
Accelerated Neuroimmune Dysfunction in Aged HIV-1-Infected Humanized Mice

 ,
,  ,
,  and
and {kind=link}
{kind=link}
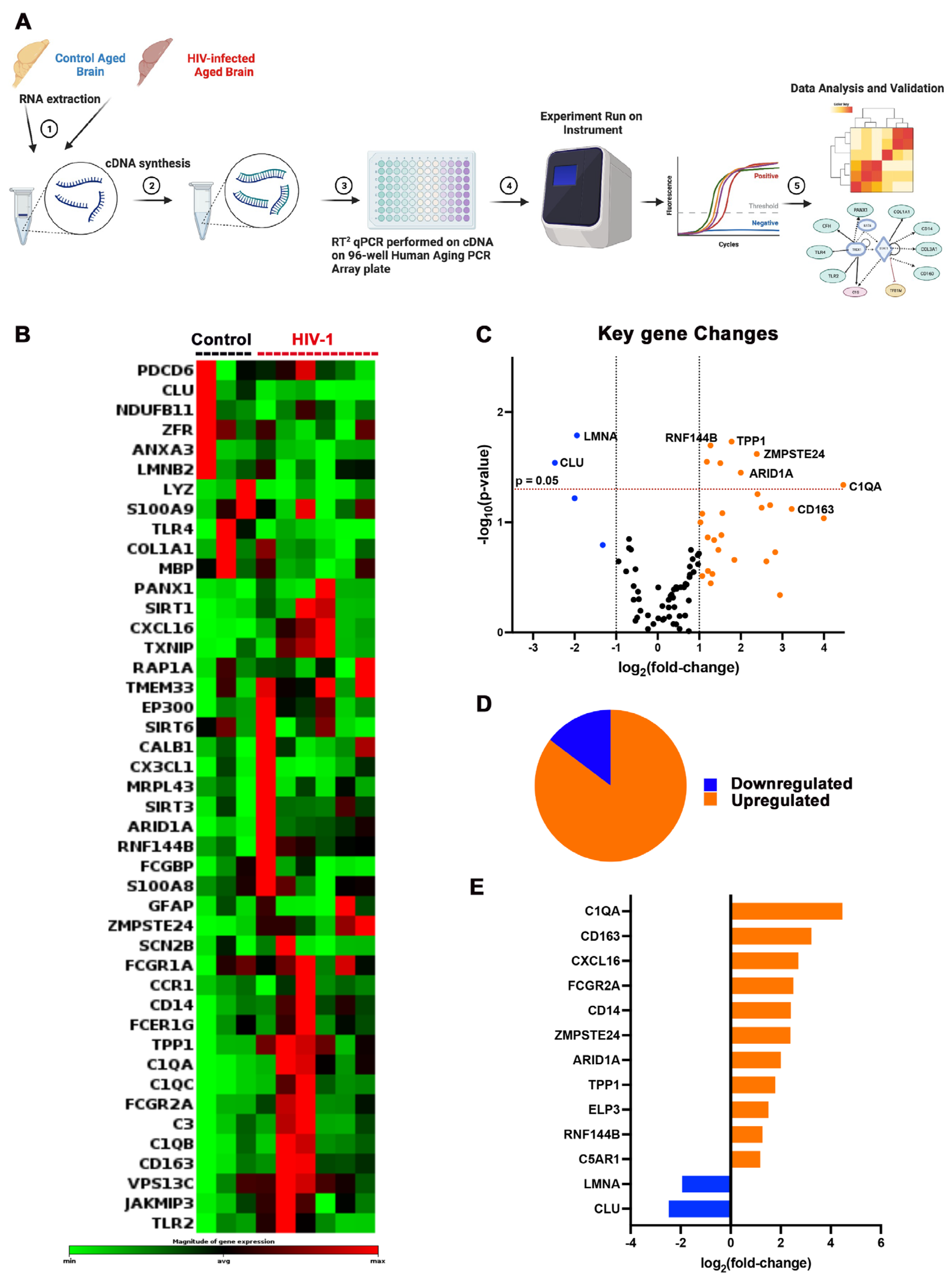{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Viral and Immune Profile in Aged HIV-1-Infected Humanized Mice (Hu-Mice)
2.2. Aging-Linked Biomarkers in HIV-1-Infected Hu-Mice
2.3. Pathway Enrichment Associated with Aged HIV-1-Infected Hu-Mice
2.4. Multispectral Immunofluorescence Imaging
3. Discussion
4. Materials and Methods
4.1. Generation and HIV-Infection of Hu-Mice
4.2. Flow Cytometry
4.3. Viral Load Measurements
4.4. Immunohistochemical and Immunofluorescent Staining
4.5. Nucleic Acid Extractions and qPCR Assays
4.6. cDNA Synthesis and RT2 Profiler PCR Array Specific for Human Aging
4.7. Transcriptomics Analyses
4.8. Statistical Evaluations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Calvanese, V.; Lara, E.; Kahn, A.; Fraga, M.F. The role of epigenetics in aging and age-related diseases. Ageing Res. Rev. 2009, 8, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Campisi, J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J. Gerontol. A Biol. Sci. Med. Sci. 2014, 69 (Suppl. S1), S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Smit, M.; Brinkman, K.; Geerlings, S.; Smit, C.; Thyagarajan, K.; van Sighem, A.; de Wolf, F.; Hallett, T.B. Future challenges for clinical care of an ageing population infected with HIV: A modelling study. Lancet Infect. Dis. 2015, 15, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Wing, E.J. HIV and aging. Int. J. Infect. Dis. 2016, 53, 61–68. [Google Scholar] [CrossRef]
- Roomaney, R.A.; van Wyk, B.; Pillay-van Wyk, V. Aging with HIV: Increased Risk of HIV Comorbidities in Older Adults. Int. J. Environ. Res. Public Health 2022, 19, 2359. [Google Scholar] [CrossRef] [PubMed]
- Abbondanzo, S.J.; Chang, S.L. HIV-1 transgenic rats display alterations in immunophenotype and cellular responses associated with aging. PLoS ONE 2014, 9, e105256. [Google Scholar] [CrossRef]
- Duffau, P.; Ozanne, A.; Bonnet, F.; Lazaro, E.; Cazanave, C.; Blanco, P.; Riviere, E.; Desclaux, A.; Hyernard, C.; Gensous, N.; et al. Multimorbidity, age-related comorbidities and mortality: Association of activation, senescence and inflammation markers in HIV adults. AIDS 2018, 32, 1651–1660. [Google Scholar] [CrossRef]
- Rosenthal, J.; Tyor, W. Aging, comorbidities, and the importance of finding biomarkers for HIV-associated neurocognitive disorders. J. Neurovirol. 2019, 25, 673–685. [Google Scholar] [CrossRef]
- Zhang, C.; Zaman, L.A.; Poluektova, L.Y.; Gorantla, S.; Gendelman, H.E.; Dash, P.K. Humanized Mice for Studies of HIV-1 Persistence and Elimination. Pathogens 2023, 12, 879. [Google Scholar] [CrossRef]
