
Synthesis and Bioevaluation of New Stable Derivatives of Chrysin-8-C-Glucoside That Modulate the Antioxidant Keap1/Nrf2/HO-1 Pathway in Human Macrophages

 , , , , ,
, , , , ,  , and
, and 
Abstract

1. Introduction
2. Results
2.1. Isolation and Chemical Characterization of 8-C-β-D-Glucopyranosylchrysin (1) and Preparation of Derivatives 1a and 1b
2.2. Antioxidant Scavenging Activity
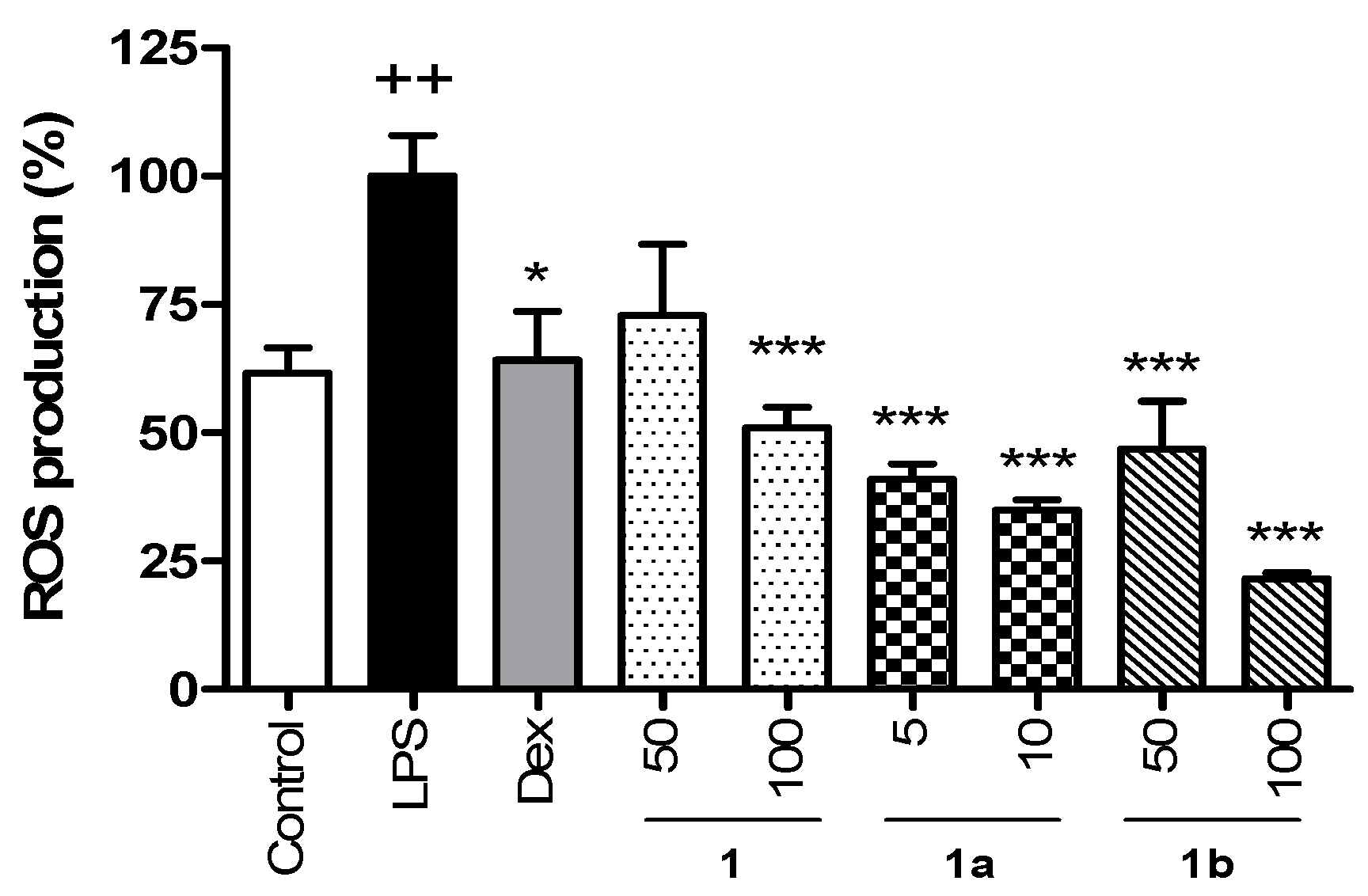
2.3. Intracellular ROS Measurements
2.4. Pro-Inflammatory Cytokine Production
2.5. Anti-Inflammatory and Antioxidant Protein Expression
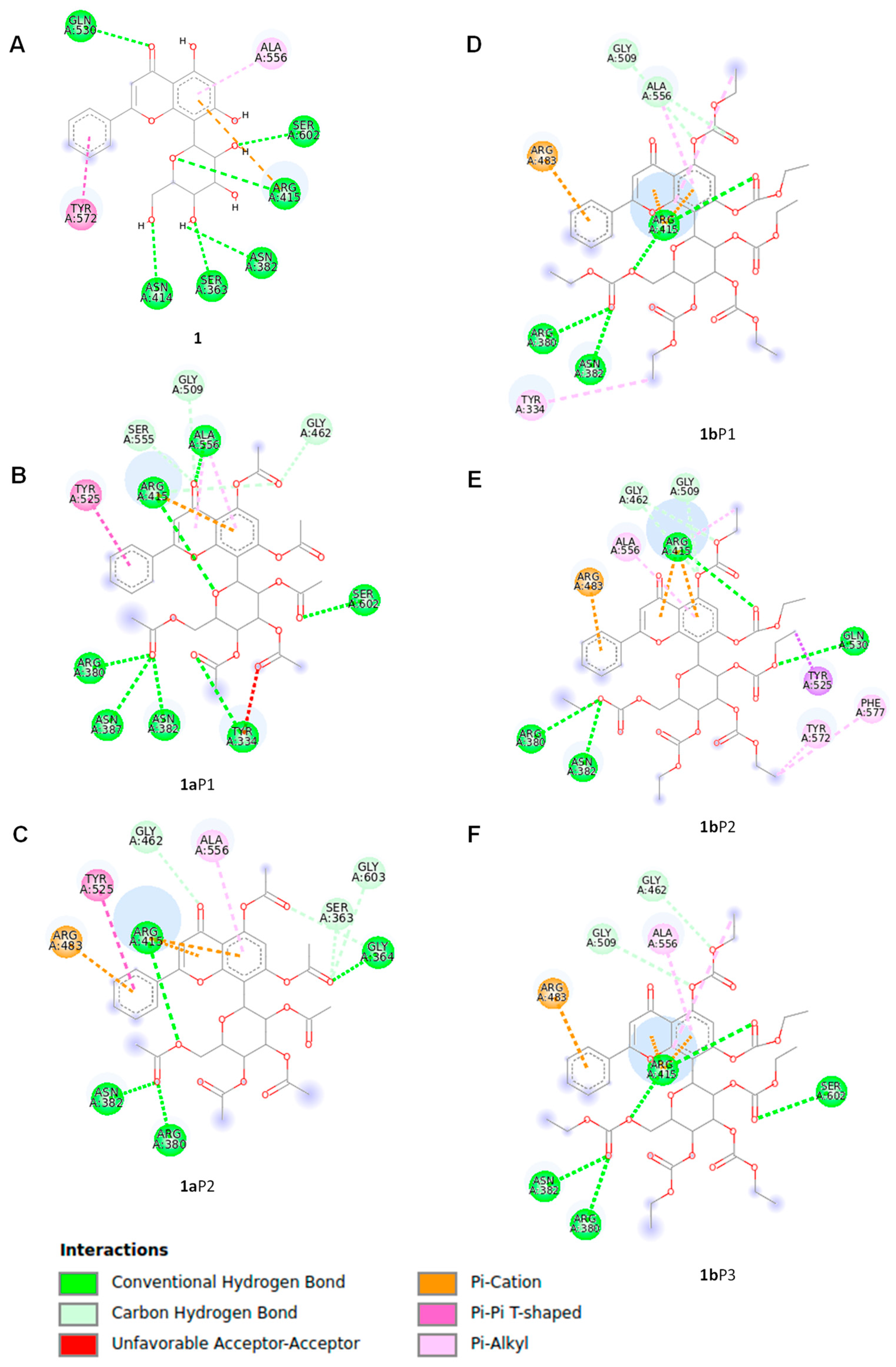
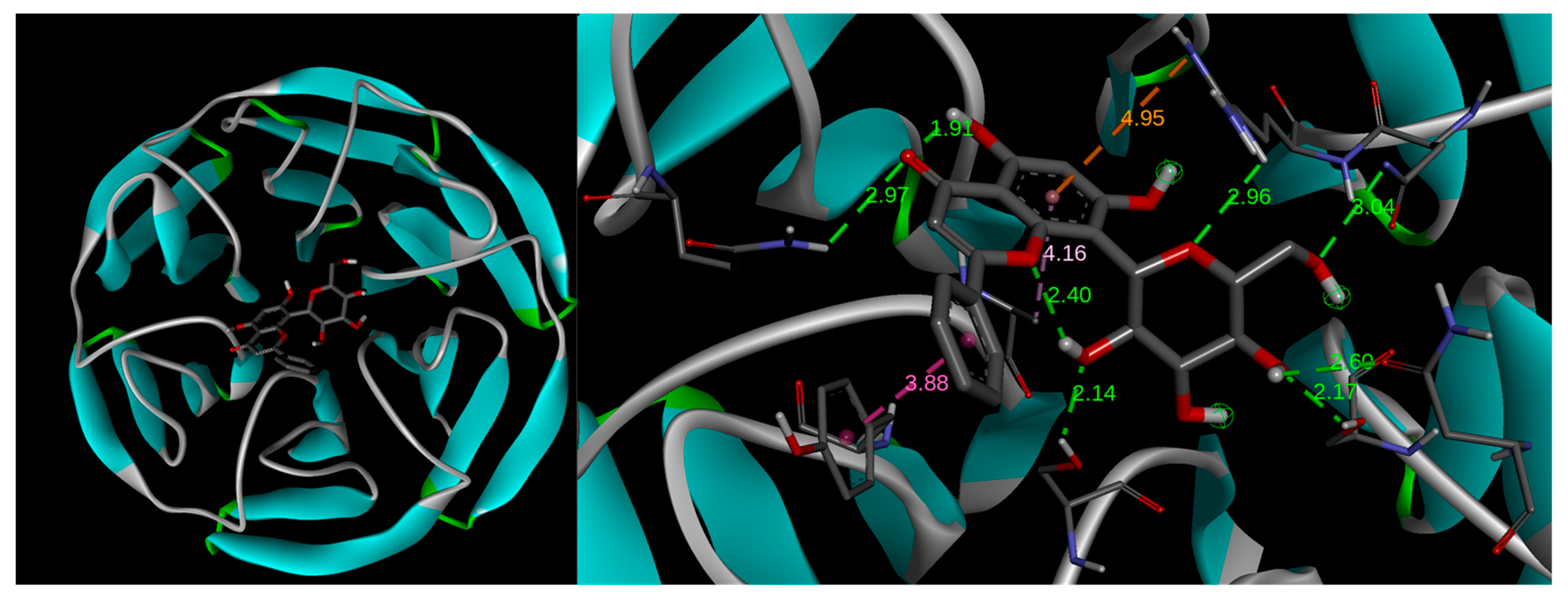
2.6. Docking Study
2.7. In Silico Analysis of the Physicochemical Properties of 1 and 1a
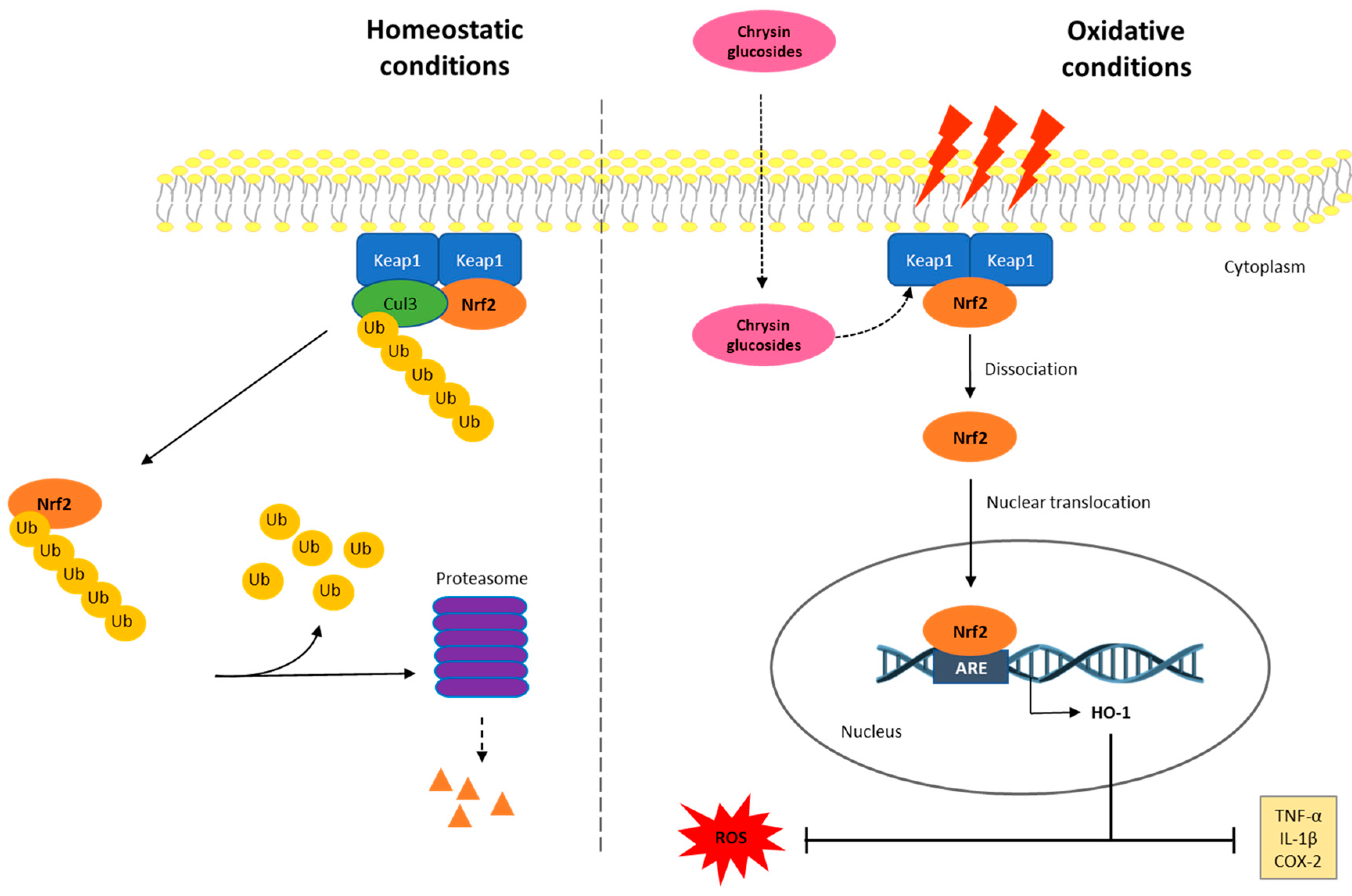
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Extraction and Isolation of 8-C-β-D-Glucopyranosylchrysin (1)
4.3. General Procedure of Acetylation and Ethyl Carbonate Derivatizations
4.4. Cell Culture
4.5. Cytotoxicity Assay
4.6. ABTS Radical Scavenging Assay
4.7. Intracellular ROS Production
4.8. Determination of Pro-Inflammatory Cytokine Levels
4.9. Protein Expression Measurement
4.10. Docking Analysis and Study of Permeability
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2018, 9, 7204–7218. [Google Scholar] [CrossRef]
- Bourgonje, A.R.; Feelisch, M.; Faber, K.N.; Pasch, A.; Dijkstra, G.; van Goor, H. Oxidative Stress and Redox-Modulating Therapeutics in Inflammatory Bowel Disease. Trends Mol. Med. 2020, 26, 1034–1046. [Google Scholar] [CrossRef]
- Wu, S.; Liao, X.; Zhu, Z.; Huang, R.; Chen, M.; Huang, A.; Zhang, J.; Wu, Q.; Wang, J.; Ding, Y. Antioxidant and anti-inflammation effects of dietary phytochemicals: The Nrf2/NF-κB signalling pathway and upstream factors of Nrf2. Phytochemistry 2022, 204, 113429. [Google Scholar] [CrossRef]
- Davinelli, S.; Medoro, A.; Intrieri, M.; Saso, L.; Scapagnini, G.; Kang, J.X. Targeting NRF2–KEAP1 axis by Omega-3 fatty acids and their derivatives: Emerging opportunities against aging and diseases. Free Radic. Biol. Med. 2022, 193, 736–750. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Sahana, G.R.; Nagella, P.; Joseph, B.V.; Alessa, F.M.; Al-Mssallem, M.Q. Flavonoids as Potential Anti-Inflammatory Molecules: A Review. Molecules 2022, 27, 2901. [Google Scholar] [CrossRef]
- Li, G.; Ding, K.; Qiao, Y.; Zhang, L.; Zheng, L.; Pan, T.; Zhang, L. Flavonoids regulate inflammation and oxidative stress in cancer. Molecules 2020, 25, 5628. [Google Scholar] [CrossRef]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and human health: The role of bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef]
