yang, H.; Wu, Q.; You, L.; Wang, Y.; Lin, Y.; et al. Chronic stress promotes colitis by disturbing the gut microbiota and triggering immune system response. Proc. Natl. Acad. Sci. USA 2018, 115, E2960–E2969. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cai, J.; Fan, P.; Dong, X.; Zhang, N.; Tai, J.; Cao, Y. Salidroside alleviates dextran sulfate sodium-induced colitis in mice by modulating the gut microbiota. Food Funct. 2023, 14, 7506–7519. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Chen, H.; Kang, D.; Shao, Y.; Shen, B.; Li, X.; Yin, X.; Zhu, Z.; Li, H.; Rao, T.; et al. Qualitatively and quantitatively investigating the regulation of intestinal microbiota on the metabolism of panax notoginseng saponins. J. Ethnopharmacol. 2016, 194, 324–336. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wang, H.; Huang, H.; Li, H.; Li, C.; Yan, C.; Yu, T.; Guo, H.; Hu, K.; Du, Y.; et al. Absolute quantitative analysis of endogenous neurotransmitters and amino acids by liquid chromatography-tandem mass spectrometry combined with multidimensional adsorption and collision energy defect. J. Chromatogr. A 2021, 1638, 461867. [Google Scholar] [CrossRef]
- Zhao, C.; Sun, R.; Cao, B.; Gu, S.; Zhao, J.; Liu, L.; Wang, X.; Zha, W.; Yu, X.; Xiao, W.; et al. An in vitro metabolic system of gut flora and the metabolism of ginsenoside Rg3 and cholic acid. Eur. J. Drug Metab. Pharmacokinet. 2014, 39, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yao, L.; Tang, C.; Jiang, J.; Ye, Y.; Liu, J. Qualitatively and quantitatively investigating the metabolism of 20(S)-protopanaxadiol-type ginsenosides by gut microbiota of different species. Biomed. Chromatogr. BMC 2021, 35, e5219. [Google Scholar] [CrossRef]
- Guo, W.; Kong, D.; Du, Y.; Shi, X.; Wang, W.; Wang, Y. Simultaneous determination of a novel diphenylpiperazine calcium channel blocker and its four metabolites in rat liver microsomes by liquid chromatography tandem mass spectrometry. Pharmacology 2012, 89, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Jin, W.; Fu, S.; Liu, K.; Xu, F.; Wu, L.; Xu, Y.; Yang, H.; Liu, H.; Wang, G.; et al. Strategies for mapping protein hydrolysate profiles and pharmacokinetics based on non-targeted proteomics combining skyline-aided quantitative techniques. Anal. Chim. Acta 2023, 1265, 341272. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Crocin | Crocetin |
|---|---|---|
| Tmax (h) | 1 | 6 |
| Cmax (μg/L) | 43.5 ± 8.6 | 4662.5 ± 586.1 |
| t1/2 (h) | 7.8 ± 1.7 | 9.4 ± 5.1 |
| AUC (μg·h/L) | 151 ± 20.8 | 33,451.9 ± 3323.6 |
| Vd (L/kg) | 12,574.45 ± 2684.7 | 86.4 ± 34.1 |
| CL (L/h/kg) | 1244 ± 285.4 | 7.8 ± 1.1 |
| Parameters | 100 mg/kg, i.g. | |||
|---|---|---|---|---|
| Crocin | Crocetin | |||
| Ctrl | ABX | Ctrl | ABX | |
| Tmax (h) | 2.8 ± 0.5 | 3.2 ± 0.8 | 7.3 ± 0.4 | 3.4 ± 0.6 * |
| Cmax (μg/L) | 198.7 ± 29.4 | 111.9 ± 11.5 * | 4441.8 ± 690.4 | 942.4 ± 166.6 * |
| t1/2 (h) | 2.1 ± 0.2 | 3.2 ± 0.2 * | 2.7 ± 0.2 | 10.4 ± 2.3 * |
| AUC (μg·h/L) | 1269.6 ± 154.7 | 738.9 ± 57.1 * | 32186.7 ± 4275.3 | 8292.4 ± 1066.4 * |
| Vd (L/kg) | 270.8 ± 46 | 696.9 ± 131.2 * | 13.8 ± 1.6 | 166.3 ± 42.5 * |
| CL (L/h/kg) | 90.4 ± 13.7 | 144.8 ± 16.8 * | 3.5 ± 0.4 | 11 ± 1.3 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, F.; Song, Y.; Wang, G.; Aa, J. Intestinal Metabolism of Crocin and a Pharmacokinetics and Pharmacodynamics Study in the Chronic Social Defeat Stress Mouse Model. Pharmaceuticals 2024, 17, 843. https://doi.org/10.3390/ph17070843
Xiao F, Song Y, Wang G, Aa J. Intestinal Metabolism of Crocin and a Pharmacokinetics and Pharmacodynamics Study in the Chronic Social Defeat Stress Mouse Model. Pharmaceuticals. 2024; 17(7):843. https://doi.org/10.3390/ph17070843
Chicago/Turabian StyleXiao, Fan, Yulong Song, Guangji Wang, and Jiye Aa. 2024. "Intestinal Metabolism of Crocin and a Pharmacokinetics and Pharmacodynamics Study in the Chronic Social Defeat Stress Mouse Model" Pharmaceuticals 17, no. 7: 843. https://doi.org/10.3390/ph17070843
APA StyleXiao, F., Song, Y., Wang, G., & Aa, J. (2024). Intestinal Metabolism of Crocin and a Pharmacokinetics and Pharmacodynamics Study in the Chronic Social Defeat Stress Mouse Model. Pharmaceuticals, 17(7), 843. https://doi.org/10.3390/ph17070843