- Shultz, L.D.; Ishikawa, F.; Greiner, D.L. Humanized mice in translational biomedical research. Nat. Rev. Immunol. 2007, 7, 118–130. [Google Scholar] [CrossRef] [PubMed]
- Dash, P.K.; Gorantla, S.; Gendelman, H.E.; Knibbe, J.; Casale, G.P.; Makarov, E.; Epstein, A.A.; Gelbard, H.A.; Boska, M.D.; Poluektova, L.Y. Loss of neuronal integrity during progressive HIV-1 infection of humanized mice. J. Neurosci. 2011, 31, 3148–3157. [Google Scholar] [CrossRef] [PubMed]
- Gorantla, S.; Makarov, E.; Finke-Dwyer, J.; Castanedo, A.; Holguin, A.; Gebhart, C.L.; Gendelman, H.E.; Poluektova, L. Links between progressive HIV-1 infection of humanized mice and viral neuropathogenesis. Am. J. Pathol. 2010, 177, 2938–2949. [Google Scholar] [CrossRef]
- Boska, M.D.; Dash, P.K.; Knibbe, J.; Epstein, A.A.; Akhter, S.P.; Fields, N.; High, R.; Makarov, E.; Bonasera, S.; Gelbard, H.A.; et al. Associations between brain microstructures, metabolites, and cognitive deficits during chronic HIV-1 infection of humanized mice. Mol. Neurodegener. 2014, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Honeycutt, J.B.; Sheridan, P.A.; Matsushima, G.K.; Garcia, J.V. Humanized mouse models for HIV-1 infection of the CNS. J. Neurovirol. 2015, 21, 301–309. [Google Scholar] [CrossRef]
- Waight, E.; Zhang, C.; Mathews, S.; Kevadiya, B.D.; Lloyd, K.C.K.; Gendelman, H.E.; Gorantla, S.; Poluektova, L.Y.; Dash, P.K. Animal models for studies of HIV-1 brain reservoirs. J. Leukoc. Biol. 2022, 112, 1285–1295. [Google Scholar] [CrossRef]
- Epstein, A.A.; Narayanasamy, P.; Dash, P.K.; High, R.; Bathena, S.P.; Gorantla, S.; Poluektova, L.Y.; Alnouti, Y.; Gendelman, H.E.; Boska, M.D. Combinatorial assessments of brain tissue metabolomics and histopathology in rodent models of human immunodeficiency virus infection. J. Neuroimmune Pharmacol. 2013, 8, 1224–1238. [Google Scholar] [CrossRef]
- Dash, P.K.; Chen, C.; Kaminski, R.; Su, H.; Mancuso, P.; Sillman, B.; Zhang, C.; Liao, S.; Sravanam, S.; Liu, H.; et al. CRISPR editing of CCR5 and HIV-1 facilitates viral elimination in antiretroviral drug-suppressed virus-infected humanized mice. Proc. Natl. Acad. Sci. USA 2023, 120, e2217887120. [Google Scholar] [CrossRef]
- Goodkin, K.; Wilkie, F.L.; Concha, M.; Asthana, D.; Shapshak, P.; Douyon, R.; Fujimura, R.K.; LoPiccolo, C. Subtle neuropsychological impairment and minor cognitive-motor disorder in HIV-1 infection. Neuroradiological, neurophysiological, neuroimmunological, and virological correlates. Neuroimaging Clin. N. Am. 1997, 7, 561–579. [Google Scholar]
- Antinori, A.; Arendt, G.; Becker, J.T.; Brew, B.J.; Byrd, D.A.; Cherner, M.; Clifford, D.B.; Cinque, P.; Epstein, L.G.; Goodkin, K.; et al. Updated research nosology for HIV-associated neurocognitive disorders. Neurology 2007, 69, 1789–1799. [Google Scholar] [CrossRef]