- Khan, A.U.; Dagur, H.S.; Khan, M.; Malik, N.; Alam, M.; Mushtaque, M. Therapeutic role of flavonoids and flavones in cancer prevention: Current trends and future perspectives. Eur. J. Med. Chem. Rep. 2021, 3, 100010. [Google Scholar] [CrossRef]
- Slika, H.; Mansour, H.; Wehbe, N.; Nasser, S.A.; Iratni, R.; Nasrallah, G.; Shaito, A.; Ghaddar, T.; Kobeissy, F.; Eid, A.H. Therapeutic potential of flavonoids in cancer: ROS-mediated mechanisms. Biomed. Pharmacother. 2022, 146, 112442. [Google Scholar] [CrossRef]
- Lin, D.; Jiang, S.; Zhang, A.; Wu, T.; Qian, Y.; Shao, Q. Structural derivatization strategies of natural phenols by semi-synthesis and total-synthesis. Nat. Prod. Bioprospect. 2022, 12, 8. [Google Scholar] [CrossRef]
- Xie, L.; Deng, Z.; Zhang, J.; Dong, H.; Wang, W.; Xing, B.; Liu, X. Comparison of Flavonoid O-Glycoside, C-Glycoside and Their Aglycones on Antioxidant Capacity and Metabolism during In Vitro Digestion and In Vivo. Foods 2022, 11, 882. [Google Scholar] [CrossRef]
- Stompor-gorący, M.; Bajek-bil, A.; Machaczka, M. Chrysin: Perspectives on contemporary status and future possibilities as pro-health agent. Nutrients 2021, 13, 2038. [Google Scholar] [CrossRef]
- Talebi, M.; Talebi, M.; Farkhondeh, T.; Samarghandian, S. Molecular mechanism-based therapeutic properties of honey. Biomed. Pharmacother. 2020, 130, 110590. [Google Scholar] [CrossRef] [PubMed]
- Liew, K.Y.; Kamise, N.I.; Ong, H.M.; Aw Yong, P.Y.; Islam, F.; Tan, J.W.; Tham, C.L. Anti-Allergic Properties of Propolis: Evidence From Preclinical and Clinical Studies. Front. Pharmacol. 2022, 12, 785371. [Google Scholar] [CrossRef] [PubMed]
- Quevedo-Tinoco, L.; Rodríguez-García, G.; del Río, R.E.; Talero, E.; Motilva, V.; Cerda-García-Rojas, C.M.; Joseph-Nathan, P.; Gómez-Hurtado, M.A. Strategy for the vibrational circular dichroism study of a glycosylflavonoid evaluated as its peracetate. The case of bioactive 7-O-β-D-glucopyranosylchrysin. J. Mol. Struct. 2021, 1225, 129147. [Google Scholar] [CrossRef]
- Liu, H.; Mou, Y.; Zhao, J.; Wang, J.; Zhou, L.; Wang, M.; Wang, D.; Han, J.; Yu, Z.; Yang, F. Flavonoids from halostachys caspica and their antimicrobial and antioxidant activities. Molecules 2010, 15, 7933–7945. [Google Scholar] [CrossRef]