- Benevides, M.; Filho, S.B.; Debona, R.; Bergamaschi, E.N.C.; Nunes, J.C. Prevalence of Peripheral Neuropathy and associated factors in HIV-infected patients. J. Neurol. Sci. 2017, 375, 316–320. [Google Scholar] [CrossRef]
- Gray, F.; Scaravilli, F.; Everall, I.; Chretien, F.; An, S.; Boche, D.; Adle-Biassette, H.; Wingertsmann, L.; Durigon, M.; Hurtrel, B.; et al. Neuropathology of early HIV-1 infection. Brain Pathol. 1996, 6, 1–15. [Google Scholar] [CrossRef]
- Meir-Shafrir, K.; Pollack, S. Accelerated Aging in HIV Patients. Rambam Maimonides Med. J. 2012, 3, e0025. [Google Scholar] [CrossRef]
- Horvath, S.; Levine, A.J. HIV-1 Infection Accelerates Age According to the Epigenetic Clock. J. Infect. Dis. 2015, 212, 1563–1573. [Google Scholar] [CrossRef]
- Angelovich, T.A.; Hearps, A.C.; Maisa, A.; Martin, G.E.; Lichtfuss, G.F.; Cheng, W.J.; Palmer, C.S.; Landay, A.L.; Crowe, S.M.; Jaworowski, A. Viremic and Virologically Suppressed HIV Infection Increases Age-Related Changes to Monocyte Activation Equivalent to 12 and 4 Years of Aging, Respectively. J. Acquir. Immune Defic. Syndr. 2015, 69, 11–17. [Google Scholar] [CrossRef]
- Rothermundt, M.; Peters, M.; Prehn, J.H.; Arolt, V. S100B in brain damage and neurodegeneration. Microsc. Res. Tech. 2003, 60, 614–632. [Google Scholar] [CrossRef]
- Cohen, J.; Torres, C. HIV-associated cellular senescence: A contributor to accelerated aging. Ageing Res. Rev. 2017, 36, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Martin-Iguacel, R.; Llibre, J.M.; Friis-Moller, N. Risk of Cardiovascular Disease in an Aging HIV Population: Where Are We Now? Curr. HIV/AIDS Rep. 2015, 12, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Borges, Á.H.; Silverberg, M.J.; Wentworth, D.; Grulich, A.E.; Fätkenheuer, G.; Mitsuyasu, R.; Tambussi, G.; Sabin, C.A.; Neaton, J.D.; Lundgren, J.D. Predicting risk of cancer during HIV infection: The role of inflammatory and coagulation biomarkers. AIDS 2013, 27, 1433–1441. [Google Scholar] [CrossRef]
- Gupta, S.K.; Kitch, D.; Tierney, C.; Melbourne, K.; Ha, B.; McComsey, G.A. Markers of renal disease and function are associated with systemic inflammation in HIV infection. HIV Med. 2015, 16, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Warriner, A.H.; Burkholder, G.A.; Overton, E.T. HIV-related metabolic comorbidities in the current ART era. Infect. Dis. Clin. N. Am. 2014, 28, 457–476. [Google Scholar] [CrossRef] [PubMed]
- The Lancet Infectious Diseases. The challenge of HIV associated neurocognitive disorder. Lancet Infect. Dis. 2013, 13, 907. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.; Hellmuth, J.; Spudich, S.; Valcour, V. Cognitive Impairment and Persistent CNS Injury in Treated HIV. Curr. HIV/AIDS Rep. 2016, 13, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Schank, M.; Zhao, J.; Wang, L.; Nguyen, L.N.T.; Zhang, Y.; Wu, X.Y.; Zhang, J.; Jiang, Y.; Ning, S.; El Gazzar, M.; et al. ROS-Induced Mitochondrial Dysfunction in CD4 T Cells from ART-Controlled People Living with HIV. Viruses 2023, 15, 1061. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Longchamps, R.J.; Piggott, D.A.; Castellani, C.A.; Sumpter, J.A.; Brown, T.T.; Mehta, S.H.; Arking, D.E.; Kirk, G.D. Association between HIV Infection and Mitochondrial DNA Copy Number in Peripheral Blood: A Population-Based, Prospective Cohort Study. J. Infect. Dis. 2019, 219, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- ToVinh, M.; Hörr, G.; Dobrikova, K.; Gotter, C.; Rommel, C.; Hoffmeister, C.; Raabe, J.; Kaiser, K.M.; Finnemann, C.; Bischoff, J.; et al. Mitochondrial Dysfunction Contributes to Impaired Cytokine Production of CD56bright Natural Killer Cells from Human Immunodeficiency Virus-Infected Individuals Under Effective Antiretroviral Therapy. J. Infect. Dis. 2022, 226, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Willig, A.L.; Overton, E.T. Metabolic Complications and Glucose Metabolism in HIV Infection: A Review of the Evidence. Curr. HIV/AIDS Rep. 2016, 13, 289–296. [Google Scholar] [CrossRef]
- Premeaux, T.A.; Ndhlovu, L.C. Decrypting biological hallmarks of aging in people with HIV. Curr. Opin. HIV AIDS 2023, 18, 237–245. [Google Scholar] [CrossRef]
- Aberg, J.A. Aging, inflammation, and HIV infection. Top. Antivir. Med. 2012, 20, 101–105. [Google Scholar]
- Deeks, S.G. HIV infection, inflammation, immunosenescence, and aging. Ann. Rev. Med. 2011, 62, 141–155. [Google Scholar] [CrossRef]
- Sundermann, E.E.; Hussain, M.A.; Moore, D.J.; Horvath, S.; Lin, D.T.S.; Kobor, M.S.; Levine, A. Inflammation-related genes are associated with epigenetic aging in HIV. J. Neurovirol. 2019, 25, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Alcaide, M.L.; Parmigiani, A.; Pallikkuth, S.; Roach, M.; Freguja, R.; Della Negra, M.; Bolivar, H.; Fischl, M.A.; Pahwa, S. Immune activation in HIV-infected aging women on antiretrovirals--implications for age-associated comorbidities: A cross-sectional pilot study. PLoS ONE 2013, 8, e63804. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.L.; de Boer, R.; Brul, S.; Budovskaya, Y.; van Spek, H. Premature and accelerated aging: HIV or HAART? Front. Genet. 2012, 3, 328. [Google Scholar] [CrossRef]