- Cherkaoui-Tangi, K.; Lachkar, M.; Wibo, M.; Morel, N.; Gilani, A.H.; Lyoussi, B. Pharmacological studies on hypotensive, diuretic and vasodilator activities of chrysin glucoside from Calycotome villosa in rats. Phyther. Res. 2008, 22, 356–361. [Google Scholar] [CrossRef]
- Berthier, A.; Girard, C.; Grandvuillemin, A.; Muyard, F.; Skaltsounis, A.L.; Jouvenot, M.; Delage-Mourroux, R. Effect of 7-O-beta-D-glucopyranosylchrysin and its aglycone chrysin isolated from Podocytisus caramanicus on estrogen receptor alpha transcriptional activity. Planta Med. 2007, 73, 1447–1451. [Google Scholar] [CrossRef]
- Ji, S.; Li, R.; Wang, Q.; Miao, W.J.; Li, Z.W.; Si, L.L.; Qiao, X.; Yu, S.W.; Zhou, D.M.; Ye, M. Anti-H1N1 virus, cytotoxic and Nrf2 activation activities of chemical constituents from Scutellaria baicalensis. J. Ethnopharmacol. 2015, 176, 475–484. [Google Scholar] [CrossRef]
- Jiang, H.-Y.; Gu, W.-W.; Gan, J.; Yang, Q.; Shi, Y.; Lian, W.-B.; Xu, H.-R.; Yang, S.-H.; Yang, L.; Zhang, X.; et al. MNSFβ promotes LPS-induced TNFα expression by increasing the localization of RC3H1 to stress granules, and the interfering peptide HEPN2 reduces TNFα production by disrupting the MNSFβ-RC3H1 interaction in macrophages. Int. Immunopharmacol. 2024, 142, 113053. [Google Scholar] [CrossRef]
- Li, H.; Liang, J.; Li, P.; Li, X.; Liu, Q.; Yang, S.; Zhang, C.; Liu, S.; He, Y.; Tan, C. Schizonepeta tenuifolia Briq-Saposhnikovia divaricata decoction alleviates atopic dermatitis via downregulating macrophage TRPV1. Front. Pharmacol. 2024, 15, 1413513. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Román, J.; Talero, E.; de los Reyes, C.; García-Mauriño, S.; Motilva, V. Microalgae-derived oxylipins decrease inflammatory mediators by regulating the subcellular location of NFκB and PPAR-γ. Pharmacol. Res. 2018, 128, 220–230. [Google Scholar] [CrossRef]
- López-García, E.; Marín-Gordillo, A.; Sánchez-Hidalgo, M.; Ávila-Román, J.; Romero-Gil, V.; Bermúdez-Oria, A.; Benítez-Cabello, A.; Garrido-Fernández, A.; Rodríguez-Gómez, F.; Arroyo-López, F.N. Functional features of the exopolysaccharide extracts produced by Lactiplantibacillus strains isolated from table olives. Food Funct. 2024, 15, 1938–1947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-Y.; Guo, Y.-Z.; Ageta, H.; Harigaya, Y.; Onda, M.; Hashimoto, K.; Ikeya, Y.; Okada, M.; Maruno, M. A New Flavone C-glycoside from Scutellaria baicalensis. J. Chin. Pharm. Sci. 1997, 6, 182–185. [Google Scholar]
- Pallesen, J.S.; Narayanan, D.; Tran, K.T.; Solbak, S.M.Ø.; Marseglia, G.; Sørensen, L.M.E.; Høj, L.J.; Munafò, F.; Carmona, R.M.C.; Garcia, A.D.; et al. Deconstructing Noncovalent Kelch-like ECH-Associated Protein 1 (Keap1) Inhibitors into Fragments to Reconstruct New Potent Compounds. J. Med. Chem. 2021, 64, 4623–4661. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Song, H.Y.; Sik Kim, W.; Kim, J.M.; Bak, D.H.; Moo Han, J.; Lim, S.T.; Byun, E.B. A hydroxyethyl derivative of chrysin exhibits anti-inflammatory activity in dendritic cells and protective effects against dextran sodium salt-induced colitis in mice. Int. Immunopharmacol. 2019, 77, 105958. [Google Scholar] [CrossRef]
- Lim, H.; Jin, J.H.; Park, H.; Kim, H.P. New synthetic anti-inflammatory chrysin analog, 5,7-dihydroxy-8-(pyridine-4yl)flavone. Eur. J. Pharmacol. 2011, 670, 617–622. [Google Scholar] [CrossRef]
- Pałasz, A.; Cież, D.; Trzewik, B.; Miszczak, K.; Tynor, G.; Bazan, B. In the Search of Glycoside-Based Molecules as Antidiabetic Agents. Top. Curr. Chem. 2019, 377, 19. [Google Scholar] [CrossRef]
- Adesina, A.F.; Adewuyi, A.; Otuechere, C.A. Exploratory studies on chrysin via antioxidant, antimicrobial, ADMET, PASS and molecular docking evaluations. Pharmacol. Res.-Mod. Chin. Med. 2024, 11, 100413. [Google Scholar] [CrossRef]
- Zheng, Y.Z.; Deng, G.; Zhang, Y.C. Multiple free radical scavenging reactions of flavonoids. Dye. Pigment. 2022, 198, 109877. [Google Scholar] [CrossRef]
- Xu, W.; Lu, H.; Yuan, Y.; Deng, Z.; Zheng, L.; Li, H. The Antioxidant and Anti-Inflammatory Effects of Flavonoids from Propolis via Nrf2 and NF-κB Pathways. Foods 2022, 11, 2439. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.L.; Fang, J.Y.; Chen, M.; Wu, C.J.; Huang, C.C.; Hung, C.F. Chrysin protects epidermal keratinocytes from UVA- and UVB-induced damage. J. Agric. Food Chem. 2011, 59, 8391–8400. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F. Targeting immune cell circuits and trafficking in inflammatory bowel disease. Nat. Immunol. 2019, 20, 970–979. [Google Scholar] [CrossRef]
- Kofla-Dłubacz, A.; Akutko, K.; Krzesiek, E.; Jamer, T.; Braksator, J.; Pytrus, T.; Stawarski, A.; Grębska, P. Selective Forms of Therapy in the Treatment of Inflammatory Bowel Diseases. J. Clin. Med. 2022, 11, 994. [Google Scholar] [CrossRef]
- Abbasi, M.; Mousavi, M.J.; Jamalzehi, S.; Alimohammadi, R.; Bezvan, M.H.; Mohammadi, H.; Aslani, S. Strategies toward rheumatoid arthritis therapy; the old and the new. J. Cell. Physiol. 2019, 234, 10018–10031. [Google Scholar] [CrossRef]