- Knibbe-Hollinger, J.S.; Fields, N.R.; Chaudoin, T.R.; Epstein, A.A.; Makarov, E.; Akhter, S.P.; Gorantla, S.; Bonasera, S.J.; Gendelman, H.E.; Poluektova, L.Y. Influence of age, irradiation and humanization on NSG mouse phenotypes. Biol. Open 2015, 4, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Hearps, A.C.; Martin, G.E.; Angelovich, T.A.; Cheng, W.J.; Maisa, A.; Landay, A.L.; Jaworowski, A.; Crowe, S.M. Aging is associated with chronic innate immune activation and dysregulation of monocyte phenotype and function. Aging Cell 2012, 11, 867–875. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Cui, Y.; Hou, L.; Bian, Z.; Yang, Z.; Xu, R.; Jia, Y.; Wu, Z.; Yang, X. Diffusion kurtosis imaging-derived histogram metrics for prediction of resistance to neoadjuvant chemoradiotherapy in rectal adenocarcinoma: Preliminary findings. Eur. J. Radiol. 2021, 144, 109963. [Google Scholar] [CrossRef] [PubMed]
- Cassol, E.; Misra, V.; Dutta, A.; Morgello, S.; Gabuzda, D. Cerebrospinal fluid metabolomics reveals altered waste clearance and accelerated aging in HIV patients with neurocognitive impairment. AIDS 2014, 28, 1579–1591. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.J.; Chenna, A.; Petropoulos, C.J.; Lie, Y.; Curanovic, D.; Crescini, M.; Winslow, J.; Sundermann, E.; Tang, B.; Letendre, S.L. Higher cerebrospinal fluid biomarkers of neuronal injury in HIV-associated neurocognitive impairment. J. Neurovirol. 2022, 28, 438–445. [Google Scholar] [CrossRef]
- Mäkitalo, S.; Mellgren, Å.; Borgh, E.; Kilander, L.; Skillbäck, T.; Zetterberg, H.; Gisslén, M. The cerebrospinal fluid biomarker profile in an HIV-infected subject with Alzheimer’s disease. AIDS Res. Ther. 2015, 12, 23. [Google Scholar] [CrossRef]
- Shive, C.L.; Jiang, W.; Anthony, D.D.; Lederman, M.M. Soluble CD14 is a nonspecific marker of monocyte activation. Aids 2015, 29, 1263–1265. [Google Scholar] [CrossRef]
- Lyons, J.L.; Uno, H.; Ancuta, P.; Kamat, A.; Moore, D.J.; Singer, E.J.; Morgello, S.; Gabuzda, D. Plasma sCD14 is a biomarker associated with impaired neurocognitive test performance in attention and learning domains in HIV infection. J. Acquir. Immune Defic. Syndr. 2011, 57, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Poesen, R.; Ramezani, A.; Claes, K.; Augustijns, P.; Kuypers, D.; Barrows, I.R.; Muralidharan, J.; Evenepoel, P.; Meijers, B.; Raj, D.S. Associations of Soluble CD14 and Endotoxin with Mortality, Cardiovascular Disease, and Progression of Kidney Disease among Patients with CKD. Clin. J. Am. Soc. Nephrol. 2015, 10, 1525–1533. [Google Scholar] [CrossRef] [PubMed]
- Castley, A.; Williams, L.; James, I.; Guelfi, G.; Berry, C.; Nolan, D. Plasma CXCL10, sCD163 and sCD14 Levels Have Distinct Associations with Antiretroviral Treatment and Cardiovascular Disease Risk Factors. PLoS ONE 2016, 11, e0158169. [Google Scholar] [CrossRef] [PubMed]