- Zeinali, M.; Rezaee, S.A.; Hosseinzadeh, H. An overview on immunoregulatory and anti-inflammatory properties of chrysin and flavonoids substances. Biomed. Pharmacother. 2017, 92, 998–1009. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Liao, T.; Yang, N.; Wei, Y.; Xing, R.; Wu, P.; Li, X.; Mao, J.; Wang, P. Chrysin ameliorates synovitis and fibrosis of osteoarthritic fibroblast-like synoviocytes in rats through PERK/TXNIP/NLRP3 signaling. Front. Pharmacol. 2023, 14, 1170243. [Google Scholar] [CrossRef]
- Küçükler, S.; Kandemir, F.M.; Yıldırım, S. Protective effect of chrysin on indomethacin induced gastric ulcer in rats: Role of multi-pathway regulation. Biotech. Histochem. 2022, 97, 490–503. [Google Scholar] [CrossRef]
- Ye, B.; Ling, W.; Wang, Y.; Jaisi, A.; Olatunji, O.J. Protective Effects of Chrysin against Cyclophosphamide-Induced Cardiotoxicity in Rats: A Biochemical and Histopathological Approach. Chem. Biodivers. 2022, 19, e202100886. [Google Scholar] [CrossRef]
- Akaras, N.; Ileriturk, M.; Gur, C.; Kucukler, S.; Oz, M.; Kandemir, F.M. The protective effects of chrysin on cadmium-induced pulmonary toxicity; a multi-biomarker approach. Environ. Sci. Pollut. Res. Int. 2023, 30, 89479–89494. [Google Scholar] [CrossRef] [PubMed]
- Sayed, H.M.; Awaad, A.S.; Abdel Rahman, F.E.Z.S.; Al-Dossari, M.; Abd El-Gawaad, N.S.; Ahmed, O.M. Combinatory Effect and Modes of Action of Chrysin and Bone Marrow-Derived Mesenchymal Stem Cells on Streptozotocin/Nicotinamide-Induced Diabetic Rats. Pharmaceuticals 2022, 16, 34. [Google Scholar] [CrossRef] [PubMed]
- Koc, F.; Tekeli, M.Y.; Kanbur, M.; Karayigit, M.Ö.; Liman, B.C. The effects of chrysin on lipopolysaccharide-induced sepsis in rats. J. Food Biochem. 2020, 44, e13359. [Google Scholar] [CrossRef] [PubMed]
- Samini, M.; Farkhondeh, T.; Azimi-Nezhad, M.; Samarghandian, S. Chrysin’s Impact on Oxidative and Inflammation Damages in the Liver of Aged Male Rats. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 743–748. [Google Scholar] [CrossRef]
- Temel, Y.; Kucukler, S.; Yıldırım, S.; Caglayan, C.; Kandemir, F.M. Protective effect of chrysin on cyclophosphamide-induced hepatotoxicity and nephrotoxicity via the inhibition of oxidative stress, inflammation, and apoptosis. Naunyn. Schmiedebergs. Arch. Pharmacol. 2020, 393, 325–337. [Google Scholar] [CrossRef]
- Oršolić, N.; Nemrava, J.; Jeleč, Ž.; Kukolj, M.; Odeh, D.; Jakopović, B.; Jembrek, M.J.; Bagatin, T.; Fureš, R.; Bagatin, D. Antioxidative and Anti-Inflammatory Activities of Chrysin and Naringenin in a Drug-Induced Bone Loss Model in Rats. Int. J. Mol. Sci. 2022, 23, 2872. [Google Scholar] [CrossRef]
- Eldutar, E.; Kandemir, F.M.; Kucukler, S.; Caglayan, C. Restorative effects of Chrysin pretreatment on oxidant-antioxidant status, inflammatory cytokine production, and apoptotic and autophagic markers in acute paracetamol-induced hepatotoxicity in rats: An experimental and biochemical study. J. Biochem. Mol. Toxicol. 2017, 31, e21960. [Google Scholar] [CrossRef]
- Jiang, Y.; Gong, F.L.; Zhao, G.B.; Li, J. Chrysin suppressed inflammatory responses and the inducible nitric oxide synthase pathway after spinal cord injury in rats. Int. J. Mol. Sci. 2014, 15, 12270–12279. [Google Scholar] [CrossRef]
- Li, R.; Zang, A.; Zhang, L.; Zhang, H.; Zhao, L.; Qi, Z.; Wang, H. Chrysin ameliorates diabetes-associated cognitive deficits in Wistar rats. Neurol. Sci. 2014, 35, 1527–1532. [Google Scholar] [CrossRef]
- Ramírez-Espinosa, J.J.; Salda a-Ríos, J.; García-Jiménez, S.; Villalobos-Molina, R.; Ávila-Villarreal, G.; Rodríguez-Ocampo, A.N.; Bernal-Fernández, G.; Estrada-Soto, S. Chrysin Induces Antidiabetic, Antidyslipidemic and Anti-Inflammatory Effects in Athymic Nude Diabetic Mice. Molecules 2017, 23, 67. [Google Scholar] [CrossRef]
- Ferrer, M.D.; Busquets-Cortés, C.; Capó, X.; Tejada, S.; Tur, J.A.; Pons, A.; Sureda, A. Cyclooxygenase-2 Inhibitors as a Therapeutic Target in Inflammatory Diseases. Curr. Med. Chem. 2018, 26, 3225–3241. [Google Scholar] [CrossRef] [PubMed]
- Byun, E.B.; Song, H.Y.; Kim, W.S.; Han, J.M.; Seo, H.S.; Park, W.Y.; Kim, K.; Byun, E.H. Chrysin derivative CM1 and exhibited anti-inflammatory action by upregulating toll-interacting protein expression in lipopolysaccharide-stimulated RAW264.7 macrophage cells. Molecules 2021, 26, 1532. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Tao, Z.; Cai, L.; Chen, C.; Zhang, C.; Wang, Q.; Ying, X.; Hu, W.; Chen, H. Chrysin Attenuates IL-1β-Induced Expression of Inflammatory Mediators by Suppressing NF-κB in Human Osteoarthritis Chondrocytes. Inflammation 2017, 40, 1143–1154. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.H.; Suh, B.; Shin, I.; Kim, E.H.; Kim, D.; Shin, Y.J.; Chang, S.Y.; Baek, S.H.; Kim, H.; Bae, O.N. Inhibitory effects of a novel chrysin-derivative, CPD 6, on acute and chronic skin inflammation. Int. J. Mol. Sci. 2019, 20, 2607. [Google Scholar] [CrossRef]