- Lichterfeld, M.; Cung, T.; Seiss, K.; Rosenberg, E.S.; Pereyra, F.; Yu, X.G. Shelterin dysfunction and p16(INK4a)-mediated growth inhibition in HIV-1-specific CD8 T cells. J. Virol. 2012, 86, 5533–5540. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Zhao, J.; Cao, D.; Dang, X.; Wang, L.; Lian, J.; Zhang, Y.; Jia, Z.; Wu, X.Y.; Morrison, Z.; et al. Inhibition of TRF2 accelerates telomere attrition and DNA damage in naïve CD4 T cells during HCV infection. Cell Death Dis. 2018, 9, 900. [Google Scholar] [CrossRef]
- Messner, M.; Ghadge, S.K.; Maurer, T.; Graber, M.; Staggl, S.; Christine Maier, S.; Pölzl, G.; Zaruba, M.M. ZMPSTE24 Is Associated with Elevated Inflammation and Progerin mRNA. Cells 2020, 9, 1981. [Google Scholar] [CrossRef]
- Caron-Debarle, M.; Lagathu, C.; Boccara, F.; Vigouroux, C.; Capeau, J. HIV-associated lipodystrophy: From fat injury to premature aging. Trends Mol. Med. 2010, 16, 218–229. [Google Scholar] [CrossRef]
- Mlambo, Z.P.; Varaden, D.; Moodley, J.; Naicker, T. Are concentrations of clusterin and beta-2-glycoprotein I dysregulated in HIV associated preeclampsia? Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 251, 1–7. [Google Scholar] [CrossRef]
- Torres-Muñoz, J.E.; Redondo, M.; Czeisler, C.; Roberts, B.; Tacoronte, N.; Petito, C.K. Upregulation of glial clusterin in brains of patients with AIDs. Brain Res. 2001, 888, 297–301. [Google Scholar] [CrossRef]
- Mogensen, T.H.; Melchjorsen, J.; Larsen, C.S.; Paludan, S.R. Innate immune recognition and activation during HIV infection. Retrovirology 2010, 7, 54. [Google Scholar] [CrossRef]
- Mohan, T.; Bhatnagar, S.; Gupta, D.L.; Rao, D.N. Current understanding of HIV-1 and T-cell adaptive immunity: Progress to date. Microb. Pathog. 2014, 73, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Manches, O.; Frleta, D.; Bhardwaj, N. Dendritic cells in progression and pathology of HIV infection. Trends Immunol. 2014, 35, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Altfeld, M.; Fadda, L.; Frleta, D.; Bhardwaj, N. DCs and NK cells: Critical effectors in the immune response to HIV-1. Nat. Rev. Immunol. 2011, 11, 176–186. [Google Scholar] [CrossRef]
- Morrison, J.H.; Baxter, M.G. The ageing cortical synapse: Hallmarks and implications for cognitive decline. Nat. Rev. Neurosci. 2012, 13, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Levi, G.; Minghetti, L.; Aloisi, F. Regulation of prostanoid synthesis in microglial cells and effects of prostaglandin E2 on microglial functions. Biochimie 1998, 80, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Fiebich, B.L.; Schleicher, S.; Spleiss, O.; Czygan, M.; Hüll, M. Mechanisms of prostaglandin E2-induced interleukin-6 release in astrocytes: Possible involvement of EP4-like receptors, p38 mitogen-activated protein kinase and protein kinase C. J. Neurochem. 2001, 79, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Wu, L.; Wang, Q.; Hand, T.; Bilak, M.; McCullough, L.; Andreasson, K. Function of COX-2 and prostaglandins in neurological disease. J. Mol. Neurosci. 2007, 33, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Perry, C.J.; Tsui, Y.C.; Staron, M.M.; Parish, I.A.; Dominguez, C.X.; Rosenberg, D.W.; Kaech, S.M. Prostaglandin E2 and programmed cell death 1 signaling coordinately impair CTL function and survival during chronic viral infection. Nat. Med. 2015, 21, 327–334. [Google Scholar] [CrossRef]