- Yang, C.; Wang, T.; Zhao, Y.; Meng, X.; Ding, W.; Wang, Q.; Liu, C.; Deng, H. Flavonoid 4,4’-dimethoxychalcone induced ferroptosis in cancer cells by synergistically activating Keap1/Nrf2/HMOX1 pathway and inhibiting FECH. Free Radic. Biol. Med. 2022, 188, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Ji, L.L.; Sheng, Y.C.; Zheng, Z.Y.; Shi, L.; Wang, Z.T. The involvement of p62-Keap1-Nrf2 antioxidative signaling pathway and JNK in the protection of natural flavonoid quercetin against hepatotoxicity. Free Radic. Biol. Med. 2015, 85, 12–23. [Google Scholar] [CrossRef]
- Lo, S.C.; Li, X.; Henzl, M.T.; Beamer, L.J.; Hannink, M. Structure of the Keap1:Nrf2 interface provides mechanistic insight into Nrf2 signaling. EMBO J. 2006, 25, 3605–3617. [Google Scholar] [CrossRef]
- Canning, P.; Sorrell, F.J.; Bullock, A.N. Structural basis of Keap1 interactions with Nrf2. Free Radic. Biol. Med. 2015, 88, 101–107. [Google Scholar] [CrossRef]
- Wen, X.; Walle, T. Methylated flavonoids have greatly improved intestinal absorption and metabolic stability. Drug Metab. Dispos. 2006, 34, 1786–1792. [Google Scholar] [CrossRef]
- Isika, D.K.; Özkömeç, F.N.; Çeşme, M.; Sadik, O.A. Synthesis, biological and computational studies of flavonoid acetamide derivatives. RSC Adv. 2022, 12, 10037–10050. [Google Scholar] [CrossRef]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef] [PubMed]
- Twentyman, P.R.; Luscombe, M. A study of some variables in a tetrazolium dye (MTT) based assay for cell growth and chemosensitivity. Br. J. Cancer 1987, 56, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Adelusi, T.I.; Abdul-Hammed, M.; Idris, M.O.; Oyedele, Q.K.; Adedotun, I.O. Molecular dynamics, quantum mechanics and docking studies of some Keap1 inhibitors—An insight into the atomistic mechanisms of their antioxidant potential. Heliyon 2021, 7, e07317. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ligand | Binding Affinity ΔG, (kcal/mol) | Aminoacids of Keap-1 Receptor Forming Intermolecular Interactions | Types of Interaction |
|---|---|---|---|
| 1 | −8.9 | Ser363, Asn382, Asn414, Arg415, Gln530, Ser602 | Conventional Hydrogen Bond |
| Arg415 | Pi-Cation | ||
| Tyr572 | Pi-Pi Stacked | ||
| Ala556 | Pi-Alkyl | ||
| 1aP1 | −8.2 | Tyr334, Arg380, Asn382, Asn 387, Arg415, Ala556, Ser602 | Conventional Hydrogen Bond |
| Arg415, Gly462, Gly509, Ser555 | Carbon Hydrogen Bond | ||
| Arg415 | Pi-Cation | ||
| Tyr525 | Pi-Pi T-shaped | ||
| Ala556 | Pi-Alkyl | ||
| Tyr334 | Unfavorable Acceptor-Acceptor | ||
| 1aP2 | Gly364, Arg380 Asn382, Arg415 | Conventional Hydrogen Bond | |
| Ser363, gly,364, gly462, gly603 | Carbon Hydrogen Bond | ||