- Gorantla, S.; Sneller, H.; Walters, L.; Sharp, J.G.; Pirruccello, S.J.; West, J.T.; Wood, C.; Dewhurst, S.; Gendelman, H.E.; Poluektova, L. Human immunodeficiency virus type 1 pathobiology studied in humanized BALB/c-Rag2-/-gammac-/-mice. J. Virol. 2007, 81, 2700–2712. [Google Scholar] [CrossRef]
- Su, H.; Sravanam, S.; Sillman, B.; Waight, E.; Makarov, E.; Mathews, S.; Poluektova, L.Y.; Gorantla, S.; Gendelman, H.E.; Dash, P.K. Recovery of Latent HIV-1 from Brain Tissue by Adoptive Cell Transfer in Virally Suppressed Humanized Mice. J. Neuroimmune Pharmacol. 2021, 16, 796–805. [Google Scholar] [CrossRef]
- Dash, P.K.; Kaminski, R.; Bella, R.; Su, H.; Mathews, S.; Ahooyi, T.M.; Chen, C.; Mancuso, P.; Sariyer, R.; Ferrante, P.; et al. Sequential LASER ART and CRISPR Treatments Eliminate HIV-1 in a Subset of Infected Humanized Mice. Nat. Commun. 2019, 10, 2753. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Cheng, Y.; Sravanam, S.; Mathews, S.; Gorantla, S.; Poluektova, L.Y.; Dash, P.K.; Gendelman, H.E. Immune Activations and Viral Tissue Compartmentalization During Progressive HIV-1 Infection of Humanized Mice. Front. Immunol. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Arainga, M.; Su, H.; Poluektova, L.Y.; Gorantla, S.; Gendelman, H.E. HIV-1 cellular and tissue replication patterns in infected humanized mice. Sci. Rep. 2016, 6, 23513. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Su, H.; Waight, E.; Poluektova, L.Y.; Gorantla, S.; Gendelman, H.E.; Dash, P.K. Accelerated Neuroimmune Dysfunction in Aged HIV-1-Infected Humanized Mice. Pharmaceuticals 2024, 17, 149. https://doi.org/10.3390/ph17020149
Zhang C, Su H, Waight E, Poluektova LY, Gorantla S, Gendelman HE, Dash PK. Accelerated Neuroimmune Dysfunction in Aged HIV-1-Infected Humanized Mice. Pharmaceuticals. 2024; 17(2):149. https://doi.org/10.3390/ph17020149
Chicago/Turabian StyleZhang, Chen, Hang Su, Emiko Waight, Larisa Y. Poluektova, Santhi Gorantla, Howard E. Gendelman, and Prasanta K. Dash. 2024. "Accelerated Neuroimmune Dysfunction in Aged HIV-1-Infected Humanized Mice" Pharmaceuticals 17, no. 2: 149. https://doi.org/10.3390/ph17020149
APA StyleZhang, C., Su, H., Waight, E., Poluektova, L. Y., Gorantla, S., Gendelman, H. E., & Dash, P. K. (2024). Accelerated Neuroimmune Dysfunction in Aged HIV-1-Infected Humanized Mice. Pharmaceuticals, 17(2), 149. https://doi.org/10.3390/ph17020149