| Arg415, Arg483 | Pi-Cation | ||
| Tyr525 | Pi-Pi Stacked | ||
| Ala556 | Pi-Alkyl | ||
| 1bP1 | −7.9 | Arg380, Asn382, Arg415 | Conventional Hydrogen Bond |
| Gly509, Ala556 | Carbon Hydrogen Bond | ||
| Arg415, Arg483 | Pi-Cation | ||
| Tyr334, Arg415 | Alkyl | ||
| Ala556 | Pi-Alkyl | ||
| 1bP2 | Arg380, Asn382, Arg415, Gln530 | Conventional Hydrogen Bond | |
| Gly462, Gly509 | Carbon Hydrogen Bond | ||
| Arg415, Arg483 | Pi-Cation | ||
| Tyr525 | Pi-Sigma | ||
| Arg415, Tyr572 | Alkyl | ||
| Ala556, Phe577 | Pi-Alkyl | ||
| 1bP3 | Arg380, Asn382, Arg415, Ser602 | Conventional Hydrogen Bond | |
| Gly462, Gly509 | Carbon Hydrogen Bond | ||
| Arg415, Arg483 | Pi-Cation | ||
| Arg415 | Alkyl | ||
| Ala556 | Pi-Alkyl | ||
| Native ligand | −9.3 | Ser508, Ser602 | Conventional Hydrogen Bond |
| Arg415 | Pi-Cation | ||
| Ala556 | Pi-Sigma | ||
| Tyr334, Tyr525, Phe577 | Pi-Pi | ||
| Tyr572 | Pi-Alkyl |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ávila-Román, J.; Quevedo-Tinoco, L.; Oliveros-Ortiz, A.J.; García-Gil, S.; Rodríguez-García, G.; Motilva, V.; Gómez-Hurtado, M.A.; Talero, E. Synthesis and Bioevaluation of New Stable Derivatives of Chrysin-8-C-Glucoside That Modulate the Antioxidant Keap1/Nrf2/HO-1 Pathway in Human Macrophages. Pharmaceuticals 2024, 17, 1388. https://doi.org/10.3390/ph17101388
Ávila-Román J, Quevedo-Tinoco L, Oliveros-Ortiz AJ, García-Gil S, Rodríguez-García G, Motilva V, Gómez-Hurtado MA, Talero E. Synthesis and Bioevaluation of New Stable Derivatives of Chrysin-8-C-Glucoside That Modulate the Antioxidant Keap1/Nrf2/HO-1 Pathway in Human Macrophages. Pharmaceuticals. 2024; 17(10):1388. https://doi.org/10.3390/ph17101388
Chicago/Turabian StyleÁvila-Román, Javier, Lirenny Quevedo-Tinoco, Antonio J. Oliveros-Ortiz, Sara García-Gil, Gabriela Rodríguez-García, Virginia Motilva, Mario A. Gómez-Hurtado, and Elena Talero. 2024. "Synthesis and Bioevaluation of New Stable Derivatives of Chrysin-8-C-Glucoside That Modulate the Antioxidant Keap1/Nrf2/HO-1 Pathway in Human Macrophages" Pharmaceuticals 17, no. 10: 1388. https://doi.org/10.3390/ph17101388
APA StyleÁvila-Román, J., Quevedo-Tinoco, L., Oliveros-Ortiz, A. J., García-Gil, S., Rodríguez-García, G., Motilva, V., Gómez-Hurtado, M. A., & Talero, E. (2024). Synthesis and Bioevaluation of New Stable Derivatives of Chrysin-8-C-Glucoside That Modulate the Antioxidant Keap1/Nrf2/HO-1 Pathway in Human Macrophages. Pharmaceuticals, 17(10), 1388. https://doi.org/10.3390/ph17101388